
Perspectives in the Application of High, Medium, and Low Molecular Weight Oat β-d-Glucans in Dietary Nutrition and Food Technology—A Short Overview
 , , , ,
, , , ,
Abstract
:1. Introduction
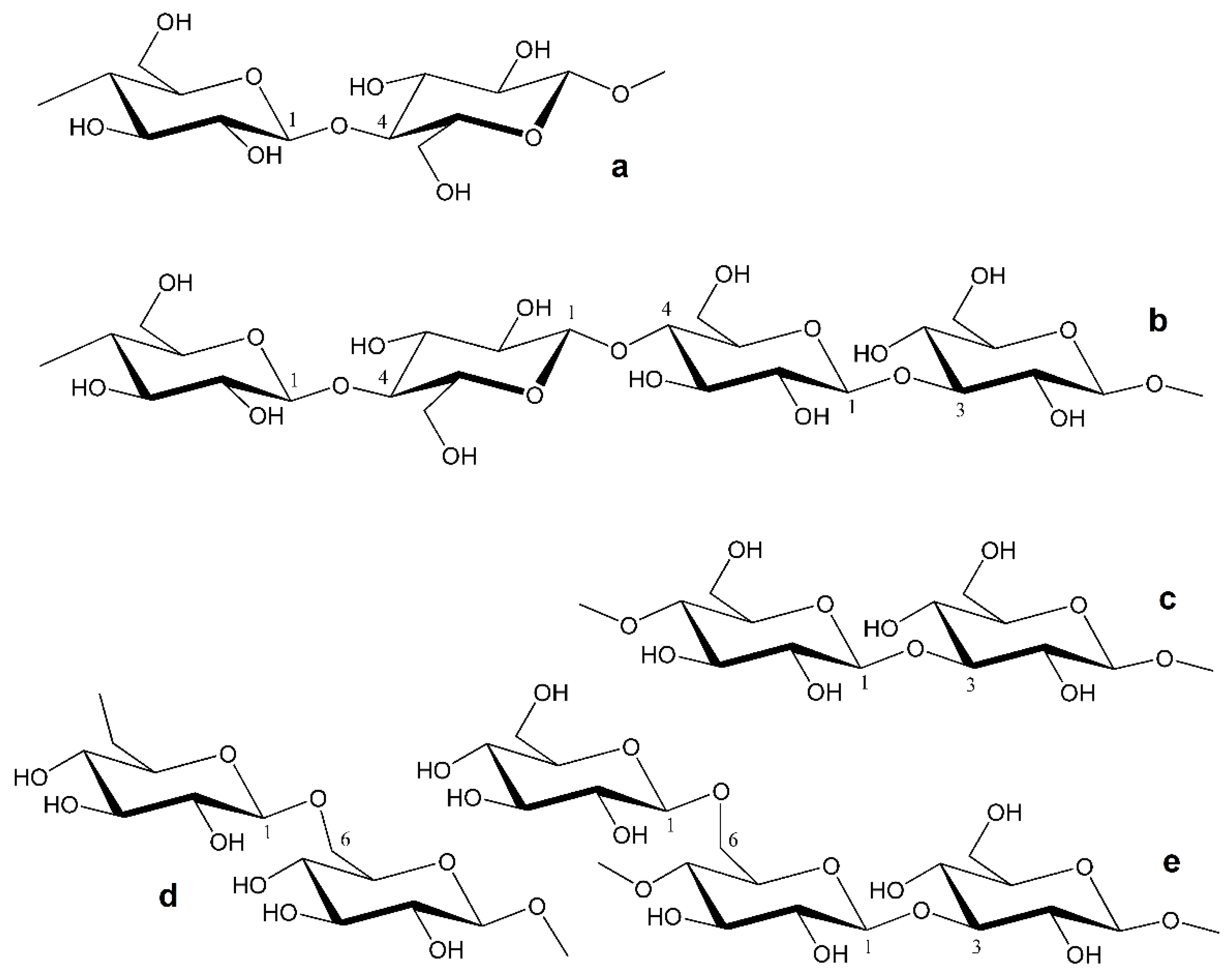
- branched (1 → 3) (1 → 6)- β-d-glucans of various complexity (in terms of branching) from fungi [19,20,21,22,23,24,25,26,27,28,29] and algae [5,6], usually consisting of 1,3-linked β-d-glucopyranosyl units in the backbone and terminal β-d-glucopyranosyls as side chains linked at the O-6 position (Figure 1e).
2. Oats: Health Benefits and the Source of Mixed-Linkage β-d-Glucan
3. Physical Properties and Mw of β-d-Glucans
4. Possible Molecular Mechanisms of Oat β-d-Glucan Activities
5. Preparation of Oat β-d-Glucans with Various Mw
- endo-β-1,3-1,4-glucanases or lichenases (EC 3.2.1.73);
- endo-β-1,3(4)-glucanase (EC 3.2.1.6);
- endo-β-1,3-glucanases or laminarinases (EC 3.2.1.39);
- endo-β-1,4-glucanases or cellulases (EC 3.2.1.4).

{kind=link}
| Enzyme | EC Number | Source | Cleavage of Bonds | Effect on Oat β-d-Glucans | References |
|---|---|---|---|---|---|
| Pancreatin | EC 3.4.21.70 | Pork pancreas | Internal peptide bonds | Release of β-d-glucan from oat products (the upper intestine model) | [72] |
| Endo-1,4-α- glucanase (α-amylase) | EC 3.2.1.1 | Bacillus licheniformis | Internal 1 → 4 bonds | ||
| Endo-1,4-β- glucanase (cellulase) | EC 3.2.1.4 | Aspergillus niger Trichoderma sp. | Internal 1 → 4 bonds | More potent activation of Dectin-1 | [88,89,90] |
| Decrease in Mw Reduction of viscosity | |||||
| Endo-1,3(4)-β- glucanase | EC 3.2.1.6 | Wheat, rye, barley | Internal 1 → 3- or 1 → 4 bonds | [85,86,87] | |
| Endo-1,3:1,4-β- glucanase (lichenase) | EC 3.2.1.73 | Bacillus subtilis | Internal 1 → 4 bonds | [72] |
6. Functionalized Oat β-d-Glucans
| Reaction | Reagent | Product | DS | Properties Tested | Ref. |
|---|---|---|---|---|---|
| Oxidation | TEMPO/NaClO/NaClO2 | 6-carboxy-β-d-glucan | 0.65 | bile acid binding | [106] |
| NaIO4 | β-d-glucan dialdehyde | ||||
| Oxidation Reductive amination | HCOH NH4OAc/NaBH4CN | cationic β-d-glucan | 0.48 | [99] | |
| antimicrobial | |||||
| ACE inhibitor | |||||
| Etherification | NaCH3COCl | CM β-d-glucan | 0.06 | paper improving | [100] |
| Esterification | Ac2O | O-acetylated β-d-glucan | 0.03–0.12 | swelling, rheology | [82] |
| bile acid binding | |||||
| HCOH, HSO3Cl propylene oxide | sulfated β-d-glucan | 0.68 | [101] | ||
| solubility | |||||
| anticoagulant | |||||
| ferulic acid | O-feruloylated β-d-glucan | 0.03–0.18 | stability, anticancer | [103] | |
| antioxidant | |||||
| Maillard reaction | amino acids | conjugates | rheology | [107] | |
| peptides |
7. Influence on Gastrointestinal Diseases and Inflammation
8. Cholesterol Regulation and Bile Acid Binding
9. Anti-Diabetic Activities
10. Antioxidant Activity
11. Prebiotic Activity
12. Anticancer Activity
13. The Application Prospect of Food Technology
14. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Synytsya, A.; Novák, M. Structural Diversity of Fungal Glucans. Carbohydr. Polym. 2013, 92, 792–809. [Google Scholar] [CrossRef]
- Synytsya, A.; Novak, M. Structural Analysis of Glucans. Ann. Transl. Med. 2014, 2, 17. [Google Scholar] [PubMed]
- Ohno, N. Structural Diversity and Physiological Functions of β-Glucans. Int. J. Med. Mushrooms 2005, 7, 167–174. [Google Scholar] [CrossRef]
- Hu, X.; Zhao, J.; Zhao, Q.; Zheng, J. Structure and Characteristic of β-Glucan in Cereal: A Review. J. Food Process. Preserv. 2015, 39, 3145–3153. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, Structure and Biofunctional Activities of Laminarin from Brown Algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Belik, A.A.; Kusaykin, M.I.; Malyarenko, O.S.; Zvyagintseva, T.N.; Ermakova, S.P. Laminarans and 1,3-β-d-Glucanases. Int. J. Biol. Macromol. 2020, 163, 1010–1025. [Google Scholar] [CrossRef] [PubMed]
- Lazaridou, A.; Biliaderis, C.G.; Micha-Screttas, M.; Steele, B.R. A Comparative Study on Structure–Function Relations of Mixed-Linkage (1→3), (1→4) Linear β-d-Glucans. Food Hydrocol. 2004, 18, 837–855. [Google Scholar] [CrossRef]
- Eichhorn, S.J.; Baillie, C.A.; Zafeiropoulos, N.; Mwaikambo, L.Y.; Ansell, M.P.; Dufresne, A.; Entwistle, K.M.; Herrera-Franco, P.J.; Escamilla, G.C.; Groom, L.; et al. Current International Research into Cellulosic Fibres and Composites. J. Mater. Sci. 2001, 36, 2107–2131. [Google Scholar] [CrossRef]
- Chawla, P.R.; Bajaj, I.B.; Survase, S.A.; Sin ghal, R.S. Microbial Cellulose: Fermentative Production and Applications. Food Technol. Biotechnol. 2009, 47, 107–124. [Google Scholar]
- Cheng, Y.; Mondal, A.K.; Wu, S.; Xu, D.; Ning, D.; Ni, Y.; Huang, F. Study on the Anti-Biodegradation Property of Tunicate Cellulose. Polymers 2020, 12, 3071. [Google Scholar] [CrossRef]
- Limberger-Bayer, V.M.; de Francisco, A.; Chan, A.; Oro, T.; Ogliari, P.J.; Barreto, P.L. Barley β-Glucans Extraction and Partial Characterization. Food Chem. 2014, 154, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Wood, P.J. Oat and Rye β-Glucan: Properties and Function. Cereal Chem. 2010, 87, 315–330. [Google Scholar] [CrossRef]
- Cui, W.; Wood, P.J.; Weisz, J.; Beer, M.U. Nonstarch Polysaccharides from Preprocessed Wheat Bran: Carbohydrate Analysis and Novel Rheological Properties. Cereal Chem. 1999, 76, 129–133. [Google Scholar] [CrossRef]
