1. Introduction
The latest estimation by the Food and Agriculture Organization of the United Nations (FAO) shows continuously growth of the protein demand around the world due to population growth [
1]. Lean meat is considered protein-rich, with an average protein content of about 22%, and has a balanced essential amino acid composition, playing an essential role in the human diet [
2]. However, the increasing global production and consumption of meat has triggered global environmental concerns in terms of greenhouse gas emissions, land use, pollution, and biodiversity loss [
3,
4,
5,
6]. To address this issue, calls for a reduction in meat consumption and an increase in the production of alternative proteins have been proposed [
7].
In this respect, insects have gained interest as sustainable protein sources, mainly due to their environmental and nutritional benefits. Insect farming, compared to farming cattle or pigs, generally requires less feed, water, and land, and emits fewer greenhouse gases and less ammonia [
8]. Insect rearing for food has a high feed-to-protein conversion efficiency: for example, house crickets (
Acheta domesticus) are at least 4 times more efficient than pigs and 12 times more efficient than cattle [
8,
9]. From a nutritional perspective,
A. domesticus has a crude protein content of ~13–25% as a wet basis and ~42–75% as dry matter [
10]. All essential amino acids are present in
A. domesticus, and the contents of essential amino acids are comparable to those of pork and beef [
11,
12]. Additionally,
A. domesticus has a protein digestibility-corrected amino acid score of 0.69–0.92, which is comparable to the 0.92 score of meat and higher than those of plant proteins such as lentils, beans, peas, and chickpeas, which have scores of 0.57–0.71 [
13]. The digestible indispensable amino acid score of
A. domesticus was found to be >75, with sulfur amino acids (cysteine and methionine) identified as the first limiting amino acids for children, adolescents, and adults [
14].
There are 2111 edible insect species consumed by 2 billion people in 113 countries, most species being collected wild, and a few species being farmed [
15,
16]. Entomophagy (the practice of eating insects) is already a part of the diets in some regions, and insects are even considered delicacies in many cultures located in Africa, Asia, and South America [
8]. In Europe, however, consumer acceptance of eating insects is still a challenge, due to reasons such as cultural taboos and safety concerns (possible toxicity, allergenicity, or microbial loads) [
17,
18]. Research suggests that adding insects, in such a way that they become invisible in food preparations, might help to reduce insect food neophobia [
19]. For example, insects could be incorporated into hybrid meat products, which may be used to persuade predominant meat eaters to reduce their meat consumption.
Past studies incorporating insect-based ingredients into processed meat products have focused on the compositional and technological aspects of hybrid products. Insect flour from mealworms (
Tenebrio molitor), silkworms (
Bombyx mori pupae), and house crickets (
A. domesticus) have been reported to be suitable to replace up to 20% of the pork in cooked sausages or patties without negative effects on nutritional or technological properties [
20,
21,
22,
23]. Furthermore, Kim et al. (2020) [
24] suggested the use of protein extract from yellow mealworms (
T. molitor) to replace about 40% of the pork myofibrillar protein in a hybrid emulsion system. Meat-insect hybrid products containing pork and superworms (
Zophobas morio larvae) have been reported to have similar viscoelastic properties, water-holding capacities, and gel-forming abilities as compared to meat products, provided that a higher heating temperature and the proper insect:meat ratio were used [
25,
26]. However, the desired texture tends to pose a challenge when developing meat-insect hybrid products. Kim et al. (2020) and Scholliers, Steen, and Fraeye (2020) pointed out that the interaction between insect protein and myofibrillar protein resulted in degraded rheological properties and thermal stability of the hybrid emulsion system in comparison to pure meat systems [
24,
27]. F. Zhang et al. (2022) found a lowered sensory score for cooked sausages when replacing lean meat with defatted mealworm (
T. molitor) flour even at a low ratio, indicating that the processing condition (heating temperature, ionic strength) is crucial for the desired texture [
28]. The use of functional ingredients (such as hydrocolloids, starch, and plant-based proteins) was suggested to improve the texture of the final products when developing meat-insect hybrid products [
26]. In addition to meat-insect hybrid products, combinations of plant-based materials and insects to produce hybrid products (such as extrudates, jerky, breads, pasta, 3D-printed snacks, and chocolate chip cookies) have also been suggested, aiming to improve the textural and nutritional quality of the final products under controlled processing conditions [
29,
30,
31,
32,
33,
34,
35,
36,
37,
38,
39].
Oxidation reactions are crucial to the quality of the final products because they can result in various compounds altering the sensory, structural, and nutritional properties of foods. Currently, there are limited research papers studying the oxidative stability of hybrid products made from combined meat and insect products during storage. Kim et al. (2016) reported a slight increase in lipid oxidation during 7 days of storage of pork-insect sausages which were made by adding defatted mealworm (
T. molitor) or silkworm (
B. mori pupae) flour to a pork sausage recipe [
22]. However, there is no existing literature regarding protein oxidation of a hybrid product combining meat and insects. Therefore, the aim of the current study was to investigate the effects of added cricket flour on the textural properties and oxidative stability of hybrid sausages in order to clarify the technological properties of insect meal as an ingredient in meat products. The specific aims were to (1) examine the functional properties of cricket flour (CF), (2) evaluate the effects of CF addition on the textural properties of cooked sausages, and (3) evaluate the effects of CF addition on lipid and protein oxidation of the cooked sausages during storage.
