
Analytical Studies of Antimicrobial Peptides as Diagnostic Biomarkers for the Detection of Bacterial and Viral Pneumonia



Abstract
:1. Introduction
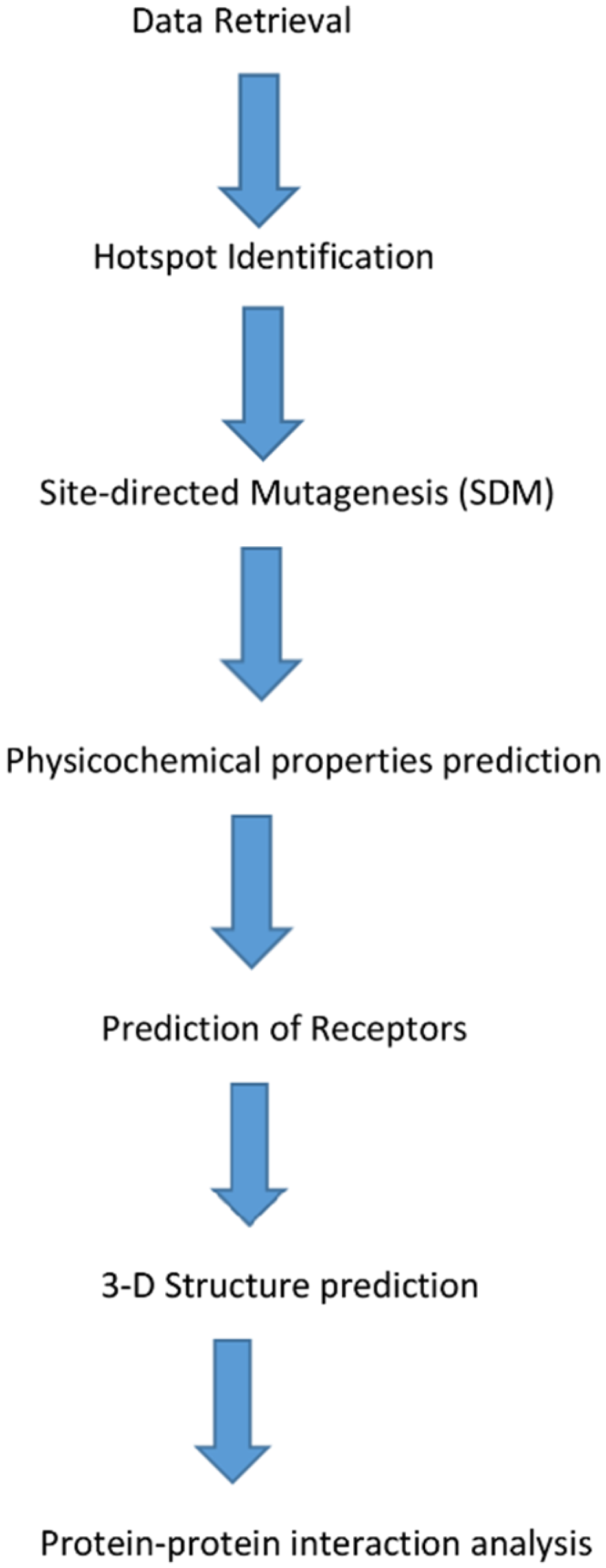
2. Materials and Methods
2.1. Data Retrieval
2.2. Identification of Hotspot Residues
2.3. In Silico Site-Directed Mutagenesis (SDM)
2.4. Physicochemical Properties
2.5. Prediction of Receptors
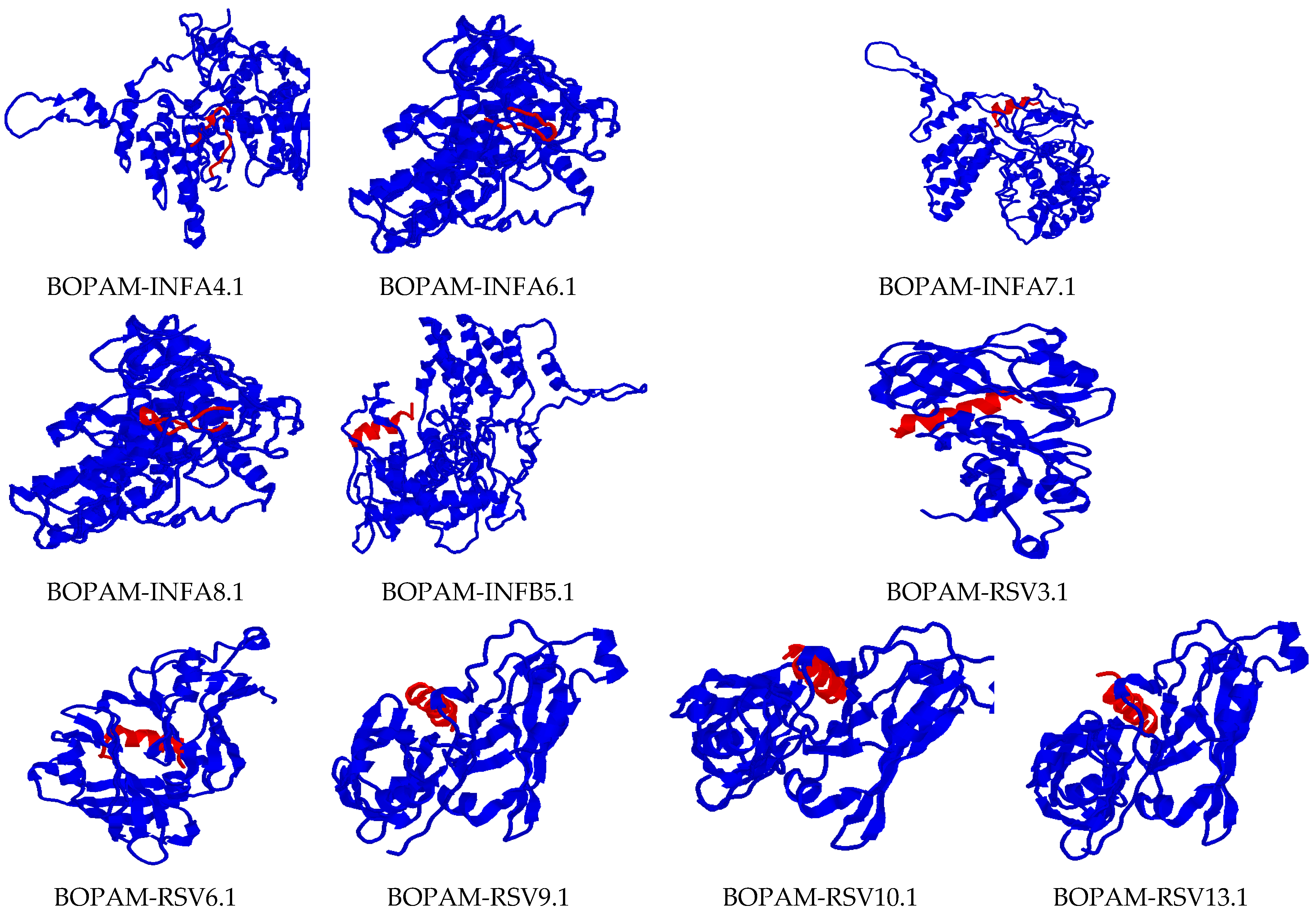
2.6. In Silico 3D Structure Prediction
2.7. In Silico Protein–Protein Interaction Study
3. Results
3.1. Knowledge-Based FADE and Contacts (KFC) Analysis
3.2. In Silico Site-Directed Mutagenesis of the AMPs
3.3. Physicochemical Properties of the Derivative AMPs
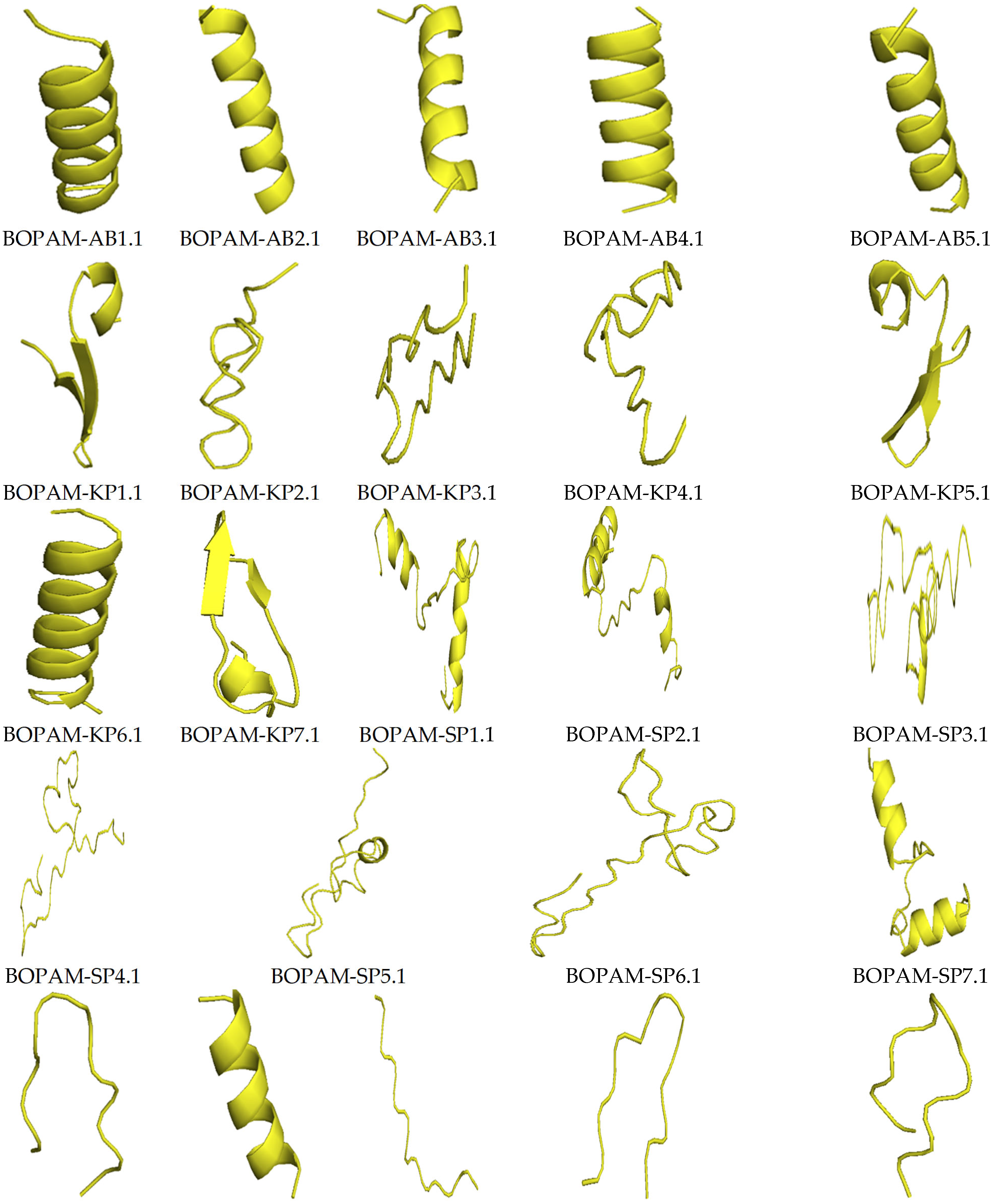
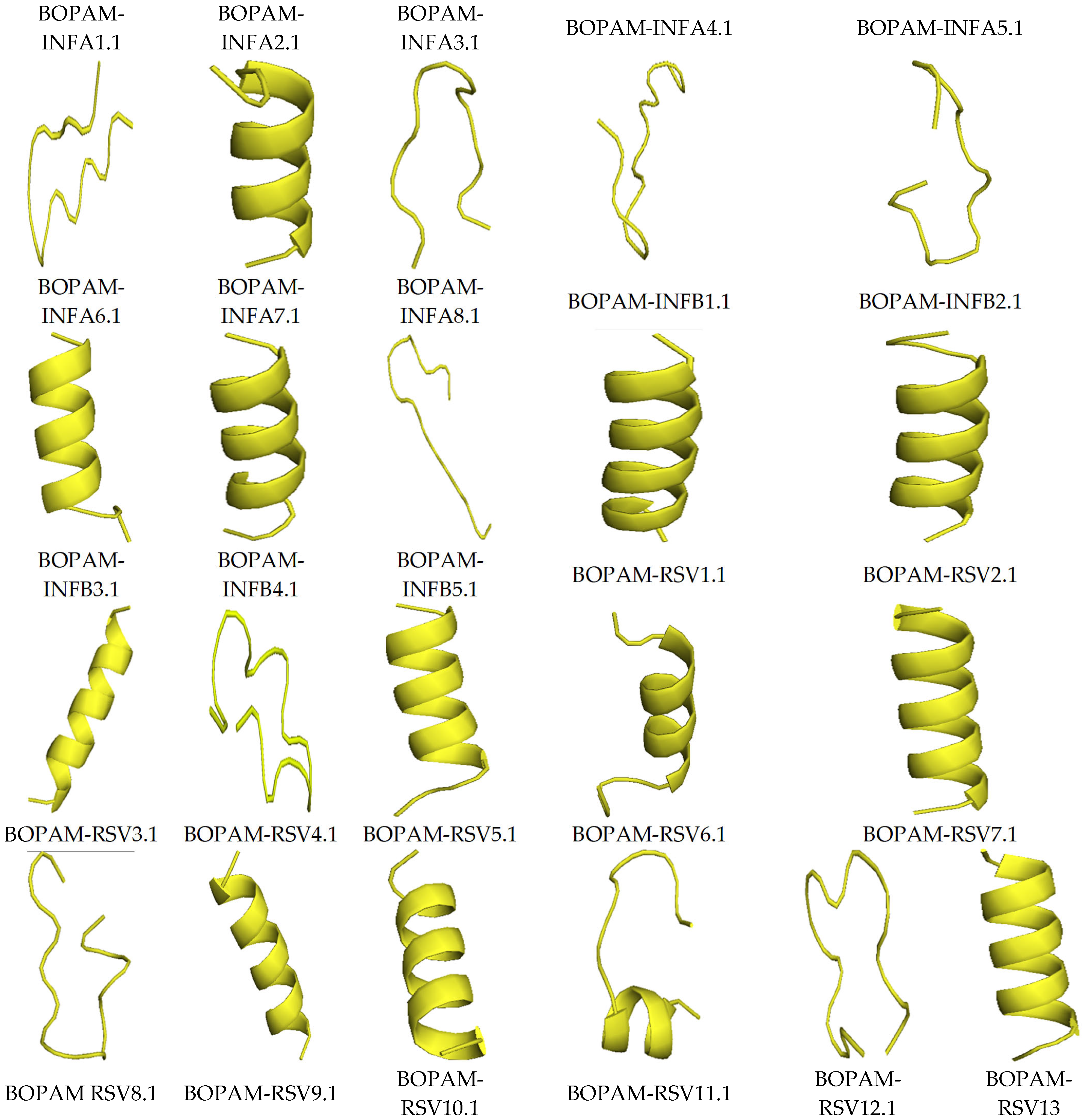
3.4. Structure Prediction Using I-TASSER
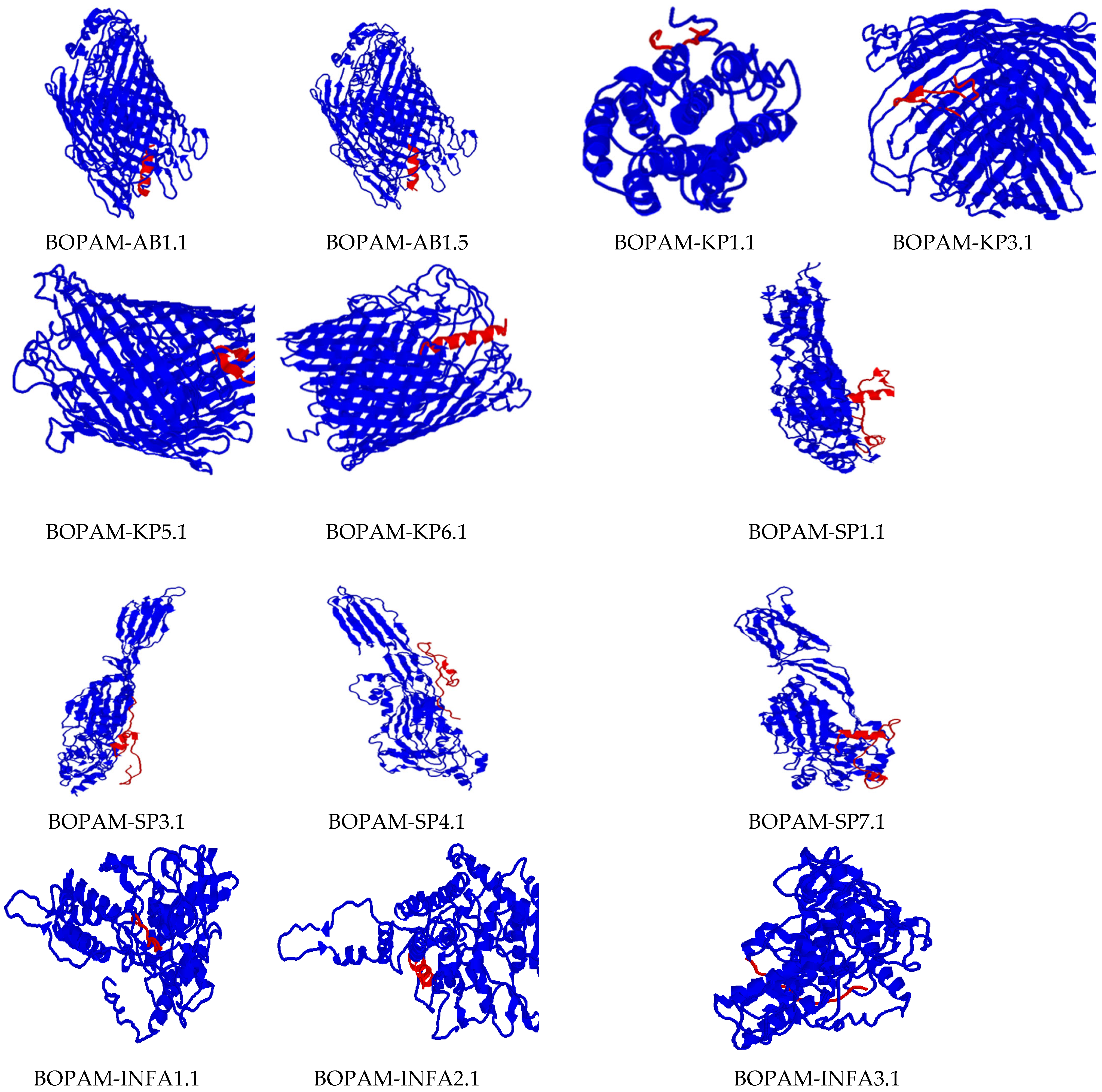
3.5. Docking Interaction Analysis of the Derivative AMPs with Bacterial and Viral Receptors Using PATCHDOCK
4. Discussion
5. Conclusions
6. Future Work
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics approval and Consent to Participate
Consent for Publication
References
- Peres, M.A.; Macpherson, L.M.; Weyant, R.J.; Daly, B.; Venturelli, R.; Mathur, M.R.; Listl, S.; Celeste, R.K.; Guarnizo-Herreño, C.C.; Kearns, C.; et al. Oral diseases: A global public health challenge. Lancet 2019, 394, 249–260. [Google Scholar] [CrossRef]
- Atkinson, T.P.; Balish, M.F.F.; Waites, K.B. Epidemiology, clinical manifestations, pathogenesis and laboratory detection of Mycoplasma pneumoniae infections. FEMS Microbiol. Rev. 2008, 32, 956–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinton, L.J.; Walkey, A.J.; Mizgerd, J.P. Integrative physiology of pneumonia. Physiol. Rev. 2018, 98, 1417–1464. [Google Scholar] [CrossRef] [PubMed]
- Shin, B.; Kim, S.-H.; Yong, S.J.; Lee, W.-Y.; Park, S.; Lee, S.J.; Lee, S.J.; Lee, M.K. Early readmission and mortality in acute exacerbation of chronic obstructive pulmonary disease with community-acquired pneumonia. Chronic Respir. Dis. 2018, 16, 1479972318809480. [Google Scholar] [CrossRef]
- Libby, P.; Ridker, P.M.; Hansson, G.K.; Leducq Transatlantic Network On Atherothrombosis. Inflammation in atherosclerosis: From pathophysiology to practice. J. Am. Coll. Cardiol. 2009, 54, 2129–2138. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Youn, Y.S.; Rhim, J.W.; Kang, J.H.; Lee, K.Y. Early serologic diagnosis of Mycoplasma pneumoniae pneumonia: An observational study on changes in titers of specific-IgM antibodies and cold agglutinins. Medicine 2016, 95, e3605. [Google Scholar] [CrossRef]
- Bakare, O.O.; Fadaka, A.O.; Klein, A.; Pretorius, A. Dietary effects of antimicrobial peptides in therapeutics. All Life 2020, 13, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919. [Google Scholar]
- Guryanova, S.V.; Ovchinnikova, T.V. Immunomodulatory and allergenic properties of antimicrobial peptides. Int. J. Mol. Sci. 2022, 23, 2499. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Ovchinnikova, T.V. Improved strategy for recombinant production and purification of antimicrobial peptide tachyplesin I and its analogs with high cell selectivity. Biotechnol. Appl. Biochem. 2017, 64, 35–42. [Google Scholar] [CrossRef]
- Lockhart, D.J.; Winzeler, E. Genomics, gene expression and DNA arrays. Nature 2000, 405, 827. [Google Scholar] [CrossRef] [PubMed]
- Edelheit, O.; Hanukoglu, A.; Hanukoglu, I. Simple and efficient site-directed mutagenesis using two single-primer reactions in parallel to generate mutants for protein structure-function studies. BMC Biotechnol. 2009, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlgemuth, R. Session 1: Technical Biochemistry Lectures L1. 2. ChemBioChem 2009, 10, 1888–1896. [Google Scholar]
