1. Introduction
Traditionally, suckler bull beef production was based on the provision of concentrate-based diets until slaughter (i.e., intensive indoor systems) [
1,
2]. The profitability of such systems in temperate regions, can be enhanced by the use of cheaper feedstuffs such as conserved or grazed pasture [
3]. Thus, O’Riordan et al. [
2] showed that introducing a pasture grazing period of 100 days for suckler bulls prior to finishing on concentrates decreased the cost of production. It was also reported that in such suckler beef systems, slaughtering bulls at under 20 months of age was desirable, as some markets have an age restriction [
1]. However, suckler bulls from such systems, which usually involve late maturing breed types, risked not achieving the desired carcass fat classification [
2,
4]. Similarly, such modifications (i.e., incorporating a grazing period) can affect the compositional and eating quality of beef [
4].
The longer bulls are at pasture, the cheaper the cost of production. However, the trade-off between the duration of grazing, carcass fat classification and the extent of the concentrate feeding during finishing is important economically. There is a need therefore to investigate the effect of extending the grazing period combined with finishing suckler bulls on a cereal concentrate on carcass fat classification, as well as other important carcass and meat quality traits. In addition to maximising economic returns, beef from grass fed animals is favoured by some consumers, as it is considered more ‘natural’ [
5] and reported to enhance the proportion of nutritionally important fatty acids [
6,
7,
8]. However, the provision of grass for a prolonged period in bull beef production systems would likely increase the age at slaughter, which may influence consumer preferences and the sensory qualities of beef [
9,
10].
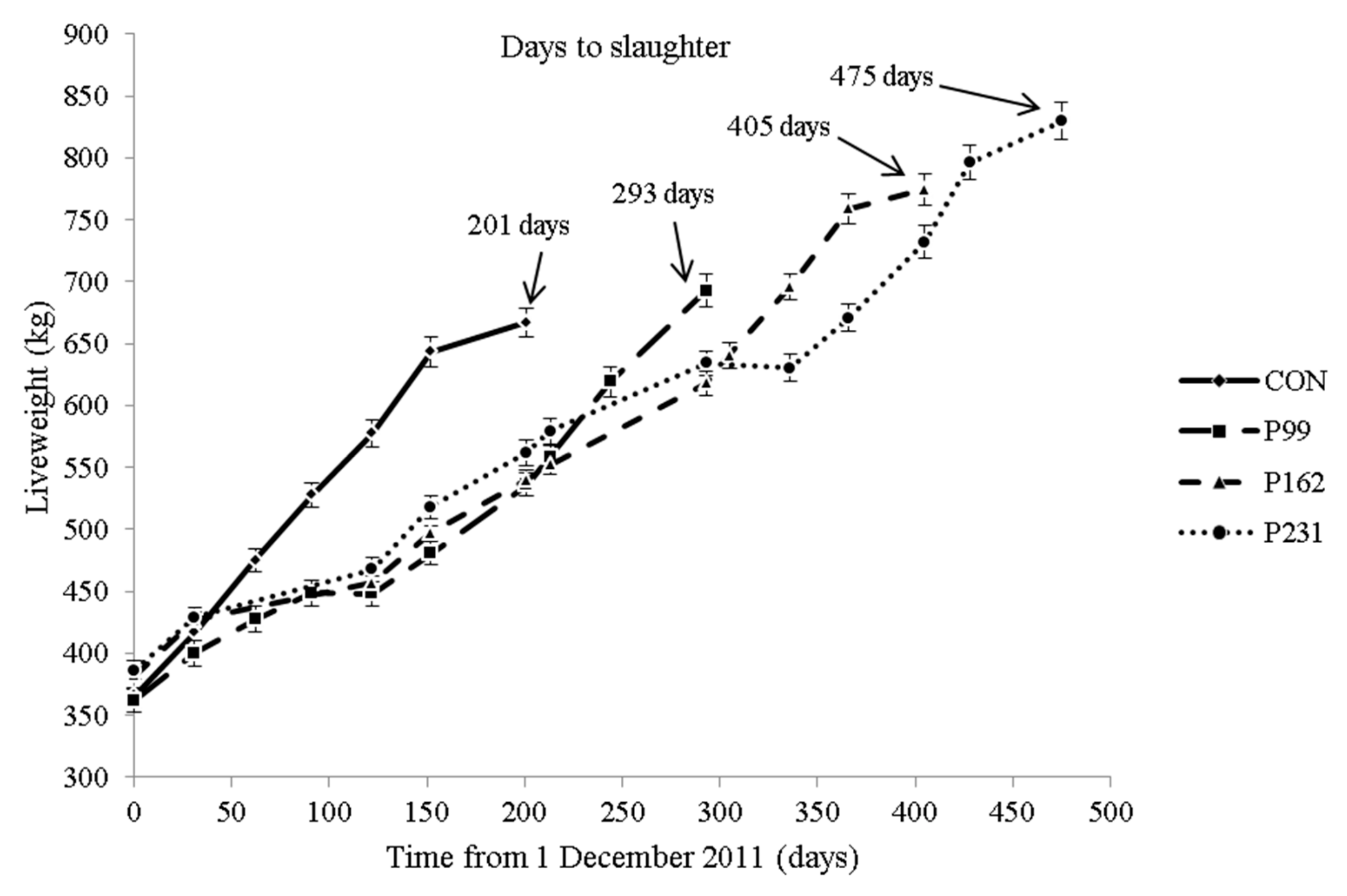
Therefore, the objective of the study was to examine the effect of varying the grazing period from 0 to approximately 8 months prior to finishing on a cereal-based concentrate (to achieve a carcass fat classification similar to that achieved by the intensively concentrate fed bulls) on fat and muscle colour, biochemical and sensory characteristics of beef. It was hypothesized that extending the grazing period up to a full season and its associated increase in slaughter age would negatively influence the quality of beef, particularly sensory tenderness.