2. Materials and Methods
2.1. Materials
House crickets (A. domesticus) were purchased as whole and frozen at the adult stage from Villa Mangs Ltd. (Raasepori, Finland). After lyophilization (–80 °C, 0 mbar, ≥4 days) (Gamma 2–20 Freeze Dryer, Martin Christ, Osterode, Lower Saxony, Germany) and grinding (GM200, Retsch, Haan, North Rhine-Westphalia, Germany), CF was stored in desiccator at –20 °C in the dark until use. Freeze-drying was chosen to improve the mixing and comminution of the crickets, in order to avoid visible brown particles on the cut surface and overall browning of the insect ingredients, which would be caused by heating. Lean pork and pork fat were ordered from Lihatukku Heikin Liha Ltd. (Helsinki, Finland); after pre-chopping, lean pork and pork fat were vacuum-packed and kept at –20 °C in the dark until use. All chemicals and reagents were of analytical grade and obtained from either Sigma-Aldrich (Seelze, Germany) or Merck (Darmstadt, Germany).
2.2. Functional Properties of CF
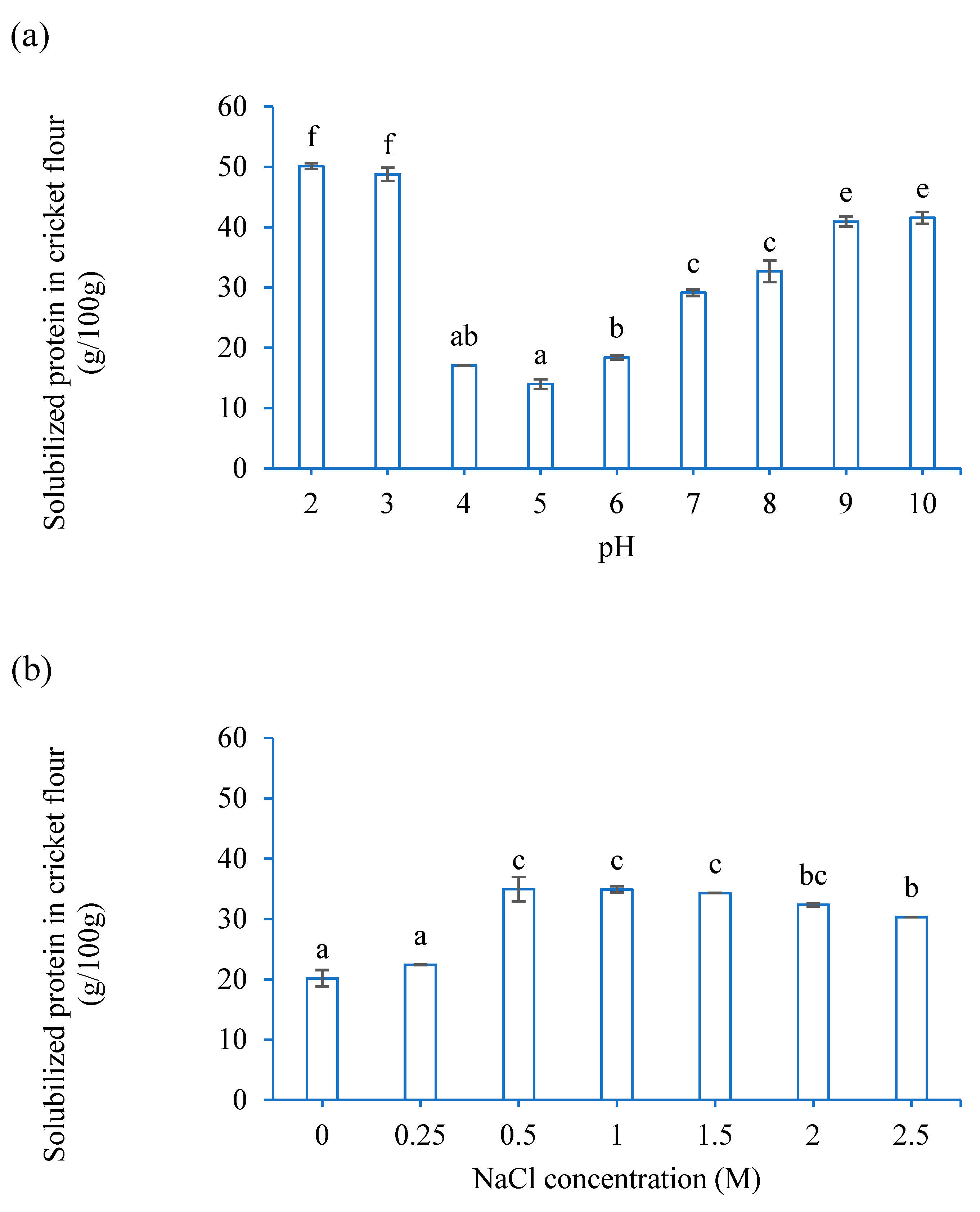
The protein solubility of CF at different pH levels (2–10) and NaCl concentrations (0–2.5 M) was determined using the method outlined by Kim et al. (2017) [