- Goh, K.M.; Hong, G.P.; Chyi, N.H.; Ng, P.; Piaw, C.K.; Rahman, R.N.Z.R.A. Trends and tips in protein engineering, a review. J. Teknol. 2012, 59, 21–31. [Google Scholar]
- Bakare, O.O.; Keyster, M.; Pretorius, A. Identification of biomarkers for the accurate and sensitive diagnosis of three bacterial pneumonia pathogens using in silico approaches. BMC Mol. Cell Biol. 2020, 21, 82. [Google Scholar] [CrossRef]
- Bakare, O.O.; Keyster, M.; Pretorius, A. Building HMM and molecular docking analysis for the sensitive detection of anti-viral pneumonia antimicrobial peptides (AMPs). Sci. Rep. 2021, 11, 20621. [Google Scholar] [CrossRef]
- Williams, M.E.; Tincho, M.; Gabere, M.; Uys, A.; Pretorius, M.M.A.A. Molecular validation of putative antimicrobial peptides for improved human immunodeficiency virus diagnostics via HIV protein p24. J. AIDS Clin. Res. 2016, 7, 571. [Google Scholar] [CrossRef] [Green Version]
- Darnell, S.J.; Legault, L.H.; Mitchell, J.C. KFC Server: Interactive forecasting of protein interaction hot spots. Nucleic Acids Res. 2008, 36 (Suppl. 2), W265–W269. [Google Scholar] [CrossRef] [Green Version]
- Fu, E.; Lutz, B.; Kauffman, P.; Yager, P. Controlled reagent transport in disposable 2D paper networks. Lab Chip 2010, 10, 918–920. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, G. APD: The antimicrobial peptide database. Nucleic Acids Res. 2004, 32 (Suppl. 1), D590–D592. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, Y. I-TASSER server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Chan, H.S.; Hu, Z. Using PyMOL as a platform for computational drug design. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2017, 7, e1298. [Google Scholar] [CrossRef]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33 (Suppl. 2), W363–W367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, M. Monoclonal antibodies as diagnostics: An appraisal. Indian J. Pharm. Sci. 2010, 72, 12. [Google Scholar] [CrossRef] [Green Version]
- Biro, J. Amino acid size, charge, hydropathy indices and matrices for protein structure analysis. Theor. Biol. Med Model. 2006, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Tachi, T.; Epand, R.F.; Epand, R.M.; Matsuzaki, K. Position-dependent hydrophobicity of the antimicrobial magainin peptide affects the mode of peptide−lipid interactions and selective toxicity. Biochemistry 2002, 41, 10723–10731. [Google Scholar] [CrossRef]
- Cuthbert, T.J.; Hisey, B.; Harrison, T.D.; Trant, J.F.; Gillies, E.R.; Ragogna, P.J. Surprising antibacterial activity and selectivity of hydrophilic polyphosphoniums featuring sugar and hydroxy substituents. Angew. Chem. Int. Ed. 2018, 57, 12707–12710. [Google Scholar] [CrossRef]
- Dathe, M.; Nikolenko, H.; Meyer, J.; Beyermann, M.; Bienert, M. Optimization of the antimicrobial activity of magainin peptides by modification of charge. FEBS Lett. 2001, 501, 146–150. [Google Scholar] [CrossRef] [Green Version]
- Jodoin, J.; Hincke, M.T. Histone H5 is a potent antimicrobial agent and a template for novel antimicrobial peptides. Sci. Rep. 2018, 8, 2411. [Google Scholar] [CrossRef]
- Bakare, O.O.; Gokul, A.; Keyster, M. PR-1-like protein as a potential target for the identification of Fusarium oxysporum: An in silico approach. Biotech 2021, 10, 8. [Google Scholar] [CrossRef]
- Tincho, M.; Gabere, M.N.; Pretorius, A. In silico identification and molecular validation of putative antimicrobial peptides for HIV therapy. J. AIDS Clin. Res. 2016, 7, 606. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/N | Interacting Amino Acids of the Receptors | Interacting Amino Acids of the Putative AMPs |
