
Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective

Abstract
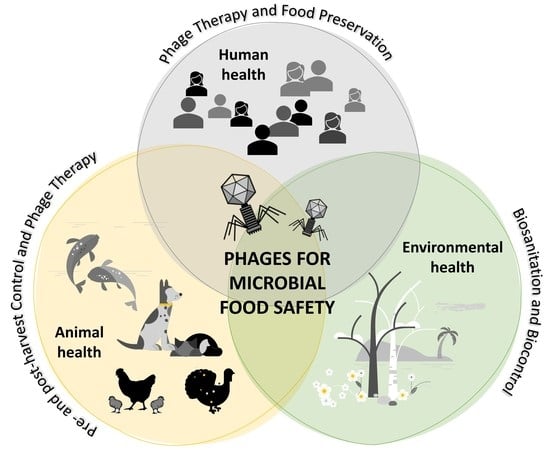
:
1. Introduction
2. Phage Biocontrol in Animal Husbandry for Food Production
2.1. Control of Campylobacter and Salmonella in Broilers

{kind=link}
{kind=link}
{kind=link}
| Animal (Age) | Bacteria Load 1,2 | Phage | Application Method and Dose 3 | Bacterial Reduction | Ref. |
|---|---|---|---|---|---|
| Campylobacter spp. | |||||
| Chickens (38 days old) | C. jejuni AMR * 108 (1) | ϕ16-izsam ϕ7-izsam | Oral (37 dpi); single dose; sequential application (24 h) 1 day before slaughtering. 107 108 | 1 log reduction 2 log reduction | [38] |
| Chickens (24 days old) | C. jejuni HPC5 108 (1) | Cocktail (2): CP20 GII CP30A GIII | Oral (4 dpi); single dose; 107 | 2.4 and 1.3 log reduction after 2 and 5 dpt | [39] |
| Chickens (25 days old) | C. jejuni HPC5 107 (1) C. jejuni GIIC8 107 (1) | CP34 GIII CP8 GIII CP8 GIII | Oral (5 dpi); single dose; 105–107–109 | 0.5–4 log reduction Marginal reductions Initial 5 log reduction and 2 log reduction after 5 dpt | [40] |
| Chickens (38 days old) | C. jejuni 2140CD1 107 (1) C. coli A11 106 (1) | Cocktail (3): ϕCcoIBB35 GII ϕCcoIBB37 GII ϕCcoIBB12 GII | Oral (7 dpi); single dose; 106 In feed (7dpi); single dose; 107 | 1.2 and 1.7 log reduction after 2 and 7 dpt 2 log reduction after 2 and 7 dpt | [41] |
| 3 field trials Chickens (36 days old) | C. jejuni 102–107 (1) | Cocktail (4): NCTC12672 GIII NCTC12673 GIII NCTC12674 GIII NCTC12678 GIII | Drinking water (7 dpi); single dose; 107 | Up to 3.2 log reduction in one field trial No reduction in two field trials | [42] |
| Chickens (47 days old) | Naturally colonized chickens | Cocktail (4): PH5, PH8, PH11, PH13 | Oral; single dose; 107 | 1.3 log reduction after 1 dpt | [43] |
| Chickens (10 days old) Chickens (32 days old) | C. jejuni C356 108–109 (1) | NCTC12671 GIII Cocktail (2): NCTC12671 GIII NCTC12669 GIII | Oral (5dpi); 5 doses (24 h interval); 1010–1011 Oral (7dpi); 4 doses (24 h interval); 1010–1011 | Initial 3 log reduction followed by 1 log reduction over 20 dpt Initial 1.5 log reduction followed by 1 log reduction over 20 dpt | [44] |
| Chickens (25 days old) | C. jejuni HPC5 107 (1) C. coli OR12 109 (1) | CP220 GII | Oral (5 dpi); single dose; 107 109 | 2 log reduction 2 log reduction | [45] |
| Chickens (27 days old) | C. jejuni 3871 109 (1) | CP14 GIII Cocktail (2): CP14 GIII CP81 GIII CP14 GIII CP68 GII | Oral (7 dpi); single dose; 5 × 108 Oral (7 dpi); single dose; 5 × 108 Oral (7 dpi); single dose; sequential application (24 h); 5 × 108–5 × 1010 | 1 log reduction after 3 dpt No reduction 3 log reduction after 3 dpt | [46] |
| Salmonella spp. | |||||
| Layer hens (6 weeks old) | S. Gallinarum KVCC BA00722 108 (2) | ST4 L13 SG3 | Feed additive 108 7 days before and 21 dpi | 50% reduction in liver and spleen after 7 dpi; 70% survival rate 75% and 50% reduction in liver and spleen after 7 and 14 dpi, respectively; 75% survival rate 25% and 50% reduction in liver and spleen after 7 and 14 dpi, respectively; 50% survival rate | [49] |
| Chickens (36 days old) | S. Enteritidis P125109 S. Typhimurium 4/74 108 (2) | Phage cocktail: ϕ151, ϕ25 ϕ10 | Oral; single dose 1011 | 1.53 log and 3.48 log reduction of S. Enteritidis and S. Typhimurium, respectively | [50] |
| Chickens (6-10 days) Chickens (31-35 days) | S. Enteritidis (PT4) 6 × 106 (2) | Phage cocktail: CNPSA1 CNPSA3 CNPSA4 | Early treatment after challenge: drinking water for 5 consecutive days (from 6 to 10 dpi) Later treatment after challenge: drinking water for 5 consecutive days (from 31 to 35 dpi) 109 | 1.08 log reduction after later treatment | [51] |
| Chickens (1 day old) (6 days old) | S. Enteritidis 103 (2) | Single phage or cocktail: CB4ϕ WT45ϕ | Cloacal drop 1 h pi: WT45ϕ: 109 Oral delivery 1 h pi: WT45ϕ: 108 CB4ϕ: 108 Cocktail: 108 | Reduction in Salmonella detection to 36% Reduction in Salmonella detection to 70%, 65%, and 45% after 1 dpt No significant differences after 48 h | [52] |
| Chickens (10 days old) | S. Enteritidis 105 (2) | Phage cocktail | Coarse spray or drinking water 108 | Reduction in Salmonella detection to 72.7% by aerosol-spray | [53] |
| Layer hens (40 weeks old) | S. Enteritidis (SENAR) 108 (2) | Phage cocktail: SP-1 STP-1 | Feed additive: 0.2% of the phage cocktail | 0.9, 0.57, and 0.38 log reduction in cecum, liver, and spleen at 7 dpt 0.86 log reduction in cecum at 6 dpt | [54] |
| Layer hens (60 weeks old) | Natural infection | Autophage (AP) Wild-type phage | Spray applications 108 Two single applications in 24 h intervals | 1.78 log reduction in feces samples Total elimination of Salmonella from the environment | [55] |
| Chickens (1 to 35 days) | S. Enteritidis 104 (2) | Bafasal (4 phages cocktail) | Feed additive daily 106 | 1 log reduction at day 35 of study | [56] |
2.2. Control of Listeria Monocytogenes in Animals
2.3. Control of Vibrio spp. in Aquaculture
3. Phage Biocontrol at the Post-Harvest and Post-Slaughtering Stage
3.1. Campylobacter
| Food | Bacteria Load 1 | Phage | Application MOI * and Method | Result/Bacterial Reduction | Ref. |
|---|---|---|---|---|---|
| Campylobacter spp. | |||||
| Chicken skin | C. jejuni PT14 4 and 6 log | ϕ2: NCTC12674 GIII | MOI: 0.01–1 MOI: 10–103 spread on surface | Negligible reduction 1 log reduction after 30 min, 3 d and 5 d at 4 °C 2.5 log reduction after 5 d at −20 °C | [26] |
| Chicken skin | C. jejuni C222 4 log | NCTC12673 GIII | MOI: 102 spread on surface | 1 log reduction after 1 d at 4 °C | [94] |
| Chicken neck skin | C. jejuni NCTC12662 4 log | F356GIII F357 GIII F379 GIII Cocktail (2): F356 GIII F357 GIII | MOI: 103 spread on surface | 0.5 log reduction at 5 °C 0.5 log reduction at 5 °C Negligible reduction at 5 °C 0.7 log reduction after 1 d at 5 °C | [95] |
| Chicken meat | C. coli NCTC 126683 C. jejuni NCTC 11168 3 log | NCTC12684 GII CP81 GIII | MOI: 104 spread on surface | No reduction at 4 °C No reduction at 4 °C | [96] |
| Raw and cooked beef | C. jejuni 4 log | Cj6 | MOI: 104 spread on surface | 1.5 and 2 log reduction after 1 d at 5 °C in raw and cooked beef, respectively | [97] |
| Chicken meat | C. jejuni 4 log | CJ01 | MOI: 102 spread on surface | 1.7 log reduction after 2 d at 4 °C | [98] |
| Salmonella spp. | |||||
