

Genetic Characterization of Listeria from Food of Non-Animal Origin Products and from Producing and Processing Companies in Bavaria, Germany
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Isolates from Supermarkets
2.2. Origin of Isolates from FNAO-Producing and -Processing Plants
2.3. WGS
2.4. Data Analysis
2.5. Phenotypic AMR Testing
3. Results
3.1. Occurrence of Listeria spp. in FNAO from Supermarkets
3.2. Occurrence of Listeria spp. in FNAO-Producing and Processing Plants
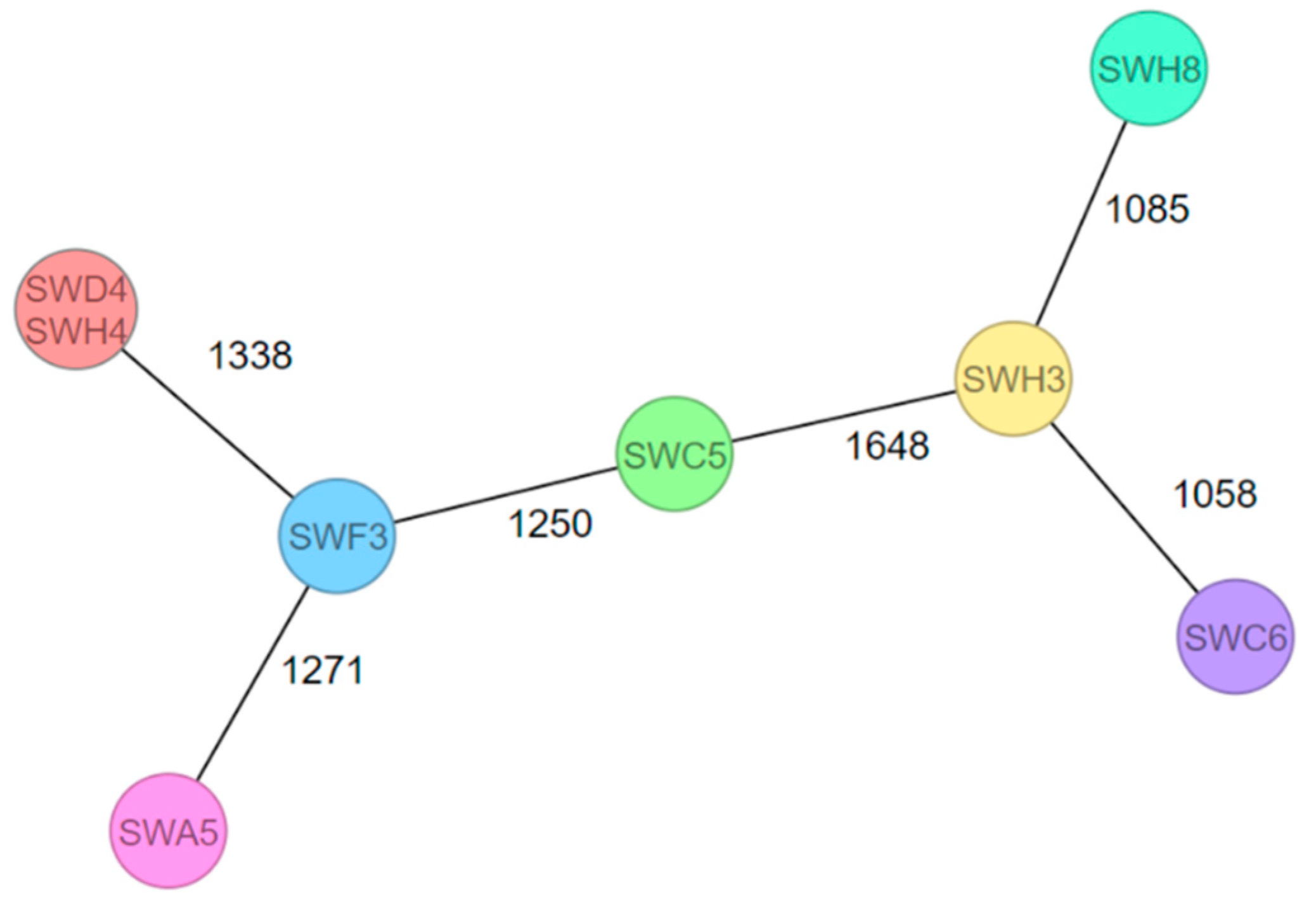
3.3. Genetic Relation of 64 Listeria Isolates Sequenced by WGS
3.4. Serogroup Determination of L. monocytogenes
3.5. MLST Analysis of L. monocytogenes
3.6. cgMLST Analysis of L. monocytogenes
3.7. Prevalence of Genetic and Phenotypic AMRs in Listeria spp.
3.8. Prevalence of Virulence Genes in Listeria spp.
4. Discussion
4.1. Distribution and Genetic Relation of Listeria spp. in FNAO from Supermarkets and FNAO-Producing and Processing Plants
4.2. Serogroup and MLST Analysis of L. monocytogenes
4.3. cgMLST Analysis of L. monocytogenes
4.4. Genetic and Phenotypic AMRs of Listeria spp.
4.5. Prevalence of Virulence Genes in Listeria spp.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newell, D.G.; Koopmans, M.; Verhoef, L.; Duizer, E.; Aidara-Kane, A.; Sprong, H.; Opsteegh, M.; Langelaar, M.; Threfall, J.; Scheutz, F.; et al. Food-borne diseases—The challenges of 20 years ago still persist while new ones continue to emerge. Int. J. Food Microbiol. 2010, 139 (Suppl. 1), S3–S15. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Multistate Outbreaks. Available online: https://www.cdc.gov/foodsafety/outbreaks/multistate-outbreaks/index.html (accessed on 16 January 2023).
- Montero, D.; Bodero, M.; Riveros, G.; Lapierre, L.; Gaggero, A.; Vidal, R.M.; Vidal, M. Molecular epidemiology and genetic diversity of Listeria monocytogenes isolates from a wide variety of ready-to-eat foods and their relationship to clinical strains from listeriosis outbreaks in Chile. Front. Microbiol. 2015, 6, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Whole Cantaloupes from Jensen Farms, Colorado (FINAL UPDATE). Available online: https://www.cdc.gov/listeria/outbreaks/cantaloupes-jensen-farms/index.html (accessed on 16 January 2023).
- Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Frozen Vegetables (Final Update). Available online: https://www.cdc.gov/listeria/outbreaks/frozen-vegetables-05-16/index.html (accessed on 16 January 2023).
- Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Packaged Salads Produced at Springfield, Ohio Dole Processing Facility (Final Update). Available online: https://www.cdc.gov/listeria/outbreaks/bagged-salads-01-16/index.html (accessed on 16 January 2023).
- EFSA. Multi-country outbreak of Listeria monocytogenes serogroup IVb, multi-locus sequence type 6, infections linked to frozen corn and possibly to other frozen vegetables—First update. EFS3 2018, 15, 1448E. [Google Scholar] [CrossRef]
- Gaul, L.K.; Farag, N.H.; Shim, T.; Kingsley, M.A.; Silk, B.J.; Hyytia-Trees, E. Hospital-acquired listeriosis outbreak caused by contaminated diced celery--Texas, 2010. Clin. Infect. Dis. 2013, 56, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.R.; Salter, M.; Tarr, C.; Conrad, A.; Harvey, E.; Steinbock, L.; Saupe, A.; Sorenson, A.; Katz, L.; Stroika, S.; et al. Notes from the field: Listeriosis associated with stone fruit--United States, 2014. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 282–283. [Google Scholar]
- Angelo, K.M.; Conrad, A.R.; Saupe, A.; Dragoo, H.; West, N.; Sorenson, A.; Barnes, A.; Doyle, M.; Beal, J.; Jackson, K.A.; et al. Multistate outbreak of Listeria monocytogenes infections linked to whole apples used in commercially produced, prepackaged caramel apples: United States, 2014-2015. Epidemiol. Infect. 2017, 145, 848–856. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Listeria Outbreaks. Available online: https://www.cdc.gov/listeria/outbreaks/index.html (accessed on 16 January 2023).
- Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Associated with Jensen Farms Cantaloupe: United States, August—September 2011. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm6039a5.htm (accessed on 16 January 2023).
