
Comparative Genomic Analysis Reveals the Functional Traits and Safety Status of Lactic Acid Bacteria Retrieved from Artisanal Cheeses and Raw Sheep Milk

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains Used in the Study
2.2. Whole Genome Sequencing, Assembly, and Quality Control
2.3. In Silico Typing and Comparative Genomic Analysis
2.4. Statistical Analysis
3. Results and Discussion
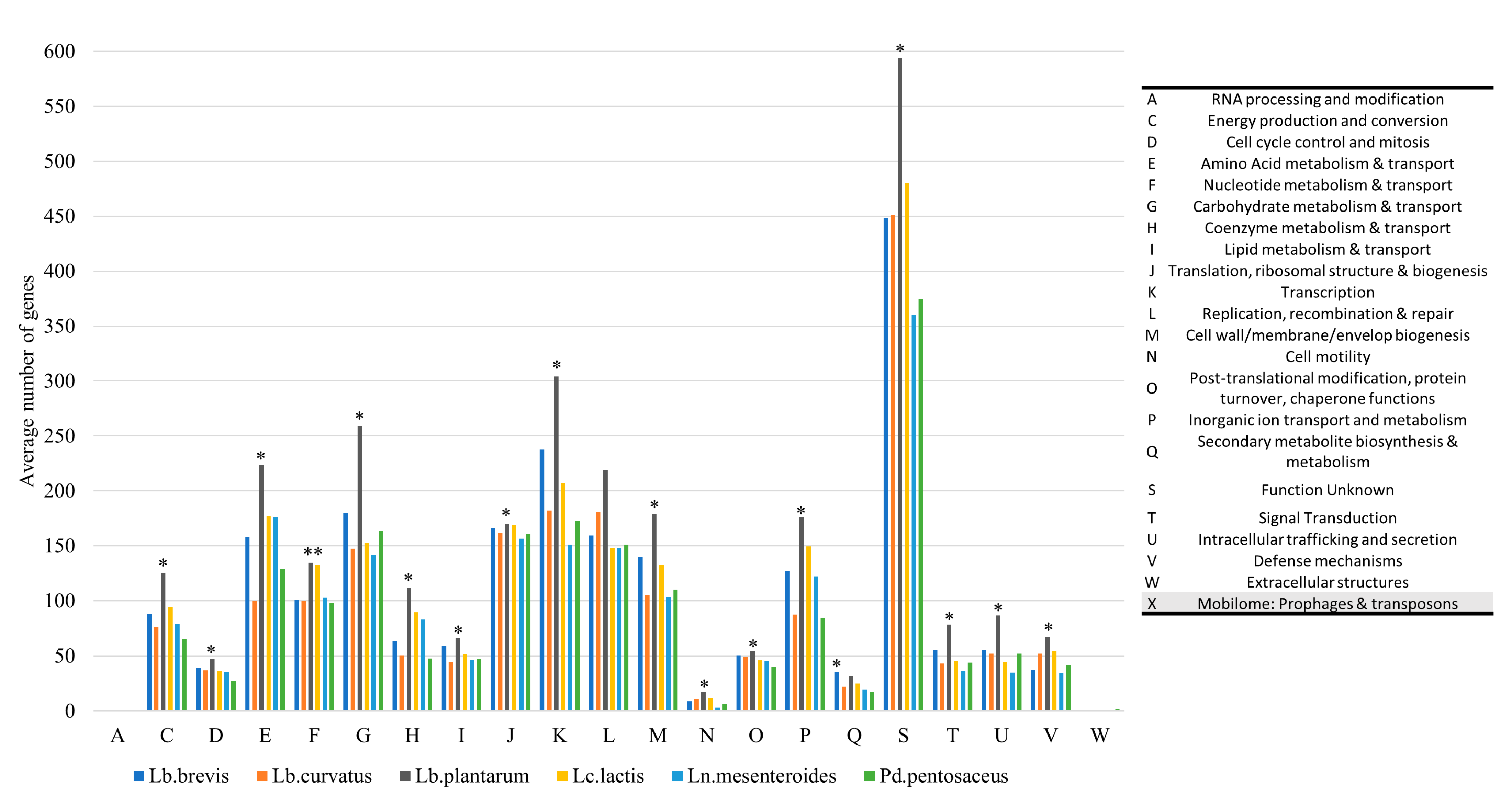
3.1. Assembly Statistics and Subsystem Analysis
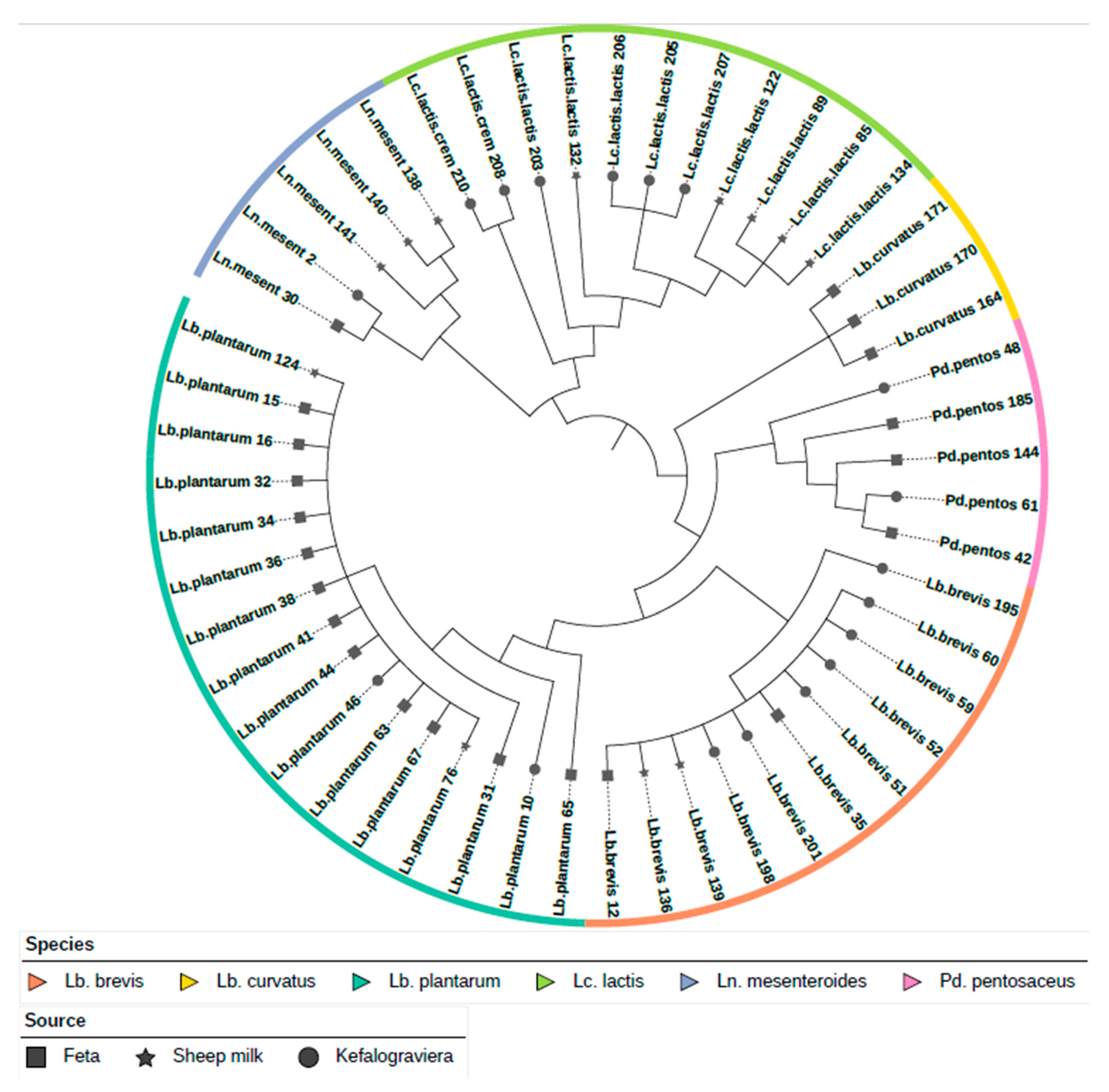
3.2. Phylogenetic Analysis and Assessment of RGs and VGs
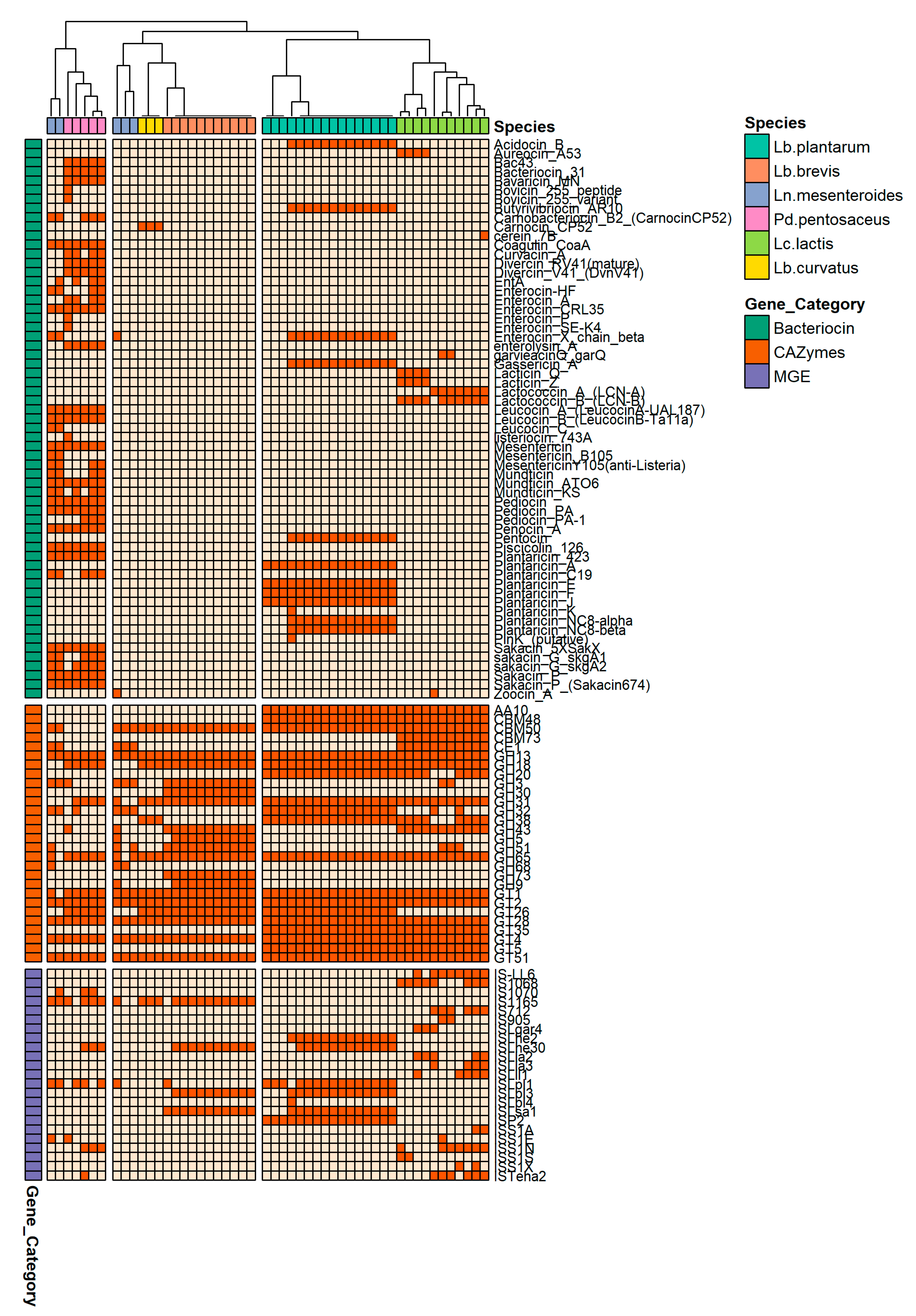
3.3. Comparative Genomics
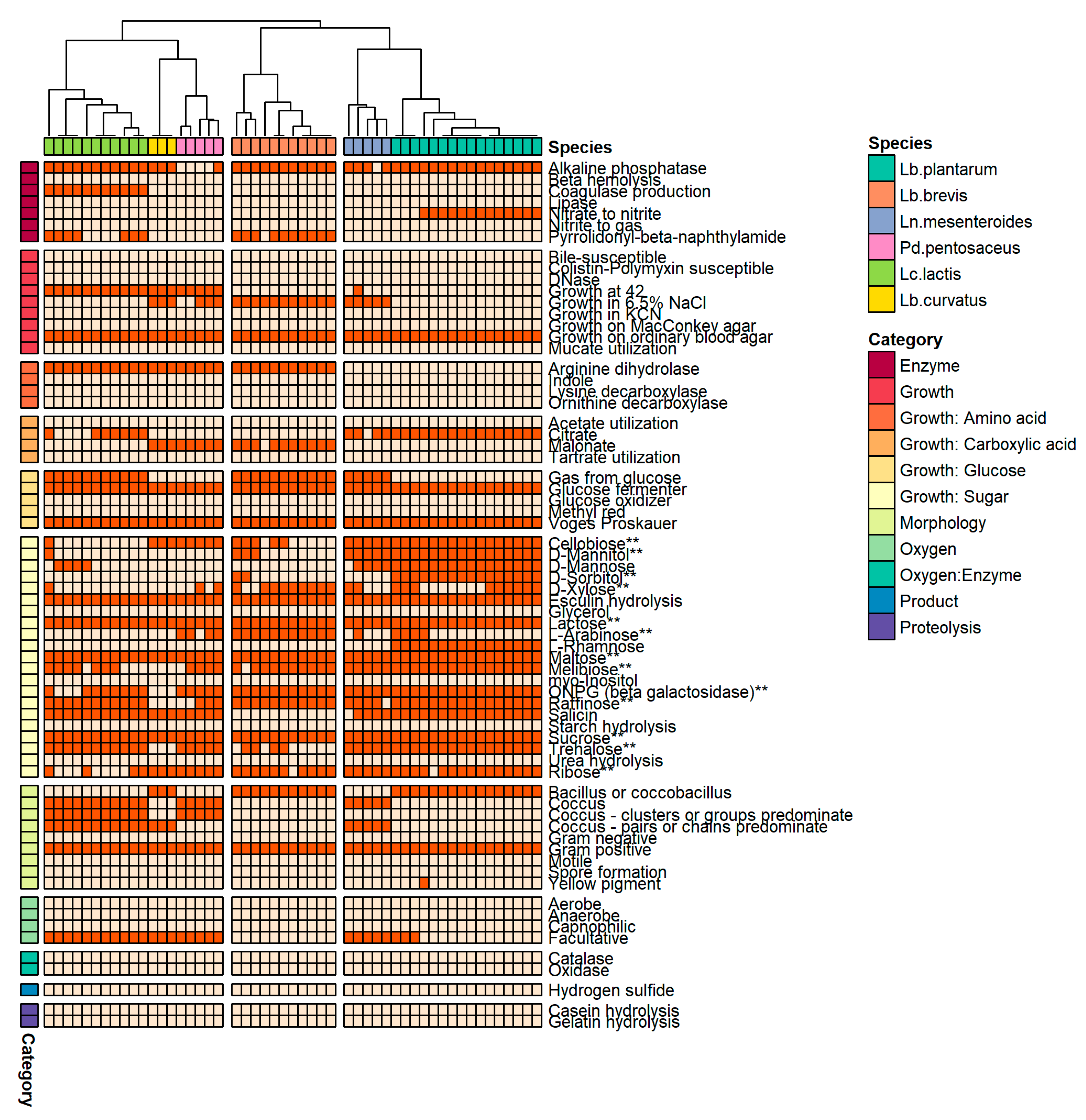
3.4. Analysis of Phenotypic Traits
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cano-Garrido, O.; Seras-Franzoso, J.; Garcia-Fruitós, E. Lactic Acid Bacteria: Reviewing the Potential of a Promising Delivery Live Vector for Biomedical Purposes. Microb. Cell Fact. 2015, 14, 137. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Harris, H.M.B.; McCann, A.; Guo, C.; Argimón, S.; Zhang, W.; Yang, X.; Jeffery, I.B.; Cooney, J.C.; Kagawa, T.F.; et al. Expanding the Biotechnology Potential of Lactobacilli through Comparative Genomics of 213 Strains and Associated Genera. Nat. Commun. 2015, 6, 8322. [Google Scholar] [CrossRef] [PubMed]
- Mayo, B.; Rodríguez, J.; Vázquez, L.; Flórez, A.B. Microbial Interactions within the Cheese Ecosystem and Their Application to Improve Quality and Safety. Foods 2021, 10, 602. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, P.; Murphy, J.; Mahony, J.; van Sinderen, D. Next-Generation Sequencing as an Approach to Dairy Starter Selection. Dairy Sci. Technol. 2015, 95, 545–568. [Google Scholar] [CrossRef]
- Tofalo, R.; Perpetuini, G.; Battistelli, N.; Pepe, A.; Ianni, A.; Martino, G.; Suzzi, G. Accumulation γ-Aminobutyric Acid and Biogenic Amines in a Traditional Raw Milk Ewe’s Cheese. Foods 2019, 8, 401. [Google Scholar] [CrossRef]
- Tofalo, R.; Schirone, M.; Fasoli, G.; Perpetuini, G.; Patrignani, F.; Manetta, A.C.; Lanciotti, R.; Corsetti, A.; Martino, G.; Suzzi, G. Influence of Pig Rennet on Proteolysis, Organic Acids Content and Microbiota of Pecorino Di Farindola, a Traditional Italian Ewe’s Raw Milk Cheese. Food Chem. 2015, 175, 121–127. [Google Scholar] [CrossRef]
- Buron-Moles, G.; Chailyan, A.; Dolejs, I.; Forster, J.; Mikš, M.H. Uncovering Carbohydrate Metabolism through a Genotype-Phenotype Association Study of 56 Lactic Acid Bacteria Genomes. Appl. Microbiol. Biotechnol. 2019, 103, 3135–3152. [Google Scholar] [CrossRef]
- Sałański, P.; Kowalczyk, M.; Bardowski, J.K.; Szczepankowska, A.K. Health-Promoting Nature of Lactococcus lactis IBB109 and Lactococcus lactis IBB417 Strains Exhibiting Proliferation Inhibition and Stimulation of Interleukin-18 Expression in Colorectal Cancer Cells. Front. Microbiol. 2022, 13, 822912. [Google Scholar] [CrossRef]
