
Redspotted Grouper Nervous Necrosis Virus and the Reassortant RGNNV/SJNNV In Vitro Susceptibility against a Commercial Peroxy-Acid Biocide under Different Conditions of Use

{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Suspension Virucidal Activity Test
2.3. Net Virucidal Activity Test
2.4. Statistical Analyses
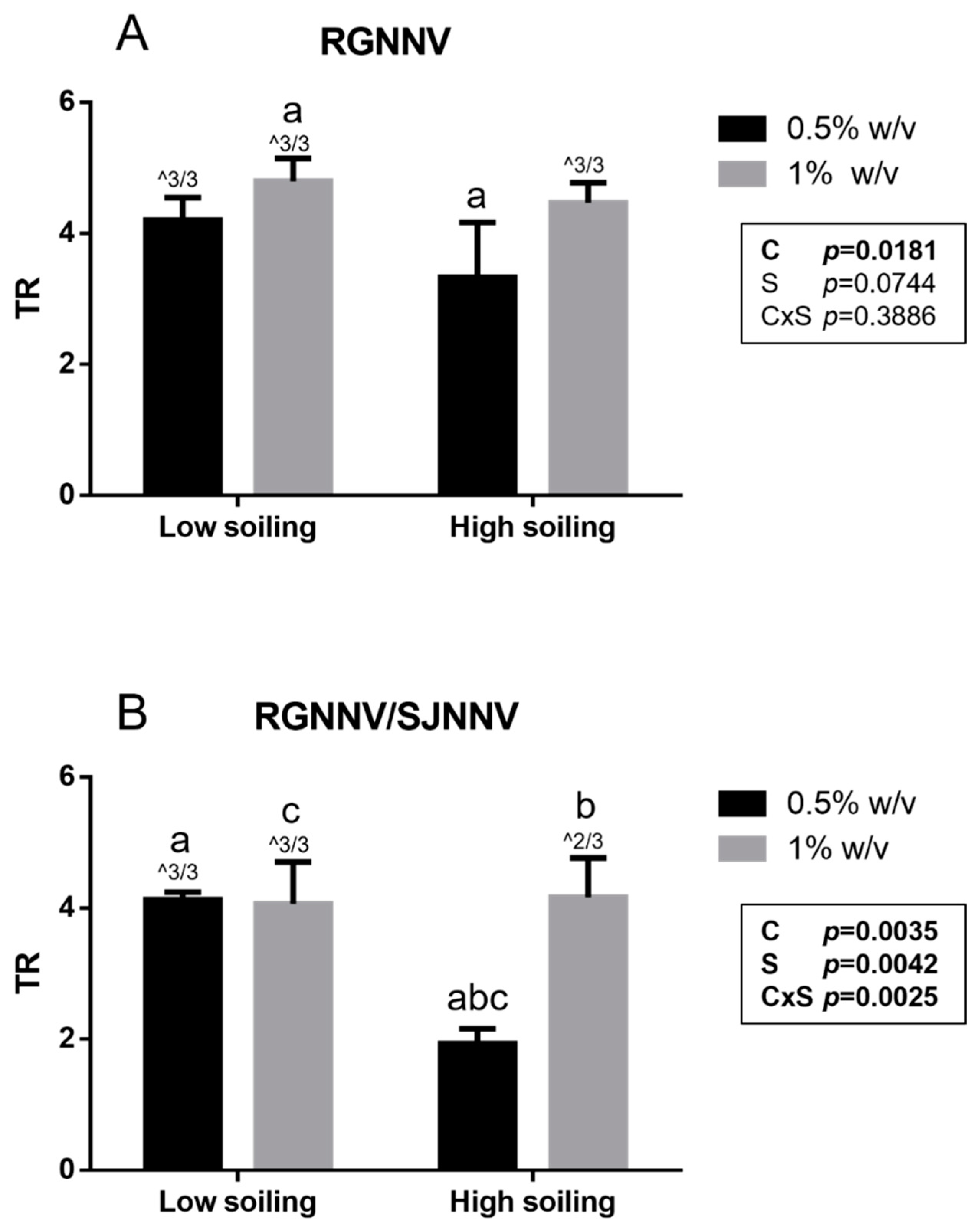
3. Results
3.1. Suspension Virucidal Activity Test
3.2. Net Virucidal Activity Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020; FAO: Rome, Italy, 2020. [Google Scholar]
- Padrós, F.; Caggiano, M.; Toffan, A.; Constenla, M.; Zarza, C.; Ciulli, S. Integrated Management Strategies for Viral Nervous Necrosis (VNN) Disease Control in Marine Fish Farming in the Mediterranean. Pathogens 2022, 11, 330. [Google Scholar] [CrossRef] [PubMed]
- Bandín, I.; Souto, S. Betanodavirus and VER Disease: A 30-Year Research Review. Pathogens 2020, 9, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toffan, A.; Pascoli, F.; Pretto, T.; Panzarin, V.; Abbadi, M.; Buratin, A.; Quartesan, R.; Gijón, D.; Padrós, F. Viral Nervous Necrosis in Gilthead Sea Bream (Sparus aurata) Caused by Reassortant Betanodavirus RGNNV/SJNNV: An Emerging Threat for Mediterranean Aquaculture. Sci. Rep. 2017, 7, 46755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doan, Q.K.; Vandeputte, M.; Chatain, B.; Morin, T.; Allal, F. Viral Encephalopathy and Retinopathy in Aquaculture: A Review. J. Fish Dis. 2017, 40, 717–742. [Google Scholar] [CrossRef] [Green Version]
- Thiéry, R.; Johnson, K.L.; Nakai, T.; Schneemann, A.; Bonami, J.R.; Lightner, D.V. Family Nodaviridae. In Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: San Diego, CA, USA, 2011; pp. 1061–1067. [Google Scholar]
- Mori, K.; Nakai, T.; Muroga, K.; Arimoto, M.; Mushiake, K.; Furusawa, I. Properties of a New Virus Belonging to Nodaviridae Found in Larval Striped Jack (Pseudocaranx dentex) with Nervous Necrosis. Virology 1992, 187, 368–371. [Google Scholar] [CrossRef]
- Nagai, T.; Nishizawa, T. Sequence of the Non-Structural Protein Gene Encoded by RNA1 of Striped Jack Nervous Necrosis Virus. J. Gen. Virol. 1999, 80, 3019–3022. [Google Scholar] [CrossRef]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic Classification of Fish Nodaviruses by Molecular Phylogenetic Analysis of the Coat Protein Gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef] [Green Version]