- Nakashima, A.; Yamada, K.; Iwata, O.; Sugimoto, R.; Atsuji, K.; Ogawa, T.; Ishibashi-Ohgo, N.; Suzuki, K. β-Glucan in Foods and Its Physiological Functions. J. Nutr. Sci. Vitaminol. 2018, 64, 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barsanti, L.; Gualtieri, P. Paramylon, a Potent Immunomodulator from WZSL Mutant of Euglena Gracilis. Molecules 2019, 24, 3114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Q.; Zhang, L.; Xu, X.; Zhang, X.; Wu, C. Solution Properties of Pachyman from Poria cocos Mycelia in Dimethyl Sulfoxide. J. Macromol. Sci. B 2001, 40, 147–156. [Google Scholar] [CrossRef]
- Pereyra, M.T.; Prieto, A.; Bernabé, M.; Leal, J.A. Studies of New Polysaccharides from Lasallia pustulata (L.) Hoffm. Lichenologist 2003, 35, 177–185. [Google Scholar] [CrossRef]
- Alves da Cunha, M.A.; Turmina, J.A.; Ivanov, R.C.; Barroso, R.R.; Marques, P.T.; Fonseca, E.A.; Fortes, Z.B.; Dekker, R.F.; Khaper, N.; Barbosa, A.M. Lasiodiplodan, an Exocellular (1→ 6)-β-d-Glucan from Lasiodiplodia theobromae MMPI: Production on Glucose, Fermentation Kinetics, Rheology and Anti-Proliferative Activity. J. Ind. Microbiol. Biotechnol. 2012, 39, 1179–1188. [Google Scholar] [CrossRef]
- Mandal, S.; Maity, K.K.; Bhunia, S.K.; Dey, B.; Patra, S.; Sikdar, S.R.; Islam, S.S. Chemical Analysis of New Water-Soluble (1→ 6)-,(1→ 4)-α, β-Glucan and Water-Insoluble (1→ 3)-,(1→ 4)-β-Glucan (Calocyban) from Alkaline Extract of an Edible Mushroom, Calocybe indica (Dudh Chattu). Carbohydr. Res. 2010, 345, 2657–2663. [Google Scholar] [CrossRef]
- Barbosa, A.M.; Steluti, R.M.; Dekker, R.F.; Cardoso, M.S.; Da Silva, M.C. Structural Characterization of Botryosphaeran: A (1→ 3; 1→ 6)-β-d-Glucan Produced by the Ascomyceteous Fungus, Botryosphaeria sp. Carbohydr. Res. 2003, 338, 1691–1698. [Google Scholar] [CrossRef]
- Schmid, F.; Stone, B.A.; Brownlee, R.T.; McDougall, B.M.; Seviour, R.J. Structure and Assembly of Epiglucan, the Extracellular (1→ 3; 1→ 6)-β-Glucan Produced by the Fungus Epicoccum nigrum Strain F19. Carbohydr. Res. 2006, 341, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Tada, R.; Adachi, Y.; Ishibashi, K.; Ohno, N. An Unambiguous Structural Elucidation of a 1, 3-β-d-Glucan Obtained from Liquid-Cultured Grifola Frondosa by Solution NMR Experiments. Carbohydr. Res. 2009, 344, 400–404. [Google Scholar] [CrossRef]
- Kobayashi, H.; Matsunaga, K.; Oguchi, Y. Antimetastatic Effects of PSK (Krestin), a Protein-Bound Polysaccharide Obtained from Basidiomycetes: An Overview. Cancer Epidemiol. Biomarkers Prev. 1995, 4, 275–281. [Google Scholar] [PubMed]
- Zhang, Y.; Li, S.; Wang, X.; Zhang, L.; Cheung, P.C. Advances in Lentinan: Isolation, Structure, Chain Conformation and Bioactivities. Food Hydrocol. 2011, 25, 196–206. [Google Scholar] [CrossRef]
- Misaki, A.; Kawaguchi, K.; Miyaji, H.; Nagae, H.; Hokkoku, S.; Kakuta, M.; Sasaki, T. Structure of Pestalotan, a Highly Branched (1→ 3)-β-D-Glucan Elaborated by Pestalotia Sp. 815, and the Enhancement of Its Antitumor Activity by Polyol Modification of the Side Chains. Carbohydr. Res. 1984, 129, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Karácsonyi, Š.; Kuniak, L. Polysaccharides of Pleurotus ostreatus: Isolation and Structure of Pleuran, an Alkali-Insoluble β-d-Glucan. Carbohydr. Polym. 1994, 24, 107–111. [Google Scholar] [CrossRef]
- Zhang, Y.; Kong, H.; Fang, Y.; Nishinari, K.; Phillips, G.O. Schizophyllan: A Review on Its Structure, Properties, Bioactivities and Recent Developments. Bioact. Carbohydr. Diet. Fibre 2013, 1, 53–71. [Google Scholar] [CrossRef]
- Coviello, T.; Palleschi, A.; Grassi, M.; Matricardi, P.; Bocchinfuso, G.; Alhaique, F. Scleroglucan: A Versatile Polysaccharide for Modified Drug Delivery. Molecules 2005, 10, 6–33. [Google Scholar] [CrossRef] [Green Version]
- Fizpatrick, F.W.; DiCarlo, F.J. Zymosan. Ann. N. Y. Acad. Sci. 1964, 118, 235–261. [Google Scholar] [CrossRef]
- Khan, A.A.; Gani, A.; Khanday, F.A.; Masoodi, F.A. Biological and Pharmaceutical Activities of Mushroom β-Glucan Discussed as a Potential Functional Food Ingredient. Bioact. Carbohydr. Diet. Fibre 2018, 16, 1–13. [Google Scholar] [CrossRef]
- Zielke, C.; Kosik, O.; Ainalem, M.-L.; Lovegrove, A.; Stradner, A.; Nilsson, L. Characterization of Cereal β-Glucan Extracts from Oat and Barley and Quantification of Proteinaceous Matter. PLoS ONE 2017, 12, e0172034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielke, C.; Stradner, A.; Nilsson, L. Characterization of Cereal β-Glucan Extracts: Conformation and Structural Aspects. Food Hydrocol. 2018, 79, 218–227. [Google Scholar] [CrossRef]
- Ronda, F.; Perez-Quirce, S.; Lazaridou, A.; Biliaderis, C.G. Effect of Barley and Oat β-Glucan Concentrates on Gluten-Free Rice-Based Doughs and Bread Characteristics. Food Hydrocol. 2015, 48, 197–207. [Google Scholar] [CrossRef]
- Stewart, D.; McDougall, G. Oat Agriculture, Cultivation and Breeding Targets: Implications for Human Nutrition and Health. Br. J. Nutr. 2014, 112, S50–S57. [Google Scholar] [CrossRef] [Green Version]
- Angelov, A.; Yaneva-Marinova, T.; Gotcheva, V. Oats as a Matrix of Choice for Developing Fermented Functional Beverages. J. Food Sci. Technol. 2018, 55, 2351–2360. [Google Scholar] [CrossRef]
- Tsopmo, A. Processing Oats and Bioactive Components. In Processing and Impact on Active Components in Food; Elsevier: Amsterdam, The Netherlands, 2015; pp. 361–368. [Google Scholar]
- Comino, I.; de Lourdes Moreno, M.; Sousa, C. Role of Oats in Celiac Disease. World J. Gastroenterol. 2015, 21, 11825. [Google Scholar] [CrossRef]
- Sargautiene, V.; Zariņš, Z.; Ligere, R. Effects of the Modulation Gut Microbiota by Oat β-Glucan on Type 2 Diabetes Mellitus. Int. J. Res. Stud. Biosci. 2015, 3, 2349–2357. [Google Scholar]