- Pightling, A.W.; Pettengill, J.B.; Luo, Y.; Baugher, J.D.; Rand, H.; Strain, E. Interpreting Whole-Genome Sequence Analyses of Foodborne Bacteria for Regulatory Applications and Outbreak Investigations. Front. Microbiol. 2018, 9, 01482. [Google Scholar] [CrossRef] [Green Version]
- Bull, M.J.; Jolley, K.A.; Bray, J.E.; Aerts, M.; Vandamme, P.; Maiden, M.C.J.; Marchesi, J.R.; Mahenthiralingam, E. The Domestication of the Probiotic Bacterium Lactobacillus acidophilus. Sci. Rep. 2015, 4, 7202. [Google Scholar] [CrossRef]
- Yu, J.; Sun, Z.; Liu, W.; Bao, Q.; Zhang, J.; Zhang, H. Phylogenetic Study of Lactobacillus acidophilus Group, L. casei Group and L. plantarum Group Based on Partial hsp60, pheS and tuf Gene Sequences. Eur. Food Res. Technol. 2012, 234, 927–934. [Google Scholar] [CrossRef]
- Tsigkrimani, M.; Bakogianni, M.; Paramithiotis, S.; Bosnea, L.; Pappa, E.; Drosinos, E.H.; Skandamis, P.N.; Mataragas, M. Microbial Ecology of Artisanal Feta and Kefalograviera Cheeses, Part I: Bacterial Community and Its Functional Characteristics with Focus on Lactic Acid Bacteria as Determined by Culture-Dependent Methods and Phenotype Microarrays. Microorganisms 2022, 10, 161. [Google Scholar] [CrossRef]
- Tsigkrimani, M.; Panagiotarea, K.; Paramithiotis, S.; Bosnea, L.; Pappa, E.; Drosinos, E.H.; Skandamis, P.N.; Mataragas, M. Microbial Ecology of Sheep Milk, Artisanal Feta, and Kefalograviera Cheeses. Part II: Technological, Safety, and Probiotic Attributes of Lactic Acid Bacteria Isolates. Foods 2022, 11, 459. [Google Scholar] [CrossRef]
- Syrokou, M.K.; Themeli, C.; Paramithiotis, S.; Mataragas, M.; Bosnea, L.; Argyri, A.A.; Chorianopoulos, N.G.; Skandamis, P.N.; Drosinos, E.H. Microbial Ecology of Greek Wheat Sourdoughs, Identified by a Culture-Dependent and a Culture-Independent Approach. Foods 2020, 9, 1603. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2019. [Google Scholar]
- Arkin, A.P.; Cottingham, R.W.; Henry, C.S.; Harris, N.L.; Stevens, R.L.; Maslov, S.; Dehal, P.; Ware, D.; Perez, F.; Canon, S.; et al. KBase: The United States Department of Energy Systems Biology Knowledgebase. Nat. Biotechnol. 2018, 36, 566–569. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing Bacterial Genome Assemblies with Multiplex MinION Sequencing. Microb. Genomics 2017, 3. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef]
- Bosi, E.; Donati, B.; Galardini, M.; Brunetti, S.; Sagot, M.F.; Lió, P.; Crescenzi, P.; Fani, R.; Fondi, M. MeDuSa: A Multi-Draft Based Scaffolder. Bioinformatics 2015, 31, 2443–2451. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Salzberg, S.L. SkewIT: The Skew Index Test for Large-Scale GC Skew Analysis of Bacterial Genomes. PLoS Comput. Biol. 2020, 16, e1008439. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Lu, J.; Langmead, B. Improved Metagenomic Analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS Is an Automated High-Throughput Platform for State-of-the-Art Genome-Based Taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Smirnov, S.; Nikolskaya, A.N.; et al. The COG Database: An Updated Vesion Includes Eukaryotes. BMC Bioinform. 2003, 4, 1–14. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. EggNOG-Mapper v2: Functional Annotation, Orthology Assignments, and Domain Prediction at the Metagenomic Scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an Update of CRISRFinder, Includes a Portable Version, Enhanced Performance and Integrates Search for Cas Proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef]
- Arndt, D.; Marcu, A.; Liang, Y.; Wishart, D.S. PHAST, PHASTER and PHASTEST: Tools for Finding Prophage in Bacterial Genomes. Brief. Bioinform. 2019, 20, 1560–1567. [Google Scholar] [CrossRef]
- Seemann, T. Abricate; Github: San Francisco, CA, USA, 2020. [Google Scholar]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and Refined Dataset for Big Data Analysis—10 Years On. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of Mobile Genetic Elements Associated with Antibiotic Resistance in Salmonella enterica Using a Newly Developed Web Tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garciá-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids Using Plasmidfinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. DbCAN2: A Meta Server for Automated Carbohydrate-Active Enzyme Annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef]
- Weimann, A.; Mooren, K.; Frank, J.; Pope, P.B.; Bremges, A.; McHardy, A.C. From Genomes to Phenotypes: Traitar, the Microbial Trait Analyzer. mSystems 2016, 1. [Google Scholar] [CrossRef]
- Brynildsrud, O.; Bohlin, J.; Scheffer, L.; Eldholm, V. Rapid Scoring of Genes in Microbial Pan-Genome-Wide Association Studies with Scoary. Genome Biol. 2016, 17, 238. [Google Scholar] [CrossRef]
- Reijnders, M.J.M.F.; Waterhouse, R.M. Summary Visualizations of Gene Ontology Terms with GO-Figure! Front. Bioinform. 2021, 1, 638255. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles Instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Overbeek, R.; Begley, T.; Butler, R.M.; Choudhuri, J.V.; Chuang, H.Y.; Cohoon, M.; de Crécy-Lagard, V.; Diaz, N.; Disz, T.; Edwards, R.; et al. The Subsystems Approach to Genome Annotation and Its Use in the Project to Annotate 1000 Genomes. Nucleic Acids Res. 2005, 33, 5691–5702. [Google Scholar] [CrossRef] [PubMed]
- Tenea, G.N.; Ortega, C. Genome Characterization of Lactiplantibacillus Plantarum Strain UTNGt2 Originated from Theobroma grandiflorum (White Cacao) of Ecuadorian Amazon: Antimicrobial Peptides from Safety to Potential Applications. Antibiotics 2021, 10, 383. [Google Scholar] [CrossRef] [PubMed]