4. Discussion
In temperate climates, maximising the proportion of grazed grass in the diet of beef cattle generally decreases the cost of production. The primary objective of the present study was to exploit a full grazing season in a suckler bull production system. From a carcass quality perspective, the main concern in this study was meeting the carcass weight and fat classification required in several European markets. From a meat quality perspective, the main concern was whether the increasing age/weight at slaughter as the period at pasture increased would have a negative effect. The slaughter strategies chosen were based on the modification of the more traditional sucker bull beef production (high concentrate feeding indoors to less than 16 months of age) whereby a relatively short grazing period is included and animals are slaughtered to a carcass weight target (C v P99) [
2] and then extending the grazing period but slaughtering at a similar fat classification as the traditional system (C, P162, P231). These strategies were achieved with respect to carcass weight, similar for C and P99 and fat classification, similar for C, P162 and P231. The low average carcass fat classification for the P99 bulls reflects the shorter finishing period on concentrate to achieve the target carcass weight. Some lucrative bull beef markets require a carcass fat classification of ≥6 (on a 1–15 scale). Of the 14 carcasses in P99, 7 were in fat class 6 i.e., borderline acceptable (6 were in fat class 3 and 1 in fat class 9), which highlights the dependence of good grass growth and management in maximizing the role of grazed grass in producing “lighter” carcasses within this system. However, from a lean meat production perspective, the lower average fat score, at a similar carcass weight, was reflected in lower fat deposition in the cube roll joint highlighting the positive effect of grazing in this regard. Achieving the current market specification for fat score at 18 months of age is most easily done by using an early maturing sire breed [
21].
