
Effects and Prognostic Values of Circadian Genes CSNK1E/GNA11/KLF9/THRAP3 in Kidney Renal Clear Cell Carcinoma via a Comprehensive Analysis

 , , and
, , and
Abstract
:1. Introduction
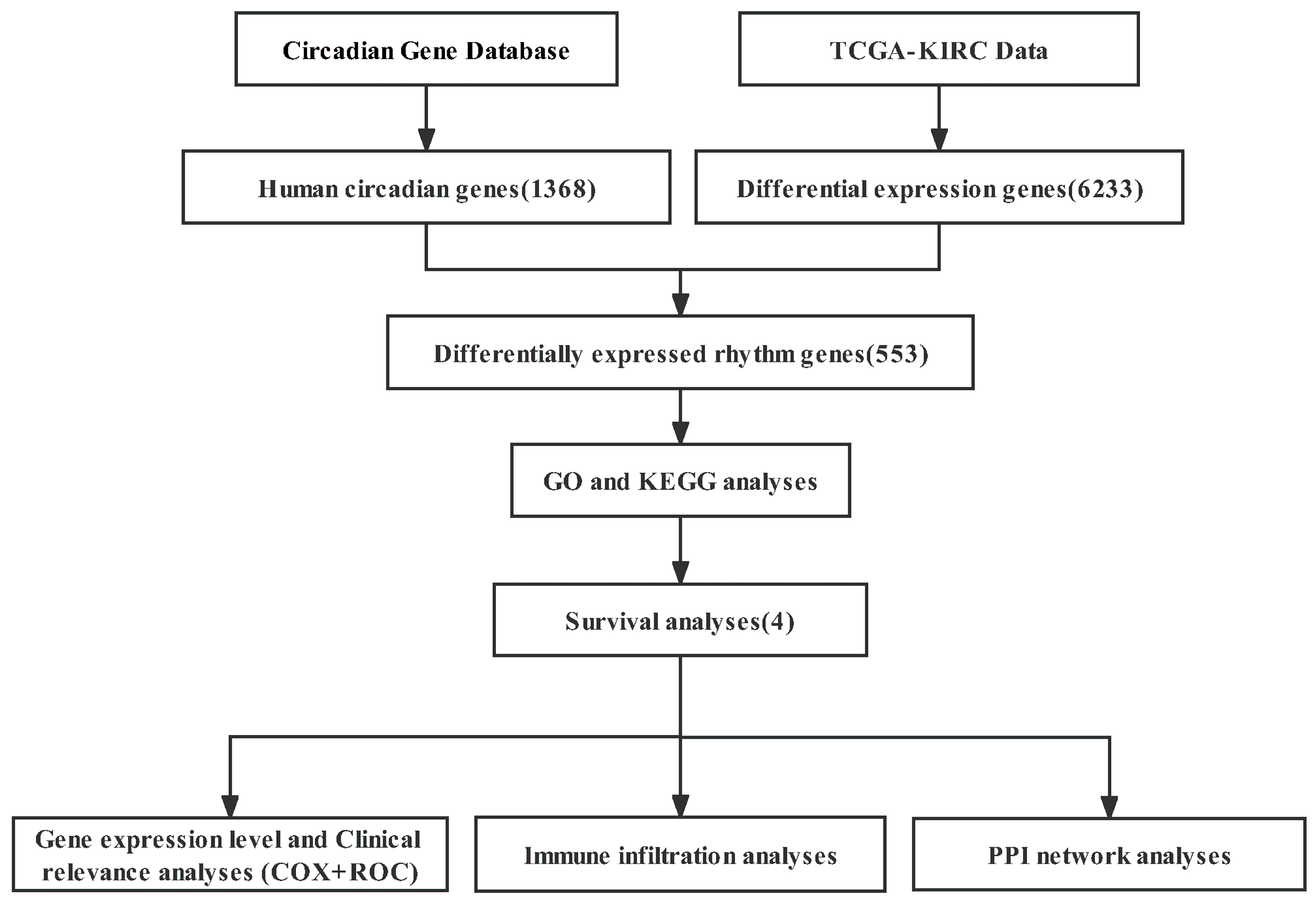
2. Materials and Methods
2.1. Data Collection
2.2. DERGs Identification
2.3. Functional Enrichment Analyses
2.4. Survival Analyses
2.5. Matching Analyses
2.6. Immunohistochemical Analyses
2.7. Clinical Correlation Analyses
2.8. Immune Infiltration Analyses
2.9. Protein–Protein Interaction Network Analyses
2.10. Cell Culture
2.11. Quantitative Real-Time PCR
2.12. Statistical Analysis
3. Results
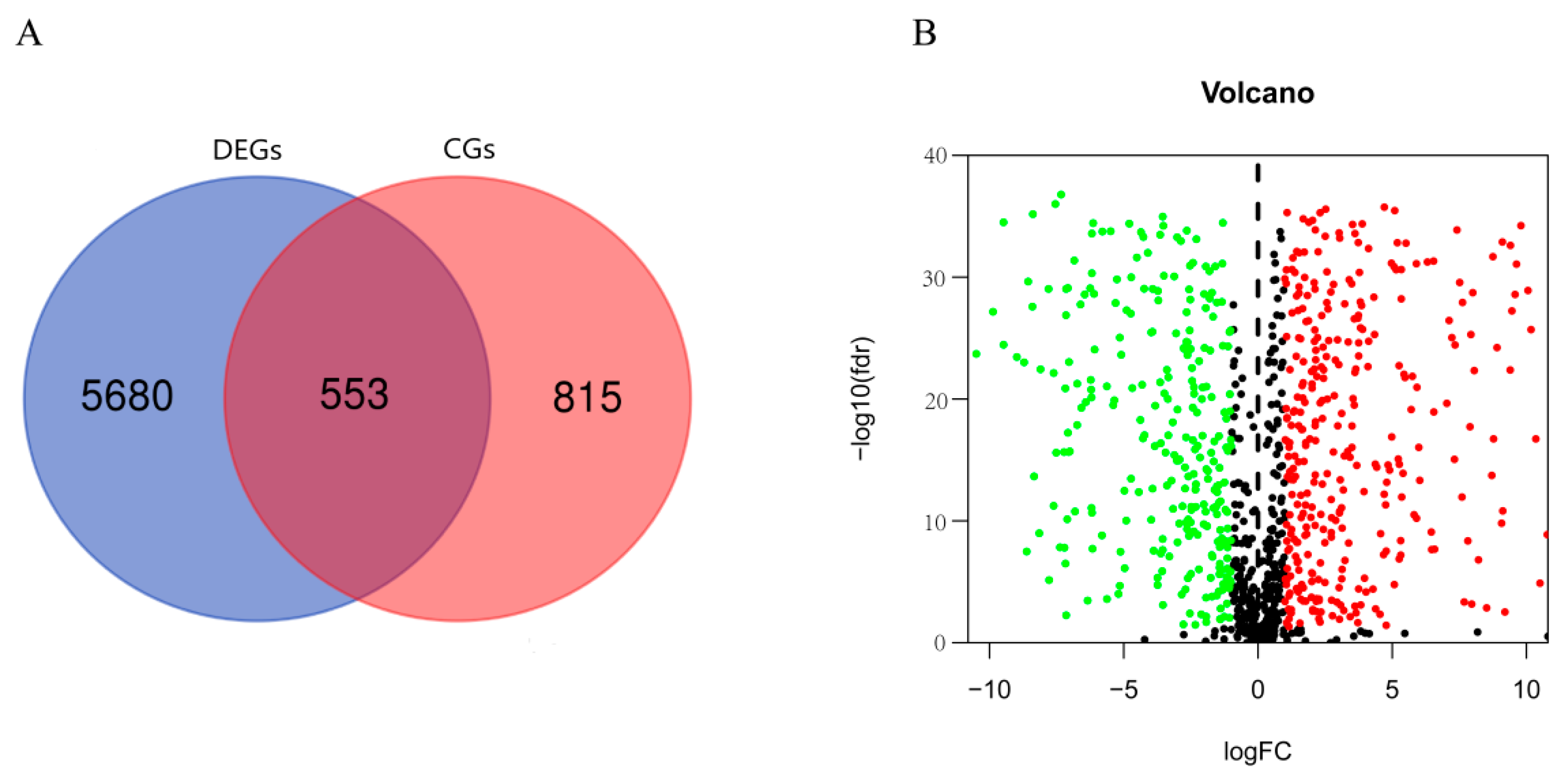
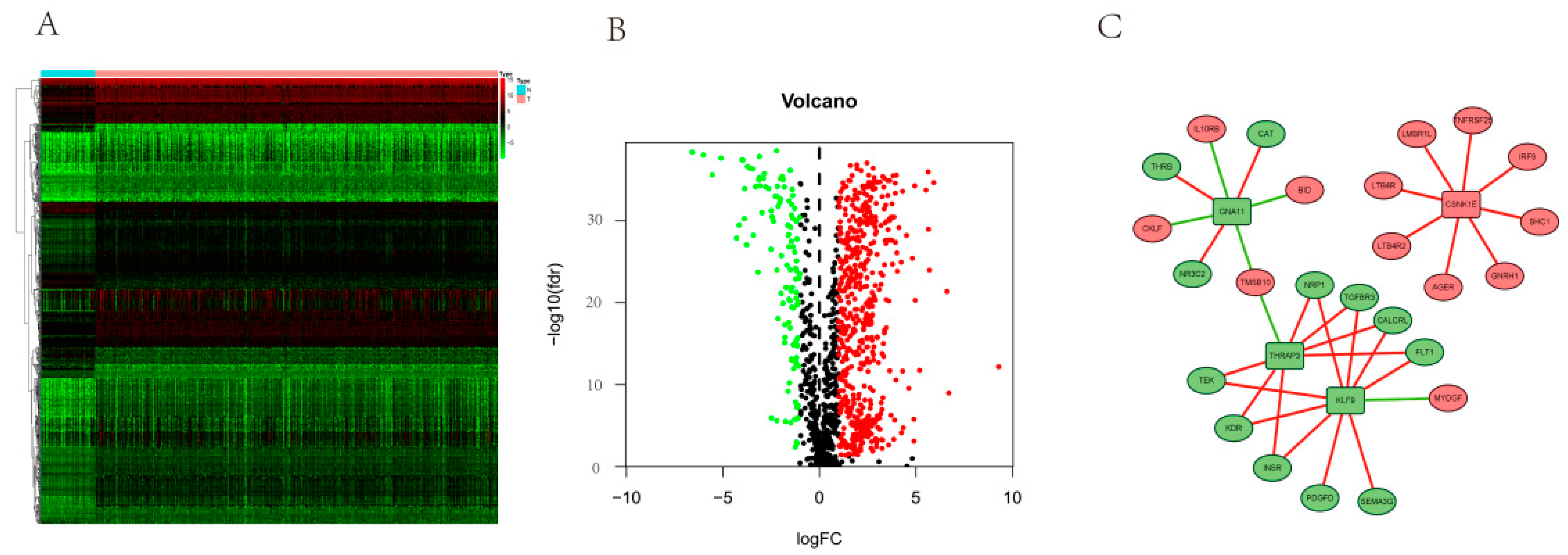
3.1. DERGs Identification
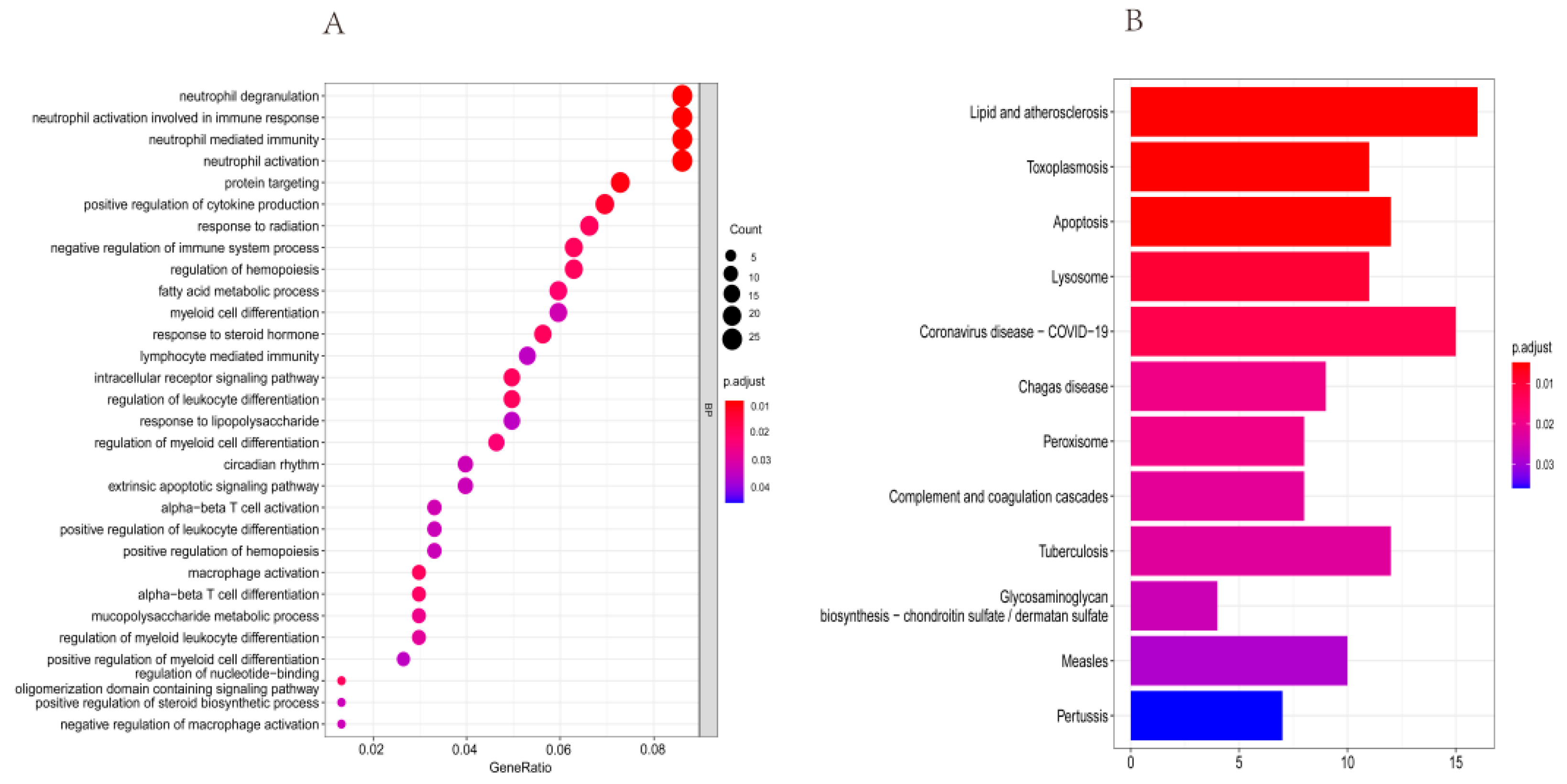
3.2. Functional Enrichment Analyses of DERGs
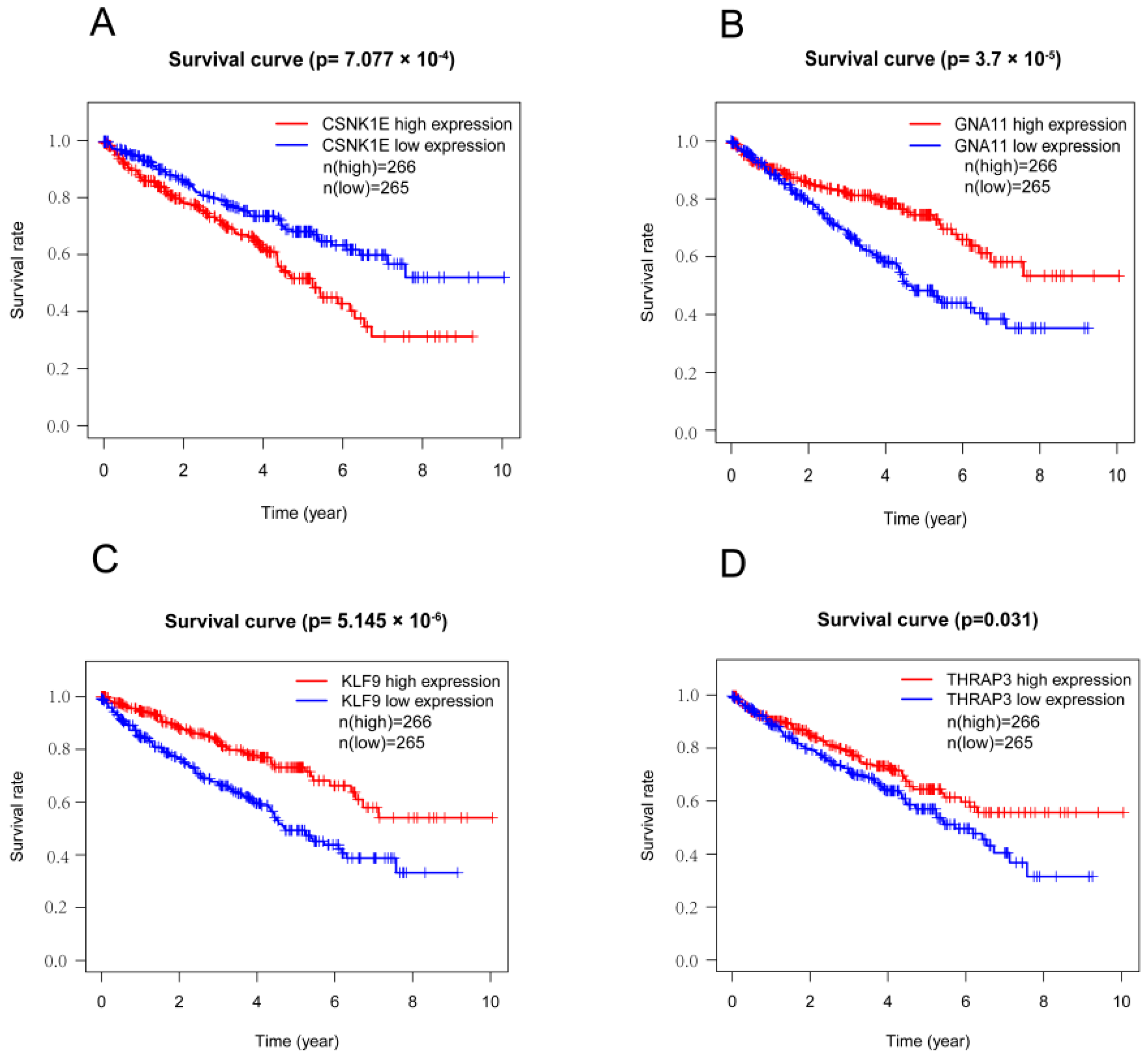
3.3. Survival Analyses of DERGs
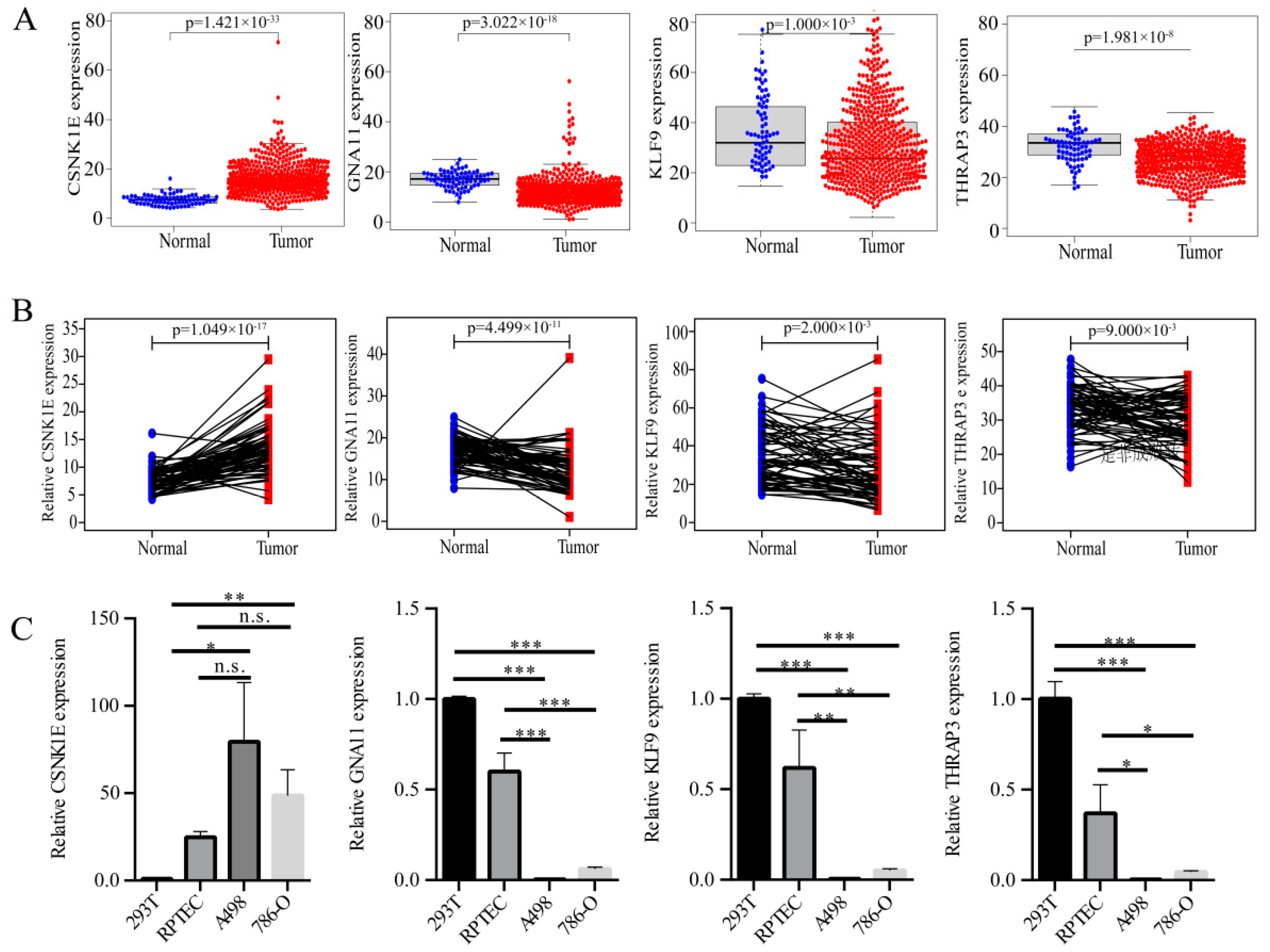
3.4. Expression Level Analyses of DERGs
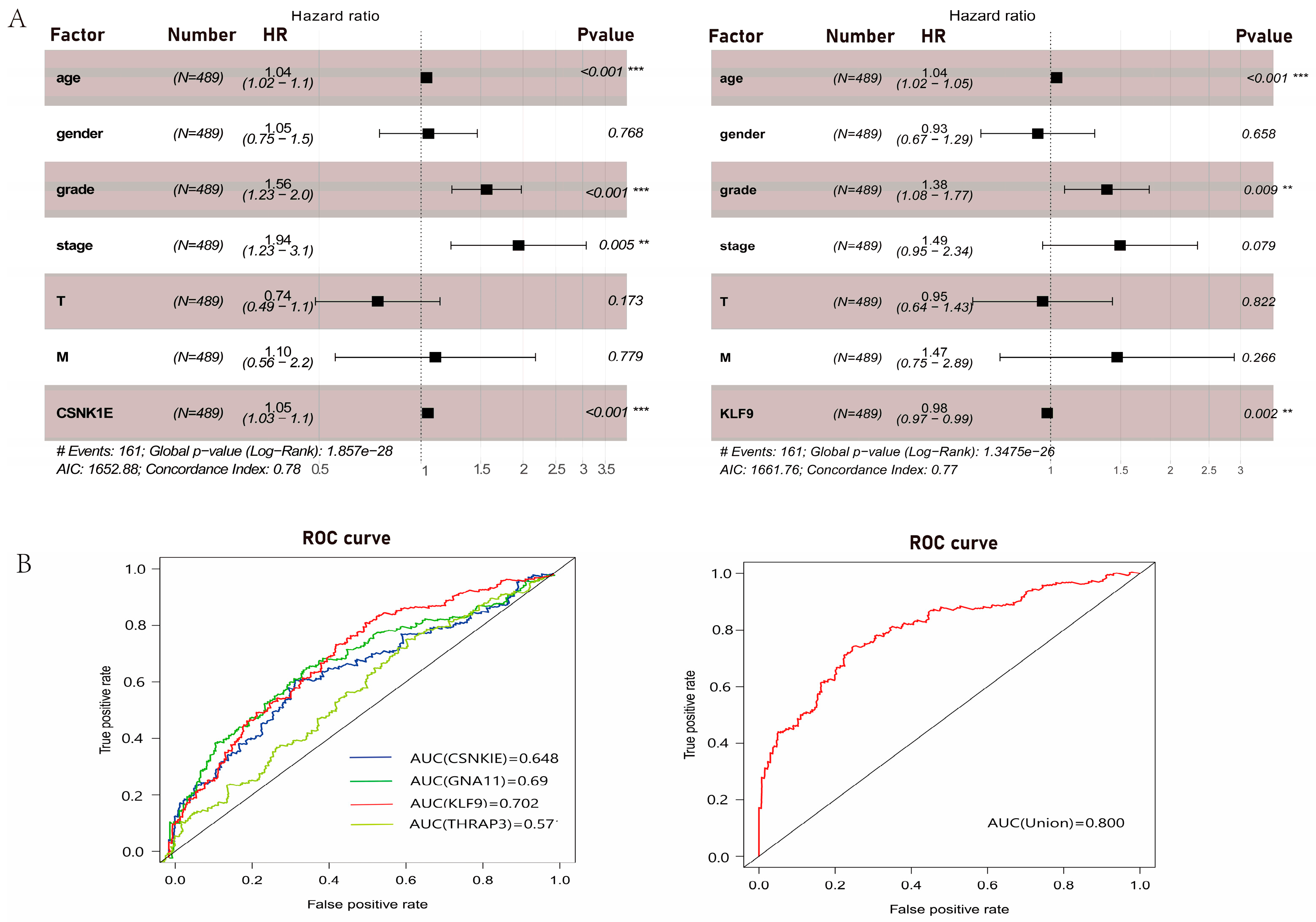
3.5. Clinical Relevance Analyses of DERGs
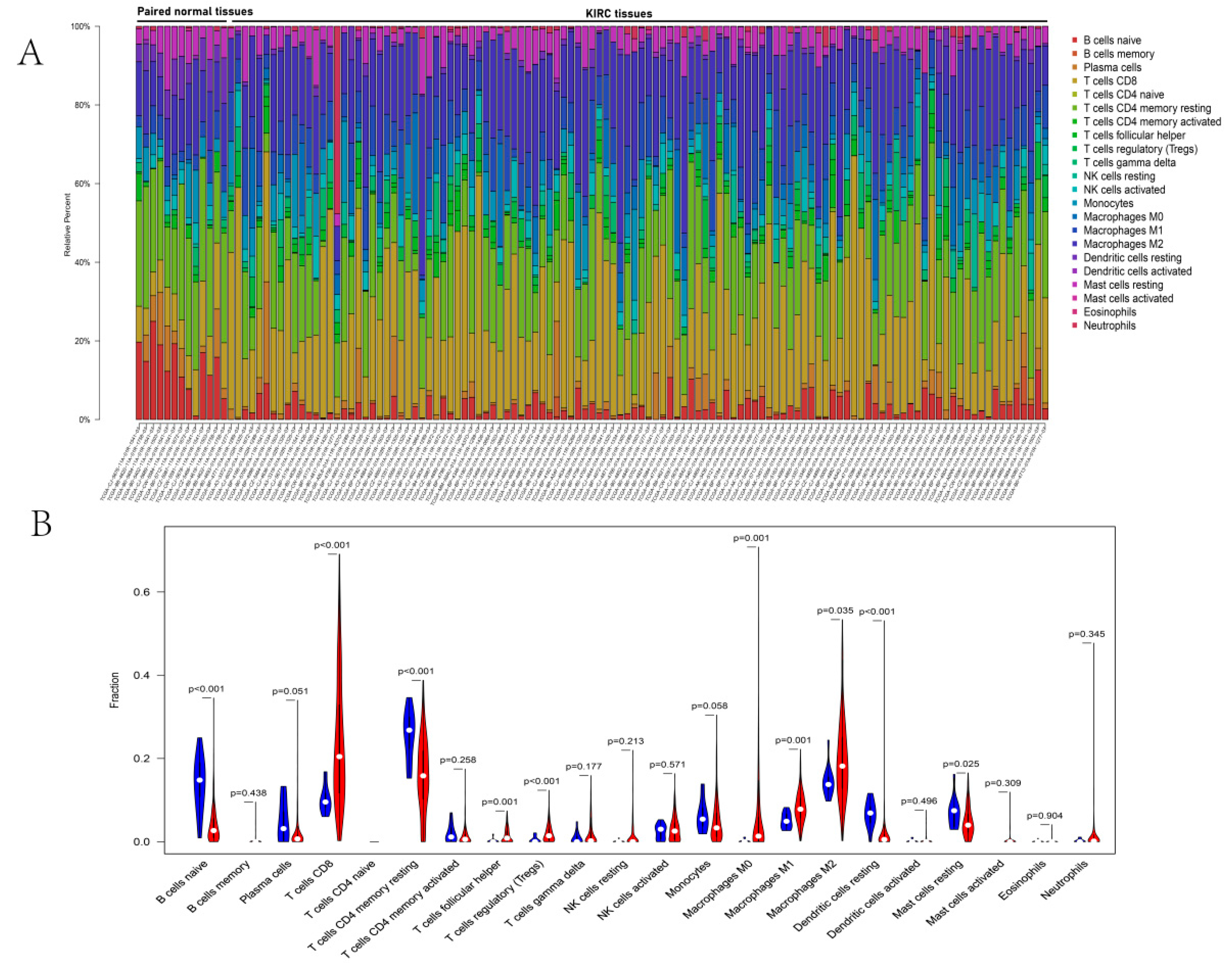
3.6. Immune Infiltration Analyses of DERGs
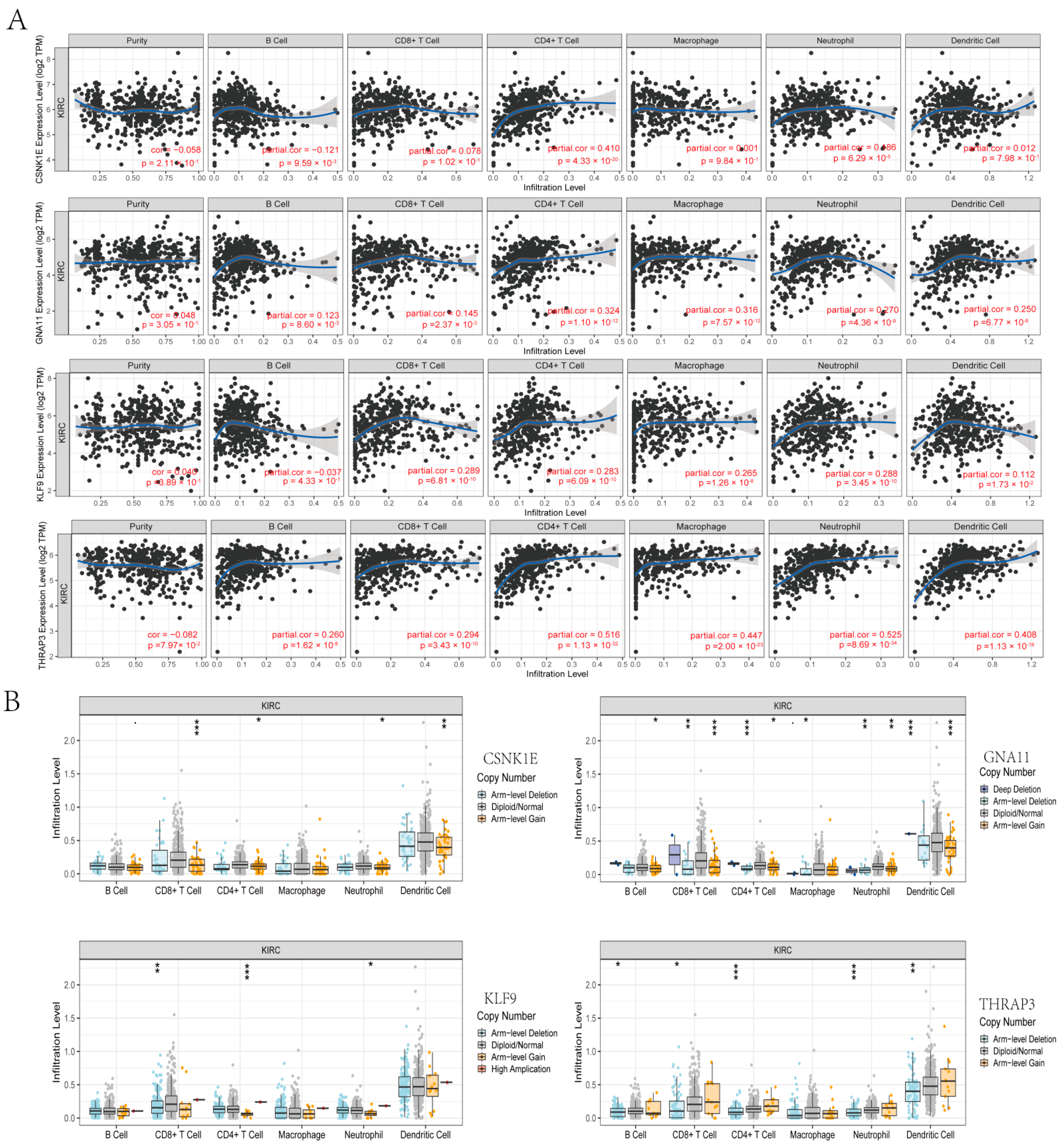
3.7. Correlation Analyses between DERGs and Immune Infiltration in KIRC
3.8. PPI Network Analyses of DERGs and Immune Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, Y.; Zhang, G. Identification of a Necroptosis-Related Prognostic Index and Associated Regulatory Axis in Kidney Renal Clear Cell Carcinoma. Int. J. Gen. Med. 2022, 3, 5407–5423. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, B.; Bensalah, K.; Canfield, S.; Dabestani, S.; Hofmann, F.; Hora, M.; Kuczyk, M.A.; Lam, T.; Marconi, L.; Merseburger, A.S.; et al. EAU Guidelines on Renal Cell Carcinoma: 2014 Update. Eur. Urol. 2015, 67, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Tao, W.; Guo, X.; Jing, C.; Zhang, M.; Wang, Z.; Kong, F.; Suo, N.; Jiang, S.; Wang, H. Construction of a Lactate-Related Prognostic Signature for Predicting Prognosis, Tumor Microenvironment, and Immune Response in Kidney Renal Clear Cell Carcinoma. Front. Immunol. 2022, 13, 818984. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Kannourakis, G.; Prithviraj, P.; Ahmed, N. Precision Medicine: An Optimal Approach to Patient Care in Renal Cell Carcinoma. Front. Med. 2022, 14, 766869. [Google Scholar] [CrossRef] [PubMed]
- Mollica, V.; Santoni, M.; Matrana, M.R.; Basso, U.; De Giorgi, U.; Rizzo, A.; Maruzzo, M.; Marchetti, A.; Rosellini, M.; Bleve, S.; et al. Concomitant Proton Pump Inhibitors and Outcome of Patients Treated with Nivolumab Alone or Plus Ipilimumab for Advanced Renal Cell Carcinoma. Target. Oncol. 2021, 17, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Gamble, K.L.; Berry, R.; Frank, S.J.; Young, M.E. Circadian Clock Control of Endocrine Factors. Nat. Rev. Endocrinol. 2014, 10, 466–475. [Google Scholar] [CrossRef] [Green Version]
- Kiessling, S.; Beaulieu-Laroche, L.; Blum, I.D.; Landgraf, D.; Welsh, D.K.; Storch, K.-F.; Labrecque, N.; Cermakian, N. Enhancing Circadian Clock Function in Cancer Cells Inhibits Tumor Growth. BMC. Biol. 2017, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Chen, Z.; Jiang, T.; Yang, X.; Du, Y.; Liang, J.; Wang, L.; Xi, J.; Lin, M.; Feng, M. Circadian Rhythm-Associated Clinical Relevance and Tumor Microenvironment of Non-small Cell Lung Cancer. J. Cancer 2021, 12, 2582–2597. [Google Scholar] [CrossRef]