23]. To investigate how pH affects protein solubility, 1 g of CF was weighed in a centrifuge tube and mixed (vortex 30 s) with 10 mL Milli-Q water. The pH of the mixture was adjusted with a few drops of 6 M HCl or 6 M NaOH solution. To study the effect of NaCl concentration on protein solubility, 1 g of CF was mixed with 10 mL NaCl solutions (0, 0.25, 0.5, 1, 1.5, 2, 2.5 M) at pH 6.8. The mixture was mixed well (vortexed for 2 min) and shaken overnight (4 °C). After centrifuging (3220×
g, 4 °C, 15 min), the protein content in the supernatant was determined according to DC Protein Assay (Bio-Rad, Hercules, CA, USA) against BSA at a wavelength of 750 nm using a spectrophotometer (UV-1800, Shimadzu, Tokyo, Japan). The protein solubility of CF was expressed as g protein in the supernatant per g sample.
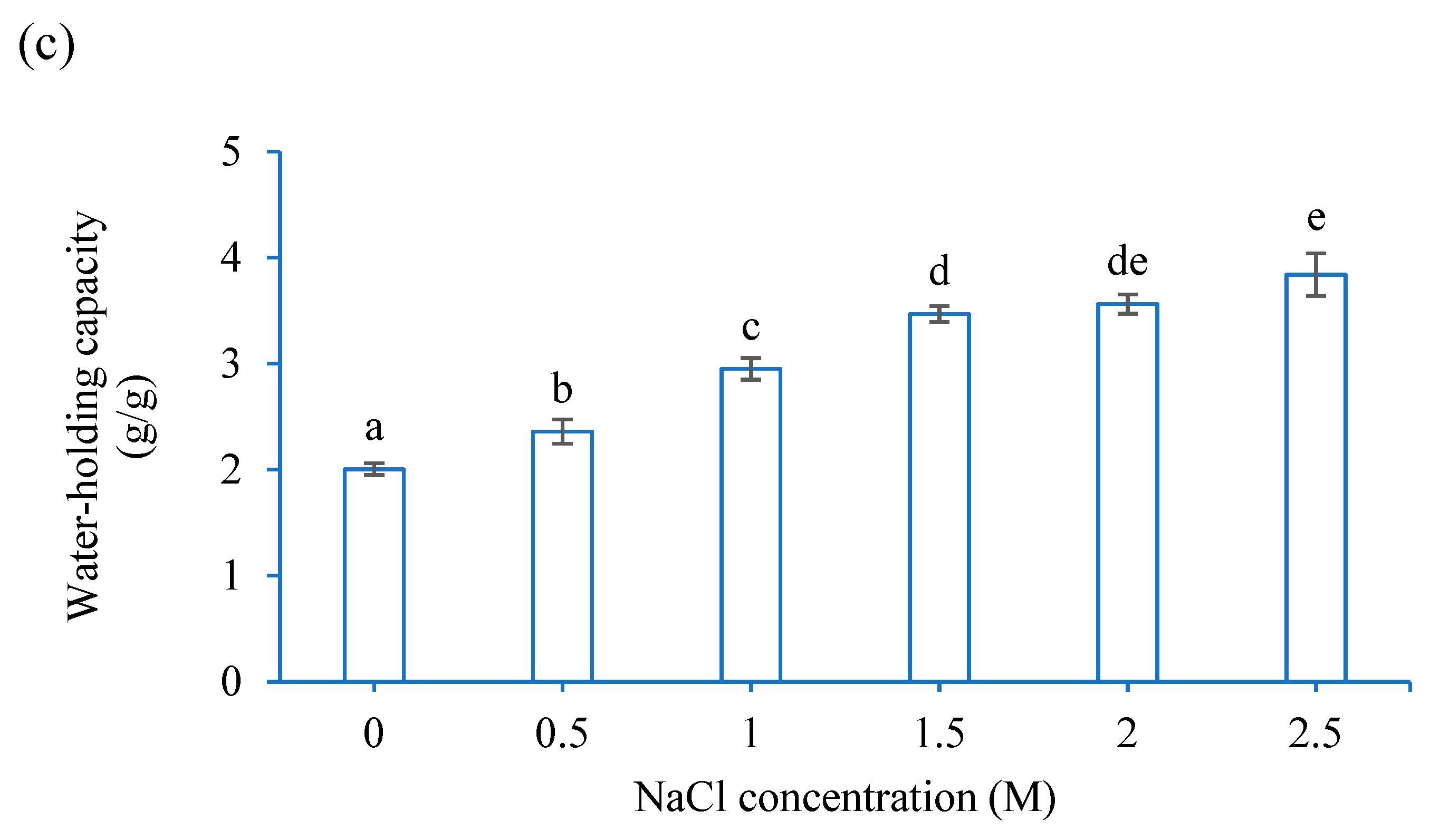
Water-holding capacity of CF was determined by a laboratory method. First, 1 g of CF was mixed with Milli-Q water or NaCl solutions (0.5, 1.0, 1.5, 2.0, or 2.5 M) at pH 6.8 (vortex, 2 min, room temperature (r.t.)). The mixture was heated at 70 °C (water bath, 30 min). After cooling and centrifuging (5000× g, 21 °C, 30 min), the supernatant was discarded. The water-holding capacity was calculated as the weight difference between the pellet and the 1 g sample.