- Flórez, A.B.; de Los Reyes-Gavilán, C.G.; Wind, A.; Mayo, B.; Margolles, A. Ubiquity and Diversity of Multidrug Resistance Genes in Lactococcus lactis Strains Isolated between 1936 and 1995. FEMS Microbiol. Lett. 2006, 263, 21–25. [Google Scholar] [CrossRef]
- Almeida, M.; Hébert, A.; Abraham, A.L.; Rasmussen, S.; Monnet, C.; Pons, N.; Delbès, C.; Loux, V.; Batto, J.M.; Leonard, P.; et al. Construction of a Dairy Microbial Genome Catalog Opens New Perspectives for the Metagenomic Analysis of Dairy Fermented Products. BMC Genomics 2014, 15. [Google Scholar] [CrossRef]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Henning, C.; Gautam, D.; Muriana, P. Identification of Multiple Bacteriocins in Enterococcus Spp. Using an Enterococcus-Specific Bacteriocin PCR Array. Microorganisms 2015, 3, 1–16. [Google Scholar] [CrossRef]
- Franz, C.M.A.P.; Van Belkum, M.J.; Holzapfel, W.H.; Abriouel, H.; Gálvez, A. Diversity of Enterococcal Bacteriocins and Their Grouping in a New Classification Scheme. FEMS Microbiol. Rev. 2007, 31, 293–310. [Google Scholar] [CrossRef]
- Todokoro, D.; Tomita, H.; Inoue, T.; Ike, Y. Genetic Analysis of Bacteriocin 43 of Vancomycin-Resistant Enterococcus faecium. Appl. Environ. Microbiol. 2006, 72, 6955–6964. [Google Scholar] [CrossRef] [Green Version]
- Abdulhussain Kareem, R.; Razavi, S.H. Plantaricin Bacteriocins: As Safe Alternative Antimicrobial Peptides in Food Preservation—A Review. J. Food Saf. 2020, 40. [Google Scholar] [CrossRef]
- Leer, R.J.; van der Vossen, J.M.B.M.; van Giezen, M.; van Noort Johannes, M.; Pouwels, P.H. Genetic Analysis of Acidocin B, a Novel Bacteriocin Produced by Lactobacillus acidophilus. Microbiology 1995, 141, 1629–1635. [Google Scholar] [CrossRef]
- Hammami, R.; Zouhir, A.; Le Lay, C.; Ben Hamida, J.; Fliss, I. BACTIBASE Second Release: A Database and Tool Platform for Bacteriocin Characterization. BMC Microbiol. 2010, 10, 22. [Google Scholar] [CrossRef]
- Markusková, B.; Lichvariková, A.; Szemes, T.; Koreňová, J.; Kuchta, T.; Drahovská, H. Genome Analysis of Lactic Acid Bacterial Strains Selected as Potential Starters for Traditional Slovakian Bryndza Cheese. FEMS Microbiol. Lett. 2018. [Google Scholar] [CrossRef]
- Tarrah, A.; Pakroo, S.; Lemos Junior, W.J.F.; Guerra, A.F.; Corich, V.; Giacomini, A. Complete Genome Sequence and Carbohydrates-Active EnZymes (CAZymes) Analysis of Lactobacillus paracasei DTA72, a Potential Probiotic Strain with Strong Capability to Use Inulin. Curr. Microbiol. 2020, 77, 2867–2875. [Google Scholar] [CrossRef]
- Shen, J.; Zheng, L.; Chen, X.; Han, X.; Cao, Y.; Yao, J. Metagenomic Analyses of Microbial and Carbohydrate-Active Enzymes in the Rumen of Dairy Goats Fed Different Rumen Degradable Starch. Front. Microbiol. 2020, 11, 1003. [Google Scholar] [CrossRef]
- He, Q.; Hou, Q.; Wang, Y.; Li, J.; Li, W.; Kwok, L.-Y.; Sun, Z.; Zhang, H.; Zhong, Z. Comparative Genomic Analysis of Enterococcus faecalis: Insights into Their Environmental Adaptations. BMC Genomics 2018, 19, 527. [Google Scholar] [CrossRef]