|---|---|---|
| 1 | Acinetobacter baumannii Outer Membrane Protein: Arg188A, Arg358A, Asn597A, Arg600A, Val612A, Asp644A, Gln646A, Asp739A, Gln787A, Leu789A, Asn792A, Asn794A | BOPAM-AB1: Phe1a, Leu2a, Val5a, Leu9a, Ser11a, Ser14a, Gly15a, Leu16a, Leu17a |
| 2 | Acinetobacter baumannii Outer Membrane Protein: Glu180A, Arg188A, Ser189A, Arg203A, Ser206A, Arg207A, Arg600A, Val645A, Gln646A, Tyr736A, Arg737A, Gln787A, Leu789A, Arg790A | BOPAM-AB2: Phe2a, Gly6a, Lys7a, Leu9a, Ser11a, Gly15a, Leu16a, Leu17a |
| 3 | Acinetobacter baumannii Outer Membrane Protein: Arg176A, Arg188A, Ser189A, Arg203A, Asp204A, Asp431A, Ile433A, Leu473A, Leu595A, Asn597A, Ala598A, Arg600A, Asp644A, Leu789A, Arg790A, Asn793A | BOPAM-AB3: Phe1a, Phe2a, Pro3a, Leu8a, Leu9a, Leu13a, Phe14a, Leu17a |
| 4 | Acinetobacter baumannii Outer Membrane Protein: Arg188A, Ser189A, Gln587A, Asn597A, Ala598A, Val612A, Asp644A, Gln646A, Phe648A, Tyr736A, Asp739A, Gln787A, Leu789A | BOPAM-AB4: Phe1a, Phe2a, Ile4a, Val5a, Lys7a, Leu8a, Leu9a, Lys10a, Ser14a |
| 5 | Acinetobacter baumannii Outer Membrane Protein: Thr184A, Arg188A, Ser189A, Arg600A, Val612A, Asp644A, Val645Aa, Gln646A, Phe648A, Tyr736A, Asp739A, Gln787A, Leu789A | BOPAM-AB5: Thr4a, Gly6a, Lys7a, Ala8a, Lys11a, Arg14a, Ala15a, Asn17a |
| 6 | Influenza A Virus Nucleoprotein: Ser165A, Leu166A, Arg267A, Gly268A, Val270A, His272A, Phe338A, Glu339A. Asp340A, Arg342A, Val343A, Pro453A, Ser457A, Leu479A, Tyr487A | BOPAM-INFA1: Pro2a, Phe4a, Ile5a, Asp6a, Gly7a, Gln8a, Val9a |
| 7 | Influenza A Virus Nucleoprotein: Ser165A, Leu264A, Ile265A, Arg267A, Asp340A, Arg342A, Val343A, Pro453A, Val456A, Ser457A, Phe458A, Pro477A, Leu479A, Tyr487A | BOPAM-INFA2: Pro2a, Val3a, Leu5a, Ile9a, Gln10a, Lys14a |
| 8 | Influenza A Virus Nucleoprotein: Glu339A, Arg342A, Val343A, Ala387A, Thr390A, Asn395A, Pro453A, Glu454A, Asp455A, Val456A, Ser457A, Phe458A, Gly462A, Val463A, Leu479A | BOPAM-INFA4: Phe4a, Ile5a, Asp6a, Gly7a, Gln8a, Val9a, Pro10a, Gln14a |
| 9 | Influenza A Virus Nucleoprotein: Leu166A, Arg267A, Val270A, His272A, Glu339A, Asp340A, Val343A, Ser457A, Phe458A, Val463A, Pro477A, Leu479A | BOPAM-INFA5: Thr1a, Thr3a, Phe4a, Ile5a, Val9a, Ile11a, Gln13a, Gln14a |
| 10 | Influenza A Virus Nucleoprotein: Ser165A, Arg267A, Glu339A, Asp340A, Val343A, Glu454A, Asp455A, Ser457A, Phe458A, Val463A, Pro477A, Leu479A | BOPAM-INFA6: Pro2a, Phe4a, Ile5a, Asp6a, Gly7a, Val9a, Pro12a |
| 11 | Influenza A Virus Nucleoprotein: Ser165A, Leu166A, Leu264A, Ile265A, Arg267A, Phe338A, Asp340A, Val343A, Asn395A, Pro453A, Asp455A, Val456A, Ser457A, Phe458A, Tyr487A | BOPAM-INFA7: Pro2a, Val3a, Ile4a, Asp6a, Gln10a, Val11a, Phe12a |
| 12 | Influenza A Virus Nucleoprotein: Ser165A, Ile265A, Arg267A, Gly268A, Val270A, His272A, Ala336A, Glu339A, Arg342A, Val343A, Ser344A, Ile347A, Ala387A, Arg389A, Pro453A, Phe458A | BOPAM-INFA8: Thr1a, Pro2a, Thr3a, Ile5a, Val5a, Glu13a, Gln14a |
| 13 | Influenza B Virus Nucleoprotein: Ser289A, Ala290A, Val323A, Val324A, Arg325A, Arg398A, Arg447A, Glu452A, Met503A, Ser507A, Gly514A | BOPAM-INFB1: Met1a, Val3a, Ser4a, Arg6a, Trp7a, Thr8a, Phe9a, Leu10a, Val12a |
| 14 | Influenza B Virus Nucleoprotein: Arg116A, Lys125A, Gly151A, Arg170A, Arg235A, Glu253A, Arg256A, Phe257A | BOPAM-INFB2: Ser4a, Arg6a, Thr8a, Phe9a, Met11a, Val12a, Pro13a, Pro14a |
| 15 | Influenza B Virus Nucleoprotein: Arg116A, Leu119A, Ala120A, Asp123A, Lys125A, Asp137A, Glu140A, Lys142A, Glu143A, Thr150A, Gly151A, Pro234A | BOPAM-INFB3: Leu1a, Asn2a, Pro5a, Gln6a, Leu7a, Leu8a, Leu10A, |
