
β-Lactamase Producing Escherichia coli Encoding blaCTX-M and blaCMY Genes in Chicken Carcasses from Egypt
 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Isolation and Identification of Escherichia coli
2.3. Molecular Identification of E. coli
2.4. Antimicrobial Susceptibility Testing
2.5. PCR Screening for β-Lactamase Genes
2.6. Phylogenetic Typing of E. coli Isolates
3. Results and Discussion
3.1. Prevalence of E. coli in the Chicken Samples Examined
3.2. Antimicrobial Resistance Profiles of E. coli Isolates
3.3. Determination of β-Lactamase Genes
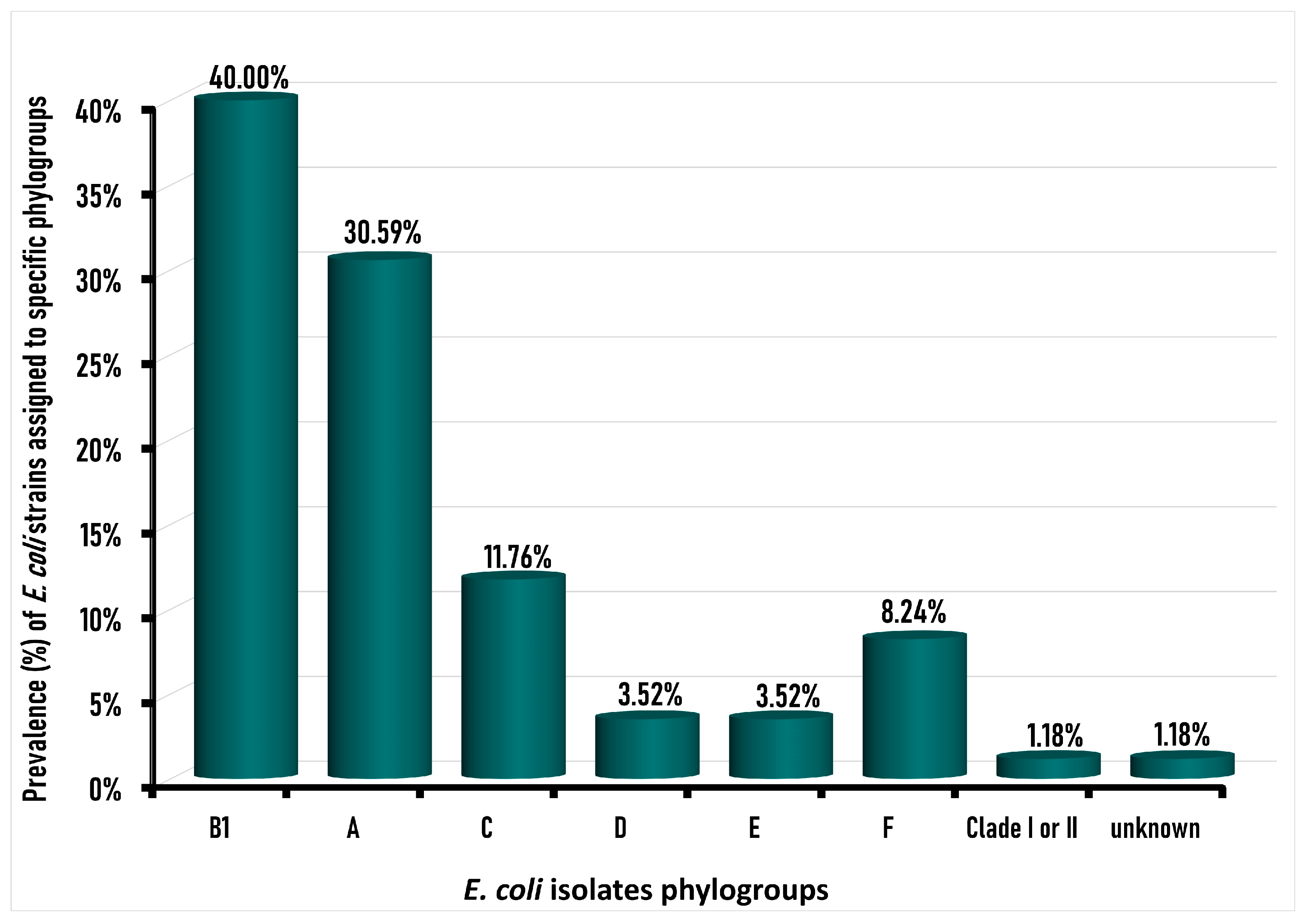
3.4. Phylogroup Characterization of Isolated E. coli Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Adenipekun, E.O.; Jackson, C.R.; Oluwadun, A.; Iwalokun, B.A.; Frye, J.G.; Barrett, J.B.; Hiott, L.M.; Woodley, T.A. Prevalence and Antimicrobial Resistance in Escherichia coli from Food Animals in Lagos, Nigeria. Microb. Drug Resist. 2015, 21, 358–365. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Draft Global Action Plan on Antimicrobial Resistance WHA68.7; World Health Organization: Geneva, Switzerland, 2015; pp. 1–4. Available online: http://apps.who.int/gb/ebwha/pdf_files/WHA68/A68_20-en.pdf?ua¼1 (accessed on 24 November 2022).