As designed, the increase in the duration of grazing followed by concentrate feeding to meet the fat classification target increased the age at slaughter resulting in a range in mean age of 9 months approximately. The higher growth rate prior to slaughter (finishing period) for the P99 bulls can be explained by the shorter indoor finishing period as an increase in the length of the finishing period results in a progressive decrease in live weight gain [
22]. With regard to subcutaneous fat colour, the higher lightness for C, P162 and P231 bulls may be attributed to the higher fat scores of their carcasses compared to the P99 bulls. A similar observation on the relationship between fat classification and fat lightness was made by Mezgebo et al. [
4] Based on the review of factors influencing fat colour [
23] we hypothesised that fat yellowness would increase as the duration of the grazing period increased. The data do not support this hypothesis. If the fat had become more yellow due to grazing and deposition of carotenoids from the grass [
23], it seems the length of the finishing period on concentrates was sufficient to remove this effect.
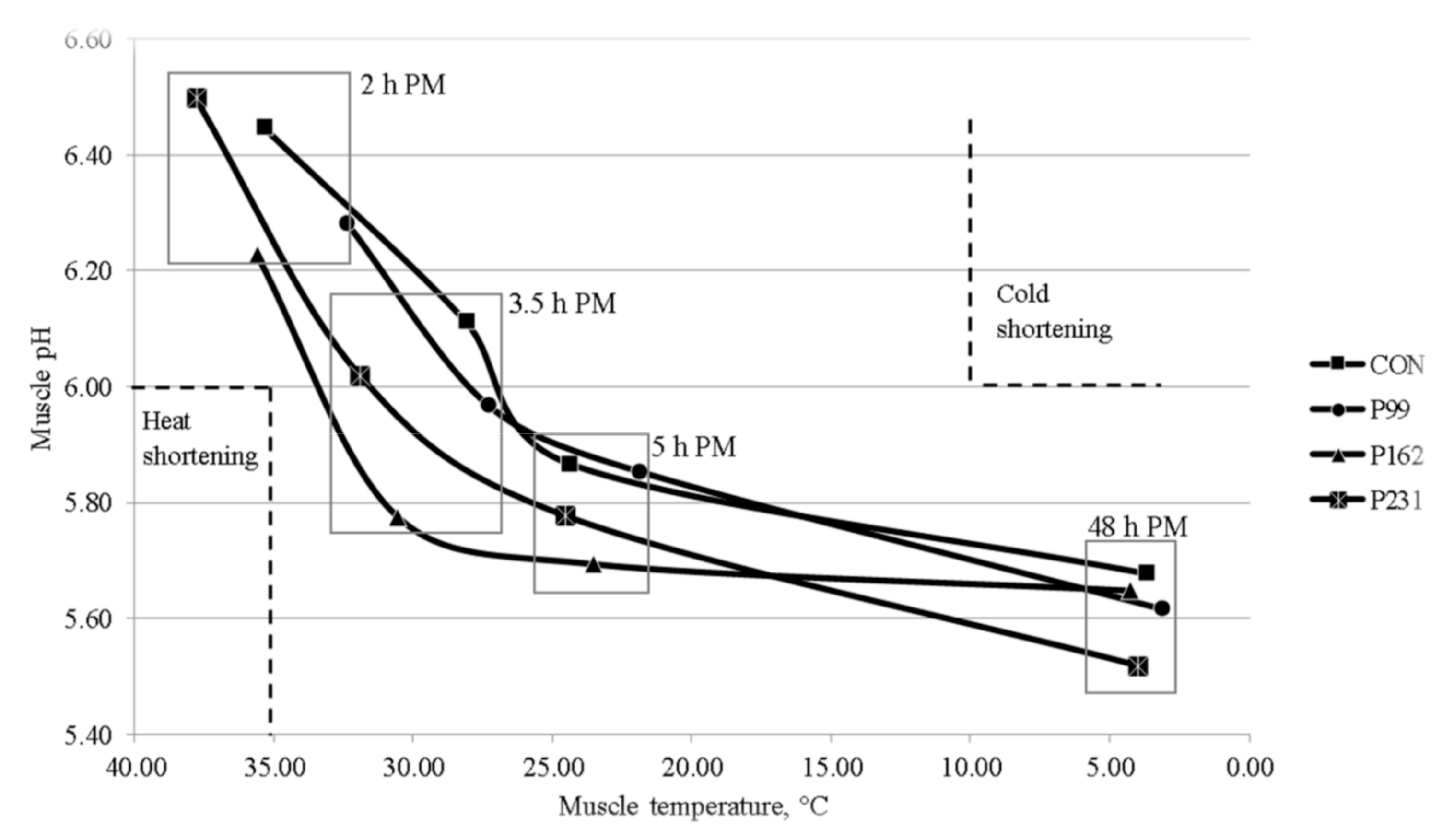
The lower fat classification of the P99 bulls may have contributed to the faster post-mortem decline in LT temperature of these bulls compared to the C, P162 and P231 bulls. The post-mortem pH profile is mainly determined by muscle glycogen content at slaughter, which in turn is influenced by pre-slaughter nutrition and stress levels of the animal before and at slaughter [
24]. In the present study, the animals were managed to avoid stress-related loss of glycogen mainly by finishing indoors and careful handling during transport and in the lairage. While variations in the pattern of decline in the pH of LT were observed between the groups, none of the carcasses were likely to have been affected by ‘cold shortening’, muscle pH > 6 at muscle temperature < 10 °C [
25] or ‘heat shortening’, muscle pH < 6 at muscle temperature >35 °C [
26,
27] (
Figure 2).
The lower pH at 48 h post-mortem for the P231 bulls than for the C, P99 and P231 bulls suggests that the glycogen level was higher before slaughter in LT from these bulls. However, LT pH values from all groups were within the ‘normal’ pH range of 5.4–5.8 [
28] and no carcasses were deemed “dark cutters” by abattoir personnel. Nevertheless, when adjusted for production system effects i.e., partial correlation, pH was negatively correlated with LT lightness (r = −0.45,
p < 0.001) and ‘hue’ angle (r = −0.41,
p < 0.01).