- Wang, Z.; Su, G.; Dai, Z.; Meng, M.; Zhang, H.; Fan, F.; Liu, Z.; Zhang, L.; Weygant, N.; He, F.; et al. Circadian Clock Genes Promote Glioma Progression by Affecting Tumour Immune Infiltration and Tumour Cell Proliferation. Cell Prolif. 2021, 54, e12988. [Google Scholar] [CrossRef]
- Liu, H.; Gao, Y.; Hu, S.; Fan, Z.; Wang, X.; Li, S. Bioinformatics Analysis of Differentially Expressed Rhythm Genes in Liver Hepatocellular Carcinoma. Front. Genet. 2021, 12, 680528. [Google Scholar] [CrossRef]
- Zhou, L.; Luo, Z.; Li, Z.; Huang, Q. Circadian Clock is Associated with Tumor Microenvironment in Kidney Renal Clear Cell Carcinoma. Aging 2020, 14, 14620–14632. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Huang, S.-P.; Hsueh, Y.-M.; Xhen, L.-C.; Lu, T.-L.; Bao, B.-Y. Genetic Analysis Identifies the Role of HLF in Renal Cell Carcinoma. Cancer Genom. Proteomics. 2020, 17, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Shui, K.; Zhang, Y.; Lv, Y.; Deng, W.; Ullah, S.; Zhang, L.; Xue, Y. CGDB: A database of circadian genes in eukaryotes. Nucleic Acids Res. 2017, 45, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.A.; Schmithals, C.; von Harten, M.; Piiper, A.; Korf, H.W.; von Gall, C. Time-Dependent Changes in Proliferation, DNA Damage and Clock Gene Expression in Hepatocellular Carcinoma and Healthy Liver of a Transgenic Mouse Model. Int. J. Cancer. 2021, 148, 226–237. [Google Scholar] [CrossRef]
- Li, B.; Yang, X.; Li, J.; Yang, Y.; Yan, Z.; Zhang, H.; Mu, J. Elevated Expression of CLOCK is Associated with Poor Prognosis in Hepatocellular Carcinoma. Chin. J. Cell. Mol. Immunol. 2018, 34, 163–168. [Google Scholar]