- Meena, H.R.; Kumar, V. Antimicrobial Resistance and Rational Use of Antimicrobials in Livestock: Developing Countries’ Perspective. In Livestock Health and Farming; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- von Wintersdorff, C.J.H.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of Antimicrobial Resistance in Microbial Ecosystems through Horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef]
- Hughes, L.A.; Pinchbeck, G.; Callaby, R.; Dawson, S.; Clegg, P.; Williams, N. Antimicrobial prescribing practice in UK equine veterinary practice. Equine Vet. J. 2013, 45, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. β-Lactams and β-lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- CIPARS. Canadian Integrated Program for Antimicrobial Resistance Surveillance. Federal Organisation of CIPARS, Canada. 2006. Available online: https://www.canada.ca/en/public-health/services/surveillance/canadian-integrated-program-antimicrobial-resistance-surveillance-cipars.html (accessed on 24 December 2022).
- Ungureanu, V.; Corcionivoschi, N.; Gundogdu, O.; Stef, L.; Pet, I.; Pacala, N.; Madden, R.H. The emergence of β lactamase producing Escherichia coli and the problems in assessing their potential contribution to foodborne illness: A review. AgroLife Sci. J. 2019, 8, 248–260. [Google Scholar]
- Imre, K.; Ban-Cucerzan, A.; Herman, V.; Sallam, K.I.; Cristina, R.T.; Abd-Elghany, S.M.; Morar, D.; Popa, S.A.; Imre, M.; Morar, A. Occurrence, Pathogenic Potential and Antimicrobial Resistance of Escherichia coli Isolated from Raw Milk Cheese Commercialized in Banat Region, Romania. Antibiotics 2022, 11, 721. [Google Scholar] [CrossRef] [PubMed]
- Essack, S.Y. The Development of β-Lactam Antibiotics in Response to the Evolution of β-Lactamases. Pharm. Res. 2001, 18, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M.; Canton, R.; Gniadkowski, M.; Nordmann, P.; Rossolini, G.M.; Arlet, G.; Ayala, J.; Coque, T.M.; Kern-Zdanowicz, I.; Luzzaro, F.; et al. CTX-M: Changing the face of ESBLs in Europe. J. Antimicrob. Chemother. 2007, 59, 165–174. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated Functional Classification of β-Lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Braun, S.D.; Ahmed, M.F.E.; El-Adawy, H.; Hotzel, H.; Engelmann, I.; Weiß, D.; Monecke, S.; Ehricht, R. Surveillance of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in Dairy Cattle Farms in the Nile Delta, Egypt. Front. Microbiol. 2016, 7, 1020. [Google Scholar] [CrossRef] [PubMed]
- ElBaradei, A.; Maharem, D.A.; Kader, O.; Ghareeb, M.K.; Naga, I.S. Fecal carriage of ESBL-producing Escherichia coli in Egyptian patients admitted to the Medical Research Institute hospital, Alexandria University. AIMS Microbiol. 2020, 6, 422–433. [Google Scholar] [CrossRef]
- Panisello, P.J.; Rooney, R.; Quantick, P.C.; Stanwell-Smith, R. Application of foodborne disease outbreak data in the development and maintenance of HACCP systems. Int. J. Food Microbiol. 2000, 59, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; McCabe, J.S.; White, D.G.; Johnston, B.; Kuskowski, M.A.; McDermott, P. Molecular Analysis of Escherichia coli from Retail Meats (2002–2004) from the United States National Antimicrobial Resistance Monitoring System. Clin. Infect. Dis. 2009, 49, 195–201. [Google Scholar] [CrossRef]