| Commercial broiler and turkey carcasses | S. Enteritidis (PT 13A) 20 CFU S. Enteritidis (PT 13A) 31 CFU S. Enteritidis host S. Enteritidis field (S9, S14) | PHL 4 72 wild-type phages | Broiler carcasses: MOI: 104 to 1010 spray Turkey carcasses: MOI: 106 to 108 rinsed | 50–100% reduction 60% reduction | [100] |
| Breast and eggs | S. Enteritidis LK5, UA1894 Breast: 106 Eggs: 107 | UAB_Phi 20 UAB_Phi78 UAB_Phi87 | 109 PFU (MOI: 103) rinse 1010 (MOI:103) spray | 2.0 log reduction 0.9 log reduction | [101] |
| Liquid eggs and chicken meat | S. Enteritidis Liquid eggs: 104 Chicken meat: 105 | SE07 | 1011 (MOI 107) Direct addition of 100 mL 1012 (MOI 107) spray | 2 log reduction after 12, 24, and 48 h 2 log reduction after 12, 24, and 48 h | [102] |
| Breast samples | S. Enteritidis ATCC13076 CVCC2184 4 × 105 | PA13076 PC2184 | Single phage: 4 × 109 (MOI: 104) Cocktail: 4 × 109 (MOI: 104) | 2 log reduction Phage PC2184 better than phage PA13076 at 4 °C and 25 °C 2 log reduction | [103] |
| Chicken breast | S. Typhimurium ATCC 14,028 S. Enteritidis ATCC 4931 S. Heidelberg ATCC 8326 3 logs | SalmoFresh TM (6 phages) | MOI: 106 spray | 0.7 and 0.9 log reduction on day 0 and 1, at 4 °C 1 log reduction on day 7 with modified atmosphere at 4 °C 0.8, 0.9, and 0.4 log reduction at 0, 4, and 8 h at room temperature, respectively | [104] |
| Chicken and turkey meat | S. Enteritidis ATCC 13,076 S. Typhimurium ATCC 6539 S. Heidelberg ATCC 8326 1.5 × 103 1.25 × 103 | SalmoLyse® | 2 × 106, 4 × 106, 9 × 106 MOI: 2 × 103, 3 × 103, 6 × 103 spray 9 × 106 and 2 × 107 MOI: 7 × 103, 1 × 104 spray | 60%, 71%, and 88% reduction from chicken meat at 2 × 106, 4 × 106, 9 × 106 PFU/mL, respectively 68% and 86% reduction from turkey meat at 106 and 107 PFU/g, respectively | [105] |
| Chicken meat | S. Typhimurium JCW-3001 S. Enteritidis VDL-133 S. Dublin SP-598 5 log | SalmoFREE® (6 phages) | 108, 109 (MOI: 103, 104) immersion | 1.9–2.0 log reduction in combination with plant-based essential oils | [106] |
| Chicken meat | S. Enteritidis 104 | PhageGuard S® (2 phages) | 107 (MOI: 103) immersion | 1.5 log reduction after 24 h | [107] |
| Listeria monocytogenes | |||||
| Raw salmon | 4 log 2 log | Listex™ P100 | MOI: 1, 10, 102, 103, 104 spread on surface MOI: 106 spread on surface | Marginal reductions at lower MOIs, but up to 3 log reduction at higher MOIs 1.4 log reduction (1 d) No regrowth after 10 d at 4 °C | [108] |
| Raw hake Raw salmon Smoked salmon | 3 log | Listex™ P100 | Automated spray MOI: 104 | 1.2 and 2.0 log reduction after 1 d and 7 d at 4°C (hake) 0.8 and 1.0 log reduction after 1 d and 7 d at 4°C (raw salmon) 0.8 and 1.6 log reduction after 1 d and 30 d at 4°C (smoked salmon) | [109] |
| Smoked salmon | 3 log | ListShield™ (6 phages) | MOI: 103 spray | 0.4 and 1 log reduction | [110] |
| RTE chicken breast roll | 2, 4, and 5 log | FWLLm1 | MOI: 105, 103, 102 spread on surface | Rapid 1.5–2.5 log at 5–30 °C. Regrowth prevented over 21 d at higher MOI and 5 °C (vacuum packed) | [111] |
| Cooked turkey and roast beef | 3 log | Listex™ P100 | MOI:104 spread on surface | 1.7 log and 2.1 log, respectively, after 28 d at 4 °C | [112] |
| Sliced cooked ham | 4 log | Listex™ P100 | MOI: 104 spread on surface | Rapid 1 log reduction 2 log reduction after 28 d at 4 °C | [113] |
| Dry-cured ham | 2, 3, 4 log | Listex™ P100 | MOI: 102–106 spread on surface | 2.5 log to undetectable (highest MOI) after 14 d at 4 °C | [114] |
| Milk | 5 log | Monophages LMP1 and LMP7 | MOI:10 addition to milk | 0.5–3.3 log at 4 °C | [115] |
| “Queso fresco” cheese | 4 log | Listex™ P100 | MOI: 104 spread on surface | 2 log reduction | [116] |
| Soft cheeses | 3 log 1, 2 log | A511 | MOI: 105 in the smearing solution MOI: 106, 107 | 2.5–3 log reduction during the 21 d ripening period >6 log reduction (below the limit of detection) | [117] |
| Hard cheese | 4 log | ListShield™ (6 phages) | MOI: 104 spray | 0.7 log reduction | [110] |
| Lettuce Apple slices | 3 log 4 log | ListShield™ (6 phages) | MOI: 104, 105 spray MOI: 102 spray | 1.1 log reduction at higher MOI 1 log reduction | [110] |
| Fresh-cut apple and melon | 5.5 log | Cocktail (12 phages) LM-103 Cocktail (6 phages) LMP-102 | MOI: 102 spray | Below 0.4 log reduction in apple 2.0–4.6 log reduction in melon | [118] |
| Fresh-cut apple, pear, and melon slices. Apple, pear, and melon juices | 5 log 5 log | Listex™ P100 | MOI: 103 spread on surface MOI: 103 addition to juice | None, 0.6, and 1.5 log reduction in apple, pear, and melon slices after 8 d at 10 °C None, 2, and 8 log reduction in apple, pear, and melon juices after 8 d at 10 °C | [119] |
| Celery and enoki mushroom | 5 log | Mix of 3 phages: LMPC01 LMPC02 LMPC03 | MOI: 10 | 2.2 and 1.8 log reduction in celery and enoki mushroom after 7 d at 4 °C | [120] |
| Vibriospp. | |||||
| Oysters Crassostrea gigas | V. parahaemolyticus CRS 09-17, AMR* 1.6 × 106 CFU in each oyster | pVp-1 | 2 × 107 PFU/oyster (MOI: 10) surface of flesh | 6 log CFU/mL growth reduction after 12 h | [80] |
| Fresh fish flesh | V. parahaemolyticus FORC_023 3 × 104 | VPG01 | MOI: 1 MOI: 10 surface direct application | 1 log reduction (MOI: 1) Counts under the detection limit after 6 h (MOI: 10) | [81] |
| Cutting board | V. parahaemolyticus FORC_023 3 × 104 CFU/cm2 | VPG01 | MOI: 103 surface direct application | 3 log reduction in utensil surface | [81] |
| Raw fish flesh slices | V. parahaemolyticus FORC_023 3 × 104 | VPT02 | MOI of 0, 1, or 10 surface direct application | 2 log reduction after 6 h at 25 °C (MOI: 10) | [121] |
| Shrimp | V. parahaemolyticus F23 | F23s1 Recombinant endolysin ORF52 | MOI: 103 in vitro 20 µmol/L | Growth inhibition at 25 °C for 12 h Decreased OD600 after 60 min The endolysin also showed lytic activity against a panel of 23 drug-resistant V. parahaemolyticus | [122] |
| Manila clams | V. parahaemolyticus Vp-KF4 1 × 104 | Vpp2 | MOI of 1, 10, or 100 | 2.1 log reduction at 25 °C until 24 h No effect of treatment at 4 °C | [123] |
| Oysters | V. parahaemolyticus ATCC 17802) 104 | vB_VpaS_OMN | MOI: 103 surface direct application | 1 log and 2 log reduction after 48 and 72 h of incubation, respectively | [124] |
| Oysters | V. vulnificus 106 | Phage pool (9 phages): S1, P3, P38, P53, P65, P68, P108, P111, P147 | Unknown | 5 log reduction after 18 h of incubation at 4 °C | [125] |
| Abalone flesh | V. vulnificus MO6-24/O 2 × 103 | VVP001 | MOI: 105 MOI: 106 | 2.06 log reduction 2.51 log reduction | [126] |