- Noble, E.E.; Hsu, T.M.; Kanoski, S.E. Gut to Brain Dysbiosis: Mechanisms Linking Western Diet Consumption, the Microbiome, and Cognitive Impairment. Front. Behav. Neurosci. 2017, 11, 9. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Song, R.; Li, Y.; Zhang, W.; Wan, Z.; Wang, Y.; Zhang, H.; Han, S. Effects of Oat Fiber Intervention on Cognitive Behavior in LDLR–/–Mice Modeling Atherosclerosis by Targeting the Microbiome–Gut–Brain Axis. J. Agric. Food Chem. 2020, 68, 14480–14491. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Peñas, E. Health Benefits of Oat: Current Evidence and Molecular Mechanisms. Curr. Opin. Food Sci. 2017, 14, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M. Cereal β-glucans: An underutilized health endorsing food ingredient. Crit. Rev. Food Sci. Nutr. 2022, 62, 3281–3300. [Google Scholar] [CrossRef] [PubMed]
- Izydorczyk, M.S. Functional Properties of Cereal Cell Wall Polysaccharides. In Carbohydrates in Food; CRC Press: Boca Raton, FL, USA, 2017; pp. 215–278. [Google Scholar]
- Lazaridou, A.; Biliaderis, C.G. Molecular Aspects of Cereal β-Glucan Functionality: Physical Properties, Technological Applications and Physiological Effects. J. Cereal Sci. 2007, 46, 101–118. [Google Scholar] [CrossRef]
- Atanasov, J.; Schloermann, W.; Trautvetter, U.; Glei, M. The Effects of β-Glucans on Intestinal Health. Ernahr. Umsch. 2020, 67, 52–59. [Google Scholar]
- Mathews, R.; Kamil, A.; Chu, Y. Global Review of Heart Health Claims for Oat β-Glucan Products. Nutr. Rev. 2020, 78, 78–97. [Google Scholar] [CrossRef] [PubMed]
- DJukić, N.H.; Knežević, D.S. Molecular Characterization and Genetic Diversity Analysis β-Glucan Content Variability in Grain of Oat (Avena sativa L.). Genetika 2014, 46, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Ciecierska, A.; Drywien, M.; Hamulka, J.; Sadkowski, T. Nutraceutical Functions of β-Glucans in Human Nutrition. Roczniki Państwowego Zakl. Hig. 2019, 70, 315–324. [Google Scholar]
- Wang, Q.; Sheng, X.; Shi, A.; Hu, H.; Yang, Y.; Liu, L.; Fei, L.; Liu, H. β-Glucans: Relationships between Modification, Conformation and Functional Activities. Molecules 2017, 22, 257. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Sharma, M.; Ji, D.; Xu, M.; Agyei, D. Structural Features, Modification, and Functionalities of β-Glucan. Fibers 2019, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Suchecka, D.; Gromadzka-Ostrowska, J.; Żyla, E.; Harasym, J.; Oczkowski, M. Selected Physiological Activities and Health Promoting Properties of Cereal β-Glucans. A Review. J. Anim. Feed Sci. 2017, 26, 183–191. [Google Scholar] [CrossRef]
- Bhosale, S.; Singru, S.; Khismatrao, D. Oats: Prospects and Challenges in India. US Nat. Libr. Med. Enlist. J. 2022, 8, 15–155. [Google Scholar]
- El Khoury, D.; Cuda, C.; Luhovyy, B.L.; Anderson, G.H. Beta glucan: Health benefits in obesity and metabolic syndrome. J. Nutr. Metab. 2012, 2012, 851362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacic, A.; Fincher, G.B.; Stone, B. (Eds.) Chemistry, Biochemistry, and Biology of 1-3 Beta Glucans and Related Polysaccharides; Academic Press: Cambridge, MA, USA, 2009; pp. 1–350. [Google Scholar]
- Pérez-Quirce, S.; Lazaridou, A.; Biliaderis, C.G.; Ronda, F. Effect of β-Glucan Molecular Weight on Rice Flour Dough Rheology, Quality Parameters of Breads and in Vitro Starch Digestibility. LWT—Food Sci. Technol. 2017, 82, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Zielke, C.; Lu, Y.; Nilsson, L. Aggregation and Microstructure of Cereal β-Glucan and Its Association with Other Biomolecules. Colloids Surf. A Physicochem. Eng. Asp. 2019, 560, 402–409. [Google Scholar] [CrossRef]
- Rose, D.J. Impact of Whole Grains on the Gut Microbiota: The next Frontier for Oats? Br. J. Nutr. 2014, 112, S44–S49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebello, C.J.; Johnson, W.D.; Martin, C.K.; Han, H.; Chu, Y.-F.; Bordenave, N.; van Klinken, B.J.W.; O’Shea, M.; Greenway, F.L. Instant Oatmeal Increases Satiety and Reduces Energy Intake Compared to a Ready-to-Eat Oat-Based Breakfast Cereal: A Randomized Crossover Trial. J. Am. Coll. Nutr. 2016, 35, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Karp, S.; Wyrwisz, J.; Kurek, M.A. The Impact of Different Levels of Oat β-Glucan and Water on Gluten-Free Cake Rheology and Physicochemical Characterisation. J. Food Sci. Technol. 2020, 57, 3628–3638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brummer, Y.; Defelice, C.; Wu, Y.; Kwong, M.; Wood, P.J.; Tosh, S.M. Textural and Rheological Properties of Oat Beta-Glucan Gels with Varying Molecular Weight Composition. J. Agric. Food Chem. 2014, 62, 3160–3167. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.A.; Davis, J.M.; Carmichael, M.D. Immune Modulating Effects of β-Glucan. Curr. Opin. Clin. Nutr. Metabol. Care 2010, 13, 656–661. [Google Scholar] [CrossRef]
- Rieder, A.; Samuelsen, A.B. Do Cereal Mixed-Linked β-Glucans Possess Immune-Modulating Activities? Mol. Nutr. Food Res. 2012, 56, 536–547. [Google Scholar] [CrossRef]
- Kremmyda, A.; MacNaughtan, W.; Arapoglou, D.; Eliopoulos, C.; Metafa, M.; Harding, S.E.; Israilides, C. The detection, purity and structural properties of partially soluble mushroom and cereal β-d-glucans: A solid-state NMR study. Carbohydr. Polym. 2021, 266, 118103. [Google Scholar] [CrossRef]