| 16 | Influenza B Virus Nucleoprotein: Arg116A, Leu119A, Ala120A, Asp123A, Lys125A, Asp137A, Glu140A, Thr150A, Gly152A, Thr153A, Pro234A, Pro422A, Ala423A | BOPAM-INFB4: Leu4a, Gln5a, Leu7a, Leu8a, Leu10a, Lys1a, Val12a, Pro13a, Leu15a |
| 17 | Influenza B Virus Nucleoprotein: Aeg116A Leu119A, Ala120A, Lys125A, Phe129A, Asp137A, Glu140A, Glu143A, Gly151A, Gly152A, Thr153A, His201A, Thr232A, Pro234A, His420A | BOPAM-INFB5: Thr2a, Ile3a, Leu4a, Leu6a, Leu7a, Leu10a, Lys11a, Gln14a, Leu15a |
| 18 | Influenza B Virus Nucleoprotein: Gln203A, Ala229A, Ile244A, Val322A, Val323A, Arg325A, Val328A, Ser330A, Tyr394A, Glu395A, Asp396A, Gly449A, Met508A, Gly514A, Ala516A | BOPAM-INFB6: Leu1a, Asn4a, Leu7a, Cys9a, Asn11a, Asn13a, Pro14a, Gln15a |
| 19 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Cys93A, Asn95A, Val96A, Tyr199A, Ser200A, Ile225A, Asp227A, Ala230A | BOPAM-RSV1: Ile1a, Ile5a, Glu7a, Glu8a, Cys12a, Lys15a, Phe16a |
| 20 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Ile92A, Cys93A, Asn95A, Val96A, Tyr199A, Ser200A, Leu203A, Ile225A, Asp227A | BOPAM-RSV2: Asn6a, Glu7a, Ile9a, Asp10a, Leu13a, Ser14a |
| 21 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Met40A, Ile92A, Cys93A, Asn95A, Tyr199A, Ser200A, Leu203A, Ile225A, Asp227A | BOPAM-RSV3: Asn1a, Asp4a, Val5a, Lys8a, Ile9a, Asn12a, Thr13a |
| 22 | Respiratory Syncytial Virus Chain A Protein: Leu27A, Ile30A, Arg31A, Asp34A | BOPAM-RSV4: Val2a, Glu4a, Ile5a, Ala7a, Asn11a |
| 23 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Ile92A, Cys93A, Leu128A, Thr130A, Tyr199A, Ser200A, Ile225A, Asp227A | BOPAM-RSV5: Ser6a, Ala7a, Gln8a, Asn10a, Lys11a, Asn15a |
| 24 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Pro39A, Met40A, Cys93A, Thr130A, Tyr199A, Ser200A, Leu203A, Val226A, Asp227A | BOPAM-RSV6: Lys1a, Gln4a, Ile5a, Ala8a, Ile9a, Gln12a |
| 25 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Cys93A, Asn95A, Thr130A, Tyr199A, Ser200A, Leu203A, Ile225A, Asp227A | BOPAM-RSV7: Lys6a, Asn10a, Gln11a, Ile13a, Asn14a |
| 26 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Pro39A, Met40A, Cys93A, Lys125A, Thr130A, Tyr199A, Ser200A, Leu203A, Ile225A, Asp227A | BOPAM-RSV8: Ile2a, Asn4a, Phe5a, Ser9a, Leu12a, Leu13a, Ser14a |
| 27 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Met40A, Cys93A, Asn95A, Lys125A, Leu128A, Thr130A, Tyr199A, Ser200A, Leu203A, Ile225A, Asp227A | BOPAM-RSV9: Ile2a, Ser3a, Lys4a, Thr6a, Asn7a, Asn10a, Thr11a |
| 28 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Met40A, Cys93A, Asn95A, Thr130A, Tyr199A, Ser200A, Leu203A, Ile225A, Asp227A | BOPAM-RSV10: Ser6a, Asn10a, Thr11a, Thr14a, Asn15a, Ile16a |
| 29 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Ser71A, Phe90A, Thr91A, Ile92A, Thr130A, Tyr199A, Ser200A, Leu203A, Ile225A, Val226A, Asp227A | BOPAM-RSV11: Asn1a, Val2a, Asp10a, Asn12a, Ala14a, Asp15a |
| 30 | Respiratory Syncytial Virus Chain A Protein: Glu1A, Lys27A, Asn56A, Asn58A, Arg79A, Pro111A, Cys112A, Glu113A, Ile114A | BOPAM-RSV12: Asn4a, Ile5a, Asn8a, Lys12a, Phe13a, Ile16a |
| 31 | Respiratory Syncytial Virus Chain A Protein: Trp37A, Tyr199A, Ser200A, Leu203A, Ile225A, Asp227A, Ala230A | BOPAM-RSV13: Leu5a, Glu7a, Lys8a, Asp11a, Arg12a |
| 32 | Streptococcus pneumoniae Pneumolysin: Thr57A, Ser58A, Asp59A, Met97A, Thr98A, Tyr99A, Ser100A, Lys196A, Ile198A, Thr201A, Ser203A, Asp205A, Ala206A, Asp212A, Ser239A, Ala241A, Ser330A, Thr332A, Phe335A, Val341A, Thr343A | BOPAM-SP1: Arg3a, Asp4a, Asp5a, Arg6a, Cys8a, Met12a, Ile24a, Thr26a, Phe27a, Ser34a, Ile38a, Cys39a, Asn43a, Gly44a |
| 33 | Streptococcus pneumoniae Pneumolysin: Ser58A, Asp59A, Thr98A, Tyr99A, Ser100A, Ile101A, Gln149A, Glu151A, Ile198A, Thr201A, Ser203A, Asp205A, Ser239A, Ala241A, Thr332A, Phe335A, Val341A, Thr343A | BOPAM-SP2: Arg3a, Asn4a, Cys8a, Met12a, Thr24a, Thr26a, Phe27a, His29a, Ser34a, Ile38a, Asn41a, Lys42a |