- Luo, Y.; Wang, F.; Chen, L.-A.; Chen, X.-W.; Chen, Z.-J.; Liu, P.-F.; Li, F.-F.; Li, C.-Y.; Liang, W. Deregulated Expression of Cry1 and Cry2 in Human Gliomas. Asian Pac. J. Cancer Prev. 2012, 13, 5725–5728. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Hanada, K.; Mizokami, M.; Yeo, A.E.T.; Shih, J.W.-K.; Gojobori, T.; Alter, H.J. A Comparison of the Molecular Clock of Hepatitis C Virus in the United States and Japan Predicts that Hepatocellular Carcinoma Incidence in the United States will Increase over the Next Two Decades. Proc. Natl. Acad. Sci. USA 2002, 99, 15584–15589. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, L.; Wang, Y. Effects of Per2 Overexpression on Growth Inhibition and Metastasis, and on MTA1, nm23-H1 and the Autophagy-Associated PI3K/PKB Signaling Pathway in Nude Mice Xenograft Models of Ovarian Cancer. Mol. Med. Rep. 2016, 13, 4561–4568. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.C.; Niu, Z.F.; Ma, H.; Cao, S.Z.; Hao, S.C.; Liu, Z.T.; Wang, F. Deregulated Expression of the Per1 and Per2 in Human Gliomas. Can. J. Neurol. Sci. 2010, 37, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Wang, S.; Zhou, F.; Wan, S.; Yang, Y.; Huang, X.; Xing, J. Functional Polymorphisms in the NPAS2 Gene are Associated with Overall Survival in Transcatheter Arterial Chemoembolization-Treated Hepatocellular Carcinoma Patients. Cancer Sci. 2014, 105, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Yuan, P.; Yang, T.; Mu, J.; Zhao, J.; Yang, Y.I.; Yan, Z.; Li, J. Circadian Clock Gene NPAS2 Promotes Reprogramming of Glucose Metabolism in Hepatocellular Carcinoma Cells. Cancer Lett. 2020, 469, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Yang, T.; Mu, J.; Zhao, J.; Yang, Y.; Yan, Z.; Li, J. Corrigendum to “Circadian Clock Gene NPAS2 Promotes Reprogramming of Glucose Metabolism in Hepatocellular Carcinoma Cells”. Cancer Lett. 2021, 521, 322–323. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.L.; Luo, H.Y.; Yang, J.; Wu, W.J.; Chen, D.L.; Huang, P.; Xu, R.H. Overexpression of the Circadian Clock Gene Bmal1 Increases Sensitivity to Oxaliplatin in Colorectal Cancer. Clin. Cancer Res. 2014, 20, 1042–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Sun, H.; Zhang, S.; Yang, X.; Zhang, G.; Su, T. Overexpression of PER3 Inhibits Self-Renewal Capability and Chemoresistance of Colorectal Cancer Stem-Like Cells via Inhibition of Notch and beta-Catenin. Signal. Oncol. Res. 2017, 25, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Knippschild, U.; Gocht, A.; Wolff, S.; Huber, N.; Löhler, J.; Stöter, M. The Casein Kinase 1 Family: Participation in Multiple Cellular Processes in Eukaryotes. Cell. Signal. 2005, 17, 675–689. [Google Scholar] [CrossRef]
- Varghese, R.T.; Liang, Y.; Guan, T.; Franck, C.T.; Kelly, D.F.; Sheng, Z. Survival Kinase Genes Present Prognostic Significance in Glioblastoma. Oncotarget 2016, 7, 20140–20151. [Google Scholar] [CrossRef] [Green Version]
- Huisman, S.A.; Ahmadi, A.R.; Ijzermans, J.N.M.; Verhoef, C.; van der Horst, G.; De Bruin, R.W.F. Disruption of Clock Gene Expression in Human Colorectal Liver Metastases. Tumor Biol. 2016, 37, 13973–13981. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.-H.; Lin, Y.-M.; Yeh, C.-M.; Chen, C.-J.; Chen, M.-W.; Hung, H.-F.; Yeh, K.-T.; Yang, S.-F. Casein Kinase 1 Epsilon Expression Predicts Poorer Prognosis in Low T-Stage Oral Cancer Patients. Int. J. Mol. Sci. 2014, 15, 2876–2891. [Google Scholar] [CrossRef] [Green Version]
- Wan, B.; Liu, B.; Huang, Y.; Yu, G.; Lv, C. Prognostic Value of Immune-Related Genes in CLear Cell Renal Cell Carcinoma. Aging 2019, 11, 11474–11489. [Google Scholar] [CrossRef]
