
Low-Dose Radiation-Induced Transcriptomic Changes in Diabetic Aortic Endothelial Cells
 , , , and
, , , and
Abstract
:1. Summary
2. Data Description
2.1. Quality Control Analysis of RNA-seq Data
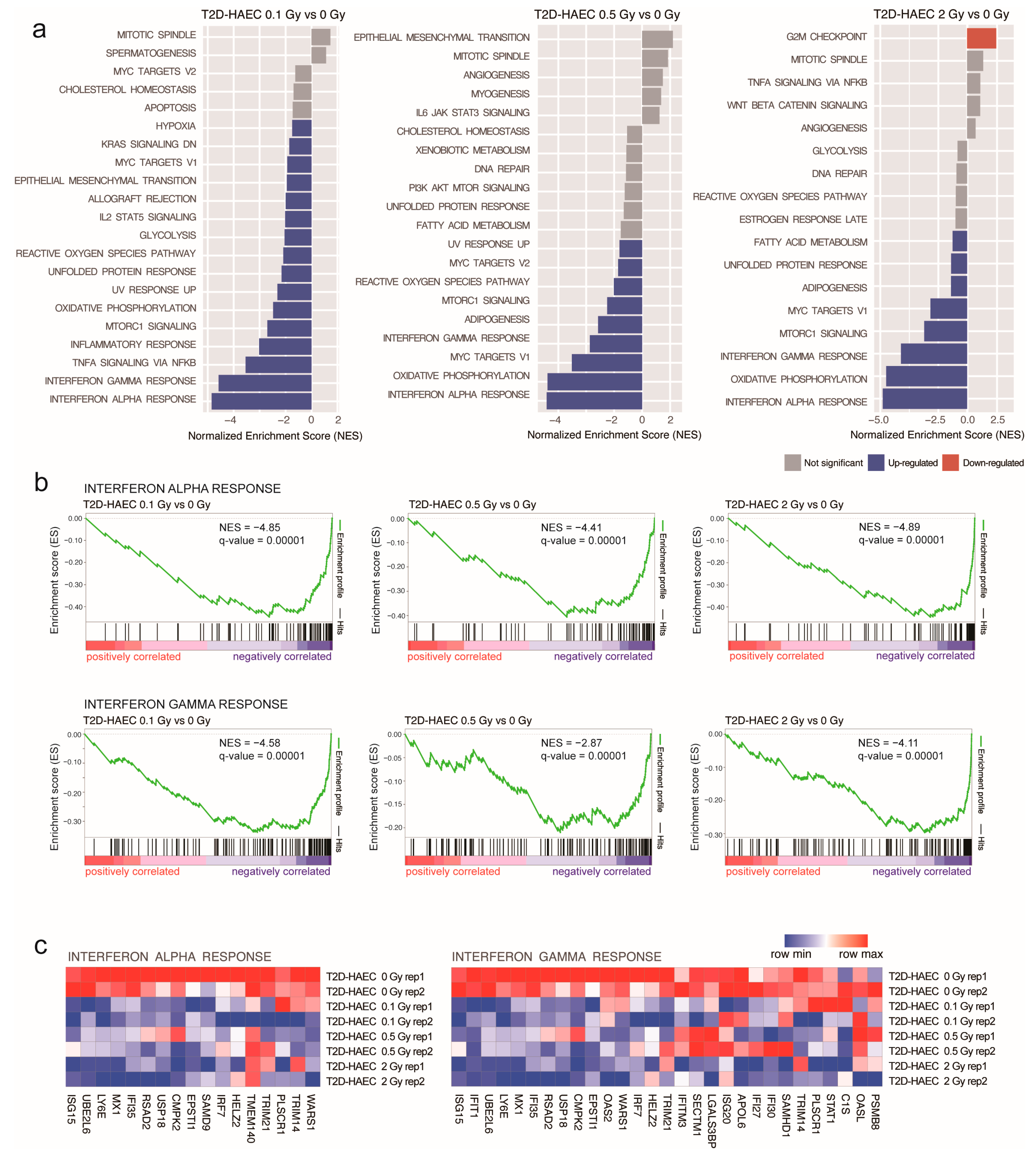
2.2. Differential Gene Expression and Pathway Analyses
3. Methods
3.1. Cell Culture
3.2. Radiation Exposure
3.3. RNA Isolation
3.4. Library Preparation and Sequencing
3.5. RNA Sequencing Data Analysis
3.6. Gene Set Enrichement Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ali, Y.F.; Cucinotta, F.A.; Ning-Ang, L.; Zhou, G. Cancer Risk of Low Dose Ionizing Radiation. Front. Phys. 2020, 8, 234. [Google Scholar] [CrossRef]
- Ghandhi, S.A.; Smilenov, L.B.; Elliston, C.D.; Chowdhury, M.; Amundson, S.A. Radiation dose-rate effects on gene expression for human biodosimetry. BMC Med. Genom. 2015, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Poonsombudlert, K.; Limpruttidham, N. Total Body Irradiation and Risk of Diabetes Mellitus; A Meta-Analysis. Asian Pac. J. Cancer Prev. 2019, 20, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lin, X.; Yu, L.; Li, W.; Qian, D.; Cheng, P.; He, L.; Yang, H.; Zhang, C. Low-dose radiation prevents type 1 diabetes-induced cardiomyopathy via activation of AKT mediated anti-apoptotic and anti-oxidant effects. J. Cell. Mol. Med. 2016, 20, 1352–1366. [Google Scholar] [CrossRef] [PubMed]
- Teinturier, C.; Tournade, M.-F.; Caillat-Zucman, S.; Boitard, C.; Amoura, Z.; Bougneres, P.-F.; Timsit, J. Diabetes mellitus after abdominal radiation therapy. Lancet 1995, 346, 633–634. [Google Scholar] [CrossRef] [PubMed]
- Alexander, Y.; Osto, E.; Schmidt-Trucksäss, A.; Shechter, M.; Trifunovic, D.; Duncker, D.J.; Aboyans, V.; Bäck, M.; Badimon, L.; Cosentino, F.; et al. Endothelial function in cardiovascular medicine: A consensus paper of the European Society of Cardiology Working Groups on Atherosclerosis and Vascular Biology, Aorta and Peripheral Vascular Diseases, Coronary Pathophysiology and