- Ghattargi, V.C.; Gaikwad, M.A.; Meti, B.S.; Nimonkar, Y.S.; Dixit, K.; Prakash, O.; Shouche, Y.S.; Pawar, S.P.; Dhotre, D.P. Comparative Genome Analysis Reveals Key Genetic Factors Associated with Probiotic Property in Enterococcus faecium Strains. BMC Genomics 2018, 19, 652. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, Z.; Menghe, B.; Zhang, H. Short Communication: Single Molecule, Real-Time Sequencing Technology Revealed Species- and Strain-Specific Methylation Patterns of 2 Lactobacillus Strains. J. Dairy Sci. 2015, 98, 3020–3024. [Google Scholar] [CrossRef]
- Chaillou, S.; Champomier-Vergès, M.-C.; Cornet, M.; Crutz-Le Coq, A.-M.; Dudez, A.-M.; Martin, V.; Beaufils, S.; Darbon-Rongère, E.; Bossy, R.; Loux, V.; et al. The Complete Genome Sequence of the Meat-Borne Lactic Acid Bacterium Lactobacillus sakei 23K. Nat. Biotechnol. 2005, 23, 1527–1533. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, S.; Stockdale, S.; Bottacini, F.; Mahony, J.; van Sinderen, D. The Lactococcus lactis Plasmidome: Much Learnt, yet Still Lots to Discover. FEMS Microbiol. Rev. 2014, 38, 1066–1088. [Google Scholar] [CrossRef] [PubMed]
- Apostolakos, I.; Paramithiotis, S.; Mataragas, M. Functional and Safety Characterization of Weissella paramesenteroides Strains Isolated from Dairy Products through Whole-Genome Sequencing and Comparative Genomics. Dairy 2022, 3, 799–813. [Google Scholar] [CrossRef]
- Durrant, M.G.; Li, M.M.; Siranosian, B.A.; Montgomery, S.B.; Bhatt, A.S. A Bioinformatic Analysis of Integrative Mobile Genetic Elements Highlights Their Role in Bacterial Adaptation. Cell Host Microbe 2020, 27, 140–153.e9. [Google Scholar] [CrossRef] [PubMed]
- Kiousi, D.E.; Efstathiou, C.; Tegopoulos, K.; Mantzourani, I.; Alexopoulos, A.; Plessas, S.; Kolovos, P.; Koffa, M.; Galanis, A. Genomic Insight Into Lacticaseibacillus paracasei SP5, Reveals Genes and Gene Clusters of Probiotic Interest and Biotechnological Potential. Front. Microbiol. 2022, 13, 2038. [Google Scholar] [CrossRef]
- Liu, D.; Liu, Y.; Zhang, D.; Chen, X.; Liu, Q.; Xiong, B.; Zhang, L.; Wei, L.; Wang, Y.; Fang, H.; et al. Quantitative Proteome Profiling Reveals Cellobiose-Dependent Protein Processing and Export Pathways for the Lignocellulolytic Response in Neurospora crassa. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Allen, J.R.; Torres-Acosta, M.A.; Mohan, N.; Lye, G.J.; Ward, J.M. Segregationally Stabilised Plasmids Improve Production of Commodity Chemicals in Glucose-Limited Continuous Fermentation. Microb. Cell Fact. 2022, 21, 229. [Google Scholar] [CrossRef]
- Krewinkel, M.; Gosch, M.; Rentschler, E.; Fischer, L. Epilactose Production by 2 Cellobiose 2-Epimerases in Natural Milk. J. Dairy Sci. 2014, 97, 155–161. [Google Scholar] [CrossRef]