3.2. Salmonella
3.3. Listeria Monocytogenes
3.4. Human Pathogenic Vibrio spp.
4. Challenges of Using Phages for Food Safety
| Challenge | Causes and/or Future Studies Needs | Refs. |
|---|---|---|
| Extreme host specificity |
| [144,145] |
| Potential development of phage resistance |
| [145,146,147] |
| Phage stability and administration routes |
| [148,149,150,151] |
| Mobilization of resistant genes between bacteria |
| [152,153] |
| Phage biobanking for immediate trials |
| [154] |
| Legislative approval |
| [155,156] |
| Consumer acceptance |
| [89] |
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Endersen, L.; O’Mahony, J.; Hill, C.; Ross, R.P.; McAuliffe, O.; Coffey, A. Phage Therapy in the Food Industry. Ann. Rev. Food Sci. Technol. 2014, 5, 327–349. [Google Scholar] [CrossRef]
- Mousavi, S.L.; Rasooli, I.; Nazarian, S.; Amani, J. Simultaneous detection of Escherichia coli O157:H7, toxigenic Vibrio cholerae, and Salmonella typhimurium by multiplex PCR. Iran. J. Clin. Infect. Dis. 2009, 4, 97–103. [Google Scholar]
- Gould, L.H.; Seys, S.; Everstine, K.; Norton, D.; Ripley, D.; Reimann, D.; Dreyfuss, M.; Chen, W.S.; Selman, C.A. Recordkeeping Practices of Beef Grinding Activities at Retail Establishments. J. Food Prot. 2011, 74, 1022–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M.; Tauxe, R.; Hednerg, C. The growing burden of foodborne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.L.; Lee, H.Y.; Mahyudin, N.A. Antimicrobial resistance of Escherichia coli and Staphylococcus aureus isolated from food handler’s hands. Food Control 2014, 44, 203–207. [Google Scholar] [CrossRef]
- WHO; FAO; OIE. Taking a Multisectoral, One Health Approach: A Tripartite Guide to Addressing Zoonotic Diseases in Countries. 2019. Available online: https://www.woah.org/app/uploads/2021/03/en-tripartitezoonosesguide-webversion.pdf (accessed on 3 August 2022).
- Gordillo-Altamirano, F.L.; Barr, J.J. Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. UK, 2014. Available online: https://amr-review.org (accessed on 29 July 2022).
- Jamal, M.; Bukhari, S.M.A.U.S.; Andleeb, S.; Ali, M.; Raza, S.; Nawaz, M.A.; Hussain, T.; Rahman, S.U.; Shah, S.S.A. Bacteriophages: An overview of the control strategies against multiple bacterial infections in different fields. J. Basic Microbiol. 2019, 59, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Nagel, T.E.; Chan, B.K.; De Vos, D.; El-Shibiny, A.; Kang’ethe, E.K.; Makumi, A.; Pirnay, J.-P. The Developing World Urgently Needs Phages to Combat Pathogenic Bacteria. Front. Microbiol. 2016, 7, 882. [Google Scholar] [CrossRef] [PubMed]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage applications for food production and processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, L.; Bolton, D.; McAuliffe, O.; Coffey, A. Bacteriophages in food applications: From foe to friend. Annu. Rev. Food Technol. 2019, 15, 151–172. [Google Scholar] [CrossRef]
- Young, I.; Wang, I.; Roof, W.D. Phages will out: Strategies of host cell lysis. Trends Microbiol. 2000, 8, 120–128. [Google Scholar] [CrossRef]
- Bernhardt, T.G.; Wang, I.N.; Struck, D.K.; Young, R. Breaking free: “protein antibiotics” and phage lysis. Res. Microbiol. 2002, 153, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Woolston, J.; Sulakvelidze, A. Phage Biocontrol Applications in Food Production and Processing. Curr. Issues Mol. Biol. 2021, 40, 267–302. [Google Scholar] [CrossRef] [PubMed]
- Pirnay, J.P.; Merabishvili, M.; Van Raemdonck, H.; DeVis, D.; Verbeken, G. Bacteriophage production in compliance with regulatory requirements. In Bacteriophage Therapy: From Lab to Clinical Practice; Azeredo, J., Sillankorva, S., Eds.; Humana Press: New York, NY, USA, 2017; Book Series: Methods in Molecular Biology; Volume 1693, pp. 233–252. [Google Scholar] [CrossRef]
- Alomari, M.M.M.; Dec, M.; Urban-Chmiel, R. Bacteriophages as an Alternative Method for Control of Zoonotic and Foodborne Pathogens. Viruses 2021, 13, 2348. [Google Scholar] [CrossRef]
- Kutateladze, M.; Adamia, R. Bacteriophages as potential new therapeutics to replace or supplement antibiotics. Trends Biotechnol. 2010, 28, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii Infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef] [Green Version]
- Doss, J.; Culbertson, K.; Hahn, D.; Camacho, J.; Barekzi, N. A Review of Phage Therapy against Bacterial Pathogens of Aquatic and Terrestrial Organisms. Viruses 2017, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodridge, L.D.; Bisha, B. Phage-based biocontrol strategies to reduce foodborne pathogens in foods. Bacteriophage 2011, 1, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Sillankorva, S.M.; Oliveira, H.; Azeredo, J. Bacteriophages and their role in food safety. Inter. J. Microbiol 2012, 12, 863945. [Google Scholar] [CrossRef] [Green Version]
- Mosimann, S.; Desiree, K.; Ebner, P. Efficacy of Phage Therapy in Poultry: A Systematic Review and Meta-Analysis. Poult. Sci. 2021, 100, 101472. [Google Scholar] [CrossRef]
- Summers, W.C. Bacteriophage therapy. Annu. Rev. Microbiol. 2001, 55, 437–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, J.R.; March, J.B. Bacteriophages and biotechnology: Vaccines, gene therapy and antibacterials. Trends Biotechnol. 2006, 24, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Atterbury, R.J.; Connerton, P.L.; Dodd, C.E.; Rees, C.E.; Connerton, I.F. Application of host specific bacteriophages to the surface of chicken skin leads to a reduction in recovery of Campylobacter jejuni. Appl. Environ. Microbiol. 2003, 69, 6302–6306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueno, E.; García, P.; Martínez, B.; Rodríguez, A. Phage inactivation of Staphylococcus aureus in fresh and hard-type cheeses. Int. J. Food Microbiol. 2012, 158, 23–27. [Google Scholar] [CrossRef] [PubMed]
- EFSA; ECDC. The European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, 7666. [Google Scholar] [CrossRef]
- Sevilla-Navarro, S.; Catalá-Gregori, P.; Marin, C. Salmonella Bacteriophage Diversity According to Most Prevalent Salmonella Serovars in Layer and Broiler Poultry Farms from Eastern Spain. Animals 2020, 10, 1456. [Google Scholar] [CrossRef]
- Sifre, E.; Salha, B.A.; Ducournau, A.; Floch, P.; Chardon, H.; Megraud, F.; Lehours, P. EUCAST recommendations for antimicrobial susceptibility testing applied to the three main Campylobacter species isolated in humans. J. Microbiol. Methods 2015, 119, 206–213. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global Priority List of Antibiotic Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 29 July 2022).
