
Aronia melanocarpa Fruit Juice Modulates ACE2 Immunoexpression and Diminishes Age-Related Remodeling of Coronary Arteries in Rats

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Aronia melanocarpa Juice
2.1.1. HPLC Analysis of Phenolic Compounds
2.1.2. HPLC Determination of Anthocyanins
2.2. Animals
2.3. Light Microscopy
2.4. Immunohistochemistry
2.5. Morphometric Analysis
2.6. Statistical Analysis
3. Results
3.1. Somatometric Parameters
3.2. Chemical Composition of AMJ
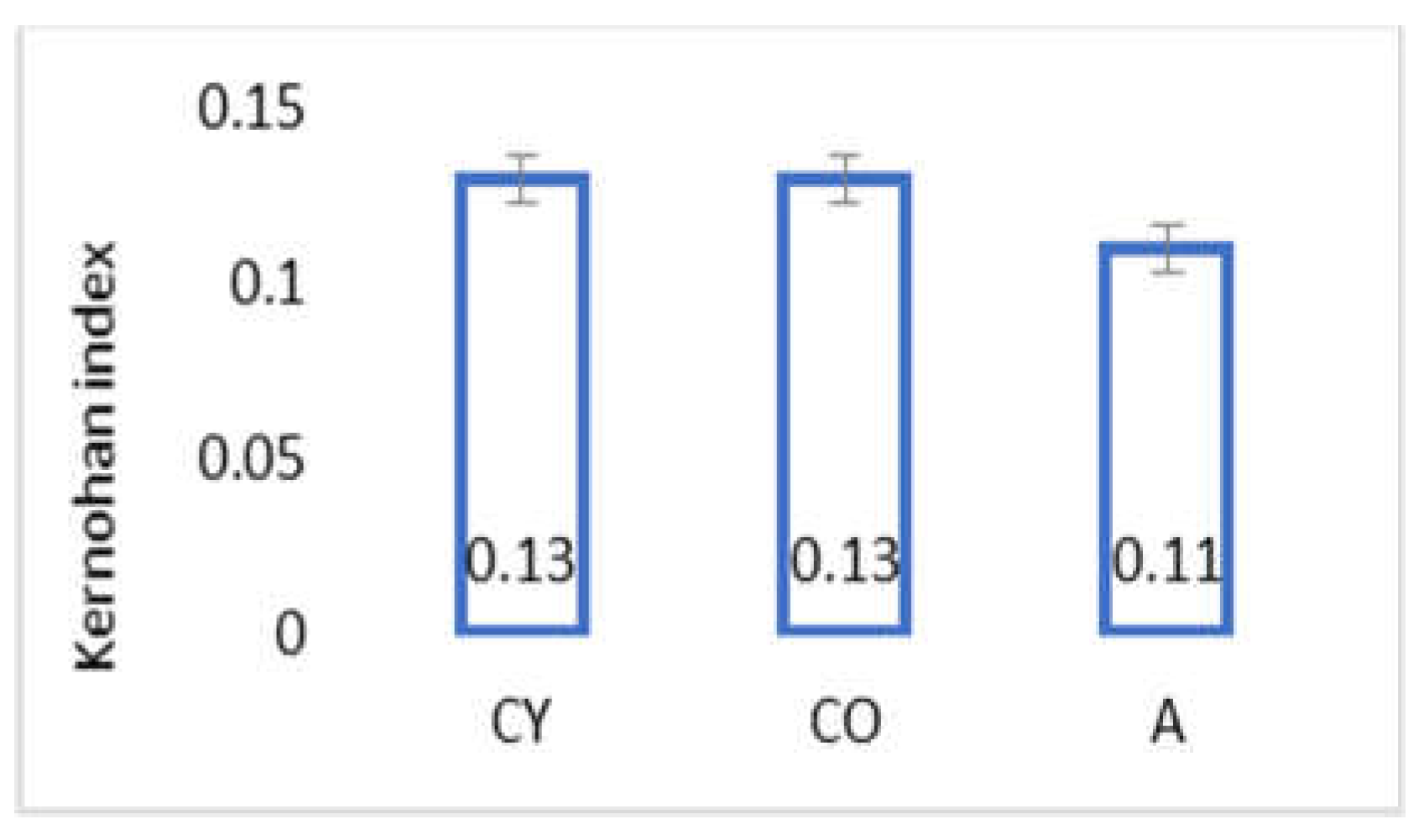
3.3. AMJ Supplementation Did Not Influence Kernohan Index
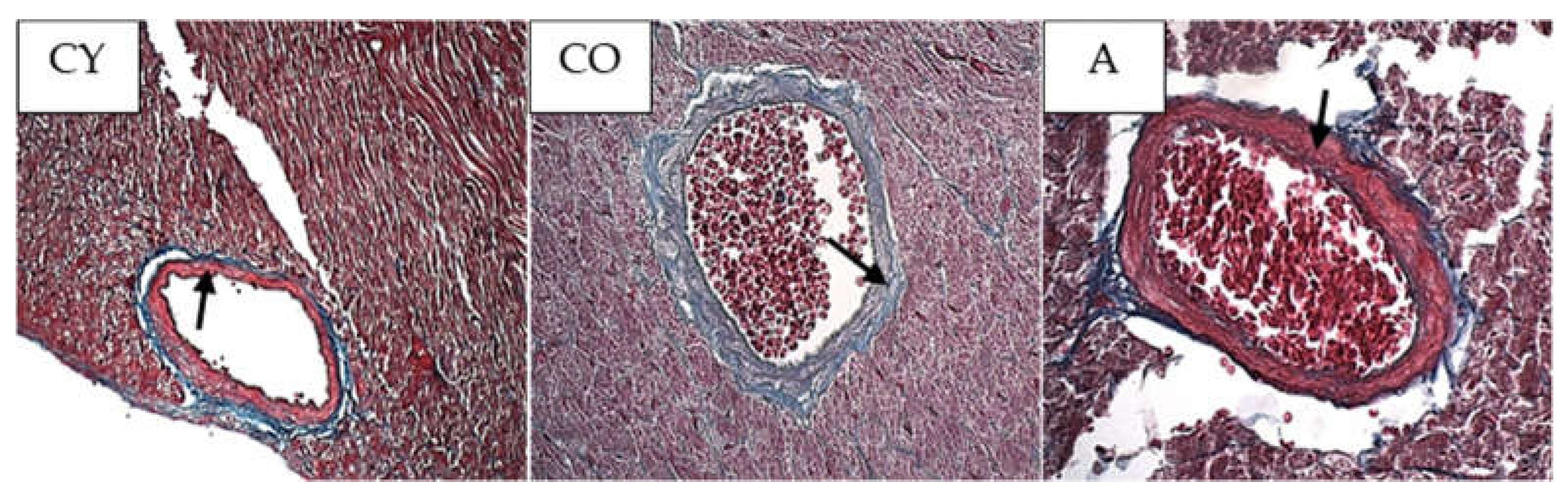
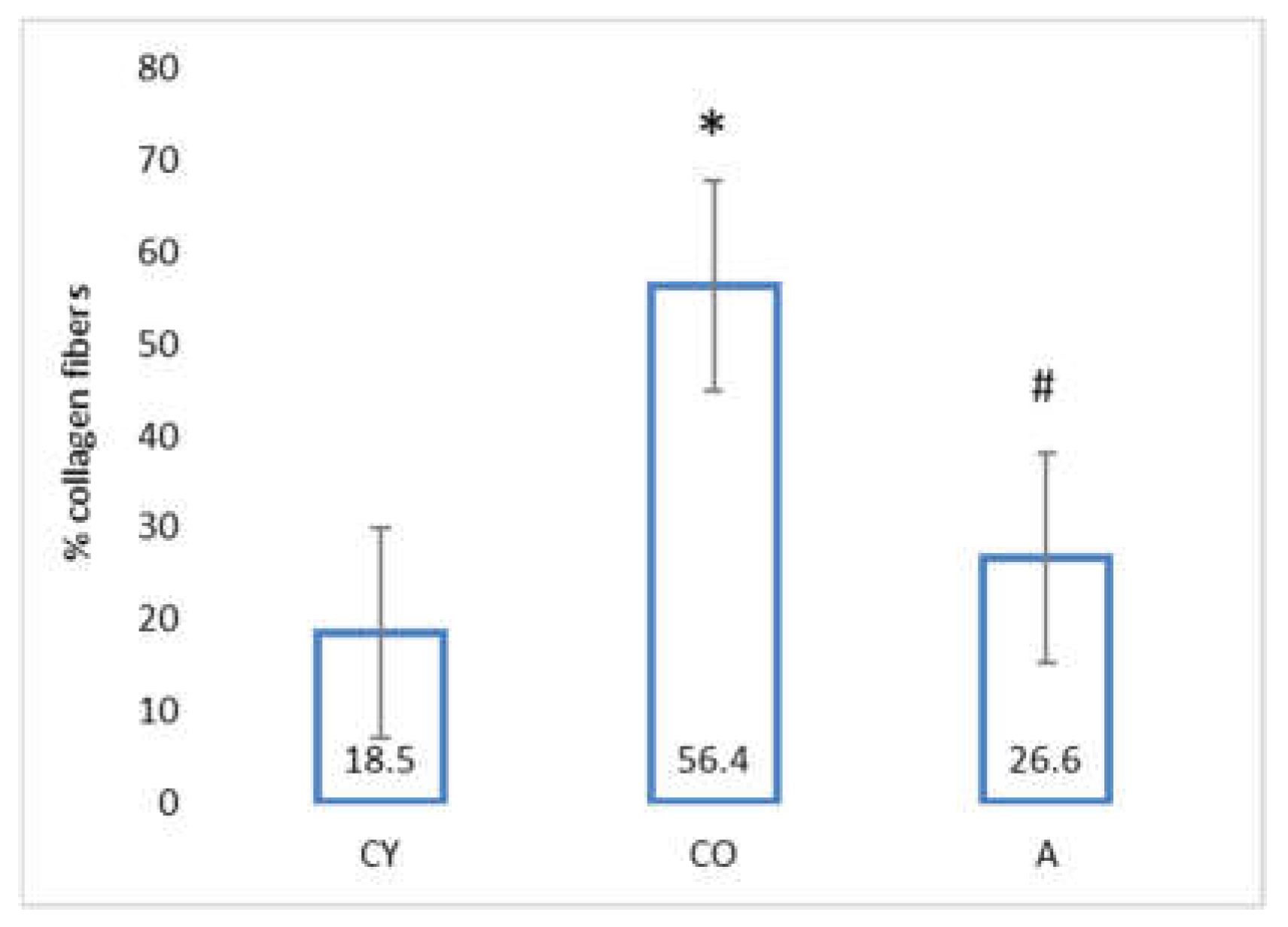
3.4. Effect of AMJ Supplementation on the Amount of Connective Tissue
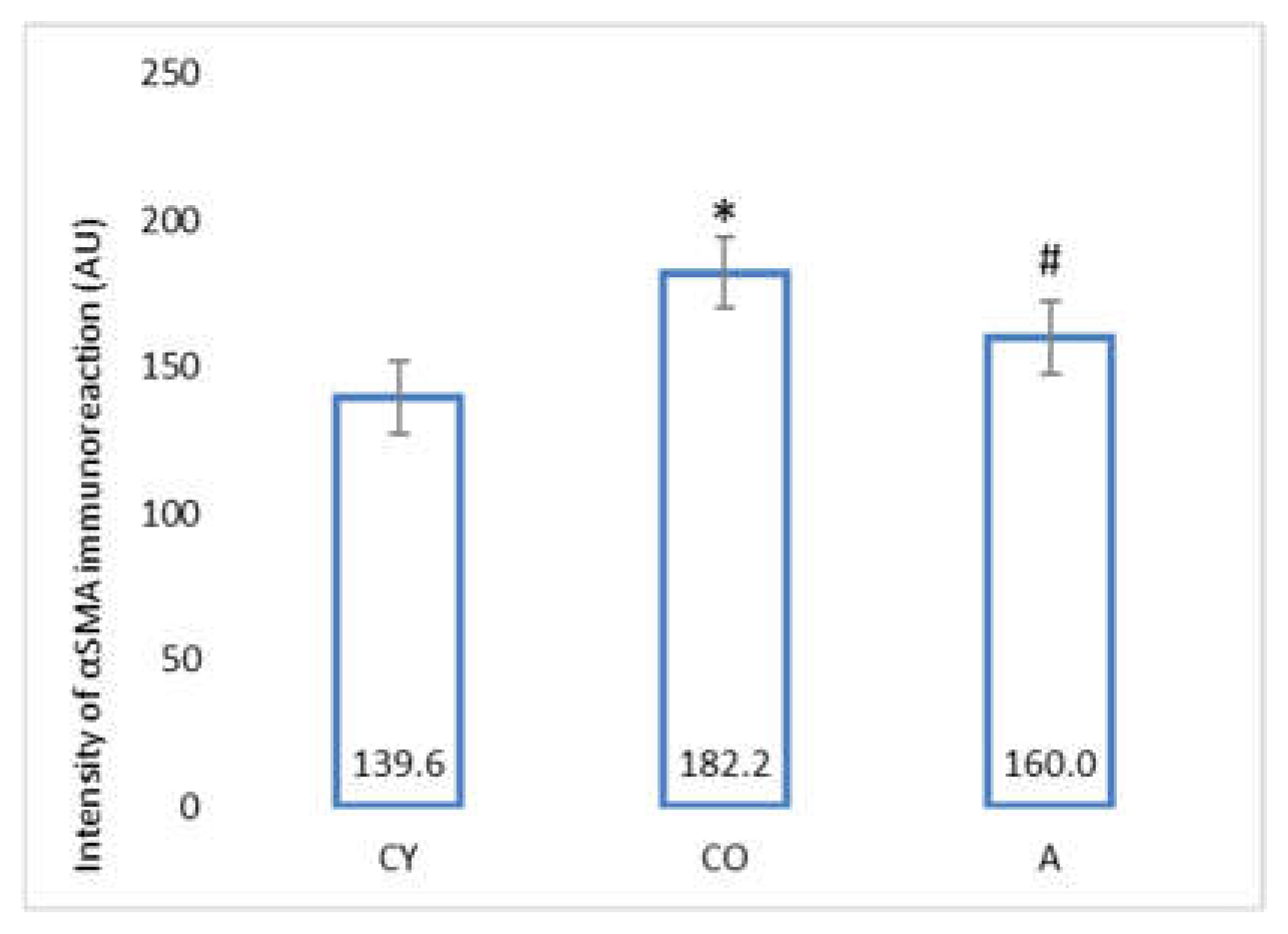
3.5. Effect of AMJ Supplementation on αSMA Immunoexpression
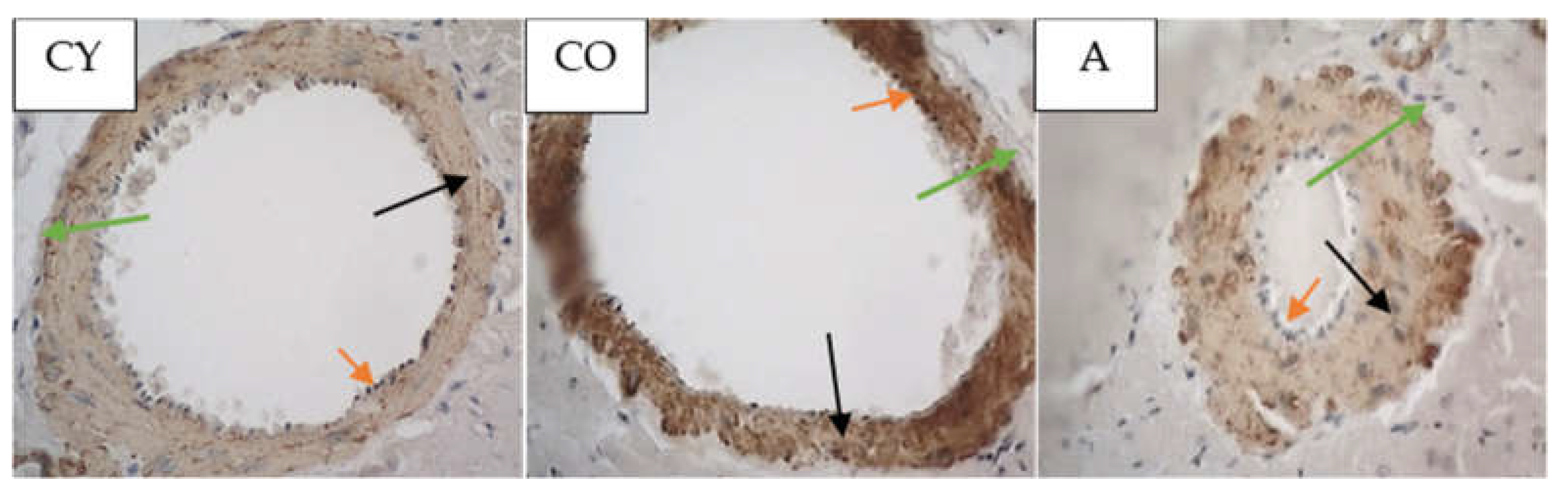
3.6. Effect of AMJ Supplementation on ACE2 Immunoexpression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Najjar, S.; Scuteri, A.; Lakatta, E. Arterial aging is it an immutable cardiovascular risk factor? Hypertension 2005, 46, 454–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, A.U.; Radaelli, A.; Centola, M. Invited review: Aging and the cardiovascular system. J. Appl. Physiol. 2003, 95, 2591–2597. [Google Scholar] [CrossRef] [PubMed]