- EFSA. The European Union One Health 2020 Zoonoses Report. EFS2 2021, 19, e05926. [Google Scholar] [CrossRef]
- EFSA. The European Union One Health 2021 Zoonoses Report. EFS2 2022, 20, e07666. [Google Scholar] [CrossRef]
- Pennone, V.; Dygico, K.L.; Coffey, A.; Gahan, C.G.M.; Grogan, H.; McAuliffe, O.; Burgess, C.M.; Jordan, K. Effectiveness of current hygiene practices on minimization of Listeria monocytogenes in different mushroom production-related environments. Food Sci. Nutr. 2020, 8, 3456–3468. [Google Scholar] [CrossRef]
- Lake, F.B.; van Overbeek, L.S.; Baars, J.J.P.; Koomen, J.; Abee, T.; den Besten, H.M.W. Genomic characteristics of Listeria monocytogenes isolated during mushroom (Agaricus bisporus) production and processing. Int. J. Food Microbiol. 2021, 360, 109438. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; Gil, M.I.; Querido-Ferreira, A.P.; Capón, C.L.; Álvarez-Ordoñez, A.; Allende, A. Frozen Vegetable Processing Plants Can Harbour Diverse Listeria monocytogenes Populations: Identification of Critical Operations by WGS. Foods 2022, 11, 1546. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Idland, L.; Heir, E.; Møretrø, T.; Aspholm, M.; Lindbäck, T.; Langsrud, S. Whole-Genome Sequencing Analysis of Listeria monocytogenes from Rural, Urban, and Farm Environments in Norway: Genetic Diversity, Persistence, and Relation to Clinical and Food Isolates. Appl. Environ. Microbiol. 2022, 88, e0213621. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.A.; Srinivasan, M.; Egholm, M.; Shen, Y.; Chen, L.; McGuire, A.; He, W.; Chen, Y.-J.; Makhijani, V.; Roth, G.T.; et al. The complete genome of an individual by massively parallel DNA sequencing. Nature 2008, 452, 872–876. [Google Scholar] [CrossRef] [Green Version]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole genome-based population biology and epidemiological surveillance of Listeria monocytogenes. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef] [Green Version]
- Ronholm, J.; Nasheri, N.; Petronella, N.; Pagotto, F. Navigating Microbiological Food Safety in the Era of Whole-Genome Sequencing. Clin. Microbiol. Rev. 2016, 29, 837–857. [Google Scholar] [CrossRef] [Green Version]
- Wartha, S.; Huber, S.; Kraemer, I.; Alter, T.; Messelhäußer, U. Presence of Listeria at primary production and processing of food of non-animal origin (FNAO) in Bavaria, Germany. J. Food Prot. 2022, 86, 100015. [Google Scholar] [CrossRef]
- International Organization for Standardization. Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria monocytogenes and of Listeria spp.; Beuth Verlag GmbH: Berlin, Germany, 2017. [Google Scholar]
- Illumina. Illumina DNA Prep Reference Guide. Available online: https://sapac.support.illumina.com/content/dam/illumina-support/documents/documentation/chemistry_documentation/illumina_prep/illumina-dna-prep-reference-guide-1000000025416-09.pdf (accessed on 16 January 2023).
- Deneke, C.; Brendebach, H.; Uelze, L.; Borowiak, M.; Malorny, B.; Tausch, S.H. Species-Specific Quality Control, Assembly and Contamination Detection in Microbial Isolate Sequences with AQUAMIS. Genes 2021, 12, 644. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Bayliss, S.C.; Thorpe, H.A.; Coyle, N.M.; Sheppard, S.K.; Feil, E.J. PIRATE: A fast and scalable pangenomics toolbox for clustering diverged orthologues in bacteria. Gigascience 2019, 8, giz119. [Google Scholar] [CrossRef]
- Simonsen, M.; Mailund, T.; Pedersen, C.N.S. Rapid Neighbour-Joining. In Algorithms in Bioinformatics; Crandall, K.A., Lagergren, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 113–122. ISBN 978-3-540-87360-0. [Google Scholar]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Abricate. Available online: https://github.com/tseemann/abricate (accessed on 16 January 2023).
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2016, 45, D566-73. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2015, 44, D694-7. [Google Scholar] [CrossRef] [PubMed]
- Hyden, P.; Pietzka, A.; Lennkh, A.; Murer, A.; Springer, B.; Blaschitz, M.; Indra, A.; Huhulescu, S.; Allerberger, F.; Ruppitsch, W.; et al. Whole genome sequence-based serogrouping of Listeria monocytogenes isolates. J. Biotechnol. 2016, 235, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragon, M.; Wirth, T.; Hollandt, F.; Lavenir, R.; Lecuit, M.; Le Monnier, A.; Brisse, S. A new perspective on Listeria monocytogenes evolution. PLoS Pathog. 2008, 4, e1000146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; von Haeseler, A.; Stoye, J.; et al. Updating benchtop sequencing performance comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef] [Green Version]
- Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Escámez, P.S.F.; Girones, R.; Herman, L.; Koutsoumanis, K.; Nørrung, B.; et al. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. Available online: https://www.efsa.europa.eu/sites/default/files/engage/170724-0.pdf (accessed on 16 January 2023). [PubMed]
- Ruppitsch, W.; Pietzka, A.; Prior, K.; Bletz, S.; Fernandez, H.L.; Allerberger, F.; Harmsen, D.; Mellmann, A. Defining and Evaluating a Core Genome Multilocus Sequence Typing Scheme for Whole-Genome Sequence-Based Typing of Listeria monocytogenes. J. Clin. Microbiol. 2015, 53, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: https://www.eucast.org/ (accessed on 16 January 2023).