- Olson, E.G.; Micciche, A.C.; Rothrock, M.J., Jr.; Yang, Y.; Ricke, S.C. Application of Bacteriophages to Limit Campylobacter in Poultry Production. Front. Microbiol. 2022, 12, 458721. [Google Scholar] [CrossRef]
- EFSA BIOHAZ Panel (European Food Safety Authority Panel on Biological Hazards). Update and review of control options for Campylobacter in broilers at primary production. EFSA J. 2020, 18, 6090. [Google Scholar] [CrossRef]
- Mota-Gutierrez, J.; Lis, L.; Lasagabaster, A.; Nafarrate, I.; Ferrocino, I.; Cocolin, L.; Rantsiou, K. Campylobacter spp. prevalence and mitigation strategies in the broiler production chain. Food Microbiol. 2022, 104, 103998. [Google Scholar] [CrossRef] [PubMed]
- Nafarrate, I.; Mateo, E.; Miranda, K.; Lasagabaster, A. Isolation, host specificity and genetic characterization of Campylobacter specific bacteriophages from poultry and swine sources. Food Microbiol. 2021, 97, 103742. [Google Scholar] [CrossRef]
- Sails, A.D.; Wareing, D.R.A.; Bolton, F.J.; Fox, A.J.; Curry, A. Characterisation of 16 Campylobacter jejuni and C. coli typing bacteriophages. J. Med. Microbiol. 1998, 47, 123–128. [Google Scholar] [CrossRef]
- D’Angelantonio, D.; Scattolini, S.; Boni, A.; Neri, D.; Di Serafino, G.; Connerton, P.; Connerton, I.; Pomilio, F.; Di Giannatale, E.; Migliorati, G.; et al. Bacteriophage Therapy to Reduce Colonization of Campylobacter jejuni in Broiler Chickens before Slaughter. Viruses 2021, 13, 1428. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.J.; Connerton, P.L.; Connerton, I. Phage Biocontrol of Campylobacter jejuni in Chickens Does Not Produce Collateral Effects on the Gut Microbiota. Front. Microbiol. 2019, 10, 476. [Google Scholar] [CrossRef] [PubMed]
- Loc-Carrillo, C.; Atterbury, R.J.; El Shibiny, A.; Connerton, P.L. Bacteriophage therapy to reduce Campylobacter jejuni colonization of broiler chickens. Appl. Environ. Microbiol. 2005, 71, 6554–6563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, C.M.; Gannon, B.W.; Halfhide, D.E.; Santos, S.B.; Hayes, C.M.; Roe, J.M.; Azeredo, J. The in vivo efficacy of two administration routes of a phage cocktail to reduce numbers of Campylobacter coli and Campylobacter jejuni in chickens. BMC Microbiol. 2010, 10, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kittler, S.; Fischer, S.; Abdulmawjood, A.; Glünder, G.; Klein, G. Effect of Bacteriophage Application on Campylobacter jejuni Loads in Commercial Broiler Flocks. Appl. Environ. Microbiol. 2013, 79, 7525–7533. [Google Scholar] [CrossRef] [Green Version]
- Chinivasagam, H.N.; Estella, W.; Maddock, L.; Mayer, D.G.; Weyand, C.; Connerton, P.L.; Connerton, I.F. Bacteriophages to control Campylobacter in commercially farmed broiler chickens in Australia. Front. Microbiol. 2020, 11, 632. [Google Scholar] [CrossRef]
- Wagenaar, J.A.; Van Bergen, M.A.; Mueller, M.A.; Wassenaar, T.M.; Carlton, R.M. Phage therapy reduces Campylobacter jejuni colonization in broilers. Veter. Microbiol. 2005, 109, 275–283. [Google Scholar] [CrossRef]
- El-Shibiny, A.; Scott, A.; Timms, A.; Metawea, Y.; Connerton, P.; Connerton, I. Application of a Group II Campylobacter Bacteriophage To Reduce Strains of Campylobacter jejuni and Campylobacter coli Colonizing Broiler Chickens. J. Food Prot. 2009, 72, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Hammerl, J.A.; Jäckel, C.; Alter, T.; Janzcyk, P.; Stingl, K.; Knüver, M.T.; Hertwig, S. Reduction of Campylobacter jejuni in broiler chicken by successive application of group II and group III phages. PLoS ONE 2014, 9, e114785. [Google Scholar] [CrossRef] [PubMed]
- WHO. Salmonella (Non-Typhoidal). Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 11 August 2022).
- Dawoud, T.M.; Davis, M.L.; Park, S.H.; Kim, S.A.; Kwon, Y.M.; Jarvis, N.; O’Bryan, C.A.; Shi, Z.; Crandall, P.G.; Ricke, S.C. The Potential Link between Thermal Resistance and Virulence in Salmonella: A Review. Front. Vet. Sci. 2017, 4, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.S.; Jeong, J.; Lee, J.; Kim, S.; Min, W.; Myung, H. Therapeutic Effects of Bacteriophages against Salmonella Gallinarum Infection in Chickens. J. Microbiol. Biotechnol. 2013, 23, 1478–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atterbury, R.J.; van Bergen, M.A.P.; Ortiz, F.; Lovell, M.A.; Harris, J.A.; de Boer, A.; Wagenaar, J.A.; Allen, V.M.; Barrow, P.A. Bacteriophage Therapy to Reduce Salmonella Colonization of Broiler Chickens. Appl. Environ. Microbiol. 2007, 73, 4543–4549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaz, C.S.L.; Voss-Rech, D.; Alves, L.; Coldebella, A.; Brentano, L.; Trevisol, I.M. Effect of Time of Therapy with Wild-Type Lytic Bacteriophages on the Reduction of Salmonella Enteritidis in Broiler Chickens. Vet. Microbiol. 2020, 240, 108527. [Google Scholar] [CrossRef]
- Andreatti, R.L.; Higgins, J.P.; Higgins, S.E.; Gaona, G.; Wolfenden, A.D.; Tellez, G.; Hargis, B.M. Ability of bacteriophages isolated from different sources to reduce Salmonella enterica serovar Enteritidis in vitro and in vivo. Poult. Sci. 2007, 86, 1904–1909. [Google Scholar] [CrossRef]
- Borie, C.; Albala, I.; Sánchez, P.; Sánchez, M.L.; Ramírez, S.; Navarro, C.; Morales, M.A.; Retamales, J.; Robeson, J. Bacteriophage treatment reduces Salmonella colonization of infected chickens. Avian Dis. 2008, 52, 64–67. [Google Scholar] [CrossRef]
- Adhikari, P.A.; Cosby, D.E.; Cox, N.A.; Lee, J.H.; Kim, W.K. Effect of dietary bacteriophage supplementation on internal organs, fecal excretion, and ileal immune response in laying hens challenged by Salmonella Enteritidis. Poult. Sci. 2017, 96, 3264–3271. [Google Scholar] [CrossRef]
- Sevilla-Navarro, S.; Marín, C.; Cortés, V.; García, C.; Vega, S.; Catalá-Gregori, P. Autophage as a control measure for Salmonella in laying hens. Poult. Sci. 2018, 97, 4367–4373. [Google Scholar] [CrossRef]
- Wójcik, E.A.; Stańczyk, M.; Wojtasik, A.; Kowalska, J.D.; Nowakowska, M.; Łukasiak, M.; Bartnicka, M.; Kazimierczak, J.; Dastych, J. Comprehensive Evaluation of the Safety and Efficacy of BAFASAL® Bacteriophage Preparation for the Reduction of Salmonella in the Food Chain. Viruses 2020, 12, 742. [Google Scholar] [CrossRef] [PubMed]
- EFSA; ECDC. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from hu-mans, animals and food in 2019–2020. EFSA J. 2022, 20, 7209. [Google Scholar] [CrossRef]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th Revision 2018, Ranking of Medically Important Antimicrobials for Risk Management of Antimicrobial Resistance Due to Non-Human Use. 2018. Available online: https://apps.who.int/iris/bitstream/handle/10665/312266/9789241515528-eng.pdf (accessed on 29 July 2022).