- Van Trijp, M.P.H.; Schutte, S.; Esser, D.; Wopereis, S.; Hoevenaars, F.P.M.; Hooiveld, G.J.E.J.; Afman, L.A. Minor Changes in the Composition and Function of the Gut Microbiota During a 12-Week Whole Grain Wheat or Refined Wheat Intervention Correlate with Liver Fat in Overweight and Obese Adults. J. Nutr. 2021, 151, 491–502. [Google Scholar] [CrossRef]
- Lai, C.-Y.; Cronan, J.E. β-Ketoacyl-Acyl Carrier Protein Synthase III (FabH) Is Essential for Bacterial Fatty Acid Synthesis. J. Biol. Chem. 2003, 278, 51494–51503. [Google Scholar] [CrossRef]
- McLeod, A.; Snipen, L.; Naterstad, K.; Axelsson, L. Global Transcriptome Response in Lactobacillus sakei during Growth on Ribose. BMC Microbiol. 2011, 11, 145. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Fukumori, F.; Nishiwaki, H.; Sakurai, Y.; Tajima, K.; Watanabe, Y. Novel Non-Phosphorylative Pathway of Pentose Metabolism from Bacteria. Sci. Rep. 2019, 9, 155. [Google Scholar] [CrossRef]
- Okano, K.; Yoshida, S.; Tanaka, T.; Ogino, C.; Fukuda, H.; Kondo, A. Homo-D-Lactic Acid Fermentation from Arabinose by Redirection of the Phosphoketolase Pathway to the Pentose Phosphate Pathway in L-Lactate Dehydrogenase Gene-Deficient Lactobacillus plantarum. Appl. Environ. Microbiol. 2009, 75, 5175–5178. [Google Scholar] [CrossRef]
- Welman, A.D.; Maddox, I.S. Exopolysaccharides from Lactic Acid Bacteria: Perspectives and Challenges. Trends Biotechnol. 2003, 21, 269–274. [Google Scholar] [CrossRef]
- Angelin, J.; Kavitha, M. Exopolysaccharides from Probiotic Bacteria and Their Health Potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef]
- Yilmaz, B.; Bangar, S.P.; Echegaray, N.; Suri, S.; Tomasevic, I.; Manuel Lorenzo, J.; Melekoglu, E.; Rocha, J.M.; Ozogul, F. The Impacts of Lactiplantibacillus plantarum on the Functional Properties of Fermented Foods: A Review of Current Knowledge. Microorganisms 2022, 10, 826. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Scientific Opinion on the Update of the List of QPS-recommended Biological Agents Intentionally Added to Food or Feed as Notified to EFSA (2017–2019). EFSA J. 2020, 18. [Google Scholar] [CrossRef]
- Teixeira, C.G.; da Silva, R.R.; Fusieger, A.; Martins, E.; de Freitas, R.; de Carvalho, A.F. Weissella Na Indústria de Alimentos: Uma Revisão. Res. Soc. Dev. 2021, 10, e8310514557. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Microorganism | Source | Genome Size (Mb) | GC Content (%) | No. of Scaffolds | N50 (Mb) | No. of CDSs |
|---|---|---|---|---|---|---|---|
| DRD-10 | Lb. plantarum | Kefalograviera | 1.7 | 38.0 | 4.0 | 1.7 | 2445 |
| DRD-15 | Lb. plantarum | Feta | 3.0 | 44.9 | 22.0 | 1.3 | 2435 |
| DRD-16 | Lb. plantarum | Feta | 2.5 | 45.8 | 23.0 | 2.3 | 2449 |
| DRD-31 | Lb. plantarum | Feta | 3.5 | 44.2 | 126.0 | 3.2 | 2170 |
| DRD-32 | Lb. plantarum | Feta | 3.5 | 44.2 | 125.0 | 3.1 | 2450 |
| DRD-34 | Lb. plantarum | Feta | 2.0 | 37.6 | 24.0 | 1.0 | 2449 |