- Martin, A.; Beutin, L. Characteristics of Shiga toxin-producing Escherichia coli from meat and milk products of different origins and association with food producing animals as main contamination sources. Int. J. Food Microbiol. 2011, 146, 99–104. [Google Scholar] [CrossRef]
- Adeyanju, G.T.; Ishola, O. Salmonella and Escherichia coli contamination of poultry meat from a processing plant and retail markets in Ibadan, Oyo State, Nigeria. Springerplus 2014, 3, 139. [Google Scholar] [CrossRef] [PubMed]
- Pavlickova, S.; Dolezalova, M.; Holko, I. Resistance and virulence factors of Escherichia coli isolated from chicken. J. Environ. Sci. Heal. Part B 2015, 50, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wang, Y.; Shi, X.; Wang, S.; Ren, H.; Shen, Z.; Wang, Y.; Lin, J.; Wang, S. Rapid rise of the ESBL and mcr-1 genes in Escherichia coli of chicken origin in China, 2008–2014. Emerg. Microbes Infect. 2018, 7, 30. [Google Scholar] [CrossRef]
- Pfeifer, Y.; Cullik, A.; Witte, W. Resistance to cephalosporins and carbapenems in Gram-negative bacterial pathogens. Int. J. Med. Microbiol. 2010, 300, 371–379. [Google Scholar] [CrossRef]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Herman, L.; Haesebrouck, F.; Butaye, P. Broad-spectrum β-lactamases among Enterobacteriaceae of animal origin: Molecular aspects, mobility and impact on public health. FEMS Microbiol. Rev. 2010, 34, 295–316. [Google Scholar] [CrossRef] [Green Version]
- Schjørring, S.; Krogfelt, K.A. Assessment of Bacterial Antibiotic Resistance Transfer in the Gut. Int. J. Microbiol. 2011, 2011, 312956. [Google Scholar] [CrossRef]
- Moawad, A.A.; Hotzel, H.; Awad, O.; Tomaso, H.; Neubauer, H.; Hafez, H.M.; El-Adawy, H. Occurrence of Salmonella enterica and Escherichia coli in raw chicken and beef meat in northern Egypt and dissemination of their antibiotic resistance markers. Gut Pathog. 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, H.H.; Jackson, C.R.; Taha, S.A.; Moawad, A.A.; Barrett, J.B.; Woodley, T.A. Contribution of Healthy Chickens to Antimicrobial-Resistant Escherichia coli Associated with Human Extraintestinal Infections in Egypt. Vector-Borne Zoonotic Dis. 2018, 18, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Bonkoungou, I.J.O.; Lienemann, T.; Martikainen, O.; Dembelé, R.; Sanou, I.; Traoré, A.S.; Siitonen, A.; Barro, N.; Haukka, K. Diarrhoeagenic Escherichia coli detected by 16-plex PCR in children with and without diarrhoea in Burkina Faso. Clin. Microbiol. Infect. 2012, 18, 901–906. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Clinical and Laboratory Standards Institute. In Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; pp. 1–294. [Google Scholar]
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Routine and Extended Internal Quality Control for MIC Determination and Disk Diffusion as Recommended by EUCAST. Version 8.0. 2018. Available online: http://www.eucast.org. (accessed on 25 November 2022).
- Ahmed, A.M.; Furuta, K.; Shimomura, K.; Kasama, Y.; Shimamoto, T. Genetic characterization of multidrug resistance in Shigella spp. from Japan. J. Med. Microbiol. 2006, 55, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Colom, K.; PãRez, J.; Alonso, R.; Fernã¡ndez-Aranguiz, A.; LariãO, E.; Cisterna, R.; Pérez, J.; Cisterna, R. Simple and reliable multiplex PCR assay for detection of blaTEM, blaSHV and blaOXA-1 genes in Enterobacteriaceae. FEMS Microbiol. Lett. 2003, 223, 147–151. [Google Scholar] [CrossRef]