- Carvalho, C.; Costa, A.R.; Silva, F.; Oliveira, A. Bacteriophages and Their Derivatives for the Treatment and Control of Food-Producing Animal Infections. Crit. Rev. Microbiol. 2017, 43, 583–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabil, N.M.; Tawakol, M.M.; Hassan, H.M. Assessing the Impact of Bacteriophages in the Treatment of Salmonella in Broiler Chickens. Infect. Ecol. Epidemiol. 2018, 8, 1539056. [Google Scholar] [CrossRef] [PubMed]
- Żbikowska, K.; Michalczuk, M.; Dolka, B. The Use of Bacteriophages in the Poultry Industry. Animals 2020, 10, 872. [Google Scholar] [CrossRef]
- Lim, T.-H.; Lee, D.-H.; Lee, Y.-N.; Park, J.-K.; Youn, H.-N.; Kim, M.-S.; Lee, H.-J.; Yang, S.-Y.; Cho, Y.-W.; Lee, J.-B.; et al. Efficacy of Bacteriophage Therapy on Horizontal Transmission of Salmonella Gallinarum on Commercial Layer Chickens. Avian Dis. 2011, 55, 435–438. [Google Scholar] [CrossRef]
- Nakari, U.M.; Rantala, L.; Pihlajasaari, A.; Toikkanen, S.; Johansson, T.; Hellsten, C.; Raulo, S.M.; Kuusi, M.; Siitonen, A.; Rimhanen-Finne, R. Investigation of increased listeriosis revealed two fishery production plants with persistent Listeria contamination in Finland in 2010. Epidemiol. Infect. 2014, 142, 2261–2269. [Google Scholar] [CrossRef]
- Skowron, K.; Kwiecinska, J.; Grudlewska, K.; Swieca, A.; Paluszak, Z.; Bauza, J.; Wałecka, E.; Gospodarek, E. The occurrence, transmission, virulence and antibiotic resistance of Listeria monocytogenes in fish processing plant. Int. J. Food Microbiol. 2018, 282, 71–83. [Google Scholar] [CrossRef]
- Morwal, S.; Sharma, S.K. Bacterial zoonosis—A public health importance. J. Dairy Vet. Anim. Res. 2017, 5, 56–59. [Google Scholar] [CrossRef]
- Loessner, M.; Carlton, R.M. Virulent Phages to Control Listeria monocytogenes in Foodstuffs and in Food Processing Plants. US Patent US7438901B2, 28 August 2008. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2004004495 (accessed on 13 October 2022).
- Li, T.; Zhao, X.; Wang, X.; Wang, Z.; Tian, C.; Shi, W.; Qi, Y.; Wei, H.; Song, C.; Xue, H.; et al. Characterization and Preliminary application of phage isolated from Listeria monocytogenes. Front. Vet. Sci. 2022, 9, 946814. [Google Scholar] [CrossRef]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef] [Green Version]
- Neetoo, H.; Reega, K.; Manoga, Z.S.; Nazurally, N.; Bhoyroo, V.; Allam, M.; Jaufeerally-Fakim, Y.; Ghoorah, A.W.; Jaumdally, W.; Hossen, A.M.; et al. Prevalence, genomic characterization, and risk assessment of human pathogenic Vibrio Species in Seafood. J. Food Prot. 2022, 85, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Prim. 2018, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, M.; LewisOscar, F.; Thajuddin, N.; Pugazhendhi, A.; Nithya, C. In vitro and in vivo biofilm forming Vibrio spp: A significant threat in aquaculture. Process Biochem. 2020, 94, 213–223. [Google Scholar] [CrossRef]
- Plaza, N.; Castillo, D.; Pérez-Reytor, D.; Higuera, G.; García, K.; Bastías, R. Bacteriophages in the control of pathogenic vibrios. Electron. J. Biotechnol. 2018, 31, 24–33. [Google Scholar] [CrossRef]
- Cascarano, M.C.; Stavrakidis-Zachou, O.; Mladineo, I.; Thompson, K.D.; Papandroulakis, N.; Katharios, P. Mediterranean Aquaculture in a Changing Climate: Temperature Effects on Pathogens and Diseases of Three Farmed Fish Species. Pathogens 2021, 10, 1205. [Google Scholar] [CrossRef]
- Harrison, J.; Nelson, K.; Morcrette, H.; Morcrette, C.; Preston, J.; Helmer, L.; Titball, R.W.; Butler, C.S.; Wagley, S. The increased prevalence of Vibrio species and the first reporting of Vibrio jasicida and Vibrio rotiferianus at UK shellfish sites. Water Res. 2022, 211, 117942. [Google Scholar] [CrossRef]
- Scarano, C.; Spanu, C.; Ziino, G.; Pedonese, F.; Dalmasso, A.; Spanu, V.; Virdis, S.; De Santis, E.P.L. Antibiotic resistance of Vibrio species isolated from Sparus aurata reared in Italian mariculture. New Microbiol. 2014, 37, 329–337. [Google Scholar]
- Elmahdi, S.; DaSilva, L.V.; Parveen, S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: A review. Food Microbiol. 2016, 57, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Yano, Y.; Hamano, K.; Satomi, M.; Tsutsui, I.; Ban, M.; Aue-umneoy, D. Prevalence and antimicrobial susceptibility of Vibrio species related to food safety isolated from shrimp cultured at inland ponds in Thailand. Food Control 2014, 38, 30–36. [Google Scholar] [CrossRef]
- European Commission. DG Maritime Affairs and Fisheries. Strategic Guidelines for a More Sustainable and Competitive EU Aquaculture for the Period 2021 to 2030; Document 52021DC0236, COM/2021/236 final; European Commission: Brussels, Belgium, 2021.