- Volpe, E.; Gustinelli, A.; Caffara, M.; Errani, F.; Quaglio, F.; Fioravanti, M.L.; Ciulli, S. Viral Nervous Necrosis Outbreaks Caused by the RGNNV/SJNNV Reassortant Betanodavirus in Gilthead Sea Bream (Sparus aurata) and European Sea Bass (Dicentrarchus labrax). Aquaculture 2020, 523, 735155. [Google Scholar] [CrossRef]
- Volpe, E.; Grodzki, M.; Panzarin, V.; Guercio, A.; Purpari, G.; Serratore, P.; Ciulli, S. Detection and Molecular Characterization of Betanodaviruses Retrieved from Bivalve Molluscs. J. Fish Dis. 2018, 41, 603–611. [Google Scholar] [CrossRef]
- Errani, F.; Ciulli, S.; Mandrioli, L.; Serratore, P.; Volpe, E. Detection of Human and Fish Viruses in Marine Gastropods. Animals 2022, 12, 2122. [Google Scholar] [CrossRef]
- Ciulli, S.; Galletti, E.; Grodzki, M.; Alessi, A.; Battilani, M.; Prosperi, S. Isolation and Genetic Characterization of Betanodavirus from Wild Marine Fish from the Adriatic Sea. Vet. Res. Commun. 2007, 31, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Volpe, E.; Pagnini, N.; Serratore, P.; Ciulli, S. Fate of Redspotted Grouper Nervous Necrosis Virus (RGNNV) in Experimentally Challenged Manila Clam Ruditapes Philippinarum. Dis. Aquat. Organ. 2017, 125, 53–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, M.; Cabon, J.; Salomoni, A.; Toffan, A.; Panzarin, V.; Bigarré, L. First Generic One Step Real-Time Taqman RT-PCR Targeting the RNA1 of Betanodaviruses. J. Virol. Methods 2015, 211, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Errani, F.; Volpe, E.; Riera-Ferrer, E.; Caffara, M.; Padrós, F.; Gustinelli, A.; Fioravanti, M.; Ciulli, S. Development and Diagnostic Validation of a One-Step Multiplex RT-PCR Assay as a Rapid Method to Detect and Identify Nervous Necrosis Virus (NNV) and Its Variants Circulating in the Mediterranean. PLoS ONE 2022, 17, e0273802. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Ellakany, H.F.; Elbestawy, A.R.; Abaza, S.S.; Geneedy, A.M.; Khafaga, A.F.; Salem, H.M.; Abd El-Aziz, A.H.; Selim, S.; et al. Inhibition of Microbial Pathogens in Farmed Fish. Mar. Pollut. Bull. 2022, 183, 114003. [Google Scholar] [CrossRef] [PubMed]