- Novák, M.; Větvička, V. β-Glucans, History, and the Present: Immunomodulatory Aspects and Mechanisms of Action. J. Immunotoxicol. 2008, 5, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, H.W. Innate Immunity Induced by Fungal β-Glucans via Dectin-1 Signaling Pathway. Int. J. Med. Mushrooms 2014, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Choromanska, A.; Lubinska, S.; Szewczyk, A.; Saczko, J.; Kulbacka, J. Mechanisms of Antimelanoma Effect of Oat β-Glucan Supported by Electroporation. Bioelectrochemistry 2018, 123, 255–259. [Google Scholar] [CrossRef]
- Choromanska, A.; Kulbacka, J.; Rembialkowska, N.; Pilat, J.; Oledzki, R.; Harasym, J.; Saczko, J. Anticancer Properties of Low Molecular Weight Oat Beta-Glucan—An in Vitro Study. Int. J. Biol. Macromol. 2015, 80, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, M.S.; Jespersen, B.M.; Mehlsen, A.; Engelsen, S.B.; Frøkiær, H. Cereal β-Glucan Immune Modulating Activity Depends on the Polymer Fine Structure. Food Res. Int. 2014, 62, 829–836. [Google Scholar] [CrossRef]
- Yun, C.-H.; Estrada, A.; Van Kessel, A.; Park, B.-C.; Laarveld, B. β-Glucan, Extracted from Oat, Enhances Disease Resistance against Bacterial and Parasitic Infections. FEMS Immunol. Med. Microbiol. 2003, 35, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.A.; Davis, J.M.; Carmichael, M.D.; Mayer, E.P.; Ghaffar, A. Benefits of Oat β-Glucan and Sucrose Feedings on Infection and Macrophage Antiviral Resistance Following Exercise Stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R1188–R1194. [Google Scholar] [CrossRef] [PubMed]
- Cheung, N.-K.V.; Modak, S.; Vickers, A.; Knuckles, B. Orally Administered β-Glucans Enhance Anti-Tumor Effects of Monoclonal Antibodies. Cancer Immunol. Immunother. 2002, 51, 557–564. [Google Scholar] [CrossRef]
- Gamel, T.H.; Abdel-Aal, E.-S.M.; Ames, N.P.; Duss, R.; Tosh, S.M. Enzymatic Extraction of β-Glucan from Oat Bran Cereals and Oat Crackers and Optimization of Viscosity Measurement. J. Cereal Sci. 2014, 59, 33–40. [Google Scholar] [CrossRef]
- Wani, S.M.; Gani, A.; Mir, S.A.; Masoodi, F.A.; Khanday, F.A. β-Glucan: A Dual Regulator of Apoptosis and Cell Proliferation. Int. J. Biol. Macromol. 2021, 182, 1229–1237. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Zhao, J.; Wang, J.; Song, Q.; Zhao, C. The Phagocytic Receptors of β-Glucan. Int. J. Biol. Macromol. 2022, 205, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Luo, H.; Ye, Y.; Chen, X.; Zou, Y.; Duan, J.; Xiang, D. Β-glucan, a Dectin-1 Ligand, Promotes Macrophage M1 Polarization via NF-κB/Autophagy Pathway. Int. J. Oncol. 2019, 54, 271–282. [Google Scholar] [PubMed]
- Sahasrabudhe, N.M.; Tian, L.; van den Berg, M.; Bruggeman, G.; Bruininx, E.; Schols, H.A.; Faas, M.M.; de Vos, P. Endo-Glucanase Digestion of Oat β-Glucan Enhances Dectin-1 Activation in Human Dendritic Cells. J. Funct. Foods 2016, 21, 104–112. [Google Scholar] [CrossRef]
- Shoukat, M.; Sorrentino, A. Cereal β-Glucan: A Promising Prebiotic Polysaccharide and Its Impact on the Gut Health. Int. J. Food Sci. 2021, 56, 2088–2097. [Google Scholar] [CrossRef]
- Maheshwari, G.; Sowrirajan, S.; Joseph, B. Extraction and Isolation of β-Glucan from Grain Sources—A Review. J. Food Sci. 2017, 82, 1535–1545. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Lahnstein, J.; James, A.P.; Fenton, H.K.; Burton, R.A.; Cato, L.; Solah, V.A. Effect of Processing on Viscosity and Molecular Weight of (1,3)(1,4)-β-Glucan in Western Australian Oat Cultivars. Cereal Chem. 2017, 94, 625–632. [Google Scholar] [CrossRef]
- Dotsenko, G.; Andersson, A.A.M.; Andersson, R. Material Disintegration Affects Enzymatic Determination of β-Glucan in Barley and Oats. J. Cereal Sci. 2019, 88, 138–144. [Google Scholar] [CrossRef]
- Decker, E.A.; Rose, D.J.; Stewart, D. Processing of Oats and the Impact of Processing Operations on Nutrition and Health Benefits. Br. J. Nutr. 2014, 112, S58–S64. [Google Scholar] [CrossRef] [Green Version]
- de Souza, N.L.; Bartz, J.; da Rosa Zavareze, E.; de Oliveira, P.D.; da Silva, W.S.V.; Alves, G.H.; Dias, A.R.G. Functional, Thermal and Rheological Properties of Oat β-Glucan Modified by Acetylation. Food Chem. 2015, 178, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Andersson, A.A.M.; Börjesdotter, D. Effects of Environment and Variety on Content and Molecular Weight of β-Glucan in Oats. J. Cereal Sci. 2011, 54, 122–128. [Google Scholar] [CrossRef]
- Ekström, L.M.N.K.; Henningsson Bok, E.A.E.; Sjöö, M.E.; Östman, E.M. Oat β-Glucan Containing Bread Increases the Glycaemic Profile. J. Funct. Foods 2017, 32, 106–111. [Google Scholar] [CrossRef]
- Mejía, S.M.V.; de Francisco, A.; Bohrer, B.M. A Comprehensive Review on Cereal β-Glucan: Extraction, Characterization, Causes of Degradation, and Food Application. Crit. Rev. Food Sci. Nutr. 2020, 60, 3693–3704. [Google Scholar] [CrossRef] [PubMed]
- Gamel, T.H.; Abdel-Aal, E.-S.M.; Tosh, S.M. Effect of Yeast-Fermented and Sour-Dough Making Processes on Physicochemical Characteristics of β-Glucan in Whole Wheat/Oat Bread. LWT—Food Sci. Technol. 2015, 60, 78–85. [Google Scholar] [CrossRef]
- Wang, X.; Storsley, J.; Thandapilly, S.J.; Ames, N. Effects of Processing, Cultivar, and Environment on the Physicochemical Properties of Oat β-Glucan. Cereal Chem. 2016, 93, 402–408. [Google Scholar] [CrossRef]
- Roubroeks, J.P.; Andersson, R.; Mastromauro, D.I.; Christensen, B.E.; Åman, P. Molecular Weight, Structure and Shape of Oat (1→3),(1→4)-β-d-Glucan Fractions Obtained by Enzymatic Degradation with (1→4)-β-d-Glucan 4-Glucanohydrolase from Trichoderma reesei. Carbohydr. Polym. 2001, 46, 275–285. [Google Scholar] [CrossRef]