- Wu, H.-H.; Yan, X.; Chen, Z.; Du, G.-W.; Bai, X.-J.; Tuoheti, K.; Liu, T.-Z. GNRH1 and LTB4R Might be Novel Immune-Related Prognostic Biomarkers in Clear Cell Renal Cell Carcinoma (ccRCC). Cancer Cell Int. 2021, 21, 1–14. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Zhao, E.; Tan, Y.; Geng, B.; Kang, C.; Li, X. PTRF/CAVIN1, Regulated by SHC1 Through the EGFR PathWay, is Found in Urine Exosomes as a Potential Biomarker of ccRCC. Carcinogenesis 2019, 41, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Spörl, F.; Korge, S.; Jürchott, K.; Wunderskirchner, M.; Schellenberg, K.; Heins, S.; Specht, A.; Stoll, C.; Klemz, R.; Maier, B.; et al. Krüppel-Like Factor 9 is A Circadian Transcription Factor in Human Epidermis That Controls Proliferation of Keratinocytes. Proc. Natl. Acad. Sci. USA 2012, 109, 10903–10908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jager, M.J.; Shields, C.L.; Cebulla, C.M.; Abdel-Rahman, M.H.; Grossniklaus, H.E.; Stern, M.H.; Carvajal, R.D.; Belfort, R.N.; Jia, R.; Shields, J.A.; et al. Uveal Melanoma. Nat. Rev. Dis. Primers. 2020, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Zhou, F.; Wang, D.; Wu, M.; Zhou, W.; Zou, Y.; Li, J.; Wu, L.; Yin, X. Expression of KLF9 in Pancreatic Cancer and Its Effects on the Invasion, Migration, Apoptosis, Cell Cycle Distribution, and Proliferation of Pancreatic Cancer Cell Lines. Oncol. Rep. 2018, 40, 3852–3860. [Google Scholar] [CrossRef] [Green Version]
- Dulong, S.; Ballesta, A.; Okyar, A.; Lévi, F. Identification of Circadian Determinants of Cancer Chronotherapy through In Vitro Chronopharmacology and Mathematical Modeling. Mol. Cancer Ther. 2015, 14, 2154–2164. [Google Scholar] [CrossRef] [Green Version]