- Ouellette, M.; Bissonnette, L.; Roy, P.H. Precise insertion of antibiotic resistance determinants into Tn21-like transposons: Nucleotide sequence of the OXA-1. Proc. Natl. Acad. Sci. USA 1987, 84, 7378–7382. [Google Scholar] [CrossRef]
- Zhao, S.; Qaiyumi, S.; Friedman, S.; Singh, R.; Foley, S.L.; White, D.G.; McDermott, P.F.; Donkar, T.; Bolin, C.; Munro, S.; et al. Characterization of Salmonella enterica Serotype Newport Isolated from Humans and Food Animals. J. Clin. Microbiol. 2003, 41, 5366–5371. [Google Scholar] [CrossRef]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Lescat, M.; Clermont, O.; Woerther, P.L.; Glodt, J.; Dion, S.; Skurnik, D.; Djossou, F.; Dupont, C.; Perroz, G.; Picard, B.; et al. Commensal Escherichia coli strains in Guiana reveal a high genetic diversity with host-dependant population structure. Environ. Microbiol. Rep. 2012, 5, 49–57. [Google Scholar] [CrossRef]
- Zweifel, C.; Althaus, D.; Stephan, R. Effects of slaughter operations on the microbiological contamination of broiler carcasses in three abattoirs. Food Control. 2015, 51, 37–42. [Google Scholar] [CrossRef]
- El Tawab, A.A.A.; Ammar, A.M.; Nasef, S.A.; Reda, R.M. Prevalence of E. coli in diseased chickens with its antibiogram pattern. Benha Vet. Med. J. 2015, 28, 224–230. [Google Scholar] [CrossRef]
- Abd El Aziz, E.E.; Hassan, A.M.; Badr, J.M. In vitro efficacy of some antimicrobials on the E. coli and Mycoplasma isolates from cases of chronic respiratory disease in broilers in Egypt. Zag. Vet. J. 2007, 35, 40–49. [Google Scholar]
- Eltai, N.O.; Yassine, H.M.; El-Obeid, T.; Al-Hadidi, S.H.; Al Thani, A.A.; Alali, W.Q. Prevalence of Antibiotic-Resistant Escherichia coli Isolates from Local and Imported Retail Chicken Carcasses. J. Food Prot. 2020, 83, 2200–2208. [Google Scholar] [CrossRef] [PubMed]
- Eyý, A.; Arslan, S. Prevalence of Escherichia coli in retail poultry meat, ground beef, and beef. Med. Weter 2012, 68, 238. [Google Scholar]
- Hossain, M.T.; Siddique, M.P.; Hossain, F.M.A.; Zinnah, M.A.; Hossain, M.M.; Alam, M.K.; Rahman, M.T.; Choudury, K.A. Isolation, identification, toxin profile and antibiogram of Escherichia coli isolated from broilers and layers in Mymensingh district of Bangladesh. Bangladesh J. Vet. Med. 2008, 6, 1–5. [Google Scholar] [CrossRef]
- Samy, A.A.; Mansour, A.S.; Khalaf, D.D.; Khairy, E.A. Development of multidrug-resistant Escherichia coli in some Egyptian veterinary farms. Vet. World 2022, 15, 488–495. [Google Scholar] [CrossRef]
- Gwida, M.; El-Gohary, A. Prevalence and Characterization of Antibiotic Resistance Food Borne Pathogens Isolated from Locally Produced Chicken Raw Meat and their Handlers. J. Dairy Vet. Anim. Res. 2015, 2, 238–244. [Google Scholar] [CrossRef]
- Jana, A.; Mondal, A. Serotyping, pathogenicity and antibiogram of Escherichia coli isolated from raw poultry meat in West Bengal, India. Vet. Ital. 2013, 49, 361–365. [Google Scholar] [CrossRef]
- Zhao, C.; Ge, B.; De Villena, J.; Sudler, R.; Yeh, E.; Zhao, S.; White, D.G.; Wagner, D.; Meng, J. Prevalence of Campylobacter spp., Escherichia coli, and Salmonella Serovars in Retail Chicken, Turkey, Pork, and Beef from the Greater Washington, D.C., Area. Appl. Environ. Microbiol. 2001, 67, 5431–5436. [Google Scholar] [CrossRef]
- Bhattacharjee, P.S.; Kundu, R.L.; Mazumder, J.U.; Hossain, E.; Miah, A.H. A retrospective analysis of chicken diseases diagnosed at the Central Disease Investigation Laboratory, Dhaka, Bangladesh. Bangladesh Vet. Jr. 1996, 30, 105–113. [Google Scholar]