- Troxler, R.; Von Graevenitz, A.; Funke, G.; Wiedemann, B.; Stock, I. Natural antibiotic susceptibility of Listeria species: L. grayi, L. innocua, L. ivanovii, L. monocytogenes, L. seeligeri and L. welshimeri strains. Clin. Microbiol. Infect. 2000, 6, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Noll, M.; Kleta, S.; Al Dahouk, S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J. Infect. Public Health 2018, 11, 572–577. [Google Scholar] [CrossRef]
- Guerreiro, D.N.; Arcari, T.; O’Byrne, C.P. The σB-Mediated General Stress Response of Listeria monocytogenes: Life and Death Decision Making in a Pathogen. Front. Microbiol. 2020, 11, 1505. [Google Scholar] [CrossRef] [PubMed]
- Tiensuu, T.; Guerreiro, D.N.; Oliveira, A.H.; O’Byrne, C.; Johansson, J. Flick of a switch: Regulatory mechanisms allowing Listeria monocytogenes to transition from a saprophyte to a killer. Microbiology 2019, 165, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Poimenidou, S.V.; Dalmasso, M.; Papadimitriou, K.; Fox, E.M.; Skandamis, P.N.; Jordan, K. Virulence Gene Sequencing Highlights Similarities and Differences in Sequences in Listeria monocytogenes Serotype 1/2a and 4b Strains of Clinical and Food Origin From 3 Different Geographic Locations. Front. Microbiol. 2018, 9, 1103. [Google Scholar] [CrossRef] [Green Version]
- Matereke, L.T.; Okoh, A.I. Listeria monocytogenes Virulence, Antimicrobial Resistance and Environmental Persistence: A Review. Pathogens 2020, 9, 528. [Google Scholar] [CrossRef]
- Chico-Calero, I.; Suárez, M.; González-Zorn, B.; Scortti, M.; Slaghuis, J.; Goebel, W.; Vázquez-Boland, J.A. Hpt, a bacterial homolog of the microsomal glucose- 6-phosphate translocase, mediates rapid intracellular proliferation in Listeria. Proc. Natl. Acad. Sci. USA 2002, 99, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, D.E.; Agaisse, H. The Metalloprotease Mpl Supports Listeria monocytogenes Dissemination through Resolution of Membrane Protrusions into Vacuoles. Infect. Immun. 2016, 84, 1806–1814. [Google Scholar] [CrossRef] [Green Version]
- Strawn, L.K.; Fortes, E.D.; Bihn, E.A.; Nightingale, K.K.; Gröhn, Y.T.; Worobo, R.W.; Wiedmann, M.; Bergholz, P.W. Landscape and meteorological factors affecting prevalence of three food-borne pathogens in fruit and vegetable farms. Appl. Environ. Microbiol. 2013, 79, 588–600. [Google Scholar] [CrossRef] [Green Version]
- Barnett-Neefs, C.; Sullivan, G.; Zoellner, C.; Wiedmann, M.; Ivanek, R. Using agent-based modeling to compare corrective actions for Listeria contamination in produce packinghouses. PLoS ONE 2022, 17, e0265251. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.; Dessai, U.; McGarry, S.; Gerner-Smidt, P. Use of Whole-Genome Sequencing for Food Safety and Public Health in the United States. Foodborne Pathog. Dis. 2019, 16, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Lakicevic, B.Z.; den Besten, H.M.W.; de Biase, D. Landscape of Stress Response and Virulence Genes Among Listeria monocytogenes Strains. Front. Microbiol. 2021, 12, 738470. [Google Scholar] [CrossRef]
- Ferreira, V.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes persistence in food-associated environments: Epidemiology, strain characteristics, and implications for public health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Mafuna, T.; Matle, I.; Magwedere, K.; Pierneef, R.E.; Reva, O.N. Whole Genome-Based Characterization of Listeria monocytogenes Isolates Recovered From the Food Chain in South Africa. Front. Microbiol. 2021, 12, 669287. [Google Scholar] [CrossRef] [PubMed]
- Šteingolde, Ž.; Meistere, I.; Avsejenko, J.; Ķibilds, J.; Bergšpica, I.; Streikiša, M.; Gradovska, S.; Alksne, L.; Roussel, S.; Terentjeva, M.; et al. Characterization and Genetic Diversity of Listeria monocytogenes Isolated from Cattle Abortions in Latvia, 2013-2018. Vet. Sci. 2021, 8, 195. [Google Scholar] [CrossRef]
- Cabal, A.; Pietzka, A.; Huhulescu, S.; Allerberger, F.; Ruppitsch, W.; Schmid, D. Isolate-Based Surveillance of Listeria monocytogenes by Whole Genome Sequencing in Austria. Front. Microbiol. 2019, 10, 2282. [Google Scholar] [CrossRef]
- Zuber, I.; Lakicevic, B.; Pietzka, A.; Milanov, D.; Djordjevic, V.; Karabasil, N.; Teodorovic, V.; Ruppitsch, W.; Dimitrijevic, M. Molecular characterization of Listeria monocytogenes isolates from a small-scale meat processor in Montenegro, 2011–2014. Food Microbiol. 2019, 79, 116–122. [Google Scholar] [CrossRef] [PubMed]
- McCollum, J.T.; Cronquist, A.B.; Silk, B.J.; Jackson, K.A.; O’Connor, K.A.; Cosgrove, S.; Gossack, J.P.; Parachini, S.S.; Jain, N.S.; Ettestad, P.; et al. Multistate outbreak of listeriosis associated with cantaloupe. N. Engl. J. Med. 2013, 369, 944–953. [Google Scholar] [CrossRef] [Green Version]
- CHEN, Y.I.; Burall, L.S.; Macarisin, D.; Pouillot, R.; Strain, E.; de Jesus, A.J.; Laasri, A.; Wang, H.U.; Ali, L.; Tatavarthy, A.; et al. Prevalence and Level of Listeria monocytogenes in Ice Cream Linked to a Listeriosis Outbreak in the United States. J. Food Prot. 2016, 79, 1828–1832. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Blue Bell Creameries Products (Final Update). Available online: https://www.cdc.gov/Listeria/outbreaks/ice-cream-03-15/ (accessed on 16 January 2023).