- Bae, I.Y.; Lee, S.; Kim, S.M.; Lee, H.G. Effect of Partially Hydrolyzed Oat β-Glucan on the Weight Gain and Lipid Profile of Mice. Food Hydrocoll. 2009, 23, 2016–2021. [Google Scholar] [CrossRef]
- Immerstrand, T.; Andersson, K.; Wange, C.; Rascon, A.; Hellstrand, P.; Nyman, M.; Cui, S.W.; Bergenståhl, B.; Trägårdh, C.; Öste, R. Effects of oat bran, processed to different molecular weights of β-glucan, on plasma lipids and caecal formation of SCFA in mice. Brit. J. Nutr. 2010, 104, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Pradeep, N.S.; Edison, L.K. (Eds.) Microbial Beta Glucanases: Molecular Structure, Functions and Applications; Springer Nature: Berlin/Heidelberg, Germany, 2022; pp. 1–195. [Google Scholar]
- Chaari, F.; Belghith-Fendri, L.; Zaouri-Ellouzi, S.; Driss, D.; Blibech, M.; Kallel, F.; Bouaziz, F.; Mehdi, Y.; Ellouz-Chaabouni, S.; Ghorbel, R. Antibacterial and antioxidant properties of mixed linkage β-oligosaccharides from extracted β-glucan hydrolysed by Penicillium occitanis EGL lichenase. Nat. Product Res. 2016, 30, 1353–1359. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.-U.; Ko, M.-J.; Chung, M.-S. Hydrolysis of Beta-Glucan in Oat Flour during Subcritical-Water Extraction. Food Chem. 2020, 308, 125670. [Google Scholar] [CrossRef]
- Harasym, J.; Suchecka, D.; Gromadzka-Ostrowska, J. Effect of Size Reduction by Freeze-Milling on Processing Properties of Beta-Glucan Oat Bran. J. Cereal Sci. 2015, 61, 119–125. [Google Scholar] [CrossRef]
- Rosa-Sibakov, N.; de Oliveira Carvalho, M.J.; Lille, M.; Nordlund, E. Impact of Enzymatic Hydrolysis and Microfluidization on the Techno-Functionality of Oat Bran in Suspension and Acid Milk Gel Models. Foods 2022, 11, 228. [Google Scholar] [CrossRef]
- Shah, A.; Masoodi, F.A.; Gani, A.; Ashwar, B.A. Effect of γ-Irradiation on Antioxidant and Antiproliferative Properties of Oat β-Glucan. Radiat. Phys. Chem. 2015, 117, 120–127. [Google Scholar] [CrossRef]
- Hussain, P.R.; Rather, S.A.; Suradkar, P.P. Structural Characterization and Evaluation of Antioxidant, Anticancer and Hypoglycemic Activity of Radiation Degraded Oat (Avena sativa) β- Glucan. Radiat. Phys. Chem. 2018, 144, 218–230. [Google Scholar] [CrossRef]
- Sun, T.; Li, J.; Qin, Y.; Xie, J.; Xue, B.; Li, X.; Gan, J.; Bian, X.; Shao, Z. Rheological and Functional Properties of Oat β-Glucan with Different Molecular Weight. J. Mol. Struct. 2020, 1209, 127944. [Google Scholar] [CrossRef]
- Shin, M.S.; Lee, S.; Lee, K.Y.; Lee, H.G. Structural and Biological Characterization of Aminated-Derivatized Oat β-Glucan. J. Agric. Food Chem. 2005, 53, 5554–5558. [Google Scholar] [CrossRef]
- Song, X.; Hubbe, M.A. Enhancement of Paper Dry Strength by Carboxymethylated β-d-Glucan from Oat as Additive. Holzforschung 2014, 68, 257–263. [Google Scholar] [CrossRef]
- Chang, Y.J.; Lee, S.; Yoo, M.A.; Lee, H.G. Structural and Biological Characterization of Sulfated-Derivatized Oat β-Glucan. J. Agric. Food Chem. 2006, 54, 3815–3818. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Ming, J.; Gao, R.; Wang, Y.; Liang, Q.; Yu, H.; Zhao, G. Characterization and Antioxidant Activity of the Complex of Tea Polyphenols and Oat β-Glucan. J. Agric. Food Chem. 2011, 59, 10737–10746. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-C.; Luo, Y.; Meng, F.-B.; Li, J.; Chen, W.-J.; Liu, D.-Y.; Zou, L.-H.; Zhou, L. Preparation and Characterization of Feruloylated Oat β-Glucan with Antioxidant Activity and Colon-Targeted Delivery. Carbohydr. Polym. 2022, 279, 119002. [Google Scholar] [CrossRef]
- Inglett, G.E.; Chen, D. Antioxidant and Pasting Properties of Oat β-Glucan Hydrocolloids. Food Sci. Nutr. 2012, 3, 827–835. [Google Scholar]
- Hitayezu, R.; Baakdah, M.M.; Kinnin, J.; Henderson, K.; Tsopmo, A. Antioxidant Activity, Avenanthramide and Phenolic Acid Contents of Oat Milling Fractions. J. Cereal Sci. 2015, 63, 35–40. [Google Scholar] [CrossRef]
- Marasca, E.; Boulos, S.; Nyström, L. Bile Acid-Retention by Native and Modified Oat and Barley β-Glucan. Carbohydr. Polym. 2020, 236, 116034. [Google Scholar] [CrossRef]
- Sun, T.; Xu, H.; Zhang, H.; Ding, H.; Cui, S.; Xie, J.; Xue, B.; Hua, X. Maillard Reaction of Oat β-Glucan and the Rheological Property of Its Amino Acid/Peptide Conjugates. Food Hydrocol. 2018, 76, 30–34. [Google Scholar] [CrossRef]
- Żyła, E.; Dziendzikowska, K.; Gajewska, M.; Wilczak, J.; Harasym, J.; Gromadzka-Ostrowska, J. Beneficial Effects of Oat β-Glucan Dietary Supplementation in Colitis Depend on Its Molecular Weight. Molecules 2019, 24, 3591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Żyła, E.; Dziendzikowska, K.; Kamola, D.; Wilczak, J.; Sapierzyński, R.; Harasym, J.; Gromadzka-Ostrowska, J. Anti-Inflammatory Activity of Oat Beta-Glucans in a Crohn’s Disease Model: Time- and Molar Mass-Dependent Effects. Int. J. Mol. Sci. 2021, 22, 4485. [Google Scholar] [CrossRef] [PubMed]
- Kopiasz, Ł.; Dziendzikowska, K.; Gajewska, M.; Wilczak, J.; Harasym, J.; Żyła, E.; Kamola, D.; Oczkowski, M.; Królikowski, T.; Gromadzka-Ostrowska, J. Time-Dependent Indirect Antioxidative Effects of Oat β-Glucans on Peripheral Blood Parameters in the Animal Model of Colon Inflammation. Antioxidants 2020, 9, 375. [Google Scholar] [CrossRef]
- Kopiasz, Ł.; Dziendzikowska, K.; Gajewska, M.; Oczkowski, M.; Majchrzak-Kuligowska, K.; Królikowski, T.; Gromadzka-Ostrowska, J. Effects of Dietary Oat β-Glucans on Colon Apoptosis and Autophagy through TLRs and Dectin-1 Signaling Pathways—Crohn’s Disease Model Study. Nutrients 2021, 13, 321. [Google Scholar] [CrossRef]