| 34 | Streptococcus pneumoniae Pneumolysin: Glu42A, Ser254A, Ser256A, Glu277A, Gln280A, Ile281A, Asn284A, Thr356A, Ala357A, Thr358A, Arg359A, Leu447A, Val448A, Asn470A, Asp471A | BOPAM-SP3: Phe1a, His13a, His14a, Gln15a, Lys16a, Leu17a, Val18a, Phe19a, Asp23a, Asn28a, Cys32a, Ala33a, Ile35a, Leu37a, Met38a |
| 35 | Streptococcus pneumoniae Pneumolysin: Val45A, Glu47A, Ser254A, Ser256A, Glu277A, Gln280A, Ile281A, Lys354A, Thr356A, Tyr358A, Arg359A, Arg419A, Pro446A, Leu447A, Val448A, Arg449A, Val468A, Asn470A, Asp471A | BOPAM-SP4: Arg2a, His3a, His13a, His14a, Gln15a, Lys16a, Leu17a, Val18a, Phe20a, Asp23a, Ser27a, Asn28a, Lys32a, Met38a, Ile44a |
| 36 | Streptococcus pneumoniae Pneumolysin: Lys19A, Leu20A, His23A, Glu26A, Val78A, Asp79A, Glu80A, Leu83A, Glu84A, Glu159A, Lys162A, Ser167A, Glu170A, Glu231A, Val351A | BOPAM-SP5: Phe1a, Arg2a, His3a, Glu4a, Phe20a, Val24a, Lys29a, Gly30a, Ile34a, Ile35a, Gly36a, Met38a |
| 37 | Streptococcus pneumoniae Pneumolysin: Glu42A, Thr253A, Ser254A, Lys255A, Ser256A, Glu277A, Ile281A, Asn284A, Ala357A, Tyr358A, Arg359A, Pro446A, Leu447A, Val448A, Asn470A | BOPAM-SP6: Glu11a, His14a, Gln15a, Lys16a, Leu17a, Val18a, Phe31a, Leu37a, Met38a |
| 38 | Streptococcus pneumoniae Pneumolysin: Thr55A, Thr57A, Asp59A, Met148A, Gln149A, Tyr150A, Glu151A, Lys164A, Phe165A, Asn194A, Lys196A, Glu264A, Lys268A, Val270A, Val272A, Ile314A, Glu315A, Phe344A | BOPAM-SP7: Arg3a, Cys8a, Asn25a, Phe27a, Asp34a, Ile38a, Lys40a, Asp41a, Lys42a, Asn43a, Gly44a |
| 39 | Klebsiella pneumoniae Iron-Regulated Outer Membrane Protein: Pro3A, His4A, Glu11A, Phe34A, Ser73A, Leu113A, Val115A, Phe142A, Arg225A, Asp227A, Glu228A, Tyr229A, Glu258A, Phe301A | BOPAM-KP1: Lys3a, Glu4a, Glu5a, Gly6a, Ser8a, Ser9a, His11a, Cys12a, Ser13a, Pro14a, Trp19a, Glu21a |
| 40 | Klebsiella pneumoniae Iron-Regulated Outer Membrane Protein: Ala94A, Arg108A, Thr109A, Ser111A, Arg112A, Tyr256A, Phe319A, Pro321A, Pro323A, Ser332A, Ser334A, Phe376A, Tyr501A, Tyr507A, Ser519A, Arg578A, Val620A, Thr621A, Gln696A | BOPAM-KP2: Lys3a, Tyr4a, Val5a, Lys7a, Gly9a, Leu10a, Asn11a, Gly13a, Gln15a, Lys17a, Ile18a, Asp19a, Asn20a |
| 41 | Klebsiella pneumoniae Iron-Regulated Outer Membrane Protein: Arg93A, Lys500A, Tyr501A, Leu515A, Leu517A, Thr561A, Ile562A, Val563A, Val564A, Asp576A, Val609A, Trp618A, Val620A, Thr621A, Ala654A, Ala655A, Arg697A, Pro704A | BOPAM-KP3: Met1a, Lys3a, His4a, Val6a, Lys7a, Leu8a, Val14a, Gln15a, Cys16a |
| 42 | Klebsiella pneumoniae Iron-Regulated Outer Membrane Protein: Gln81A, Asn85A, Thr109A, Tyr256A, Phe319A, Pro321A, Leu325A, Ser332A, Ser334A, Ser336A, Gln338A, Gln374A, Tyr507A, Ile518A, Ile693A, Gln696A, Arg697A, Ala698A, Leu700A, Leu711A | BOPAM-KP4: Ile1a, His2a, His3a, Glu4a, Ala5a, Lys7a, Gly8a, Tyr10a, Pro12a, Tyr13a, Leu14a, Trp16a, Leu18a |
| 43 | Klebsiella pneumoniae Iron-Regulated Outer Membrane Protein: Ala22A, Gln23A, Gly52A, Gln53A, Glu56A, Glu123A, Lys149A, Asp158A, Glu160A, Phe590A, Asp595A, Ala635A, Ala672A, Lys677A, Thr728A | BOPAM-KP5: Arg7a, Cys12a, Ser13a, Ala14a, Ser15a, Leu16a, Lys17a, Cys18a, Trp19a, Phe20a |
| 44 | Klebsiella pneumoniae Iron-Regulated Outer Membrane Protein: Ser79A, Gln81A, Asn85A, Met88A, Ala94A, Arg108A, Thr109A, Tyr256A, Phe376A, Tyr507A, Ile518A, Ser519A, Ile693A, Gln696A, Ala698A, Leu700A | BOPAM-KP6: Ala5a, Lys6a, Ala8a, Glu11a, Lys13a, Cys15a, Lys16a, Leu18a, Ala19a, Lys20a, Lys21a |