The gel-forming ability of CF was evaluated at 1%, 2.5%, 5%, 10%, 20%, and 30% (g/100 mL) in Milli-Q water, 1.0 M NaCl, or 2.1 M NaCl using the method by Yi et al. (2013) [
40]. The mixture in the test tube was vortexed (2 min) and adjusted to pH 6.8 by slowly adding 6 M HCl or 6 M NaOH solutions. After dissolving at 4 °C overnight, gel formation was determined through visual observation after heating at 70 °C (water bath, 30 min). If the liquid mixture was not moving upon turning the test tube, it was considered as a gel.
2.3. Proximate Composition
Lean pork and CF were characterized by determining their moisture, protein, crude fat, and chitin contents, in triplicate. The moisture content was measured by an oven air-drying method [
41]. The protein content was analyzed by the Kjeldahl method with a nitrogen-to-protein conversion factor of 5.09 for CF [
42] and 6.25 for lean pork, respectively. The crude fat content was analyzed based on Soxhlet extraction using the Soxtec system (Soxtec 2050, Foss Tecator, Hillerød, Denmark). The fatty acid composition of CF was determined by the laboratory method using GC-FID (Agilent 6890 N, Agilent, Santa Clara, CA, USA). The chitin content of CF was measured according to a reversed-phase ultra-high performance liquid chromatography method outlined by Han and Heinonen (2021) using UHPLC (Acquity Ultra Performance LC, Waters, Milford, MA, USA) equipped with a fluorescence detector [
43]. The proximate compositions of the sausages were calculated from the analytical values of lean pork and CF.
2.4. Sausage Preparation
A preliminary laboratory sausage test was carried out to find the suitable levels of meat, fat, and water for reasonable sausage quality within the variation necessitated by CF additions. The sausages of a pilot plant scale were formulated according to
Table 1. The control sausage was made of 42.5% ice water, 35% lean pork, 20% pork fat, 2% salt, and 0.5% phosphate, by weight. For the hybrid sausages, CF was added at 1% (CF-1.0%), 2.5% (CF-2.5%), and 5.0% (CF-5.0%) levels on top of the base (control) recipe. The day before the preparation, lean pork, pork fat, and CF were thawed overnight at 5 °C. After thawing, lean pork was pre-homogenized with a table cutter (Seydelmann K20, Seydelmann KG, Aalen, Germany) following mixing with salt and a portion of the ice (<0 °C). Chopping was continued by adding pork fat and part of the cold tap water/ice to obtain a fine batter (6–8 °C). Subsequently, CF and the remaining cold tap water/ice were added. Further chopping of the final batter was continued until the batter temperature reached 14 °C. The uncooked batter was stuffed (VINS/22 sausage filler, la Minerva di Chiodini Mario, Bologna, Italy) into a Walsrode K flex polymer casing (45 mm in diameter), (Wolff Walsrode AG, Bomltiz, Germany) with string-closed ends. Sausages were cooked at 75 °C using liquid steam equipment (Kerres CS 350, Kerres GmbH, Backnang, Germany) for 45 min (until the sausage core temperature reached 72 °C). Finally, the cooked sausages were quickly cooled down in an ice bath and stored at 4 °C for further analyses. The preparation of the sausages of each group was conducted once to make the four different batters, and thereafter, the sampling of the sausages was repeated three times. To determine the textural properties and oxidative stability, samples were collected on days 0, 1, 4, 7, and 14 to determine the textural properties and oxidative stability during storage.
The control sausages were formulated with lean pork, pork fat, salt, phosphate, and ice water; the CF sausages were hybrid sausages formulated by adding CF at 1% (CF-1.0%), 2.5% (CF-2.5%), and 5.0% (CF-5.0%) levels to the base (control) recipe.