- Kopiasz, Ł.; Dziendzikowska, K.; Gromadzka-Ostrowska, J. Colon Expression of Chemokines and Their Receptors Depending on the Stage of Colitis and Oat β-Glucan Dietary Intervention—Crohn’s Disease Model Study. Int. J. Mol. Sci. 2022, 23, 1406. [Google Scholar] [CrossRef]
- Wilczak, J.; Błaszczyk, K.; Kamola, D.; Gajewska, M.; Harasym, J.P.; Jałosińska, M.; Gudej, S.; Suchecka, D.; Oczkowski, M.; Gromadzka-Ostrowska, J. The Effect of Low or High Molecular Weight Oat β-Glucans on the Inflammatory and Oxidative Stress Status in the Colon of Rats with LPS-Induced Enteritis. Food Funct. 2015, 6, 590–603. [Google Scholar] [CrossRef]
- Błaszczyk, K.; Gajewska, M.; Wilczak, J.; Kamola, D.; Majewska, A.; Harasym, J.; Gromadzka-Ostrowska, J. Oral Administration of Oat β-Glucan Preparations of Different Molecular Weight Results in Regulation of Genes Connected with Immune Response in Peripheral Blood of Rats with LPS-Induced Enteritis. Eur. J. Nutr. 2019, 58, 2859–2873. [Google Scholar] [CrossRef] [Green Version]
- Suchecka, D.; Harasym, J.P.; Wilczak, J.; Gajewska, M.; Oczkowski, M.; Gudej, S.; Błaszczyk, K.; Kamola, D.; Filip, R.; Gromadzka-Ostrowska, J. Antioxidative and Anti-Inflammatory Effects of High β-Glucan Concentration Purified Aqueous Extract from Oat in Experimental Model of LPS-Induced Chronic Enteritis. J. Funct. Foods 2015, 14, 244–254. [Google Scholar] [CrossRef]
- Suchecka, D.; Błaszczyk, K.; Harasym, J.; Gudej, S.; Wilczak, J.; Gromadzka-Ostrowska, J. Impact of Purified Oat 1-3,1-4-β-d-Glucan of Different Molecular Weight on Alleviation of Inflammation Parameters during Gastritis. J. Funct. Foods 2017, 28, 11–18. [Google Scholar] [CrossRef]
- Gudej, S.; Filip, R.; Harasym, J.; Wilczak, J.; Dziendzikowska, K.; Oczkowski, M.; Jałosińska, M.; Juszczak, M.; Lange, E.; Gromadzka-Ostrowska, J. Clinical Outcomes after Oat β-Glucans Dietary Treatment in Gastritis Patients. Nutrients 2021, 13, 2791. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.J.A.; Stojanovski, E.; MacDonald-Wicks, L.; Garg, M.L. High Molecular Weight Oat β-Glucan Enhances Lipid-Lowering Effects of Phytosterols. A Randomised Controlled Trial. Clin. Nutr. 2020, 39, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Mäkelä, N.; Brinck, O.; Sontag-Strohm, T. Viscosity of β-Glucan from Oat Products at the Intestinal Phase of the Gastrointestinal Model. Food Hydrocoll. 2020, 100, 105422. [Google Scholar] [CrossRef]
- Mäkelä, N.; Rosa-Sibakov, N.; Wang, Y.-J.; Mattila, O.; Nordlund, E.; Sontag-Strohm, T. Role of β-Glucan Content, Molecular Weight and Phytate in the Bile Acid Binding of Oat β-Glucan. Food Chem. 2021, 358, 129917. [Google Scholar] [CrossRef] [PubMed]
- Bae, I.Y.; Kim, S.M.; Lee, S.; Lee, H.G. Effect of Enzymatic Hydrolysis on Cholesterol-Lowering Activity of Oat β-Glucan. New Biotechnol. 2010, 27, 85–88. [Google Scholar] [CrossRef]
- Yu, J.; Xia, J.; Yang, C.; Pan, D.; Xu, D.; Sun, G.; Xia, H. Effects of Oat β-Glucan Intake on Lipid Profiles in Hypercholesterolemic Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2022, 14, 2043. [Google Scholar] [CrossRef]
- Hakkola, S.; Nylund, L.; Rosa-Sibakov, N.; Yang, B.; Nordlund, E.; Pahikkala, T.; Kalliomäki, M.; Aura, A.-M.; Linderborg, K.M. Effect of Oat β-Glucan of Different Molecular Weights on Fecal Bile Acids, Urine Metabolites and Pressure in the Digestive Tract—A Human Cross over Trial. Food Chem. 2021, 342, 128219. [Google Scholar] [CrossRef]
- Rosa-Sibakov, N.; Mäkelä, N.; Aura, A.-M.; Sontag-Strohm, T.; Nordlund, E. In Vitro Study for Investigating the Impact of Decreasing the Molecular Weight of Oat Bran Dietary Fibre Components on the Behaviour in Small and Large Intestine. Food Funct. 2020, 11, 6680–6691. [Google Scholar] [CrossRef]
- Shen, R.-L.; Wang, Z.; Dong, J.-L.; Xiang, Q.-S.; Liu, Y.-Q. Effects of Oat Soluble and Insoluble β-Glucan on 1,2-Dimethylhydrazine-Induced Early Colon Carcinogenesis in Mice. Food Agric. Immunol. 2016, 27, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Bozbulut, R.; Sanlier, N. Promising Effects of β-Glucans on Glyceamic Control in Diabetes. Trends Food Sci. Technol. 2019, 83, 159–166. [Google Scholar] [CrossRef]
- Rieder, A.; Knutsen, S.H.; Fernandez, A.S.; Ballance, S. At a High Dose Even Partially Degraded β-Glucan with Decreased Solubility Significantly Reduced the Glycaemic Response to Bread. Food Funct. 2019, 10, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, Y.; Zhang, H.; Hu, B.; Wang, L.; Qian, H.; Qi, X. The Anti-Diabetic Activity of Oat β-d-Glucan in Streptozotocin–Nicotinamide Induced Diabetic Mice. Int. J. Biol. Macromol. 2016, 91, 1170–1176. [Google Scholar] [CrossRef]
- Regand, A.; Chowdhury, Z.; Tosh, S.M.; Wolever, T.M.S.; Wood, P. The Molecular Weight, Solubility and Viscosity of Oat β-Glucan Affect Human Glycemic Response by Modifying Starch Digestibility. Food Chem. 2011, 129, 297–304. [Google Scholar] [CrossRef]
- Noronha, J.C.; Zurbau, A.; Wolever, T.M.S. The Importance of Molecular Weight in Determining the Minimum Dose of Oat β-Glucan Required to Reduce the Glycaemic Response in Healthy Subjects without Diabetes: A Systematic Review and Meta-Regression Analysis. Eur. J. Clin. Nutr. 2022. [Google Scholar] [CrossRef]