The higher redness and colour saturation and lower ‘hue’ angle of LT from P162 and P231 bulls compared to C and P99 bulls, and the higher lightness for the C bulls compared to P99, P162 and P231 bulls, can be explained by the older age at slaughter as muscle tissue becomes redder and darker with increasing slaughter age [
29,
30]. The lack of effect of slaughter age per se on colour variables when adjusted for production system effects in the present study, however, highlights the confounding effect of other production variations on LT colour. It is likely that increasing the grazing period increased the total amount of exercise engaged in by the animals when at pasture. However, data with respect to exercise and muscle colour in cattle are equivocal [
31]. The higher proportion of LT type I fibres (characteristic of red muscle) for the P162 and P231 bulls compared to C and P99 bulls could also contribute to the higher redness.
Deposition of fat generally increases with increasing age and carcass weight of beef cattle [
32,
33]. However, in the present study, the higher IMF concentration (and the proportion of fat in the cube roll joint) for C compared to P99 bulls despite being younger at slaughter reflects their higher energy consumption and higher growth rate over the full experimental period as high nutrient/energy intake results in higher fat deposition [
32,
33,
34]. Tat the IMF concentration (and the proportion of fat in the cube roll joint) was similar for P162 and P231, despite the latter being older and having a heavier carcass at slaughter might reflect a residual effect of the longer grazing period since grazing generally results in lower fat deposition [
4,
35]. According to [
33], animal age is one of the main factors that determine the physical and chemical properties of muscle connective tissue as the animal ages, the proportion of heat stable (chemically non-reducible) crosslinks, such as pyridinoline and Ehlichs chromagen, increases [
36,
37], which in turn leads to a decrease in the proportion of heat soluble collagen [
37,
38]. On a production system basis, the data from the present study support these reports with regard to collagen solubility. However, the lack of a relationship between slaughter age, slaughter weight or carcass weight and collagen solubility, when adjusted for production system effects, indicates that the maturation of collagen is not due to one dominant influence. Moreover, while the lower collagen solubility in LT from P162 and P231 bulls compared to C and P99 bulls could be hypothesised to be due to the higher pyridinoline cross link concentration (P162 numerically but not significantly, higher than P99), when adjusted for production system effects this relationship was not significant.