- Lévi, F.; Okyar, A.; Dulong, S.; Innominato, P.F.; Clairambault, J. Circadian Timing in Cancer Treatments. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 377–421. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Xiang, Y.; Ozguc, F.M.; Kim, Y.; Liu, C.J.; Park, P.K.; Hu, Q.; Diao, L.; Lou, Y.; Lin, C.; et al. The Genomic Landscape and Pharmacogenomic Interactions of Clock Genes in Cancer Chronotherapy. Cell Syst. 2018, 6, 314–328. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Cheng, B.; Xie, M.; Chen, Y.; Zhao, J.; Zhou, X.; Chen, L. Circadian Clock Gene Bmal1 Inhibits Tumorigenesis and Increases Paclitaxel Sensitivity in Tongue Squamous Cell Carcinoma. Cancer Res. 2017, 77, 532–544. [Google Scholar] [CrossRef] [Green Version]
- Zhanfeng, N.; Yanhui, L.; Zhou, F.; Shaocai, H.; Guangxing, L.; Hechun, X. Circadian genes Per1 and Per2 Increase Radiosensitivity of Glioma in vivo. Oncotarget 2015, 6, 9951–9958. [Google Scholar] [CrossRef] [Green Version]
- Möller-Levet, C.S.; Archer, S.N.; Bucca, G.; Laing, E.E.; Slak, A.; Kabiljo, R.; Lo, J.C.Y.; Santhi, N.; von Schantz, M.; Smith, C.P.; et al. Effects of Insufficient Sleep on Circadian Rhythmicity and Expression Amplitude of the Human Blood Transcriptome. Proc. Natl. Acad. Sci. USA 2013, 110, E1132–E1141. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Zhu, S.; Zeng, L.; Liu, J.; Kang, R.; Yang, M.; Cao, L.; Wang, H.; Billiar, T.R.; Jiang, J.; et al. The Circadian Clock Controls Immune Checkpoint Pathway in Sepsis. Cell Rep. 2018, 24, 366–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Hsu, W.-H.; Chang, A.; Tan, Z.; Lan, Z.; Zhou, A.; Spring, D.J.; Lang, F.F.; Wang, Y.A.; DePinho, R.A. Circadian Regulator CLOCK Recruits Immune-Suppressive Microglia into the GBM Tumor Microenvironment. Cancer Discov. 