- Lakatta, E.G. Arterial and cardiac aging: Major shareholders in cardiovascular disease enterprises: Part III: Cellular and molecular clues to heart and arterial aging. Circulation 2003, 107, 490–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Ruiza, M.E.; Pérez-Torresb, I.; Sotoc, M.E.; Pastelínd, G.; Guarner-Lansa, V. Aging in blood vessels. Medicinal agents for systemic arterial hypertension in the elderly. Ageing Res. Rev. 2014, 18, 132–147. [Google Scholar] [CrossRef]
- Tesauro, M.; Mauriello, A.; Rovella, V.; Annicchiarico-Petruzzelli, M.; Cardillo, C.; Melino, G.; Di Daniele, N. Arterial ageing: From endothelial dysfunction to vascular calcification. Eur. J. Intern. Med. 2017, 281, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Greenwald, S.E. Ageing of the conduit arteries. J. Pathol. 2007, 211, 157–172. [Google Scholar] [CrossRef]
- Yin, H.; Pickering, J. Cellular Senescence and Vascular Disease: Novel Routes to Better Understanding and Therapy. Can. J. Cardiol. 2016, 32, 612–623. [Google Scholar] [CrossRef]
- Harvey, A.; Montezano, A.C.; Lopes, R.A.; Rios, F.; Touyz, R.M. Vascular Fibrosis in Aging and Hypertension: Molecular Mechanisms and Clinical Implications. Can. J. Cardiol. 2016, 32, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Xudong, X.; Junzhu, C.; Xingxiang, W.; Furong, Z.; Yanrong, L. Age- and gender-related difference of ACE2 expression in rat lung. Life Sci. 2006, 78, 2166–2171. [Google Scholar] [CrossRef]
- Hamming, I.; Cooper, M.E.; Haagmans, B.L.; Hooper, N.M.; Korstanje, R.; Osterhaus, A.D.; Timens, W.; Turner, A.J.; Navis, G.; van Goor, H. The emerging role of ACE2 in physiology and disease. J. Pathol. 2007, 212, 1–11. [Google Scholar] [CrossRef]
- Bartova, E.; Legartová, S.; Krejčí, J.; Arcidiacono, O. Cell differentiation and aging accompanied by depletion of the ACE2 protein. Aging 2020, 12, 22495–22508. [Google Scholar] [PubMed]
- Slevin, M.; Ahmed, N.; Wang, Q.; McDowell, G.; Badimon, L. Unique vascular protective properties of natural products: Supplements or future main-line drugs with significant anti-atherosclerotic potential? Vasc. Cell 2012, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Roman, I.; Barja, G. Regulation of longevity and oxidative stress by nutritional interventions: Role of methionine restriction. Exp. Gerontol. 