- Ibrahim, W.A.; Marouf, S.A.; Erfan, A.M.; Nasef, S.A.; El Jakee, J.K. The occurrence of disinfectant and antibiotic-resistant genes in Escherichia coli isolated from chickens in Egypt. Vet. World 2019, 12, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Nhung, N.T.; Chansiripornchai, N.; Carrique-Mas, J.J. Antimicrobial Resistance in Bacterial Poultry Pathogens: A Review. Front. Vet. Sci. 2017, 4, 126. [Google Scholar] [CrossRef]
- Hao, H.; Cheng, G.; Iqbal, Z.; Ai, X.; Hussain, H.I.; Huang, L.; Dai, M.; Wang, Y.; Liu, Z.; Yuan, Z.-H. Benefits and risks of antimicrobial use in food-producing animals. Front. Microbiol. 2014, 5, 288. [Google Scholar] [CrossRef]
- Rasmussen, M.M.; Opintan, J.A.; Frimodt-Møller, N.; Styrishave, B. Beta-Lactamase Producing Escherichia coli Isolates in Imported and Locally Produced Chicken Meat from Ghana. PLoS ONE 2015, 10, e0139706. [Google Scholar] [CrossRef]
- Ramadan, H.; Jackson, C.R.; Frye, J.G.; Hiott, L.M.; Samir, M.; Awad, A.; Woodley, T.A. Antimicrobial Resistance, Genetic Diversity and Multilocus Sequence Typing of Escherichia coli from Humans, Retail Chicken and Ground Beef in Egypt. Pathogens 2020, 9, 357. [Google Scholar] [CrossRef]
- Yassin, A.K.; Gong, J.; Kelly, P.; Lu, G.; Guardabassi, L.; Wei, L.; Han, X.; Qiu, H.; Price, S.; Cheng, D.; et al. Antimicrobial resistance in clinical Escherichia coli isolates from poultry and livestock, China. PLoS ONE 2017, 12, e0185326. [Google Scholar] [CrossRef]
- Schwarz, S.; Chaslus-Dancla, E. Use of antimicrobials in veterinary medicine and mechanisms of resistance. Vet. Res. 2001, 32, 201–225. [Google Scholar] [CrossRef]
- Bogaard, A.E.V.D.; London, N.; Driessen, C.; Stobberingh, E.E. Antibiotic resistance of faecal Escherichia coli in poultry, poultry farmers and poultry slaughterers. J. Antimicrob. Chemother. 2001, 47, 763–771. [Google Scholar] [CrossRef]
- Barton, M.D. Impact of antibiotic use in the swine industry. Curr. Opin. Microbiol. 2014, 19, 9–15. [Google Scholar] [CrossRef]
- Van den Bogaard, A.E.; Stobberingh, E.E. Antibiotic usage in animals: Impact on bacterial resistance and public health. Drugs 1999, 58, 589–607. [Google Scholar] [CrossRef] [PubMed]
- Hoelzer, K.; Wong, N.; Thomas, J.; Talkington, K.; Jungman, E.; Coukell, A. Antimicrobial drug use in food-producing animals and associated human health risks: What, and how strong, is the evidence? BMC Vet. Res. 2017, 13, 211. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Imirzalioglu, C.; Oppong, K.; Akenten, C.W.; Hogan, B.; Krumkamp, R.; Poppert, S.; Levermann, V.; Schwengers, O.; Sarpong, N.; et al. Detection and Characterization of ESBL-Producing Escherichia coli From Humans and Poultry in Ghana. Front. Microbiol. 2019, 9, 3358. [Google Scholar] [CrossRef] [PubMed]
- Reich, F.; Atanassova, V.; Klein, G. Extended-Spectrum β-Lactamase– and AmpC-Producing Enterobacteria in Healthy Broiler Chickens, Germany. Emerg. Infect. Dis. 2013, 19, 1253–1259. [Google Scholar] [CrossRef]
- Chishimba, K.; Hang’ombe, B.M.; Muzandu, K.; Mshana, S.E.; Matee, M.I.; Nakajima, C.; Suzuki, Y. Detection of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in Market-Ready Chickens in Zambia. Int. J. Microbiol. 2016, 2016, 5275724. [Google Scholar] [CrossRef]