- Onarinde, B.A.; Dixon, R.A. Prospects for Biocontrol of Vibrio parahaemolyticus Contamination in Blue Mussels (Mytilus edulus)-A Year-Long Study. Front. Microbiol. 2018, 9, 1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.W.; Kim, H.J.; Yun, S.K.; Chai, J.Y.; Park, S.C. Eating oysters without risk of vibriosis: Application of a bacteriophage against Vibrio parahaemolyticus in oysters. Int. J. Food Microbiol. 2014, 188, 31–35. [Google Scholar] [CrossRef]
- Lee, J.H.; Oh, M.; Kim, B.S. Phage biocontrol of zoonotic food-borne pathogen Vibrio parahaemolyticus for seafood safety. Food Control 2023, 144, 109334. [Google Scholar] [CrossRef]
- Vinod, M.G.; Shivu, M.M.; Umesha, K.R.; Rajeeva, B.C.; Krohneb, G. Isolation of Vibrio harveyi bacteriophage with a potential for biocontrol of luminous vibriosis in hatchery environments. Aquaculture 2006, 255, 117–124. [Google Scholar] [CrossRef]
- Droubogiannis, S.; Katharios, P. Genomic and biological profile of a novel Vibrio phage, Virtus, which improves survival of Sparus aurata larvae challenged with Vibrio harveyi. Pathogens 2022, 11, 630. [Google Scholar] [CrossRef] [PubMed]
- Higuera, G.; Bastías, R.; Tsertsvadze, G.; Romero, J.; Espejo, R.T. Recently discovered Vibrio anguillarum phages can protect against experimentally induced vibriosis in Atlantic salmon, Salmo salar. Aquaculture 2013, 395, 128–133. [Google Scholar] [CrossRef]
- Silva, Y.J.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, Â.; Calado, R.; Gomes, N.; Pardo, M.A.; Hernandez, I.; Almeida, A. Phage Therapy as an Approach to Prevent Vibrio anguillarum Infections in Fish Larvae Production. PLoS ONE 2014, 9, e114197. [Google Scholar] [CrossRef]
- Lomelí-Ortega, C.O.; Martínez-Díaz, S.F. Phage therapy against Vibrio parahaemolyticus infection in the whiteleg shrimp (Litopenaeus vannamei) larvae. Aquaculture 2014, 434, 208–211. [Google Scholar] [CrossRef]
- Karunasagar, I.; Shivu, M.M.; Girisha, S.K.; Krohne, G.; Karunasagar, I. Biocontrol of pathogens in shrimp hatcheries using bacteriophages. Aquaculture 2007, 268, 288–292. [Google Scholar] [CrossRef]
- Nakai, T.; Park, S.C. Bacteriophage therapy of infectious diseases in aquaculture. Res. Microbiol. 2002, 153, 13–18. [Google Scholar] [CrossRef]
- Lasagabaster, A.; Jiménez, E.; Lehnherr, T.; Miranda-Cadena, K.; Lehnherr, H. Bacteriophage biocontrol to fight Listeria outbreaks in seafood. Food Chem. Toxicol. 2020, 145, 111682. [Google Scholar] [CrossRef] [PubMed]
- Sulakvelidze, A. Using lytic bacteriophages to eliminate or significantly reduce contamination of food by foodborne bacterial pathogens. J. Sci. Food Agric. 2013, 93, 3137–3146. [Google Scholar] [CrossRef] [PubMed]
- Bolocan, A.S.; Callanan, J.; Forde, A.; Ross, P.; Hill, C. Phage therapy targeting Escherichia coli-a story with no end? FEMS Microbiol. Lett. 2016, 363, fnw256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, D.; Fernández, L.; Martínez, B.; Ruas-Madiedo, P.; García, P.; Rodríguez, A. Real-Time Assessment of Staphylococcus aureus Biofilm Disruption by Phage-Derived Proteins. Front. Microbiol. 2017, 8, 1632. [Google Scholar] [CrossRef]
- Hagens, S.; Loessner, M.J. Application of bacteriophages for detection and control of foodborne pathogens. Appl. Microbiol. Biotechnol. 2007, 76, 513–519. [Google Scholar] [CrossRef]
- Goode, D.; Allen, V.M.; Barrow, P.A. Reduction of experimental Salmonella and Campylobacter contamination of chicken skin by application of lytic bacteriophages. Appl. Environ. Microbiol. 2003, 69, 5032–5036. [Google Scholar] [CrossRef] [Green Version]
- Zampara, A.; Sørensen, M.C.; Elsser-Gravesen, A.; Brøndsted, L. Significance of phage-host interactions for biocontrol of Campylobacter jejuni in food. Food Control 2017, 73, 1169–1175. [Google Scholar] [CrossRef]
- Orquera, S.; Golz, G.; Hertwig, S.; Hammerl, J.; Sparborth, D.; Joldic, A.; Alter, T. Control of Campylobacter spp. and Yersinia enterocolitica by virulent bacteriophages. J. Mol. Genet. Med. 2012, 6, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Bigwood, T.; Hudson, J.A.; Billington, C.; Carey-Smith, G.V.; Heinemann, J.A. Phage inactivation of foodborne pathogens on cooked and raw meat. Food Microbiol. 2008, 25, 400–406. [Google Scholar] [CrossRef]
- Thung, T.Y.; Lee, E.; Mahyudin, N.A.; Radzi, C.W.J.; Mazlan, N.; Tan, C.W.; Radu, S. Partial characterization and in vitro evaluation of a lytic bacteriophage for biocontrol of Campylobacter jejuni in mutton and chicken meat. J. Food Saf. 2020, 40, e12770. [Google Scholar] [CrossRef]
- Siringan, P.; Connerton, P.L.; Payne, R.J.H.; Connerton, I.F. Bacteriophage-mediated dispersal of Campylobacter jejuni biofilms. Appl. Environ. Microbiol. 2011, 77, 3320–3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.P.; Higgins, S.E.; Guenther, K.L.; Huff, W.; Donoghue, A.M.; Donoghue, D.J.; Hargis, B.M. Use of a Specific Bacteriophage Treatment to Reduce Salmonella in Poultry Products. Poult. Sci. 2005, 84, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Spricigo, D.A.; Bardina, C.; Cortés, P.; Llagostera, M. Use of a Bacteriophage Cocktail to Control Salmonella in Food and the Food Industry. Int. J. Food Microbiol. 2013, 165, 169–174. [Google Scholar] [CrossRef]
- Thung, T.Y.; Krishanthi Jayarukshi Kumari Premarathne, J.M.; San Chang, W.; Loo, Y.Y.; Chin, Y.Z.; Kuan, C.H.; Tan, C.W.; Basri, D.F.; Radzi, C.W.J.W.M.; Radu, S. Use of a Lytic Bacteriophage to Control Salmonella Enteritidis in Retail Food. LWT 2017, 78, 222–225. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, P.; Zhang, H.; Zhou, Y.; Zhang, L.; Wang, R. Bio-Control of Salmonella Enteritidis in Foods Using Bacteriophages. Viruses 2015, 7, 4836–4853. [Google Scholar] [CrossRef]
- Sukumaran, A.T.; Nannapaneni, R.; Kiess, A.; Sharma, C.S. Reduction of Salmonella on chicken breast fillets stored under aerobic or modified atmosphere packaging by the application of lytic bacteriophage preparation SalmoFreshTM. Poult. Sci. 2016, 95, 668–675. [Google Scholar] [CrossRef]
- Soffer, N.; Abuladze, T.; Woolston, J.; Li, M.; Hanna, L.F.; Heyse, S.; Charbonneau, D.; Sulakvelidze, A. Bacteriophages safely reduce Salmonella contamination in pet food and raw pet food ingredients. Bacteriophage 2016, 6, e1220347. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.H.; Waite-Cusic, J.; Huang, E. Control of Salmonella in chicken meat using a combination of a commercial bacteriophage and plant-based essential oils. Food Control 2020, 110, 106984. [Google Scholar] [CrossRef]
- Hagens, S.; de Vegt, B.; Peterson, R. Efficacy of a Commercial Phage Cocktail in Reducing Salmonella Contamination on Poultry Products: Laboratory Data and Industrial Trial Data. Meat Muscle Biol. 2018, 2, 156. [Google Scholar] [CrossRef]