- Andrade, E.F.; Orlando, D.R. β-Glucans as a Therapeutic Agent: Literature Review. Madridge J. Food Technol. 2018, 3, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Thies, F.; Masson, L.F.; Boffetta, P.; Kris-Etherton, P. Oats and CVD Risk Markers: A Systematic Literature Review. Br. J. Nutr. 2014, 112, S19–S30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaremba, S.M.M.; Gow, I.F.; Drummond, S.; McCluskey, J.T.; Steinert, R.E. Effects of Oat β-Glucan Consumption at Breakfast on Ad Libitum Eating, Appetite, Glycemia, Insulinemia and GLP-1 Concentrations in Healthy Subjects. Appetite 2018, 128, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Wolever, T.M.S.; Mattila, O.; Rosa-Sibakov, N.; Tosh, S.M.; Jenkins, A.L.; Ezatagha, A.; Duss, R.; Steinert, R.E. Effect of Varying Molecular Weight of Oat β-Glucan Taken Just before Eating on Postprandial Glycemic Response in Healthy Humans. Nutrients 2020, 12, 2275. [Google Scholar] [CrossRef] [PubMed]
- Zurbau, A.; Noronha, J.C.; Khan, T.A.; Sievenpiper, J.L.; Wolever, T. The effect of oat β-glucan on postprandial blood glucose and insulin responses: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2021, 75, 1540–1554. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Wang, L.; Qian, H.; Qi, X.; Ding, X.; Hu, B.; Li, J. The Effect of Oat β-Glucan on in Vitro Glucose Diffusion and Glucose Transport in Rat Small Intestine. J. Sci. Food Agric. 2016, 96, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Henrion, M.; Francey, C.; Lê, K.-A.; Lamothe, L. Cereal β-Glucans: The Impact of Processing and How It Affects Physiological Responses. Nutrients 2019, 11, 1729. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Xu, B. Oxygen Radical Absorbance Capacity (ORAC) and Ferric Reducing Antioxidant Power (FRAP) of β-Glucans from Different Sources with Various Molecular Weight. Bioact. Carbohydr. Diet. Fibre 2014, 3, 11–16. [Google Scholar] [CrossRef]
- Błaszczyk, K.; Wilczak, J.; Harasym, J.; Gudej, S.; Suchecka, D.; Królikowski, T.; Lange, E.; Gromadzka-Ostrowska, J. Impact of Low and High Molecular Weight Oat β-Glucan on Oxidative Stress and Antioxidant Defense in Spleen of Rats with LPS Induced Enteritis. Food Hydrocoll. 2015, 51, 272–280. [Google Scholar] [CrossRef]
- Suchecka, D.; Harasym, J.; Wilczak, J.; Gromadzka-Ostrowska, J. Hepato- and Gastro- Protective Activity of Purified Oat 1–3, 1–4-β-d-Glucans of Different Molecular Weight. Int. J. Biol. Macromol. 2016, 91, 1177–1185. [Google Scholar] [CrossRef]
- Staka, A.; Bodnieks, E.; Puķītis, A. Impact of Oat-Based Products on Human Gastrointestinal Tract. Proc. Latv. Acad. Sci. B Nat. Exact Appl. Sci. 2015, 69, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Li, T.; Zhang, W.; Fan, M.; Qian, H.; Li, Y.; Wang, L. Systematic Assessment of Oat β-Glucan Catabolism during in Vitro Digestion and Fermentation. Food Chem. 2021, 348, 129116. [Google Scholar] [CrossRef]
- Dong, J.; Yang, M.; Zhu, Y.; Shen, R.; Zhang, K. Comparative Study of Thermal Processing on the Physicochemical Properties and Prebiotic Effects of the Oat β-Glucan by in Vitro Human Fecal Microbiota Fermentation. Food Res. Int. 2020, 138, 109818. [Google Scholar] [CrossRef]
- Jayachandran, M.; Chen, J.; Chung, S.S.M.; Xu, B. A Critical Review on the Impacts of β-Glucans on Gut Microbiota and Human Health. J. Nutr. Biochem. 2018, 61, 101–110. [Google Scholar] [CrossRef]
- Priyadarshini, M.; Kotlo, K.U.; Dudeja, P.K.; Layden, B.T. Role of Short Chain Fatty Acid Receptors in Intestinal Physiology and Pathophysiology. Compr. Physiol. 2018, 8, 1091–1115. [Google Scholar] [PubMed]
- Dong, J.; Yu, X.; Dong, L.; Shen, R. In Vitro Fermentation of Oat β-Glucan and Hydrolysates by Fecal Microbiota and Selected Probiotic Strains. J. Sci. Food Agric. 2017, 97, 4198–4203. [Google Scholar] [CrossRef]
- Lazaridou, A.; Serafeimidou, A.; Biliaderis, C.G.; Moschakis, T.; Tzanetakis, N. Structure Development and Acidification Kinetics in Fermented Milk Containing Oat β-Glucan, a Yogurt Culture and a Probiotic Strain. Food Hydrocoll. 2014, 39, 204–214. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Zhang, W.; Fan, M.; Qian, H.; Zhang, H.; Qi, X.; Wang, L. Source of Gut Microbiota Determines Oat β-Glucan Degradation and Short Chain Fatty Acid-Producing Pathway. Food Biosci. 2021, 41, 101010. [Google Scholar] [CrossRef]
- Akkerman, R.; Logtenberg, M.J.; An, R.; Van Den Berg, M.A.; de Haan, B.J.; Faas, M.M.; Zoetendal, E.; de Vos, P.; Schols, H.A. Endo-1,3(4)-β-Glucanase-Treatment of Oat β-Glucan Enhances Fermentability by Infant Fecal Microbiota, Stimulates Dectin-1 Activation and Attenuates Inflammatory Responses in Immature Dendritic Cells. Nutrients 2020, 12, 1660. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Ma, L.; Zhou, T.; Huang, Y.; Zhang, L.; Du, Z.; Yong, K.; Yao, X.; Shen, L.; Yu, S.; et al. Beta-Glucan Alters Gut Microbiota and Plasma Metabolites in Pre-Weaning Dairy Calves. Metabolites 2022, 12, 687. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vetvickova, J. Glucans and Cancer: Comparison of Commercially Available β-Glucans—Part IV. Anticancer Res. 2018, 38, 1327–1333. [Google Scholar]
- Shah, A.; Gani, A.; Masoodi, F.A.; Wani, S.M.; Ashwar, B.A. Structural, Rheological and Nutraceutical Potential of β-Glucan from Barley and Oat. Bioact. Carbohydr. Diet. Fibre 2017, 10, 10–16. [Google Scholar] [CrossRef]
- Baldassano, S.; Accardi, G.; Vasto, S. Beta-Glucans and Cancer: The Influence of Inflammation and Gut Peptide. Eur. J. Med. Chem. 2017, 142, 486–492. [Google Scholar] [CrossRef]
- Parzonko, A.; Makarewicz-Wujec, M.; Jaszewska, E.; Harasym, J.; Kozłowska-Wojciechowska, M. Pro-Apoptotic Properties of (1,3)(1,4)-β-d-Glucan from Avena Sativa on Human Melanoma HTB-140 Cells in Vitro. Int. J. Biol. Macromol. 2015, 72, 757–763. [Google Scholar] [CrossRef]