| 45 | Klebsiella pneumoniae Iron-Regulated Outer Membrane Protein: Ser79A, Gln81A, Asn85A, Gly87A, Met88A, Arg93A, Ala94A, Arg108A, Thr109A, Ser111A, Arg112A, Tyr256A, Ser234A, Lys500A, Tyr501A, Tyr502A, Tyr507A, Ile518A, Ile562A, Ile693A, Gln696A, Arg697A, Tyr712A | BOPAM-KP7: Leu1a, Arg3a, Glu4a, Val8a, Ser9a, His11a, Cys12a, Leu16a, Cys18a, Arg20a, Met22a |
| S/N | Parental AMP | Amino Acid and Position | Mutated AMP | Amino Acid and Position |
|---|---|---|---|---|
| 1 | BOPAM-AB1 | K-10 | BOPAM-AB1.1 | R-10 |
| 2 | BOPAM-AB2 | K-10 | BOPAM-AB2.1 | R-10 |
| 3 | BOPAM-AB3 | K-10 | BOPAM-AB3.1 | R-10 |
| 4 | BOPAM-AB4 | L-16 | BOPAM-AB4.1 | M-16 |
| 5 | BOPAM-AB5 | I-16 | BOPAM-AB5.1 | M-16 |
| 6 | BOPAM-KP1 | C-2 | BOPAM-KP1.1 | N-2 |
| 7 | BOPAM-KP2 | W-2 | BOPAM-KP2.1 | N-2 |
| 8 | BOPAM-KP3 | W-2 | BOPAM-KP3.1 | N-2 |
| 9 | BOPAM-KP4 | G-9 | BOPAM-KP4.1 | N-9 |
| 10 | BOPAM-KP5 | S-9 | BOPAM-KP5.1 | N-9 |
| 11 | BOPAM-KP6 | G-9 | BOPAM-KP6.1 | N-9 |
| 12 | BOPAM-KP7 | C-2 | BOPAM-KP7.1 | N-2 |
| 13 | BOPAM-SP1 | N-25 | BOPAM-SP1.1 | D-25 |
| 14 | BOPAM-SP2 | N-25 | BOPAM-SP2.1 | D-25 |
| 15 | BOPAM-SP3 | S-25 | BOPAM-SP3.1 | D-25 |
| 16 | BOPAM-SP4 | S-25 | BOPAM-SP4.1 | D-25 |
| 17 | BOPAM-SP5 | S-25 | BOPAM-SP5.1 | D-25 |
| 18 | BOPAM-SP6 | L-36 | BOPAM-SP6.1 | R-36 |
| 19 | BOPAM-SP7 | L-36 | BOPAM-SP7.1 | R-36 |
| 20 | BOPAM-INFA1 | L-13 | BOPAM-INFA1.1 | R-13 |
| 21 | BOPAM-INFA2 | L-13 | BOPAM-INFA2.1 | R-13 |
| 22 | BOPAM-INFA3 | G-3 | BOPAM-INFA3.1 | T-3 |
| 23 | BOPAM-INFA4 | L-13 | BOPAM-INFA4.1 | R-13 |
| 24 | BOPAM-INFA5 | G-3 | BOPAM-INFA5.1 | T-3 |
| 25 | BOPAM-INFA6 | L-13 | BOPAM-INFA6.1 | R-13 |
| 26 | BOPAM-INFA7 | L-13 | BOPAM-INFA7.1 | R-13 |
| 27 | BOPAM-INFA8 | G-3 | BOPAM-INFA8.1 | T-3 |
| 28 | BOPAM-INFB1 | H-5 | BOPAM-INFB1.1 | R-5 |
| 29 | BOPAM-INFB2 | H-5 | BOPAM-INFB2.1 | R-5 |
| 30 | BOPAM-INFB3 | C-12 | BOPAM-INFB3.1 | S-12 |
| 31 | BOPAM-INFB4 | L-15 | BOPAM-INFB4.1 | F-15 |
| 32 | BOPAM-INFB5 | L-15 | BOPAM-INFB5.1 | M-15 |
| 33 | BOPAM-INFB6 | L-8 | BOPAM-INFB6.1 | F-8 |
| 34 | BOPAM-RSV1 | S-3 | BOPAM-RSV1.1 | N-3 |
| 35 | BOPAM-RSV2 | S-3 | BOPAM-RSV2.1 | N-3 |
| 36 | BOPAM-RSV3 | V-3 | BOPAM-RSV3.1 | N-3 |
| 37 | BOPAM-RSV4 | T-3 | BOPAM-RSV4.1 | N-3 |
| 38 | BOPAM-RSV5 | T-13 | BOPAM-RSV5.1 | R-13 |
| 39 | BOPAM-RSV6 | S-3 | BOPAM-RSV6.1 | N-3 |
| 40 | BOPAM-RSV7 | S-3 | BOPAM-RSV7.1 | N-3 |
| 41 | BOPAM-RSV8 | Y-3 | BOPAM-RSV8.1 | N-3 |
| 42 | BOPAM-RSV9 | I-13 | BOPAM-RSV9.1 | R-13 |
| 43 | BOPAM-RSV10 | I-13 | BOPAM-RSV10.1 | R-13 |
| 44 | BOPAM-RSV11 | T-13 | BOPAM-RSV11.1 | R-13 |
| 45 | BOPAM-RSV12 | I-13 | BOPAM-RSV12.1 | R-13 |
| 46 | BOPAM-RSV13 | I-13 | BOPAM-RSV13.1 | R-13 |
| S/N | AMPs | Molecular Mass (Da) | % Hydrophobic | Common Amino Acid | Net Charge | PI | Boman Index (kcal/mol) | Half-Life in Mammals (Hours) |
|---|---|---|---|---|---|---|---|---|
| 1 | BOPAM-AB1.1 | 1783.33 | 52 | L | +2 | 11.65 | −1 | 1.1 |
| 2 | BOPAM-AB2.1 | 1817.249 | 52 | L | +2 | 11.65 | −0.88 | 1.1 |
| 3 | BOPAM-AB3.1 | 1937.58 | 64 | L | +2 | 11.65 | −1.63 | 1.1 |
| 4 | BOPAM-AB4.1 | 1867.50 | 58 | L | +2 | 10.81 | −1.66 | 1.1 |
| 5 | BOPAM-AB5.1 | 1714.1 | 47 | G | +4 | 11.92 | 0.72 | 4.4 |
| 6 | BOPAM-KP1.1 | 2496.90 | 30 | SC | 0 | 7.12 | 2.26 | 100 |
| 7 | BOPAM-KP2.1 | 2297.701 | 38 | N | +2 | 8.79 | 1.32 | 30 |
| 8 | BOPAM-KP3.1 | 2406.25 | 42 | N | +2 | 8.82 | 1.61 | 30 |
| 9 | BOPAM-KP4.1 | 2589.19 | 28 | K | +4 | 10.58 | 1.86 | 20 |
| 10 | BOPAM-KP5.1 | 2694.32 | 39 | K | +4 | 9.66 | 2.44 | 20 |
| 11 | BOPAM-KP6.1 | 2357.05 | 38 | K | +8 | 10.98 | 2.64 | 5.5 |
| 12 | BOPAM-KP7.1 | 2600.13 | 39 | S | +2 | 8.83 | 2.35 | 5.5 |
| 13 | BOPAM-SP1.1 | 5084.34 | 27 | R | +4 | 9.49 | 3.67 | 0.8 |
| 14 | BOPAM-SP2.1 | 4904.86 | 25 | G | +4 | 9,47 | 3.11 | 1.3 |
| 15 | BOPAM-SP3.1 | 4793.03 | 45 | G | −3 | 5.43 | 0.59 | 1.1 |