As a general trend, as an animal becomes older (especially beyond sexual maturity) its muscle undergoes less glycolytic and more oxidative metabolism [
39,
40]. The lower activities of the glycolytic enzymes, LDH and PFK (tendency), in LT from the P231 bulls in the present study could therefore be attributed to their older age at slaughter compared to the C, P99 and P162 bulls. However, these three groups had similar LDH and PFK activities in LT despite their differences in age indicating that age is not the only factor influencing the activity of these enzymes. The higher proportion of slow twitch type I oxidative fibres in LT from the P162 and P231 bulls indicates that an increase in age at slaughter can lead to an increase in oxidative fibres as reported by Jurie et al. [
39] which in turn can lead to an increase in oxidative metabolism of the muscle. However, there was no statistically significant difference in the activity of the oxidative enzymes ICDH, COX and CS. In addition, the higher proportion of type I oxidative fibres in LT from P162 and P231 bulls compared to P99 bulls could also reflect more physical activity due to the extended grazing period [
41,
42]. The higher proportion of type IIX (fast twitch glycolytic) fibres in LT from C bulls compared to P99 bulls and generally higher for P162 and P231 bulls than for the P99 bulls can be explained mainly by the longer duration of concentrate finishing as high energy intake favours glycolytic muscle metabolism [
41,
43]. In the present study, the type IIB (fast twitch glycolytic, data not shown) muscle fibre was identified in only 7 animals (1 in C, 4 in P99, none in P162, 2 in P231 bulls; 1 Charolais and 6 Limousin sired bulls)). This might indicate that expression of Type IIB fibre is breed specific as it was reported to be identified commonly in Blonde d’Aquitaine, a French beef breed [
44].
The perception that beef, and bull beef in particular, becomes less tender and less acceptable to the consumer as an animal becomes older is an important contributor to the inclusion of age limits in market specifications for bull beef. For steers and heifers, there appears to be little effect of age on tenderness, at least up to 24 months [
45]. The lack of a significant relationship between age and tenderness, when adjusted for production system effects, in the present study indicates that a similar conclusion can be made for bulls. In support of this, Dikeman et al. [
46] observed no difference in shear force or sensory tenderness in LT from early maturing bulls as age at slaughter increased from 12 to 24 months. Similarly, a literature review of mainly French production systems indicated that as age at slaughter increased from 12 to 24 months there was little evidence of an increase in shear force or decrease in sensory tenderness in meat from bulls [
47]. In Irish studies with bulls of dairy origin, there was no difference in shear force or sensory tenderness of LT from production systems similar to C, P99 and P162 where the dairy bulls were 15, 19 and 22 months of age at slaughter, respectively [
48]. It is difficult, however, in many studies to separate the effects of age per se from other production factors, which might contribute to differences in beef tenderness. In a study with similar bulls as those used in this study, when averaged across three different production systems, an increase in age from 14.5 to 21.2 months did not affect LT tenderness, however, there was an effect of production system per se [
4]. With regard to the eating quality of beef, the differences in sensory characteristics between the treatments in the present study likely reflect the combined and possibly interactive effect of the variations in pre-slaughter growth rate, IMF concentration, muscle fibre distribution and collagen crosslinks. The lower tenderness, flavour liking and overall liking scores for the P231 bulls compared to the C bulls could mainly be attributed to their higher collagen cross-link content and altered fibre distribution since the IMF concentration was similar. Similarly, the lower tenderness and flavour scores for the P99 bulls compared to the C bulls could be related to the lower IMF concentration since muscle structure was not different. The favourable juiciness scores for both P162 and P231 bulls and the higher juiciness scores for the P231 bulls compared to the P99 bulls can possibly be related to their older age at slaughter as beef from older and fatter animals is perceived to be juicier than beef from younger and leaner animals [
7,
8].




{kind=link}
{kind=link}