Microcirculation, and Thrombosis. Cardiovasc. Res. 2021, 117, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Naveed, A.; Farrukh, L.; Sana, M.K.; Naveed, B.; Randhawa, F.A. Pharmacological Primary Prevention of Diabetes Mellitus Type II: A Narrative Review. Cureus 2020, 12, e10033. [Google Scholar] [CrossRef] [PubMed]
- Leon, B.M.; Maddox, T.M. Diabetes and cardiovascular disease: Epidemiology, biological mechanisms, treatment recommendations and future research. World J. Diabetes 2015, 6, 1246–1258. [Google Scholar] [CrossRef] [PubMed]
- Meacham, L.R.; Sklar, C.A.; Li, S.; Liu, Q.; Gimpel, N.; Yasui, Y.; Whitton, J.A.; Stovall, M.; Robison, L.L.; Oeffinger, K.C.; et al. Diabetes mellitus in long-term survivors of childhood cancer. Increased risk associated with radiation therapy: A report for the childhood cancer survivor study. Arch. Intern. Med. 2009, 169, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Tatsukawa, Y.; Cordova, K.; Yamada, M.; Ohishi, W.; Imaizumi, M.; Hida, A.; Sposto, R.; Sakata, R.; Fujiwara, S.; Nakanishi, S.; et al. Incidence of Diabetes in the Atomic Bomb Survivors: 1969–2015. J. Clin. Endocrinol. Metab. 2022, 107, e2148–e2155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Jin, S.; Guo, W.; Li, C.; Li, X.; Rane, M.J.; Wang, G.; Cai, L. Attenuation of diabetes-induced cardiac inflammation and pathological remodeling by low-dose radiation. Radiat. Res. 2011, 175, 307–321. [Google Scholar] [CrossRef]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, N.; Kim, M.-J.; Kim, R.-K.; Kaushik, N.K.; Seong, K.M.; Nam, S.-Y.; Lee, S.-J. Low-dose radiation decreases tumor progression via the inhibition of the JAK1/STAT3 signaling axis in breast cancer cell lines. Sci. Rep. 2017, 7, 43361. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Nam, K.-S.; Lee, H.-J.; Kim, K.S. Ionizing Radiation-Induced GDF15 Promotes Angiogenesis in Human Glioblastoma Models by Promoting VEGFA Expression Through p-MAPK1/SP1 Signaling. Front. Oncol. 2022, 12, 801230. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinform. 2015, 51, 11.14.1–11.14.19. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Sample | Total Number of Reads | Uniquely Mapped Reads (%) | TIN (Median) | |