| 16 | BOPAM-SP4.1 | 4807.06 | 45 | G | −3 | 5.43 | 0.57 | 1.1 |
| 17 | BOPAM-SP5.1 | 4807.06 | 45 | G | −3 | 5.44 | 0.55 | 1.1 |
| 18 | BOPAM-SP6.1 | 4892.21 | 47 | V | −1 | 6.78 | 0.72 | 2.8 |
| 19 | BOPAM-SP7.1 | 5168.24 | 25 | K | +5 | 9.85 | 3.48 | 1.3 |
| 20 | BOPAM-NFA1.1 | 1569.43 | 28 | P | 0 | 6.34 | 1.57 | 7.2 |
| 21 | BOPAM-NFA2.1 | 1555.28 | 57 | VIL | +1 | 8.55 | 0.52 | 1.2 |
| 22 | BOPAM-NFA3.1 | 1562.33 | 28 | T | −2 | 3.49 | 1.63 | 7.2 |
| 23 | BOPAM-NFA4.1 | 1587.47 | 28 | P | 0 | 6.34 | 1.75 | 7.2 |
| 24 | BOPAM-NFA5.1 | 1585.69 | 28 | TOP | −1 | 3.75 | 1.15 | 7.2 |
| 25 | BOPAM-NFA6.1 | 1596.50 | 28 | P | +1 | 9.69 | 1.78 | 7.2 |
| 26 | BOPAM-NFA7.1 | 1605.29 | 50 | ISV | +1 | 8.55 | 1.03 | 1.2 |
| 27 | BOPAM-NFA8.1 | 1586.40 | 28 | PT | −2 | 3.55 | 1.24 | 7.2 |
| 28 | BOPAM-NFB1.1 | 1888.60 | 40 | R | +2 | 12.20 | 2.95 | 30 |
| 29 | BOPAM-NFB2.1 | 1863.61 | 46 | MRVP | +1 | 10.40 | 1.8 | 30 |
| 30 | BOPAM-NFB3.1 | 1671.56 | 40 | NL | 0 | 5.84 | 1.25 | 5.5 |
| 31 | BOPAM-NFB4.1 | 1798.04 | 66 | L | +2 | 10.81 | −1.44 | 5.5 |
| 32 | BOPAM-NFB5.1 | 1786.94 | 66 | L | +1 | 9.70 | −1.68 | 5.5 |
| 33 | BOPAM-NFB6.1 | 1690.34 | 46 | N | 0 | 5.76 | 0.7 | 5.5 |
| 34 | BOPAM-RSV1.1 | 1892.35 | 43 | IKN | +1 | 8.54 | 1.6 | 20 |
| 35 | BOPAM-RSV2.1 | 1803.10 | 31 | N | −1 | 4.43 | 2.49 | 1 |
| 36 | BOPAM-RSV3.1 | 1744.03 | 37 | N | 0 | 6.45 | 2.36 | 1.4 |
| 37 | BOPAM-RSV4.1 | 1797.11 | 43 | N | −1 | 4.18 | 1.95 | 1.3 |
| 38 | BOPAM-RSV5.1 | 1742.30 | 37 | N | +2 | 11.65 | 2.77 | 4.4 |
| 39 | BOPAM-RSV6.1 | 1840.79 | 50 | I | −1 | 4.18 | 1.09 | 1.3 |
| 40 | BOPAM-RSV7.1 | 1854.40 | 31 | N | 0 | 6.41 | 3.01 | 1.4 |
| 41 | BOPAM-RSV8.1 | 1734.98 | 43 | N | −1 | 3.85 | 1.41 | 1.9 |
| 42 | BOPAM-RSV9.1 | 1816.39 | 25 | N | +2 | 11.65 | 3.27 | 1.4 |
| 43 | BOPAM-RSV10.1 | 1815.40 | 31 | N | +1 | 9.69 | 2.99 | 100 |
| 44 | BOPAM-RSV11.1 | 1729.96 | 37 | N | −1 | 4.11 | 2.39 | 1.4 |
| 45 | BOPAM-RSV12.1 | 1847.16 | 37 | N | 0 | 6.45 | 2.48 | 1.1 |
| 46 | BOPAM-RSV13.1 | 1985.35 | 25 | DR | +1 | 9.53 | 5.36 | 1.3 |
| S/N | AMP Name | C-Score | TM-Score | RMSD |
|---|---|---|---|---|
| 1 | BOPAM-AB1.1 | −0.45 | 0.66 ± 0.13 | 1.5 ± 1.4 Å |
| 2 | BOPAM-AB2.1 | −0.44 | 0.66 ± 0.13 | 1.5 ± 1.4 Å |
| 3 | BOPAM-AB3.1 | −0.16 | 0.69 ± 0.12 | 1.0 ± 1.0 Å |
| 4 | BOPAM-AB4.1 | −0.33 | 0.67 ± 0.13 | 1.3 ± 1.3 Å |
| 5 | BOPAM-AB5.1 | 0.42 | 0.77 ± 0.10 | 0.5 ± 0.5 Å |
| 6 | BOPAM-KP1.1 | 0.71 | 0.81 ± 0.09 | 0.5 ± 0.5 Å |
| 7 | BOPAM-KP2.1 | −1.45 | 0.54 ± 0.15 | 3.7 ± 2.6 Å |
| 8 | BOPAM-KP3.1 | −1.42 | 0.54 ± 0.15 | 3.7 ± 2.5 Å |
| 9 | BOPAM-KP4.1 | −1.56 | 0.52 ± 0.15 | 3.9 ± 2.7 Å |
| 10 | BOPAM-KP5.1 | 0.03 | 0.72 ± 0.11 | 1.2 ± 1.2 Å |
| 11 | BOPAM-KP6.1 | −0.02 | 0.71 ± 0.12 | 1.2 ± 1.2 Å |
| 12 | BOPAM-KP7.1 | −0.57 | 0.64 ± 0.13 | 2.3 ± 1.8 Å |
| 13 | BOPAM-SP1.1 | 0.04 | 0.72 ± 0.11 | 2.3 ± 1.8 Å |
| 14 | BOPAM-SP2.1 | −0.12 | 0.70 ± 0.12 | 2.6 ± 1.9 Å |
| 15 | BOPAM-SP3.1 | −1.88 | 0.49 ± 0.15 | 6.1 ± 3.8 Å |
| 16 | BOPAM-SP4.1 | −1.88 | 0.49 ± 0.15 | 6.1 ± 3.8 Å |
| 17 | BOPAM-SP5.1 | −1.97 | 0.48 ± 0.15 | 6.3 ± 3.8 Å |
| 18 | BOPAM-SP6.1 | −1.47 | 0.53 ± 0.15 | 5.2 ± 3.4 Å |
| 19 | BOPAM-SP7.1 | −0.14 | 0.70 ± 0.12 | 2.7 ± 2.0 Å |
| 20 | BOPAM-INFA1.1 | −1.03 | 0.58 ± 0.14 | 2.2 ± 1.7 Å |
| 21 | BOPAM-INFA2.1 | −0.14 | 0.70 ± 0.12 | 0.7 ± 0.7 Å |
| 22 | BOPAM-INFA3.1 | −1.50 | 0.53 ± 0.15 | 3.1 ± 2.2 Å |
| 23 | BOPAM-INFA4.1 | −1.05 | 0.58 ± 0.14 | 2.2 ± 1.7 Å |
| 24 | BOPAM-INFA5.1 | −0.97 | 0.59 ± 0.14 | 2.1 ± 1.7 Å |
| 25 | BOPAM-INFA6.1 | −1.00 | 0.59 ± 0.14 | 2.1 ± 1.7 Å |
| 26 | BOPAM-INFA7.1 | −0.71 | 0.62 ± 0.14 | 1.6 ± 1.4 Å |
| 27 | BOPAM-INFA8.1 | −1.13 | 0.57 ± 0.14 | 2.4 ± 1.8 Å |