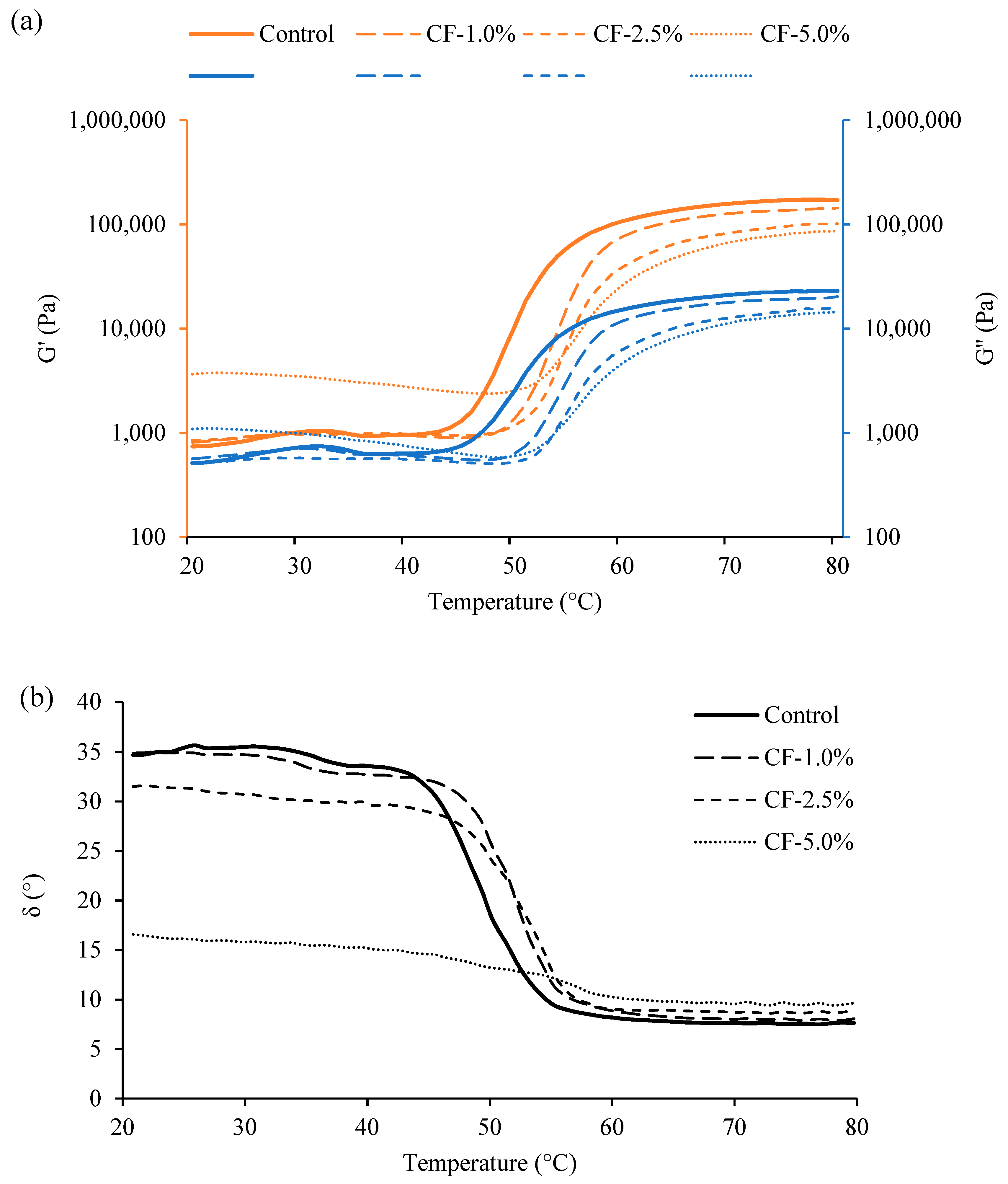
2.5. Rheological Properties of Raw Sausage Batter
The rheological properties of raw sausage batter (before cooking) were measured according to Chen et al. (2020) [
44], with modifications, using a rheometer (HAAKE MARS 40, ThermoScientific, Bremen, Germany) with parallel-plate geometry (35 mm in diameter and 1 mm in gap) in oscillatory mode. A temperature ramp sweep (20–80 °C, 2 °C/min as the heating rate) was conducted. The oscillation frequency was set at 1.0 Hz. The strain (γ
0 = 0.3%) was set according to the amplitude sweep at 20 °C for both the raw and cooked batter (after heating at 80 °C) to ensure that the measurement was conducted within the linear viscoelastic range of each sample. To prevent the sample from drying during the measurement, a sample hood with a solvent trap was used. The storage modulus (G′), loss modulus (G″) and phase angle (δ) were recorded to demonstrate the rheological properties of raw sausage batter.