|---|---|---|---|---|
| Raw Reads | Filtered Reads | |||
| T2D-HAEC 0 Gy rep1 | 31,213,313 | 30,849,903 | 67.02 | 74.69 |
| T2D-HAEC 0 Gy rep2 | 31,266,215 | 30,911,938 | 66.53 | 76.27 |
| T2D-HAEC 0.1 Gy rep1 | 31,300,635 | 30,373,825 | 69.94 | 71.40 |
| T2D-HAEC 0.1 Gy rep2 | 31,166,495 | 30,857,100 | 75.49 | 77.32 |
| T2D-HAEC 0.5 Gy rep1 | 31,308,491 | 30,076,440 | 72.15 | 70.08 |
| T2D-HAEC 0.5 Gy rep2 | 31,551,549 | 31,207,062 | 76.69 | 77.72 |
| T2D-HAEC 2 Gy rep1 | 31,390,592 | 30,915,932 | 77.76 | 74.72 |
| T2D-HAEC 2 Gy rep2 | 31,197,385 | 30,789,716 | 76.39 | 78.39 |
| HAEC 0 Gy rep1 | 26,663,037 | 26,602,485 | 65.07 | 78.38 |
| HAEC 0 Gy rep2 | 28,990,602 | 28,865,837 | 53.84 | 75.03 |
| HAEC 0.1 Gy rep1 | 31,210,032 | 31,175,477 | 75.98 | 79.38 |
| HAEC 0.1 Gy rep2 | 31,530,652 | 31,499,020 | 75.50 | 79.31 |
| HAEC 0.5 Gy rep1 | 31,075,651 | 31,043,188 | 55.25 | 78.82 |
| HAEC 0.5 Gy rep2 | 31,188,257 | 31,153,901 | 56.25 | 79.39 |
| HAEC 2 Gy rep1 | 31,359,761 | 31,279,381 | 54.61 | 77.62 |
| HAEC 2 Gy rep2 | 28,409,483 | 28,313,543 | 57.00 | 76.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Kang, K.; Son, Y.; Kim, K.S.; Kang, K.; Lee, H.-J. Low-Dose Radiation-Induced Transcriptomic Changes in Diabetic Aortic Endothelial Cells. Data 2023, 8, 92. https://doi.org/10.3390/data8050092
Park J, Kang K, Son Y, Kim KS, Kang K, Lee H-J. Low-Dose Radiation-Induced Transcriptomic Changes in Diabetic Aortic Endothelial Cells. Data. 2023; 8(5):92. https://doi.org/10.3390/data8050092
Chicago/Turabian StylePark, Jihye, Kyuho Kang, Yeonghoon Son, Kwang Seok Kim, Keunsoo Kang, and Hae-June Lee. 2023. "Low-Dose Radiation-Induced Transcriptomic Changes in Diabetic Aortic Endothelial Cells" Data 8, no. 5: 92. https://doi.org/10.3390/data8050092