| 28 | BOPAM-INFB1.1 | −0.77 | 0.62 ± 0.14 | 1.9 ± 1.5 Å |
| 29 | BOPAM-INFB2.1 | −0.78 | 0.61 ± 0.14 | 1.9 ± 1.6 Å |
| 30 | BOPAM-INFB3.1 | −1.19 | 0.57 ± 0.15 | 2.6 ± 1.9 Å |
| 31 | BOPAM-INFB4.1 | −0.03 | 0.71 ± 0.12 | 0.6 ± 0.6 Å |
| 32 | BOPAM-INFB5.1 | −0.35 | 0.67 ± 0.13 | 1.1 ± 1.1 Å |
| 33 | BOPAM-INFB6.1 | −0.91 | 0.60 ± 0.14 | 2.1 ± 1.7 Å |
| 34 | BOPAM-RSV1.1 | −0.00 | 0.71 ± 0.11 | 0.7 ± 0.7 Å |
| 35 | BOPAM-RSV2.1 | −0.01 | 0.71 ± 0.11 | 0.7 ± 0.7 Å |
| 36 | BOPAM-RSV3.1 | −0.23 | 0.68 ± 0.12 | 1.1 ± 1.1 Å |
| 37 | BOPAM-RSV4.1 | −1.53 | 0.53 ± 0.15 | 3.4 ± 2.3 Å |
| 38 | BOPAM-RSV5.1 | −0.37 | 0.67 ± 0.13 | 1.3 ± 1.3 Å |
| 39 | BOPAM-RSV6.1 | −0.85 | 0.61 ± 0.14 | 2.1 ± 1.7 Å |
| 40 | BOPAM-RSV7.1 | −0.13 | 0.70 ± 0.12 | 0.9 ± 0.9 Å |
| 41 | BOPAM-RSV8.1 | −1.42 | 0.54 ± 0.15 | 3.2 ± 2.2 Å |
| 42 | BOPAM-RSV9.1 | −0.11 | 0.70 ± 0.12 | 0.9 ± 0.9 Å |
| 43 | BOPAM-RSV10.1 | −0.78 | 0.61 ± 0.14 | 2.0 ± 1.6 Å |
| 44 | BOPAM-RSV11.1 | −1.26 | 0.56 ± 0.15 | 2.9 ± 2.1 Å |
| 45 | BOPAM-RSV12.1 | −1.46 | 0.53 ± 0.15 | 3.2 ± 2.3 Å |
| 46 | BOPAM-RSV13.1 | −0.10 | 0.70 ± 0.12 | 0.8 ± 0.8 Å |
| S/N | Receptors | Parental AMPs | Binding Score | Mutated AMPs | Binding Score |
|---|---|---|---|---|---|
| 1 | Iron-Regulated OMP | BOPAMAB1 | 10,566 | BOPAMAB1.1 | 11,826 |
| 2 | Iron-Regulated OMP | BOPAMAB5 | 11,388 | BOPAMAB5.1 | 12,952 |
| 3 | Nucleoprotein | BOPAMINFA1 | 12,134 | BOPAMINFA1.1 | 12,346 |
| 4 | Nucleoprotein | BOPAMINFA2 | 10,870 | BOPAMINFA2.1 | 10,888 |
| 5 | Nucleoprotein | BOPAMINFA3 | 10,572 | BOPAMINFA3.1 | 12,578 |
| 6 | Nucleoprotein | BOPAMINFA4 | 12,300 | BOPAMINFA4.1 | 12,458 |
| 7 | Nucleoprotein | BOPAMINFA6 | 12,110 | BOPAMINFA6.1 | 13,256 |
| 8 | Nucleoprotein | BOPAMINFA7 | 10,982 | BOPAMINFA7.1 | 11,120 |
| 9 | Nucleoprotein | BOPAMINFA8 | 12,604 | BOPAMINFA8.1 | 14,170 |
| 10 | Nucleoprotein | BOPAMINFB5 | 11,704 | BOPAMINFB5.1 | 11,932 |
| 11 | Iron-Regulated Outer Membrane Protein | BOPAMKP1 | 11,305 | BOPAMKP1.1 | 12,268 |
| 12 | Iron-Regulated Outer Membrane Protein | BOPAMKP3 | 12,384 | BOPAMKP3.1 | 13,216 |
| 13 | Iron-Regulated Outer Membrane Protein | BOPAMKP5 | 10,810 | BOPAMKP5.1 | 10,984 |
| 14 | Iron-Regulated Outer Membrane Protein | BOPAMKP6 | 13,208 | BOPAMKP6.1 | 13,870 |
| 15 | Chain A Protein | BOPAMRSV3 | 9068 | BOPAMRSV3.1 | 9156 |
| 16 | Chain A Protein | BOPAMRSV6 | 8194 | BOPAMRSV6.1 | 9236 |
| 17 | Chain A Protein | BOPAMRSV9 | 8500 | BOPAMRSV9.1 | 9278 |
| 18 | Chain A Protein | BOPAMRSV10 | 9020 | BOPAMRSV10.1 | 9158 |
| 19 | Chain A Protein | BOPAMRSV13 | 8866 | BOPAMRSV13.1 | 9072 |
| 20 | Pneumolysin | BOPAMSP1 | 12,306 | BOPAMSP1.1 | 13,164 |
| 21 | Pneumolysin | BOPAMSP3 | 12,116 | BOPAMSP3.1 | 14,134 |
| 22 | Pneumolysin | BOPAMSP4 | 12,384 | BOPAMSP4.1 | 12,934 |
| 23 | Pneumolysin | BOPAMSP7 | 11,830 | BOPAMSP7.1 | 12,378 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakare, O.O.; Gokul, A.; Keyster, M. Analytical Studies of Antimicrobial Peptides as Diagnostic Biomarkers for the Detection of Bacterial and Viral Pneumonia. Bioengineering 2022, 9, 305. https://doi.org/10.3390/bioengineering9070305
Bakare OO, Gokul A, Keyster M. Analytical Studies of Antimicrobial Peptides as Diagnostic Biomarkers for the Detection of Bacterial and Viral Pneumonia. Bioengineering. 2022; 9(7):305. https://doi.org/10.3390/bioengineering9070305
Chicago/Turabian StyleBakare, Olalekan Olanrewaju, Arun Gokul, and Marshall Keyster. 2022. "Analytical Studies of Antimicrobial Peptides as Diagnostic Biomarkers for the Detection of Bacterial and Viral Pneumonia" Bioengineering 9, no. 7: 305. https://doi.org/10.3390/bioengineering9070305