2.6. Color and Textural Properties of Cooked Sausages
The color of the sausages was measured using a chromometer (CR-400, Konica Minolta, Osaka, Japan). Each sample was cut into slices of 1 cm thick from the middle of the cooked sausage. The color space parameters
L* (lightness),
a* (red coordinate), and
b* (yellow coordinate) were recorded immediately after the cutting of the samples. The total color difference (∆
E*) between the control and CF sausages was calculated according to the following equation:
where ∆
L* is the difference in lightness and darkness (+ = lighter, − = darker), ∆
a* is the difference in red and green (+ = redder, − = greener), and ∆
b* is the difference in yellow and blue (+ = yellower, − = bluer).
Texture profile analysis was performed using a Texture Analyzer (TA XT2 i, Stable Micro Systems, Brookfield, UK) equipped with a 30 kg load cell and a cylinder probe (36 mm in diameter). A two-cycle compression test (each sample was compressed to 40% of the original height in two consecutive cycles) was applied to each sausage sample in a regular shape (26 × 26 × 26 mm). The testing conditions were as follows: pre-test speed 1 mm/s, test speed 1 mm/s, post-test speed 5 mm/s, resting time 5 s, and trigger force 5 g. Each measurement generated a two-cycle force–time curve from which the hardness, springiness, cohesiveness, chewiness, and resilience were calculated by the program.
Fracturability was measured using an Instron device (33 R 4465, Instron, High Wycombe, UK) equipped with a 100 N load cell and a pistol probe (18 mm in diameter, 2 mm in thickness) in penetration mode. Each sausage sample (50 mm in height, 45 mm in diameter) was penetrated 30 mm (60% of the sample height) at rate 100 mm/min. Each penetration generated a force–distance curve. The maximal force (Fmax) during the penetration was recorded to represent the fracturability of the sausage.
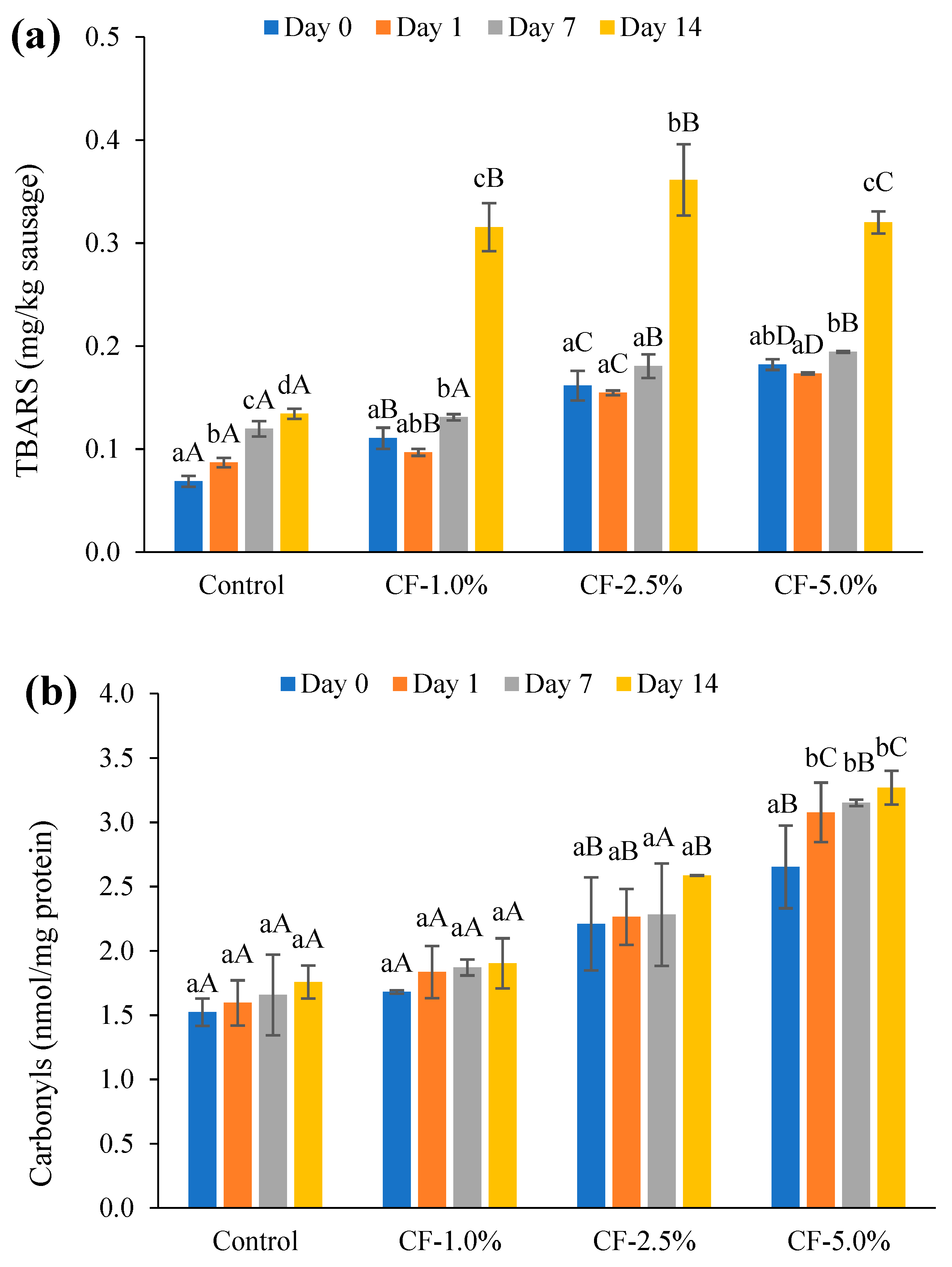
2.7. Lipid Oxidation of Cooked Sausages
Lipid oxidation was monitored by measuring the thiobarbituric acid-reactive substances (TBARS) [
45,
46]. For this process, 5 g of sausage was mixed with 15 mL trichloroacetic acid (5%, g/100 mL) and 0.5 mL butylated hydroxytoluene (4.2% in ethanol, g/100 mL) using a homogenizer (Ultra-Turrax