- Shoaib, M.; Kamboh, A.A.; Sajid, A.; Mughal, G.A.; Leghari, R.A.; Malhi, K.K.; Bughio, S.U.D.; Ali, A.; Alam, S.; Khan, S.; et al. Prevalence of Extended Spectrum Beta-Lactamase Producing Enterobacteriaceae in Commercial Broilers and Backyard Chickens. Adv. Anim. Vet. Sci. 2016, 4, 209–214. [Google Scholar] [CrossRef]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum β-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef]
- Guerra, B.; Junker, E.; Schroeter, A.; Malorny, B.; Lehmann, S.; Helmuth, R. Phenotypic and genotypic characterization of antimicrobial resistance in German Escherichia coli isolates from cattle, swine and poultry. J. Antimicrob. Chemother. 2003, 52, 489–492. [Google Scholar] [CrossRef]
- El Tawab, A.A.A.; El Aal, S.A.A.; Mazied, E.M.; Morsy, D.A.E. Prevalence of E. coli in broiler chickens in winter and summer seasons by application of PCR with its antibiogram pattern. Benha Vet. Med. J. 2015, 29, 119–128. [Google Scholar] [CrossRef]
- Overdevest, I.; Willemsen, I.; Rijnsburger, M.; Eustace, A.; Xu, L.; Hawkey, P.; Heck, M.; Savelkoul, P.; Vandenbroucke-Grauls, C.; van der Zwaluw, K.; et al. Extended-Spectrum B-Lactamase Genes of Escherichia coli in Chicken Meat and Humans, the Netherlands. Emerg. Infect. Dis. 2011, 17, 1216–1222. [Google Scholar] [CrossRef]
- Sabala, R.F.; Usui, M.; Tamura, Y.; Abd-Elghany, S.M.; Sallam, K.I.; Elgazzar, M.M. Prevalence of colistin-resistant Escherichia coli harbouring mcr-1 in raw beef and ready-to-eat beef products in Egypt. Food Control. 2020, 119, 107436. [Google Scholar] [CrossRef]
- Müller, D.; Greune, L.; Heusipp, G.; Karch, H.; Fruth, A.; Tschäpe, H.; Schmidt, M.A. Identification of unconventional intestinal pathogenic Escherichia coli isolates expressing intermediate virulence factor profiles by using a novel single-step multiplex PCR. Appl. Environ. Microbiol. 2007, 73, 3380–3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Shazly, D.A.; Nasef, S.A.; Mahmoud, F.F.; Jonas, D. Expanded spectrum β–lactamase producing Escherichia coli isolated from chickens with colibacillosis in Egypt. Poult. Sci. 2017, 96, 2375–2384. [Google Scholar] [CrossRef] [PubMed]
- Helmy, M.M.; Wasfi, R. Phenotypic and Molecular Characterization of Plasmid Mediated AmpCβ-Lactamases among Escherichia coli, Klebsiella spp., and Proteus mirabilis Isolated from Urinary Tract Infections in Egyptian Hospitals. BioMed. Res. Int. 2014, 2014, 171548. [Google Scholar] [CrossRef] [PubMed]
- Cantón, R.; González-Alba, J.M.; Galán, J.C. CTX-M Enzymes: Origin and Diffusion. Front. Microbiol. 2012, 3, 110. [Google Scholar] [CrossRef]
- Machado, E.; Cantón, R.; Baquero, F.; Galaán, J.-C.; Rollán, A.; Peixe, L.; Coque, T.M. Integron Content of Extended-Spectrum-β-Lactamase-Producing Escherichia coli Strains over 12 Years in a Single Hospital in Madrid, Spain. Antimicrob. Agents Chemother. 2005, 49, 1823–1829. [Google Scholar] [CrossRef]
- Lupo, A.; Saras, E.; Madec, J.-Y.; Haenni, M. Emergence of blaCTX-M-55 associated with fosA, rmtB and mcr gene variants in Escherichia coli from various animal species in France. J. Antimicrob. Chemother. 2018, 73, 867–872. [Google Scholar] [CrossRef]
- Carlos, C.; Pires, M.M.; Stoppe, N.C.; Hachich, E.M.; Sato, M.I.Z.; Gomes, T.A.T.; Amaral, L.A.; Ottoboni, L.M.M. Escherichia coli phylogenetic group determination and its application in the identification of the major animal source of fecal contamination. BMC Microbiol. 2010, 10, 161. [Google Scholar] [CrossRef]