2013, 48, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Denev, P.; Kratchanov, C.; Ciz, M.; Lojek, A.; Kratchanova, M. Bioavailability and antioxidant activity of black chokeberry (Aronia melanocarpa) polyphenols: In vitro and in vivo evidences and possible mechanisms of action: A review. Compr. Rev. Food Sci. Food Saf. 2012, 11, 471–489. [Google Scholar] [CrossRef]
- Zhao, C.; Giusti, M.; Malik, M.; Moyer, M.P.; Magnuson, B.A. Effects of commercial anthocyanin-rich extracts on colonic cancer and nontumorigenic colonic cell growth. J. Agric. Food Chem. 2004, 52, 6122–6128. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Gao, J.; Hao, R.; Zhang, C.; Liu, H.; Fan, J.; Wei, J. Aronia melanocarpa Elliot anthocyanins inhibit colon cancer by regulating glutamine metabolism. Food Biosci. 2021, 40, 100910. [Google Scholar] [CrossRef]
- Valcheva-Kuzmanova, S.; Belcheva, A. Current knowledge of Aronia melanocarpa as a medicinal plant. Folia Med. 2006, 48, 11–17. [Google Scholar]
- Valcheva-Kuzmanova, S.; Kuzmanov, K.; Tsanova-Savova, S. Lipid-lowering effects of Aronia melanocarpa fruit juice in rats fed cholesterol-containing diets. J. Food Biochem. 2007, 31, 589–602. [Google Scholar] [CrossRef]