® T25, IKA, Staufen, Germany) at 13,500 rpm for 30 s in an ice bath. The mixture was filtered (WhatmanTM 42, GE Healthcare Life Sciences, Gillingham, UK), mixed with 0.02 M thiobarbituric acid (1/1,
v/v), and boiled in a water bath (100 °C, 40 min). After cooling, the absorbance was recorded at a wavelength of 532 nm using a spectrophotometer (UV-1800, Shimadzu, Tokyo, Japan). A standard curve of 1,1,3,3-tetraethoxypropane was used to calculate the amount of malondialdehyde produced. The TBARS contents of the sausages were expressed as mg malondialdehyde/kg sausage.
2.8. Protein Oxidation of Cooked Sausages
The carbonyl content was measured by the 2,4-dinitrophenylhydrazine (DNPH) assay [
47,
48] with slight modifications. A sausage sample of 1 g (in duplicate) was mixed well with 10 mL 0.15 M KCl using a homogenizer (Ultra-Turrax
® T25, IKA, Staufen, Germany) at 9500 rpm for 30 s in an ice bath. An aliquot of 100 µL of the homogenate (5 replicates) was mixed with 1 mL of 10% (g/100 mL) TCA and centrifuged (5000×
g, 5 min, r.t.). The pellet was collected, mixed with 400 µL of 5% (g/100 mL) SDS, then subsequently heated (water bath at 100 °C, 10 min) and ultra-sonicated (water bath at 40 °C, 30 min). The mixture was treated with 0.8 mL of 0.3% (g/100 mL) DNPH in 3 M HCl, while the blank (in duplicate) sample was prepared by adding 0.8 mL of 3 M HCl. After incubation (30 min, r.t.), the proteins were precipitated by adding 400 µL of 40% (g/100 mL) TCA following centrifugation (5000×
g, 5 min, r.t.). The pellet was collected and washed three times with 1 mL of ethanol-ethyl acetate (1:1,
v:v) in another centrifugation (10,000×
g, 5 min, r.t.). The resulting pellets were nitrogen-dried, then subsequently dissolved in 1.5 mL of 6 M guanidine hydrochloride in 20 mM NaH
2 PO
4 (pH 6.5) and incubated overnight (4 °C, in the dark). The absorbance was read at 280 and 370 nm. The carbonyl content, expressed as nmol/mg of protein, was calculated as follows:
where 22,000 is the molar extinction coefficient and 0.43 is the coefficient for removing potential hydrazine interference at 280 nm.
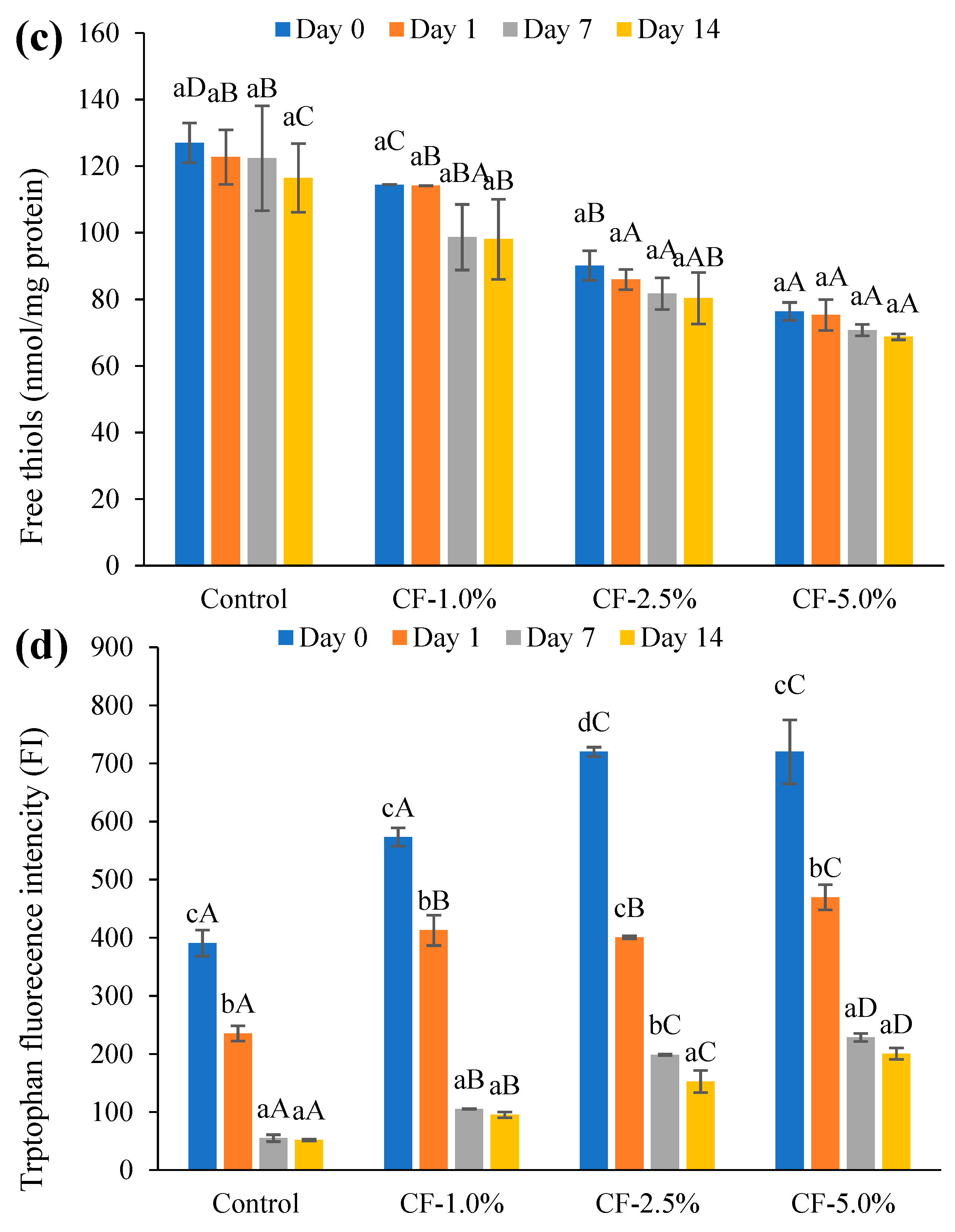
Free thiol content was measured using the 5,5′-dithio-2-nitrobenzoate (DTNB) method [