- Coura, F.M.; Diniz, S.A.; Silva, M.X.; Arcebismo, T.L.M.; Minharro, S.; Feitosa, A.C.F.; Lage, A.P.; Knöbl, T.; Mussi, J.M.S.; Heinemann, M.B. Phylogenetic Group of Escherichia coli Isolates from Broilers in Brazilian Poultry Slaughterhouse. Sci. World J. 2017, 2017, 2898701. [Google Scholar] [CrossRef]
- Ghodousi, A.; Bonura, C.; Di Noto, A.M.; Mammina, C. Extended-Spectrum ß-Lactamase, AmpC-Producing, and Fluoroquinolone-Resistant Escherichia coli in Retail Broiler Chicken Meat, Italy. Foodborne Pathog. Dis. 2015, 12, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Rafique, M.; Potter, R.F.; Ferreiro, A.; Wallace, M.A.; Rahim, A.; Malik, A.A.; Siddique, N.; Abbas, M.A.; D’Souza, A.; Burnham, C.-A.D.; et al. Genomic Characterization of Antibiotic Resistant Escherichia coli Isolated From Domestic Chickens in Pakistan. Front. Microbiol. 2020, 10, 3052. [Google Scholar] [CrossRef] [PubMed]
- Egea, P.; López-Cerero, L.; Torres, E.; Gómez-Sánchez, M.D.C.; Serrano, L.; Sánchez-Ortiz, M.D.N.; Rodriguez-Baño, J.; Pascual, A. Increased raw poultry meat colonization by extended spectrum beta-lactamase-producing Escherichia coli in the south of Spain. Int. J. Food Microbiol. 2012, 159, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, X.; Zhou, Z.; Jiang, M.; Wang, Z.; Sun, Y.; Tang, F.; Xue, F.; Ren, J.; Dai, J. Chicken-source Escherichia coli within phylogroup F shares virulence genotypes and is closely related to extraintestinal pathogenic E. coli causing human infections. Transbound. Emerg. Dis. 2020, 68, 880–895. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence | PCR Product Size | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| blaCTX-M | F: 5′-CGCTTTGCGATGTGCAG-3′ R: 5′-ACCGCGATATCGTTGGT-3′ | 550 bp | 55 | [30] |
| blaSHV | F: 5′-AGGATTGACTGCCTTTTTG-3′ R: 5′-ATTTGCTGATTTCGCTCG-3′ | 795 bp | 64 | [31] |
| blaOXA | F: 5′-TATCTACAGCAGCGCCAGTG-3′ R: 5′-CGCATCAAATGCCATAAGTG-3′ | 591 bp | 61 | [32] |
| blaTEM | F: 5′-ATAAAATTCTTGAAGACGAAA-3′ R: 5′-GACAGTTACCAATGCTTAATC-3′ | 1080bp | 51 | [30] |
| blaCMY | F: 5′-GACAGCCTCTTTCTCCACA-3′ R: 5′-TGGAACGAAGGCTACGTA-3′ | 1007 bp | 55 | [33] |
| arpA | F: 5′-AACGCTATTCGCCAGCTTGC-3′ R: 5′-TCTCCCCATACCGTACGCTA-3′ | 400 bp | 59 | [34] |
| chuA | F: 5′-ATGGTACCGGACGAACCAAC-3′ R: 5′-TGCCGCCAGTACCAAAGACA-3′ | 288 bp | 59 | [34] |
| yjaA | F: 5′-CAAACGTGAAGTGTCAGGAG-3′ R: 5′-AATGCGTTCCTCAACCTGTG-3′ | 211 bp | 59 | [34] |
| TspE4C2 | F: 5′-CACTATTCGTAAGGTCATCC-3′ R: 5′-AGTTTATCGCTGCGGGTCGC-3′ | 152 bp | 59 | [34] |
| trpA (Group C) | F: 5′-AGTTTTATGCCCAGTGCGAG-3′ R: 5′-TCTGCGCCGGTCACGCCC-3′ | 219 bp | 59 | [35] |
| arpA (Group E) | F 5′-GATTCCATCTTGTCAAAATATGCC-3′ R 5′GAAAAGAAAAAGAATTCCCAAGAG-3′ | 301 bp | 57 | [35] |
| trpA (Internal control) | F: 5′-CGGCGATAAAGACATCTTCAC-3′ R: 5′-GCAACGCGGCCTGGCGGAAG-3′ | 489 bp | 59 | [34] |
| Isolate Number | blaSHV | blaCTX-M | blaCMY | blaTEM | blaOXA | Phylogroup | Antibiotic-Resistance Profile |
|---|---|---|---|---|---|---|---|
| 5 | ND | + | ND | ND | ND | B1 | AMP, ATM, CEZ, CRO, CIP, GM, MIN, LVX, PIP, TET |
| 6 | ND | + | ND | ND | ND | B1 | AMP, CEZ, CRO, GM, PIP, TET |
| 9 | ND | + | ND | ND | ND | B1 | AMP, SAM, CEZ, CRO, GM, PIP, TET |
| 10 | ND | + | ND | ND | ND | B1 | AMP, CEZ, CRO, GM, PIP, TET |
| 13 | ND | + | ND | ND | ND | B1 | AMP, CEZ, CRO, GM, PIP, TET |
| 14 | ND | + | ND | ND | ND | B1 | AMP, CEZ, CRO, GM, PIP, TET |