| DRD-36 | Lb. plantarum | Feta | 3.0 | 44.9 | 20.0 | 2.3 | 2453 |
| DRD-38 | Lb. plantarum | Feta | 3.4 | 44.2 | 95.0 | 2.9 | 2449 |
| DRD-41 | Lb. plantarum | Feta | 3.5 | 44.2 | 105.0 | 2.1 | 2450 |
| DRD-44 | Lb. plantarum | Feta | 2.5 | 45.8 | 19.0 | 2.3 | 2454 |
| DRD-46 | Lb. plantarum | Kefalograviera | 3.5 | 44.2 | 103.0 | 2.9 | 2450 |
| DRD-63 | Lb. plantarum | Feta | 3.5 | 44.2 | 106.0 | 3.2 | 2143 |
| DRD-65 | Lb. plantarum | Feta | 3.5 | 44.2 | 109.0 | 3.2 | 2147 |
| DRD-67 | Lb. plantarum | Feta | 1.7 | 37.6 | 10.0 | 1.3 | 2147 |
| DRD-76 | Lb. plantarum | Sheep milk | 3.5 | 44.2 | 115.0 | 3.2 | 2741 |
| DRD-124 | Lb. plantarum | Sheep milk | 3.4 | 44.2 | 87.0 | 3.2 | 3213 |
| DRD-208 | Lc. lactis cremoris | Kefalograviera | 1.9 | 37.1 | 14.0 | 1.4 | 3253 |
| DRD-210 | Lc. lactis cremoris | Kefalograviera | 2.5 | 45.8 | 21.0 | 2.3 | 3254 |
| DRD-85 | Lc. lactis lactis | Sheep milk | 2.5 | 45.8 | 22.0 | 2.3 | 2753 |
| DRD-89 | Lc. lactis lactis | Sheep milk | 2.5 | 45.8 | 21.0 | 2.3 | 3232 |
| DRD-122 | Lc. lactis lactis | Sheep milk | 2.5 | 45.8 | 22.0 | 2.3 | 3253 |
| DRD-132 | Lc. lactis lactis | Sheep milk | 1.9 | 37.1 | 9.0 | 1.9 | 3264 |
| DRD-134 | Lc. lactis lactis | Sheep milk | 3.6 | 44.1 | 227.0 | 3.1 | 3260 |
| DRD-203 | Lc. lactis lactis | Kefalograviera | 3.0 | 44.9 | 18.0 | 1.0 | 3262 |
| DRD-205 | Lc. lactis lactis | Kefalograviera | 3.5 | 44.2 | 115.0 | 3.0 | 3251 |
| DRD-206 | Lc. lactis lactis | Kefalograviera | 3.5 | 44.2 | 96.0 | 3.2 | 3243 |
| DRD-207 | Lc. lactis lactis | Kefalograviera | 2.4 | 34.9 | 19.0 | 1.7 | 3364 |
| DRD-164 | Lb. curvatus | Feta | 2.5 | 34.9 | 26.0 | 2.3 | 2754 |
| DRD-170 | Lb. curvatus | Feta | 2.5 | 34.9 | 30.0 | 0.7 | 3255 |
| DRD-171 | Lb. curvatus | Feta | 3.4 | 44.2 | 92.0 | 3.2 | 3254 |
| DRD-2 | Ln. mesenteroides | Kefalograviera | 2.6 | 35.0 | 27.0 | 2.2 | 2562 |
| DRD-30 | Ln. mesenteroides | Feta | 2.5 | 34.9 | 31.0 | 1.9 | 2560 |
| DRD-138 | Ln. mesenteroides | Sheep milk | 2.5 | 45.8 | 20.0 | 2.3 | 2507 |
| DRD-140 | Ln. mesenteroides | Sheep milk | 2.2 | 37.5 | 15.0 | 2.1 | 2630 |
| DRD-141 | Ln. mesenteroides | Sheep milk | 2.5 | 45.8 | 21.0 | 2.3 | 2458 |
| DRD-12 | Lb. brevis | Feta | 2.0 | 37.7 | 14.0 | 1.1 | 2486 |
| DRD-35 | Lb. brevis | Feta | 2.1 | 37.4 | 42.0 | 1.2 | 2555 |
| DRD-51 | Lb. brevis | Kefalograviera | 2.0 | 37.2 | 17.0 | 1.8 | 2551 |
| DRD-52 | Lb. brevis | Kefalograviera | 2.1 | 41.7 | 41.0 | 1.9 | 2506 |
| DRD-59 | Lb. brevis | Kefalograviera | 2.1 | 41.7 | 41.0 | 1.9 | 2398 |
| DRD-60 | Lb. brevis | Kefalograviera | 2.1 | 41.7 | 41.0 | 1.9 | 2504 |
| DRD-136 | Lb. brevis | Sheep milk | 2.0 | 37.2 | 7.0 | 1.8 | 2253 |
| DRD-139 | Lb. brevis | Sheep milk | 2.3 | 46.4 | 12.0 | 2.2 | 2020 |
| DRD-195 | Lb. brevis | Kefalograviera | 2.5 | 45.8 | 20.0 | 2.3 | 2113 |
| DRD-198 | Lb. brevis | Kefalograviera | 2.5 | 45.8 | 21.0 | 2.3 | 1710 |