- Arimoto, M.; Sato, J.; Maruyama, K.; Mimura, G.; Furusawa, I. Effect of Chemical and Physical Treatments on the Inactivation of Striped Jack Nervous Necrosis Virus (SJNNV). Aquaculture 1996, 143, 15–22. [Google Scholar] [CrossRef]
- Frerichs, G.N.; Tweedie, A.; Starkey, W.G.; Richards, R.H. Temperature, PH and Electrolyte Sensitivity, and Heat, UV and Disinfectant Inactivation of Sea Bass ŽDicentrarchus Labrax/ Neuropathy Nodavirus. Aquaculture 2000, 185, 13–24. [Google Scholar] [CrossRef]
- Verner-Jeffreys, D.W.; Joiner, C.L.; Bagwell, N.J.; Reese, R.A.; Husby, A.; Dixon, P.F. Development of Bactericidal and Virucidal Testing Standards for Aquaculture Disinfectants. Aquaculture 2009, 286, 190–197. [Google Scholar] [CrossRef]
- Graham, D.A.; Cherry, K.; Wilson, C.J.; Rowley, H.M. Susceptibility of Salmonid Alphavirus to a Range of Chemical Disinfectants. J. Fish Dis. 2007, 30, 269–277. [Google Scholar] [CrossRef]
- Maltese, C.; Bovo, G. Effetto Di Alcuni Trattamenti Chimico-Fisici Nei Confronti Dell’agente Causale Dell’Encefalo-Retinopatia Virale Del Branzino d’allevamento (Dicentrarchus labrax). Boll. Soc. Ital. Patol. Ittica 2001, 31, 3–16. [Google Scholar]
- Tyski, S.; Bocian, E.; Laudy, A.E. Application of Normative Documents for Determination of Biocidal Activity of Disinfectants and Antiseptics Dedicated to the Medical Area: A Narrative Review. J. Hosp. Infect. 2022, 125, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Ciulli, S.; Gallardi, D.; Scagliarini, A.; Battilani, M.; Hedrick, R.P.; Prosperi, S. Temperature-Dependency of Betanodavirus Infection in SSN-1 Cell Line. Dis. Aquat. Organ. 2006, 68, 261–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hierholzer, J.C.; Killington, R.A. 2—Virus Isolation and Quantitation. In Virology Methods Manual; Mahy, B.W., Kangro, H.O., Eds.; Academic Press: London, UK, 1996; pp. 25–46. ISBN 978-0-12-465330-6. [Google Scholar]
- BS EN 14675; Chemical Disinfectants and Antiseptics—Quantitative Suspension Test for the Evaluation of Virucidal Activity of Chemical Disinfectants and Antiseptics Used in the Veterinary Area—Test Method and Requirements (Phase 2, Step 1). BSI: London, UK, 2015.
- BS EN 17111; Chemical Disinfectants and Antiseptics—Quantitative Carrier Test for the Evaluation of Virucidal Activity for Instruments Used in the Medical Area—Test Method and Requirements (Phase 2, Step 2). BSI: London, UK, 2018.