| 16 | ND | + | ND | ND | ND | F | AMP, CEZ, CRO, CIP, GM, MIN, LVX, PIP, TET, SXT |
| 17 | ND | + | ND | ND | ND | F | AMP, CEZ, CRO, CIP, GM, LVX, PIP, TET, TOB, SXT |
| 38 | ND | + | + | ND | ND | A | AMP, SAM, CEZ, CAZ, CRO, CIP, GM, MIN, PIP, TET, SXT |
| 39 | ND | + | ND | + | ND | B1 | AMP, SAM, ATM, CEZ, CAZ, CRO, GM, MIN, PIP, TET |
| 40 | ND | + | ND | + | ND | B1 | AMP, SAM, ATM, CEZ, CAZ, CRO, CIP, MIN, PIP, TET |
| 50 | ND | + | ND | + | ND | A | AMP, SAM, CEZ, CRO, CIP, GM, MIN, LVX, PIP, TET, SXT |
| 52 | ND | + | ND | + | ND | A | AMP, SAM, CEZ, CRO, CIP, GM, LVX, PIP, TET, SXT |
| 53 | ND | + | ND | ND | ND | B1 | AMP, CEZ, CRO, CIP, TET |
| 65 | ND | + | ND | + | ND | A | AMP, SAM, CEZ, CRO, CIP, GM, LVX, PIP, TET, SXT |
| 70 | ND | + | ND | + | ND | A | AMP, SAM, CEZ, CRO, CIP, GM, LVX, PIP, TET |
| 71 | ND | + | ND | + | ND | F | AMP, SAM, CEZ, CRO, CIP, GM, PIP, TET |
| 73 | ND | + | ND | + | ND | B1 | AMP, SAM, CEZ, CRO, CIP, GM, LVX, PIP, TET, SXT |
| 80 | ND | + | ND | ND | ND | C | AMP, SAM, CEZ, CRO, CIP, PIP, TET, SXT |
| 86 | ND | + | ND | ND | ND | C | AMP, ATM, CEZ, CRO, CIP, GM, LVX, PIP, TET, TOB, SXT |
| 87 | ND | + | ND | ND | ND | C | AMP, ATM, CEZ, CRO, CIP, GM, LVX, PIP |
| 96 | ND | + | ND | ND | ND | B1 | AMP, CEZ, CRO, CIP, GM, LVX, PIP, TET, SXT |
| Strain Name | blaSHV | blaCTX-M | blaCMY | blaTEM | blaOXA | Phylogroup | Antibiotic-Resistance Profile |
|---|---|---|---|---|---|---|---|
| 3 | ND | ND | + | + | ND | F | AMP, CEZ, CAZ CRO, TET |
| 4 | ND | ND | + | + | ND | A | AMP, SAM, CEZ, CAZ CRO, GM, TET, SXT |
| 7 | ND | ND | + | + | ND | F | AMP, SAM, CEZ, CRO, GM, MIN, TET |
| 35 | ND | ND | + | ND | ND | C | AMP, CEZ, CRO, CIP, GM, LVX, TET, SXT |
| 37 | ND | ND | + | ND | ND | B1 | CIP, GM, LVX, TET, TOB |
| 38 | ND | + | + | ND | ND | A | AMP, SAM, CEZ, CRO, CIP, GM, PIP, TET, SXT |
| 57 | ND | ND | + | + | ND | B1 | AMP, TET |
| 63 | ND | ND | + | + | ND | B1 | AMP, CEZ, TET |
| 67 | ND | ND | + | + | ND | F | AMP, CEZ, CRO, TET, SXT |
| Gene Name | Numbers of the Strains | Phylogroup Numbers and (%) | |||||
|---|---|---|---|---|---|---|---|
| A | B1 | C | D | E | F | ||
| blaCTX-M | 22 | 5 (22.72%) | 11(50%) | 3 (13.63%) | - | - | 3 (13.63%) |
| blaCMY | 9 | 2 (22.22%) | 3 (33.33%) | 1 (11.11%) | - | - | 3 (33.33%) |
| blaTEM | 57 | 22 (38.59%) | 24 (42.10%) | 2 (3.5%) | 3 (5.26%) | 2 (3.5%) | 4 (7.01%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abo-Almagd, E.E.; Sabala, R.F.; Abd-Elghany, S.M.; Jackson, C.R.; Ramadan, H.; Imre, K.; Morar, A.; Herman, V.; Sallam, K.I. β-Lactamase Producing Escherichia coli Encoding blaCTX-M and blaCMY Genes in Chicken Carcasses from Egypt. Foods 2023, 12, 598. https://doi.org/10.3390/foods12030598
Abo-Almagd EE, Sabala RF, Abd-Elghany SM, Jackson CR, Ramadan H, Imre K, Morar A, Herman V, Sallam KI. β-Lactamase Producing Escherichia coli Encoding blaCTX-M and blaCMY Genes in Chicken Carcasses from Egypt. Foods. 2023; 12(3):598. https://doi.org/10.3390/foods12030598
Chicago/Turabian StyleAbo-Almagd, Elham Elsayed, Rana Fahmi Sabala, Samir Mohammed Abd-Elghany, Charlene R. Jackson, Hazem Ramadan, Kálmán Imre, Adriana Morar, Viorel Herman, and Khalid Ibrahim Sallam. 2023. "β-Lactamase Producing Escherichia coli Encoding blaCTX-M and blaCMY Genes in Chicken Carcasses from Egypt" Foods 12, no. 3: 598. https://doi.org/10.3390/foods12030598