- Autio, T.; Keto-Timonen, R.; Lundén, J.; Björkroth, J.; Korkeala, H. Characterisation of persistent and sporadic Listeria monocytogenes strains by pulsed-field gel electrophoresis (PFGE) and amplified fragment length polymorphism (AFLP). Syst. Appl. Microbiol. 2003, 26, 539–545. [Google Scholar] [CrossRef]
- Chasseignaux, E.; Toquin, M.T.; Ragimbeau, C.; Salvat, G.; Colin, P.; Ermel, G. Molecular epidemiology of Listeria monocytogenes isolates collected from the environment, raw meat and raw products in two poultry- and pork-processing plants. J. Appl. Microbiol. 2001, 91, 888–899. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance. Available online: http://www.who.int/iris/bitstream/10665/112642/1/9789241564748_eng.pdf?ua=1 (accessed on 5 March 2023).
- Poyart-Salmeron, C. Transferable plasmid-mediated antibiotic resistance in Listeria monocytogenes. Lancet 1990, 335, 1422–1426. [Google Scholar] [CrossRef] [PubMed]
- Parra-Flores, J.; Holý, O.; Bustamante, F.; Lepuschitz, S.; Pietzka, A.; Contreras-Fernández, A.; Castillo, C.; Ovalle, C.; Alarcón-Lavín, M.P.; Cruz-Córdova, A.; et al. Virulence and Antibiotic Resistance Genes in Listeria monocytogenes Strains Isolated From Ready-to-Eat Foods in Chile. Front. Microbiol. 2021, 12, 796040. [Google Scholar] [CrossRef] [PubMed]
- Tîrziu, E.; Herman, V.; Nichita, I.; Morar, A.; Imre, M.; Ban-Cucerzan, A.; Bucur, I.; Tîrziu, A.; Mateiu-Petrec, O.C.; Imre, K. Diversity and Antibiotic Resistance Profiles of Listeria monocytogenes Serogroups in Different Food Products from the Transylvania Region of Central Romania. J. Food Prot. 2022, 85, 54–59. [Google Scholar] [CrossRef] [PubMed]
- de Vasconcelos Byrne, V.; Hofer, E.; Vallim, D.C.; de Castro Almeida, R.C. Occurrence and antimicrobial resistance patterns of Listeria monocytogenes isolated from vegetables. Braz. J. Microbiol. 2016, 47, 438–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gygli, S.M.; Keller, P.M.; Ballif, M.; Blöchliger, N.; Hömke, R.; Reinhard, M.; Loiseau, C.; Ritter, C.; Sander, P.; Borrell, S.; et al. Whole-Genome Sequencing for Drug Resistance Profile Prediction in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2019, 63, e02175-18. [Google Scholar] [CrossRef] [Green Version]
- Kovačević, J.; Mesak, L.R.; Allen, K.J. Occurrence and characterization of Listeria spp. in ready-to-eat retail foods from Vancouver, British Columbia. Food Microbiol. 2012, 30, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yu, T.; Xu, P.; Xu, X.; Ji, S.; Gao, W.; Shi, L. Role of Efflux Pumps in the in vitro Development of Ciprofloxacin Resistance in Listeria monocytogenes. Front. Microbiol. 2018, 9, 2350. [Google Scholar] [CrossRef]
- Morvan, A.; Moubareck, C.; Leclercq, A.; Hervé-Bazin, M.; Bremont, S.; Lecuit, M.; Courvalin, P.; Le Monnier, A. Antimicrobial resistance of Listeria monocytogenes strains isolated from humans in France. Antimicrob. Agents Chemother. 2010, 54, 2728–2731. [Google Scholar] [CrossRef] [Green Version]
- Scortti, M.; Han, L.; Alvarez, S.; Leclercq, A.; Moura, A.; Lecuit, M.; Vazquez-Boland, J. Epistatic control of intrinsic resistance by virulence genes in Listeria. PLoS Genet. 2018, 14, e1007525. [Google Scholar] [CrossRef] [Green Version]
- Da Rocha, L.S.; Gunathilaka, G.U.; Zhang, Y. Antimicrobial-resistant Listeria species from retail meat in metro Detroit. J. Food Prot. 2012, 75, 2136–2141. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.A.; Jackson, C.R. Comparative antimicrobial susceptibility of Listeria monocytogenes, L. innocua, and L. welshimeri. Microb. Drug Resist. 2009, 15, 27–32. [Google Scholar] [CrossRef]