- Tidbury, H.J.; Joiner, C.L.; Rimmer, G.S.E.; Potter, H.V.; Taylor, N.G.H. The Effectiveness of Fishery Net Dips: Advice for the Improvement of Biosecurity Measures. J. Fish Dis. 2018, 41, 1625–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Lim, J.Y.C.; Xue, K.; Yew, P.Y.M.; Owh, C.; Chee, P.L.; Loh, X.J. Sanitizing Agents for Virus Inactivation and Disinfection. View 2020, 1, e16. [Google Scholar] [CrossRef]
- Wessels, S.; Ingmer, H. Modes of Action of Three Disinfectant Active Substances: A Review. Regul. Toxicol. Pharmacol. 2013, 67, 456–467. [Google Scholar] [CrossRef]
- Collins, A.M.; Fell, S.A.; Barchia, I.M. Cleaning and Disinfection with Virkon S Significantly Reduces Lawsonia Intracellularis Survival and Transmission to Naive Pigs. J. Swine Health Prod. 2013, 21, 144–147. [Google Scholar]
- Nishizawa, T.; Mori, K.I.; Furuhashi, M.; Nakai, T.; Furusawa, I.; Muroga, K. Comparison of the Coat Protein Genes of Five Fish Nodaviruses, the Causative Agents of Viral Nervous Necrosis in Marine Fish. J. Gen. Virol. 1995, 76, 1563–1569. [Google Scholar] [CrossRef]
- Olveira, J.G.; Souto, S.; Dopazo, C.P.; Thiéry, R.; Barja, J.L.; Bandín, I. Comparative Analysis of Both Genomic Segments of Betanodaviruses Isolated from Epizootic Outbreaks in Farmed Fish Species Provides Evidence for Genetic Reassortment. J. Gen. Virol. 2009, 90, 2940–2951. [Google Scholar] [CrossRef]
- Panzarin, V.; Toffan, A.; Abbadi, M.; Buratin, A.; Mancin, M.; Braaen, S.; Olsen, C.M.; Bargelloni, L.; Rimstad, E.; Cattoli, G. Molecular Basis for Antigenic Diversity of Genus Betanodavirus. PLoS ONE 2016, 11, e0158814. [Google Scholar] [CrossRef] [Green Version]
- Biasini, L.; Berto, P.; Abbadi, M.; Buratin, A.; Toson, M.; Marsella, A.; Toffan, A.; Pascoli, F. Pathogenicity of Different Betanodavirus RGNNV/SJNNV Reassortant Strains in European Sea Bass. Pathogens 2022, 11, 458. [Google Scholar] [CrossRef]
- Brill, H.H.F.; Becker, B.; Todt, D.; Steinmann, J.; Paulmann, D.; Bischoff, B.; Steinmann, J. Virucidal Efficacy of Glutaraldehyde for Instrument Disinfection. GMS Hyg. Infect. Control 2020, 15, Doc34. [Google Scholar] [CrossRef] [PubMed]
- Kindermann, J.; Karbiener, M.; Leydold, S.M.; Knotzer, S.; Modrof, J.; Kreil, T.R. Virus Disinfection for Biotechnology Applications: Different Effectiveness on Surface versus in Suspension. Biologicals 2020, 64, 1–9. [Google Scholar] [CrossRef] [PubMed]
- El-Athman, F.; Zehlike, L.; Kämpfe, A.; Junek, R.; Selinka, H.C.; Mahringer, D.; Grunert, A. Pool Water Disinfection by Ozone-Bromine Treatment: Assessing the Disinfectant Efficacy and the Occurrence and in Vitro Toxicity of Brominated Disinfection By-Products. Water Res. 2021, 204, 117648. [Google Scholar] [CrossRef]
- Chowdhury, D.; Rahman, A.; Hu, H.; Jensen, S.O.; Deva, A.K.; Vickery, K. Effect of Disinfectant Formulation and Organic Soil on the Efficacy of Oxidizing Disinfectants against Biofilms. J. Hosp. Infect. 2019, 103, e33–e41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, C.; Sun, G.; Tang, X.; Gan, Y.; Liang, G.; Wang, J.; Huang, Y. Bactericidal Efficacy of a Low Concentration of Vaporized Hydrogen Peroxide with Validation in a BSL-3 Laboratory. J. Hosp. Infect. 2022, 127, 51–58. [Google Scholar] [CrossRef]
- Oliveira, I.M.; Gomes, I.B.; Simões, L.C.; Simões, M. Chlorinated Cyanurates and Potassium Salt of Peroxymonosulphate as Antimicrobial and Antibiofilm Agents for Drinking Water Disinfection. Sci. Total Environ. 2022, 811, 152355. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volpe, E.; Errani, F.; Zamparo, S.; Ciulli, S. Redspotted Grouper Nervous Necrosis Virus and the Reassortant RGNNV/SJNNV In Vitro Susceptibility against a Commercial Peroxy-Acid Biocide under Different Conditions of Use. Vet. Sci. 2023, 10, 76. https://doi.org/10.3390/vetsci10020076
Volpe E, Errani F, Zamparo S, Ciulli S. Redspotted Grouper Nervous Necrosis Virus and the Reassortant RGNNV/SJNNV In Vitro Susceptibility against a Commercial Peroxy-Acid Biocide under Different Conditions of Use. Veterinary Sciences. 2023; 10(2):76. https://doi.org/10.3390/vetsci10020076
Chicago/Turabian StyleVolpe, Enrico, Francesca Errani, Samuele Zamparo, and Sara Ciulli. 2023. "Redspotted Grouper Nervous Necrosis Virus and the Reassortant RGNNV/SJNNV In Vitro Susceptibility against a Commercial Peroxy-Acid Biocide under Different Conditions of Use" Veterinary Sciences 10, no. 2: 76. https://doi.org/10.3390/vetsci10020076