49,
50]. A sample of 1 g of sausage (in duplicate) was mixed with 25 mL 5% SDS in 0.1 M Tris-HCl (pH 8.0) using a homogenizer (Ultra-Turrax
® T25, IKA, Staufen, Germany) at 12,800–13,500 rpm (30 s, r.t.). The homogenates were heated (water bath at 80 °C, 30 min), cooled to r.t., and filtered (Whatman
® filter paper, Grade 42: 2.5 μm). Thiols were measured by mixing the filtrate, 0.1 M Tris-HCl (pH 8.0), and 10 Mm DTNB in 0.1 M Tris-HCl (pH 8.0) (1/4/1,
v/v/v). A protein blank (filtrate/0.1 M Tris-HCl (pH 8.0), 1/5,
v/v) and reagent blank (0.1 M Tris-HCl (pH 8.0)/10 Mm DTNB in 0.1 M Tris-HCl (pH 8.0), 5/1,
v/v) were prepared for each sample. The mixture was incubated (30 min, r.t., dark), and then absorbance was recorded at 405 nm. The concentration of thiols in the filtrate was calculated using a molar extinction coefficient of 13,600 M
−1 CM
−1 for DTNB at the given wavelength. The protein content in the filtrate was determined according to a DC Protein Assay, where a standard curve was established using BSA at 750 nm. The free thiol content was expressed as nmol thiol/mg protein.
The loss of tryptophan fluorescence intensity (FI) was determined according to Estévez et al. (2008) [
51] using a spectrometer (LS55, PerkinElmer, Waltham, MA, USA). A sausage sample of 1 g (in duplicate) was mixed with 10 mL of ice-cold 0.15 M KCl using a homogenizer (Ultra-Turrax
® T25, IKA, Staufen, Germany) at 9500 rpm for 30 s in an ice bath. The mixture was filtered and diluted by Milli-Q to ~20 µg protein/mL. The emission spectra were recorded from 320 to 460 nm, with the excitation wavelength at 290 nm (slit = 10 nm, speed = 180 nm/min).
2.9. Statistical Analysis
All results are given as mean ± standard deviation (n = 3). To study lipid and protein oxidation of cooked sausages during storage, all data were analyzed by ANOVA under the general linear model using IBM SPSS Statistics software (Version 28.0.0.0 (190)), with storage days and CF addition (CF%) being fixed factors. Tukey’s test (p ≤ 0.05) was used to determine significant differences among the results. Pearson’s correlation test was performed between the indicators of lipid and protein oxidation.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}