- Li, Q.; Sherwood, J.S.; Logue, C.M. Antimicrobial resistance of Listeria spp. recovered from processed bison. Lett. Appl. Microbiol. 2007, 44, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.L.; Rojo, F. Metabolic regulation of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 768–789. [Google Scholar] [CrossRef] [PubMed]
- Weller, D.; Andrus, A.; Wiedmann, M.; den Bakker, H.C. Listeria booriae sp. nov. and Listeria newyorkensis sp. nov., from food processing environments in the USA. Int. J. Syst. Evol. Microbiol. 2015, 65, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, C.; Takahashi, H.; Nakamura, A.; Kuda, T.; Kimura, B. Whole-Genome Sequence of Listeria newyorkensis, Isolated from River Water in Japan. Microbiol. Resour. Announc. 2019, 8, e00488-19. [Google Scholar] [CrossRef] [Green Version]
- Gaballa, A.; Guariglia-Oropeza, V.; Wiedmann, M.; Boor, K.J. Cross Talk between SigB and PrfA in Listeria monocytogenes Facilitates Transitions between Extra- and Intracellular Environments. Microbiol. Mol. Biol. Rev. 2019, 83, e00034-19. [Google Scholar] [CrossRef]
- Jacquet, C.; Doumith, M.; Gordon, J.I.; Martin, P.M.V.; Cossart, P.; Lecuit, M. A molecular marker for evaluating the pathogenic potential of foodborne Listeria monocytogenes. J. Infect. Dis. 2004, 189, 2094–2100. [Google Scholar] [CrossRef] [Green Version]
- Quereda, J.J.; Morón-García, A.; Palacios-Gorba, C.; Dessaux, C.; García-Del Portillo, F.; Pucciarelli, M.G.; Ortega, A.D. Pathogenicity and virulence of Listeria monocytogenes: A trip from environmental to medical microbiology. Virulence 2021, 12, 2509–2545. [Google Scholar] [CrossRef]
- Vilchis-Rangel, R.E.; Del Espinoza-Mellado, M.R.; Salinas-Jaramillo, I.J.; Martinez-Peña, M.D.; Rodas-Suárez, O.R. Association of Listeria monocytogenes LIPI-1 and LIPI-3 marker llsX with invasiveness. Curr. Microbiol. 2019, 76, 637–643. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, Y.; Pettengill, J.; Timme, R.; Melka, D.; Doyle, M.; Jackson, A.; Parish, M.; Hammack, T.S.; Allard, M.W.; et al. Singleton Sequence Type 382, an Emerging Clonal Group of Listeria monocytogenes Associated with Three Multistate Outbreaks Linked to Contaminated Stone Fruit, Caramel Apples, and Leafy Green Salad. J. Clin. Microbiol. 2017, 55, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Disson, O.; Moura, A.; Lecuit, M. Making Sense of the Biodiversity and Virulence of Listeria monocytogenes. Trends Microbiol. 2021, 29, 811–822. [Google Scholar] [CrossRef]
- Guillet, C.; Join-Lambert, O.; Le Monnier, A.; Leclercq, A.; Mechaï, F.; Mamzer-Bruneel, M.F.; Bielecka, M.K.; Scortti, M.; Disson, O.; Berche, P.; et al. Human listeriosis caused by Listeria ivanovii. Emerg. Infect. Dis. 2010, 16, 136–138. [Google Scholar] [CrossRef]
- Gouin, E.; Mengaud, J.; Cossart, P. The virulence gene cluster of Listeria monocytogenes is also present in Listeria ivanovii, an animal pathogen, and Listeria seeligeri, a nonpathogenic species. Infect. Immun. 1994, 62, 3550–3553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaillard, J.-L.; Berche, P.; Frehel, C.; Gouln, E.; Cossart, P. Entry of L. monocytogenes into cells is mediated by internalin, a repeat protein reminiscent of surface antigens from gram-positive cocci. Cell 1991, 65, 1127–1141. [Google Scholar] [CrossRef]