2020, 10, 371–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollinger, T.; Leutz, A.; Leliavski, A.; Skrum, L.; Kovac, J.; Bonacina, L.; Benedict, C.; Lange, T.; Westermann, J.; Oster, H.; et al. Circadian Clocks in Mouse and Human CD4+ T Cells. PLoS ONE 2011, 6, e29801. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Univariate Analysis | ||
|---|---|---|---|
| HR | 95% CI | p | |
| age | 1.03 | 1.01–1.04 | <0.001 |
| grade | 2.29 | 1.85–2.83 | <0.001 |
| stage | 1.88 | 1.64–2.16 | <0.001 |
| gender | 0.93 | 0.67–1.28 | >0.05 |
| T | 1.94 | 1.63–2.29 | <0.001 |
| M | 4.28 | 3.10–5.90 | <0.001 |
| CSNK1E | 1.03 | 1.02–1.05 | <0.001 |
| KLF9 | 0.96 | 0.95–0.97 | <0.001 |
| THRAP3 | 0.97 | 0.95–0.99 | <0.05 |
| GNA11 | 0.92 | 0.88–0.96 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, X.; Wang, Q.; Ding, K.; Chen, X.; Zhao, Y.; Gao, Y.; Wang, Y. Effects and Prognostic Values of Circadian Genes CSNK1E/GNA11/KLF9/THRAP3 in Kidney Renal Clear Cell Carcinoma via a Comprehensive Analysis. Bioengineering 2022, 9, 306. https://doi.org/10.3390/bioengineering9070306
Li S, Wang X, Wang Q, Ding K, Chen X, Zhao Y, Gao Y, Wang Y. Effects and Prognostic Values of Circadian Genes CSNK1E/GNA11/KLF9/THRAP3 in Kidney Renal Clear Cell Carcinoma via a Comprehensive Analysis. Bioengineering. 2022; 9(7):306. https://doi.org/10.3390/bioengineering9070306
Chicago/Turabian StyleLi, Shujing, Xianggang Wang, Qingqing Wang, Kaixin Ding, Xin Chen, Yun Zhao, Yu Gao, and Yuanyuan Wang. 2022. "Effects and Prognostic Values of Circadian Genes CSNK1E/GNA11/KLF9/THRAP3 in Kidney Renal Clear Cell Carcinoma via a Comprehensive Analysis" Bioengineering 9, no. 7: 306. https://doi.org/10.3390/bioengineering9070306