- Soni, K.A.; Nannapaneni, R. Bacteriophage significantly reduces Listeria monocytogenes on raw salmon fillet tissue. J. Food Prot. 2010, 73, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Baños, A.; García-López, J.D.; Núñez, C.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Biocontrol of Listeria monocytogenes in fish by enterocin AS-48 and Listeria lytic bacteriophage P100. LWT 2016, 66, 672–677. [Google Scholar] [CrossRef]
- Perera, M.N.; Abuladze, T.; Li, M.; Woolston, J.; Sulakvelidze, A. Bacteriophage cocktail significantly reduces or eliminates Listeria monocytogenes contamination on lettuce, apples, cheese, smoked salmon and frozen foods. Food Microbiol. 2015, 52, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Bigot, B.; Lee, W.J.; McIntyre, L.; Wilson, T.; Hudson, J.A.; Billington, C.; Heinemann, J. Control of Listeria monocytogenes growth in a ready-to-eat poultry product using a bacteriophage. Food Microbiol. 2011, 28, 1448–1452. [Google Scholar] [CrossRef] [PubMed]
- Chibeu, A.; Agius, L.; Gao, A.; Sabour, P.M.; Kropinski, A.M.; Balamurugan, S. Efficacy of bacteriophage LISTEX™P100 combined with chemical antimicrobials in reducing Listeria monocytogenes in cooked turkey and roast beef. Int. J. Food Microbiol. 2013, 167, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Holck, A.; Berg, J. Inhibition of Listeria monocytogenes in cooked ham by virulent bacteriophages and protective cultures. Appl. Environ. Microbiol. 2009, 75, 6944–6946. [Google Scholar] [CrossRef] [Green Version]
- Iacumin, L.; Manzano, M.; Comi, G. Phage inactivation of Listeria monocytogenes on San Daniele dry-cured ham and elimination of biofilms from equipment and working environments. Microorganisms 2016, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, M.G.; Lee, H.S.; Heo, S.; Kwon, M.; Kim, G. Isolation and Characterization of Listeria phages for Control of Growth of Listeria monocytogenes in Milk. Korean J. Food Sci. Anim. Resour. 2017, 37, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Soni, K.A.; Desai, M.; Oladunjove, A.; Skrobot, F.; Nannapaneni, R. Reduction of Listeria monocytogenes in queso fresco cheese by a combination of listericidal and listeriostatic GRAS antimicrobials. Int. J. Food Microbiol. 2012, 155, 82–88. [Google Scholar] [CrossRef]
- Guenther, S.; Loessner, M.J. Bacteriophage biocontrol of Listeria monocytogenes on soft ripened white mold and red- smear cheeses. Bacteriophage 2011, 1, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Leverentz, B.; Conway, W.S.; Camp, M.J.; Janisiewicz, W.J.; Abuladze, T.; Yang, M.; Saftner, R.; Sulakvelidze, A. Biocontrol of Listeria monocytogenes on Fresh-Cut Produce by Treatment with Lytic Bacteriophages and a Bacteriocin. Appl. Environ. Microbiol. 2003, 69, 4519–4526. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.; Viñas, I.; Colàs, P.; Anguera, M.; Usall, J.; Abadias, M. Effectiveness of a bacteriophage in reducing Listeria monocytogenes on fresh-cut fruits and fruit juices. Food Microbiol. 2014, 38, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Byun, K.H.; Han, S.H.; Choi, M.W.; Park, S.H.; Ha, S.D. Isolation, characterization, and application of bacteriophages to reduce and inhibit Listeria monocytogenes in celery and enoki mushroom. Food Control 2022, 135, 108826. [Google Scholar] [CrossRef]
- You, H.J.; Lee, J.H.; Oh, M.; Hong, S.Y.; Kim, D.; Noh, J.; Kim, M.; Kim, B.S. Tackling Vibrio parahaemolyticus in ready-to-eat raw fish flesh slices using lytic phage VPT02 isolated from market oyster. Food Res. Int. 2021, 150, 110779. [Google Scholar] [CrossRef]
- Xia, H.; Yang, H.; Yan, N.; Hou, W.; Wang, H.; Wang, X.; Wang, H.; Zhou, M. Bacteriostatic effects of phage F23s1 and its endolysin on Vibrio parahaemolyticus. J. Basic Microbiol. 2022, 62, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.J.; Hong, J.; Lee, N.; Chun, H.S.; Kim, H.Y.; Choi, S.W.; Ok, G. Growth inhibitory effect of bacteriophages isolated from western and southern coastal areas of Korea against Vibrio parahaemolyticus in Manila clams. Appl. Biol. Chem. 2016, 59, 359–365. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Z.; Zhou, Y.; Bao, H.; Wang, R.; Li, T.; Pang, M.; Sun, L.; Zhou, X. Application of a Phage in Decontaminating Vibrio Parahaemolyticus in Oysters. Int. J. Food Microbiol. 2018, 275, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Pelon, W.; Luftig, R.B.; Johnston, K.H. Vibrio vulnificus load reduction in oysters after combined exposure to Vibrio vulnificus--specific bacteriophage and to an oyster extract component. J. Food Prot. 2005, 68, 1188–1191. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, Y.T.; Kim, H.B.; Choi, S.H.; Lee, J.H. Characterization of bacteriophage VVP001 and its application for the inhibition of V. vulnificus causing seafood-borne diseases. Food Microbiol. 2021, 94, 103630. [Google Scholar] [CrossRef]
- Aguilera, M.; Martínez, S.; Tello, M.; Gallardo, M.J.; García, V. Use of Cocktail of Bacteriophage for Salmonella Typhimurium Control in Chicken Meat. Foods 2022, 11, 1164. [Google Scholar] [CrossRef]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial Biofilms in the Food Industry—A Comprehensive Review. Int. J. Environ. Res. Public Health 2021, 18, 14. [Google Scholar] [CrossRef]
- Korzeniowski, P.; Śliwka, P.; Kuczkowski, M.; Mišić, D.; Milcarz, A.; Kuźmińska-Bajor, M. Bacteriophage cocktail can effectively control Salmonella biofilm in poultry housing. Front. Microbiol. 2022, 13, 2317. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Vermeulen, A.; Gysemans, K.P.M.; Bernaerts, K.; Geeraerd, A.H.; Van Impe, J.F.; Debevere, J.; Devlieghere, F. Influence of pH, water activity and acetic acid concentration on Listeria monocytogenes at 7 °C: Data collection for the development of a growth/no growth model. Int. J. Food Microbiol. 2007, 114, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, V.B.; Mujahid, S.; Orsi, R.H.; Bergholz, T.M.; Wiedmann, M.; Boor, K.; Destro, M. Contributions of σB and PrfA to Listeria monocytogenes salt stress under food relevant conditions. Int. J. Food Microbiol. 2014, 177, 98–108. [Google Scholar] [CrossRef]
- Makariti, I.P.; Grivokostopoulos, N.C.; Skandamis, P.N. Effect οf οxygen availability and pH οn adaptive acid tolerance response of immobilized Listeria monocytogenes in structured growth media. Food Microbiol. 2021, 99, 103826. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Langsrud, S.; Møretrø, T. Microbial diversity and ecology of biofilms in food industry environments associated with Listeria monocytogenes persistence. Curr. Opin. Food Sci. 2021, 37, 171–178. [Google Scholar] [CrossRef]
- Forauer, E.; Tongyu Wu, S.; Etter, A.J. Listeria monocytogenes in the retail deli environment: A review. Food Control 2021, 119, 107443. [Google Scholar] [CrossRef]
- Belias, A.; Sullivan, G.; Wiedmann, M.; Ivanek, R. Factors that contribute to persistent Listeria in food processing facilities and relevant interventions: A rapid review. Food Control 2022, 133, 108579. [Google Scholar] [CrossRef]