- Choromanska, A.; Kulbacka, J.; Harasym, J.; Oledzki, R.; Szewczyk, A.; Saczko, J. High- and Low-Molecular Weight Oat Beta-Glucan Reveals Antitumor Activity in Human Epithelial Lung Cancer. Pathol. Oncol. Res. 2018, 24, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Choromańska, A.; Kulbacka, J.; Harasym, J.; Dubińska-Magiera, M.; Saczko, J. Anticancer Activity of Oat β-Glucan in Combination with Electroporation on Human Cancer Cells. Acta Pol. Pharm. 2017, 74, 616–623. [Google Scholar] [PubMed]
- Zhang, M.; Chun, L.; Sandoval, V.; Graor, H.; Myers, J.; Nthale, J.; Rauhe, P.; Senders, Z.; Choong, K.; Huang, A.Y.; et al. Systemic Administration of β-Glucan of 200 KDa Modulates Melanoma Microenvironment and Suppresses Metastatic Cancer. Oncoimmunology 2018, 7, e1387347. [Google Scholar] [CrossRef]
- Brummer, Y.; Duss, R.; Wolever, T.M.; Tosh, S.M. Glycemic Response to Extruded Oat Bran Cereals Processed to Vary in Molecular Weight. Cereal Chem. 2012, 89, 255–261. [Google Scholar] [CrossRef]
- Kwong, M.G.; Wolever, T.M.; Brummer, Y.; Tosh, S.M. Attenuation of Glycemic Responses by Oat β-Glucan Solutions and Viscoelastic Gels is Dependent on Molecular Weight Distribution. Food Funct. 2013, 4, 401–408. [Google Scholar] [CrossRef]
- Mikkilä, L. Oat in Fruit Juice Drink: The Behavior of Oat. Master’s Thesis, Turku University of Applied Sciences, Turku, Finland, 2022. [Google Scholar]
- Naumann, E.; VaN Rees, A.B.; Önning, G.; Öste, R.; Wydra, M.; Mensink, R.P. β-Glucan incorporated into a fruit drink effectively lowers serum LDL-cholesterol concentrations. Am. J. Clin. Nutr. 2006, 83, 601–605. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhao, L.; Zhang, J.; Wang, L.; Liu, H. Functional Drink Powders from Vertical-stone-milled Oat and Highland Barley with High Dietary-fiber Levels Decrease the Postprandial Glycemic Response. J. Funct. Foods 2021, 83, 104548. [Google Scholar] [CrossRef]
- Kurek, M.A.; Moczkowska, M.; Karp, S.; Horbańczuk, O.K.; Rodak, E. Application of Rich in β-Glucan Flours and Preparations in Bread Baked from Frozen Dough. Food Sci. Technol. Int. 2020, 26, 53–64. [Google Scholar]
- Tosh, S.M.; Brummer, Y.; Wolever, T.M.; Wood, P.J. Glycemic Response to Oat Bran Muffins Treated to Vary Molecular Weight of β-Glucan. Cereal Chem. 2008, 85, 211–217. [Google Scholar] [CrossRef]
- Åman, P.; Rimsten, L.; Andersson, R. Molecular Weight Distribution of β-Glucan in Oat-based Foods. Cereal Chem. 2004, 81, 356–360. [Google Scholar] [CrossRef]
- Tosh, S.M.; Brummer, Y.; Miller, S.S.; Regand, A.; Defelice, C.; Duss, R.; Wolever, T.M.; Wood, P.J. Processing Affects the Physicochemical Properties of β-Glucan in Oat Bran Cereal. J. Agric. Food Chem. 2010, 58, 7723–7730. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Inglett, G.E.; Liu, S.X. Content and Molecular Weight Distribution of Oat β-Glucan in Oatrim, Nutrim, and C-trim Products. Cereal Chem. 2008, 85, 701–705. [Google Scholar] [CrossRef]
- Hamaker, B.R. (Ed.) Technology of Functional Cereal Products; Woodhead Publishing Limited: Cambridge, UK, 2007; pp. 1–568. [Google Scholar]

| Structure | Product | Origin | Mw (kDa) | References |
|---|---|---|---|---|
| (1 → 3)-β-d-Glucans | Curdlan | Agrobacterium tumefaciens | 2000 | [14] |
| Paramylon | Euglena gracilis | 500 | [15] | |
| Pachyman | Poria cocos | 89–168 | [16] | |
| (1 → 4)-β-d-Glucans | Plant cellulose | Higher plants, seaweeds | [8] | |
| Animal cellulose | Tunicata | [10] | ||
| Microbial cellulose | Acetobacter xylinum | [9] | ||
| Sarcina ventriculi | ||||
| (1 → 6)-β-d-Glucans | Pustulan | Lasallia pustulata | [17] | |
| Lasiodiplodan | Lasiodiplodia theobromae | [18] | ||
| Mixed-linkage β-d-glucans | Lichenan | Cetraria islandica | [7] | |
| Cereal β-d-glucans | Hordeum vulgare | 1700–2700 | [11] | |
| Avena sativa | 65–3100 | [12] | ||
| Secale cereale | 800–1300 | [12] | ||
| Triticum sp. | 209–487 | [13] | ||
| Branched β-d-glucans | Laminarin | Laminaria digitate Saccharina longicruris Durvillaea antarctica | 3–7 | [5,6] |
| Calocyban | Calocybe indica | 200 | [19] | |
| Botryosphaeran | Botryosphaeria sp. | [20] | ||
| Epiglucan | Epicoccum nigrum | [21] | ||
| Grifolan | Grifola frondosa | [22] | ||
| Krestin | Coriolus versicolor | 10–100 | [23] | |
| Lentinan | Lentinus edodes | 300–800 | [24] | |
| Pestalotan | Pestalotia sp. 815 | [25] | ||
| Pleuran | Pleurotus ostreatus | 600–700 | [26] | |
| Schizophyllan | Schizophyllum commune | 450 | [27] | |
| Scleroglucan | Sclerotium glucanicum | [28] | ||
| Zymosan | Saccharomices cerevisiae | 240 | [29] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sushytskyi, L.; Synytsya, A.; Čopíková, J.; Lukáč, P.; Rajsiglová, L.; Tenti, P.; Vannucci, L.E. Perspectives in the Application of High, Medium, and Low Molecular Weight Oat β-d-Glucans in Dietary Nutrition and Food Technology—A Short Overview. Foods 2023, 12, 1121. https://doi.org/10.3390/foods12061121
Sushytskyi L, Synytsya A, Čopíková J, Lukáč P, Rajsiglová L, Tenti P, Vannucci LE. Perspectives in the Application of High, Medium, and Low Molecular Weight Oat β-d-Glucans in Dietary Nutrition and Food Technology—A Short Overview. Foods. 2023; 12(6):1121. https://doi.org/10.3390/foods12061121
Chicago/Turabian StyleSushytskyi, Leonid, Andriy Synytsya, Jana Čopíková, Pavol Lukáč, Lenka Rajsiglová, Paolo Tenti, and Luca E. Vannucci. 2023. "Perspectives in the Application of High, Medium, and Low Molecular Weight Oat β-d-Glucans in Dietary Nutrition and Food Technology—A Short Overview" Foods 12, no. 6: 1121. https://doi.org/10.3390/foods12061121