| DRD-201 | Lb. brevis | Kefalograviera | 2.6 | 34.9 | 26.0 | 0.5 | 2016 |
| DRD-42 | Pd. pentosaceus | Feta | 2.5 | 34.9 | 25.0 | 1.4 | 2008 |
| DRD-48 | Pd. pentosaceus | Kefalograviera | 2.5 | 34.9 | 26.0 | 1.3 | 1998 |
| DRD-61 | Pd. pentosaceus | Kefalograviera | 2.5 | 34.9 | 15.0 | 2.3 | 1662 |
| DRD-144 | Pd. pentosaceus | Feta | 2.5 | 35.4 | 41.0 | 1.8 | 2019 |
| DRD-185 | Pd. pentosaceus | Feta | 2.5 | 35.4 | 43.0 | 1.8 | 1986 |
| Species (Number of Isolates with Plasmid/Total Number of Isolates) | Plasmids | Identity (%) | Length (bp) | Note | NCBI Accession |
|---|---|---|---|---|---|
| Lc. lactis (5/11) | repUS4 | 90 | 1108 | repA(pCI2000) | AF178424 |
| Lc. lactis (8/11) Ln. mesenteroides (1/5) | rep32 | 97 | 1151 | pli0023(pLI100) | AL592102 |
| Lc. lactis (7/11) | repUS33 | 100 | 1352 | repA(pGdh442) | AY849557 |
| Ln. mesenteroides (5/5) | rep31 | 87 | 1132 | LKI10596(LkipL4719) | CP001755 |
| Lb. plantarum (11/16) | repUS73 | 94 | 1100 | rep(pLBUC02) | CP002654 |
| Lb. plantarum (12/16) | rep38 | 81 | 1031 | rep(pLBUC03) | CP002655 |
| Lb. brevis (1/11) Pd. pentosaceus (1/5) | rep28 | 92 | 932 | repA(pCIS4) | CP003162 |
| Ln. mesenteroides (1/5) | repUS72 | 98 | 1036 | C27008541(pKLC4) | CP003855 |
| Lb. brevis (9/11) Lb. curvatus (3/3) Lb. plantarum (12/16) | rep38 | 98 | 885 | repA(LBPp1) | CP005943 |
| Lb. brevis (10/11) Lb. curvatus (3/3) Lb. plantarum (13/16) | rep28 | 99 | 915 | LBPp6g007(LBPp6) | CP005948 |
| Ln. mesenteroides (5/5) | rep31 | 88 | 1133 | LCKp400005(pLCK4) | DQ489739 |
| Lb. curvatus (3/3) | repUS51 | 91 | 662 | rep(pCPS49) | FN806792 |
| Lc. lactis (4/11) | rep33 | 83 | 1131 | rep(pSMA198) | HE613570 |
| Lc. lactis (1/11) | repUS42 | 100 | 1157 | repB(pVF18) | JN172910 |
| Lc. lactis (1/11) | rep32 | 81 | 1168 | repB(pVF22) | JN172912 |
| Lb. plantarum (1/16) Pd. pentosaceus (1/5) | repUS64 | 93 | 956 | repA(pR18) | JN601038 |
| Lc. lactis (1/11) | rep33 | 86 | 1144 | rep(pK214) | X92946 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apostolakos, I.; Paramithiotis, S.; Mataragas, M. Comparative Genomic Analysis Reveals the Functional Traits and Safety Status of Lactic Acid Bacteria Retrieved from Artisanal Cheeses and Raw Sheep Milk. Foods 2023, 12, 599. https://doi.org/10.3390/foods12030599
Apostolakos I, Paramithiotis S, Mataragas M. Comparative Genomic Analysis Reveals the Functional Traits and Safety Status of Lactic Acid Bacteria Retrieved from Artisanal Cheeses and Raw Sheep Milk. Foods. 2023; 12(3):599. https://doi.org/10.3390/foods12030599
Chicago/Turabian StyleApostolakos, Ilias, Spiros Paramithiotis, and Marios Mataragas. 2023. "Comparative Genomic Analysis Reveals the Functional Traits and Safety Status of Lactic Acid Bacteria Retrieved from Artisanal Cheeses and Raw Sheep Milk" Foods 12, no. 3: 599. https://doi.org/10.3390/foods12030599