- Carlton, R.M.; Noordman, W.H.; Biswas, B.; De Meester, E.D.; Loessner, M.J. Bacteriophage P100 for control of Listeria monocytogenes in foods: Genome sequence, bioinformatic analyses, oral toxicity study and application. Regul. Toxicol. Pharmacol. 2005, 43, 301–312. [Google Scholar] [CrossRef]
- Schmuki, M.M.; Erne, D.; Loessner, M.J.; Klumpp, J. Bacteriophage P70: Unique morphology and unrelatedness to other Listeria bacteriophages. J. Virol. 2012, 86, 13099–13102. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Zhu, M.; Wang, Y.; Yang, Z.; Ye, M.; Wu, L.; Bao, H.; Pang, M.; Zhou, Y.; Wang, R.; et al. Broad host range phage vBLmoM-SH3-3 reduces the risk of Listeria contamination in two types of ready-to-eat food. Food Control 2020, 108, 106830. [Google Scholar] [CrossRef]
- Kawacka, I.; Olejnik-Schmidt, A.; Schmidt, M.; Sip, A. Effectiveness of phage-based inhibition of Listeria monocytogenes in food products and food processing environments. Microorganisms 2020, 8, 1764. [Google Scholar] [CrossRef] [PubMed]
- EFSA BIOHAZ Panel. Scientific opinion on the evaluation of the safety and efficacy of ListexTM P100 for reduction of pathogens on different ready-to-eat (RTE) food products. EFSA J. 2016, 14, e04565. [Google Scholar] [CrossRef]
- Gray, J.A.; Chandry, P.S.; Kaur, M.; Kocharunchitt, C.; Bowman, J.P.; Fox, E.M. Novel Biocontrol Methods for Listeria monocytogenes Biofilms in Food Production Facilities. Front. Microbiol. 2018, 9, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brives, C.; Pourraz, J. Phage therapy as a potential solution in the fight against AMR: Obstacles and possible futures. Palgrave Commun. 2020, 6, 100. [Google Scholar] [CrossRef]
- Ferriol-González, C.; Domingo-Calap, P. Phages for Biofilm Removal. Antibiotics 2020, 9, 268. [Google Scholar] [CrossRef]
- Mateus, L.; Costa, L.; Silva, Y.J.; Pereira, C.; Cunha, A.; Almeida, A. Efficiency of phage cocktails in the inactivation of Vibrio in aquaculture. Aquaculture 2014, 424–425, 167–173. [Google Scholar] [CrossRef]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef] [Green Version]
- Wandro, S.; Ghatbale, P.; Attai, H.; Hendrickson, C.; Samillano, C.; Suh, J.; Pride, D.T.; Whiteson, K. Phage cocktails can prevent the evolution of phage-resistant Enterococcus. BioRxiv 2021. [Google Scholar] [CrossRef]
- Kaur, S.; Kumari, A.; Kumari Negi, A.; Galav, V.; Thakur, S.; Agrawal, M.; Sharma, V. Nanotechnology Based Approaches in Phage Therapy: Overcoming the Pharmacological Barriers. Front. Pharmacol. 2021, 12, 699054. [Google Scholar] [CrossRef]
- Loh, B.; Gondil, V.S.; Manohar, P.; Khan, F.M.; Yang, H.; Leptihn, S. Encapsulation and delivery of therapeutic phages. Appl. Environ. Microbiol 2020, 87, e01979-20. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.A.; Liu, H.; Wang, Q.; Zhong, F.; Guo, Q.; Balamurugan, S. Use of encapsulated bacteriophages to enhance farm to fork food safety. Crit. Rev. Food Sci. 2017, 57, 2801–2810. [Google Scholar] [CrossRef] [PubMed]
- Danis-Wlodarczyk, K.; Dąbrowska, K.; Abedon, S.T. Phage Therapy: The Pharmacology of Antibacterial Viruses. Curr. Issues Mol. Biol. 2021, 40, 81–164. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rubio, L.; Jofre, J.; Muniesa, M. Is Genetic Mobilization Considered When Using Bacteriophages in Antimicrobial Therapy? Antibiotics 2017, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Abedon, S.T.; Danis-Wlodarczyk, K.M.; Wozniak, D.J. Phage Cocktail Development for Bacteriophage Therapy: Toward Improving Spectrum of Activity Breadth and Depth. Pharmaceuticals 2021, 14, 1019. [Google Scholar] [CrossRef] [PubMed]
- Nagel, T.; Musila, L.; Muthoni, M.; Nikolich, M.; Nakavuma, J.L.; Clokie, M.R. Phage banks as potential tools to rapidly and cost-effectively manage antimicrobial resistance in the developing world. Curr. Opin. Virol. 2022, 53, 101208. [Google Scholar] [CrossRef]
- Domingo-Calap, P.; Delgado-Martínez, J. Bacteriophages: Protagonists of a Post-Antibiotic Era. Antibiotics 2018, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Leptihn, S.; Loh, B. Complexity, challenges and costs of implementing phage therapy. Future Microbiol. 2022, 17, 643–646. [Google Scholar] [CrossRef]
- Vázquez, R.; Díez-Martínez, R.; Domingo-Calap, P.; García, P.; Gutiérrez, D.; Muniesa, M.; Ruiz-Ruigómez, M.; Sanjuán, R.; Tomás, M.; Tormo-Mas, M.; et al. Essential topics for the regulatory consideration of phages as clinically valuable therapeutic agents: A perspective from Spain. Microorganisms 2022, 10, 717. [Google Scholar] [CrossRef]


| Animal | Bacteria Load 1 | Phage | Application Method and Dose 2 | Bacterial Reduction | Ref. |
|---|---|---|---|---|---|
| Mussels | V. parahaemolyticus, naturally infected | Phage cocktail: VP10 | Immersion ~103 | Undetectable levels in seawater, sediment, or mussels after 48 h | [79] |
| Oysters | V. parahaemolyticus CRS 09-17, AMR * 2.7 × 106 | pVp-1 | Immersion 1.6 × 107 | Growth reduction >5 log after 72 h | [80] |
| Aquatic crustacean Artemia franciscana | V. parahaemolyticus FORC_023 104 | VPG01 | Immersion 103, 104, 105 | 2 log reduction Increased survival from 10% to 40% with higher phage concentration (104 and 105 PFU/mL). | [81] |
| Shrimps (Penaeus monodon) larvae | V. harveyi 105 | Bacteriophage of V. harveyi | Immersion 2 × 105 | 3 log reduction in bacterial counts Increased larvae survival from 17% to 86% More effective than antibiotics (40% survival) | [82] |
| Gilthead seabream larvae | V. harveyi VH2 106 | Virtus | Immersion 107 | Increased survival of larvae: from <6% to >40% | [83] |
| Atlantic salmon | V. anguillarum PF4 5 × 105 | CHOED | Immersion (100L) 5 × 105 Immersion (100L) 106 Immersion (farm conditions) 5 × 107 | Increased survival from 5% to 70% after 10 d Increased survival from 5% to 100% after 10 d Increased survival from 65% to 100% after 9 d (protection up to 20 d) | [84] |
| Zebrafish larvae | V. anguillarum 106 | VP-2 phage | Immersion 108 | Increased survival from 83% to 98% after 72 h | [85] |
| Whiteleg shrimp larvae | V. parahaemolyticus ATCC 17802 2 × 106 | A3S Vpms1 | Immersion 105, 106,107 | Increased survival of larvae from <60% to 80% (A3S phage), depending on the conditions | [86] |
| Shrimps (Penaeus monodon) | V. harveyi | Viha10 Viha8 | Immersion 2 × 106 | Increased survival of larvae to >86% vs. ~65% survival with antibiotics | [87] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavilla, M.; Domingo-Calap, P.; Sevilla-Navarro, S.; Lasagabaster, A. Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective. Foods 2023, 12, 552. https://doi.org/10.3390/foods12030552
Lavilla M, Domingo-Calap P, Sevilla-Navarro S, Lasagabaster A. Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective. Foods. 2023; 12(3):552. https://doi.org/10.3390/foods12030552
Chicago/Turabian StyleLavilla, Maria, Pilar Domingo-Calap, Sandra Sevilla-Navarro, and Amaia Lasagabaster. 2023. "Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective" Foods 12, no. 3: 552. https://doi.org/10.3390/foods12030552