- Valcheva-Kuzmanova, S.; Kuzmanov, A.; Kuzmanova, V.; Tzaneva, M. Aronia melanocarpa fruit juice ameliorates the symptoms of inflammatory bowel disease in TNBS-induced colitis in rats. Food Chem. Toxicol. 2018, 113, 33–39. [Google Scholar] [CrossRef]
- Gao, J.; Yu, W.; Zhang, C.; Liu, H.; Fan, J.; Wei, J. The protective effect and mechanism of Aronia melanocarpa Elliot anthocyanins on IBD model mice. Food Biosci. 2021, 41, 101075. [Google Scholar] [CrossRef]
- Daskalova, E.; Delchev, S.; Topolov, M.; Denev, P.; Dimitrova, S.; Uzunova, Y.; Valcheva-Kuzmanova, S.; Vladimirova-Kitova, L. Assessment of cognitive and locomotor functions after Aronia melanocarpa supplementation on aged rats. Food Chem. Toxicol. 2019, 132, 110674. [Google Scholar] [CrossRef] [PubMed]
- Denev, P.; Kratchanova, M.; Petrova, I.; Klisurova, D.; Georgiev, Y.; Ognyanov, M.; Yanakieva, I. Black chokeberry (Aronia melanocarpa (Michx.) Elliot) fruits and functional drinks differ significantly in their chemical composition and antioxidant activity. J. Chem. 2018, 13, 9574587. [Google Scholar] [CrossRef] [Green Version]
- Wolden-Hanson, T. Changes in body composition in response to challenges during aging in rats. Interdiscip. Top. Gerontol. 2010, 37, 64–83. [Google Scholar]