- Beye, M.; Gouriet, F.; Michelle, C.; Casalta, J.-P.; Habib, G.; Raoult, D.; Fournier, P.-E. Genome analysis of Listeria ivanovii strain G770 that caused a deadly aortic prosthesis infection. New Microbes New Infect. 2016, 10, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreft, J.; Vázquez-Boland, J.A. Regulation of virulence genes in Listeria. Int. J. Med. Microbiol. 2001, 291, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Karunasagar, I.; Lampidis, R.; Goebel, W.; Kreft, J. Complementation of Listeria seeligeri with the plcA-prfA genes from L. monocytogenes activates transcription of seeligerolysin and leads to bacterial escape from the phagosome of infected mammalian cells. FEMS Microbiol. Lett. 1997, 146, 303–310. [Google Scholar] [CrossRef]
- Orsi, R.H.; Wiedmann, M. Characteristics and distribution of Listeria spp., including Listeria species newly described since 2009. Appl. Microbiol. Biotechnol. 2016, 100, 5273–5287. [Google Scholar] [CrossRef] [Green Version]
- Rocourt, J.; Schrettenbrunner, A.; Hof, H.; Espaze, E.P. Une nouvelle espèce du genre Listeria: Listeria seeligeri. Pathol. Biol. 1987, 35, 1075–1080. [Google Scholar]


{kind=link}
| Isolate | Plant (PP or PC) | Sample | Serogroup and Serotype (In Silico) | Sequence Type (ST) (MLST) | Clonal Complex (CC) (MLST) | Complex Type (CT) (cgMLST) |
|---|---|---|---|---|---|---|
| SWH3 | K (PC) | Drain—delivery raw goods (S) | IVb (4b, 4d, and 4e) | ST1 | CC1 | 16888 |
| SWH8 | I (PP) | Drain—cooling area lettuce (S) | IVb (4b, 4d, and 4e) | ST2 | CC2 | 16889 |
| SWC6 | M (PP) | Condensate ponding of a cooling unit (S) | IVb (4b, 4d, and 4e) | ST6 | CC6 | 7504 |
| SWA5 | N (PC) | Drain—frozen food packaging (S) | IIa (1/2a and 3a) | ST7 | CC7 | 16884 |
| SWF3 | G (PC) | Drain—packaging area lettuce (S) | IIa (1/2a and 3a) | ST21 | CC21 | 16887 |
| SWD4 | I (PP) | Processing water for a salad-washing system (PW) | IIa (1/2a and 3a) | ST504 | CC475 | 16886 |
| SWH4 | I (PP) | Processing water for a salad-washing system (PW) | IIa (1/2a and 3a) | ST504 | CC475 | 16886 |
| SWC5 | E (PC) | Drain—prewashing area (S) | IIa (1/2a and 3a) | ST1413 | CC739 | 16885 |
| No. | Isolate | Listeria spp. | Sample | Resistance Gene(s) | Resistance | Phenotypical Resistance |
|---|---|---|---|---|---|---|
| 1 | SWA5 | L. monocytogenes | Swab | fosX, mprF, lin, norB | Fosfomycin, cationic peptide, lincomycin, fluoroquinolones | PEN, FOS, MOX |
| 2 | SWC5 | L. monocytogenes | Swab | fosX, mprF, lin, norB | Fosfomycin, cationic peptide, lincomycin, fluoroquinolones | PEN, FOS, CIP, MOX |
| 3 | SWC6 | L. monocytogenes | Swab | fosX, mprF, lin, norB | Fosfomycin, cationic peptide, lincomycin, fluoroquinolones | PEN, GEN, FOS, MOX |
| 4 | SWH8 | L. monocytogenes | Swab | fosX, mprF, lin, norB | Fosfomycin, cationic peptide, lincomycin, fluoroquinolones | PEN, FOS, CIP, MOX |
| 5 | SWH3 | L. monocytogenes | Swab | fosX, mprF, lin, norB | Fosfomycin, cationic peptide, lincomycin, fluoroquinolones | PEN, FOS, CIP, MOX |
| 6 | SWF3 | L. monocytogenes | Swab | fosX, mprF, lin, norB | Fosfomycin, cationic peptide, lincomycin, fluoroquinolones | PEN, FOS, CIP, MOX |
| 7 | SWD4 | L. monocytogenes | Water | fosX, mprF, lin, norB | Fosfomycin, cationic peptide, lincomycin, fluoroquinolones | PEN, FOS, MOX |
| 8 | SWH4 | L. monocytogenes | Water | fosX, mprF, lin, norB | Fosfomycin, cationic peptide, lincomycin, fluoroquinolones | PEN, FOS, CIP, MOX |
| 9 | SWA11 | L. innocua | Swab | norB | Fluoroquinolones | PEN, FOS, CIP, MOX |
| 10 | SWB4 | L. innocua | Swab | norB | Fluoroquinolones | PEN, FOS, CIP, MOX |
| 11 | SWB11 | L. innocua | Swab | norB | Fluoroquinolones | PEN, Trim-sulfa, FOS, CIP, MOX |
| 12 | SWA1 | L. innocua | Swab | norB | Fluoroquinolones | PEN, FOS, CIP, MOX |
| 13 | SWF1 | L. innocua | Swab | norB | Fluoroquinolones | PEN, FOS, CIP, MOX |
| 14 | SWF5 | L. innocua | Food | norB | Fluoroquinolones | PEN, FOS, CIP, MOX |
| 15 | SWG7 | L. innocua | Food | norB | Fluoroquinolones | PEN, FOS, CIP, MOX |
| 16 | SWH6 | L. innocua | Swab | norB, tetM, ANT(6)-la | Fluoroquinolones, tetracycline, aminoglycoside nucleotidyltransferase | PEN, Trim-sulfa, GEN, TET, FOS, CIP, MOX |
| 17 | SWG3 | L. seeligeri | Swab | norB | Fluoroquinolones | PEN, FOS, CIP, MOX |
| Antibiotics | MIC Breakpoints (mg/L) | Number of Resistant Isolates (n) | ||||
|---|---|---|---|---|---|---|
| S ≤ 4 | R > 4 | L. monocytogenes | L. innocua | L. seeligeri | L. newyorkensis | |
| PEN 2 | 0.125 | 0.125 | 8/8 | 8/8 | 1/1 | 5/5 |
| ERY 1 | 1 | 1 | 0/8 | 0/8 | 0/1 | 5/5 |
| Trim-sulfa 1, 3 | 0.06 | 0.06 | 0/8 | 2/8 | 0/1 | 0/5 |
| GEN 2 | 2 | 2 | 1/8 | 1/8 | 0/1 | 0/5 |
| TET 2 | 1 | 2 | 0/8 | 1/8 | 0/1 | 0/5 |
| FOS 2 | 32 | 32 | 8/8 | 8/8 | 1/1 | 5/5 |
| CIP 2 | 0.001 | 1 | 5/8 | 8/8 | 1/1 | 0/5 |
| MOX 2 | 0.25 | 0.25 | 8/8 | 8/8 | 1/1 | 0/5 |
| prfA % (n) | hly % (n) | plcA % (n) | plcB % (n) | hpt % (n) | actA % (n) | inlA % (n) | inlB % (n) | mpl % (n) | |
|---|---|---|---|---|---|---|---|---|---|
| L. monocytogenes | 100 (8/8) | 100 (8/8) | 100 (8/8) | 100 (8/8) | 100 (8/8) | 100 (8/8) | 100 (8/8) | 100 (8/8) | 100 (8/8) |
| L. innocua | 0.00 (0/8) | 0.00 (0/8) | 0.00 (0/8) | 0.00 (0/8) | 0.00 (0/8) | 0.00 (0/8) | 0.00 (0/8) | 0.00 (0/8) | 0.00 (0/8) |
| L. ivanovii | 66.67 (4/6) | 66.67 (4/6) | 0.00 (0/6) | 0.00 (0/6) | 100 (6/6) | 0.00 (0/6) | 66.67 (4/6) | 66.67 (4/6) | 0.00 (0/6) |
| L. seeligeri | 91.67 (33/36) | 91.67 (33/36) | 91.67 (33/36) | 91.67 (33/36) | 100 (36/36) | 0.00 (0/36) | 0.00 (0/36) | 0.00 (0/36) | 91.67 (33/36) |
| L. newyorkensis | 0.00 (0/5) | 0.00 (0/5) | 0.00 (0/5) | 0.00 (0/5) | 0.00 (0/5) | 0.00 (0/5) | 0.00 (0/5) | 0.00 (0/5) | 0.00 (0/5) |
| L. grayi | 0.00 (0/1) | 0.00 (0/1) | 0.00 (0/1) | 0.00 (0/1) | 0.00 (0/1) | 0.00 (0/1) | 0.00 (0/1) | 0.00 (0/1) | 0.00 (0/1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wartha, S.; Bretschneider, N.; Dangel, A.; Hobmaier, B.; Hörmansdorfer, S.; Huber, I.; Murr, L.; Pavlovic, M.; Sprenger, A.; Wenning, M.; et al. Genetic Characterization of Listeria from Food of Non-Animal Origin Products and from Producing and Processing Companies in Bavaria, Germany. Foods 2023, 12, 1120. https://doi.org/10.3390/foods12061120
Wartha S, Bretschneider N, Dangel A, Hobmaier B, Hörmansdorfer S, Huber I, Murr L, Pavlovic M, Sprenger A, Wenning M, et al. Genetic Characterization of Listeria from Food of Non-Animal Origin Products and from Producing and Processing Companies in Bavaria, Germany. Foods. 2023; 12(6):1120. https://doi.org/10.3390/foods12061120
Chicago/Turabian StyleWartha, Simone, Nancy Bretschneider, Alexandra Dangel, Bernhard Hobmaier, Stefan Hörmansdorfer, Ingrid Huber, Larissa Murr, Melanie Pavlovic, Annika Sprenger, Mareike Wenning, and et al. 2023. "Genetic Characterization of Listeria from Food of Non-Animal Origin Products and from Producing and Processing Companies in Bavaria, Germany" Foods 12, no. 6: 1120. https://doi.org/10.3390/foods12061120