- Aissaoui, A.; Zizi, S.; Israili, Z.; Lyoussi, B. Hypoglycemic and hypolipidemic effects of Coriandrum sativum L. in Meriones shawi rats. J. Ethnopharmacol. 2011, 137, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Man, A.; Wang, Y. Age-associated arterial remodeling. EC Cardiol. 2017, 4, 137–164. [Google Scholar]
- Daskalova, E.; Delchev, S.; Bivolarski, I.; Denev, P.; Kratchanova, M.; Cvetkov, P.; Kaluch, M. Vasoprotective properties of Aronia melanocarpa—A histological and morphometric study. Acta Morphol. Anthropol. 2015, 22, 3–8. [Google Scholar]
- Daskalova, E.; Delchev, S.; Denev, P.; Vladimirova-Kitova, L.; Valcheva-Kuzmanova, S.; Kitov, S.; Kanarev, M. Age-related changes in rat thymus connective tissue influenced by Aronia melanocarpa. Acta Morphol. Anthropol. 2019, 26, 1–2. [Google Scholar]
- Shinde, A.; Humeres, C.; Frangogiannis, N. The role of α-smooth muscle actin in fibroblast-mediated matrix contraction and remodeling. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 298–309. [Google Scholar] [CrossRef]
- Harikrishnan, V.; Titus, A.; Cowling, R.; Kailasam, S. Collagen receptor cross-talk determines -smooth muscle actin-dependent collagen gene expression in angiotensin II–stimulated cardiac fibroblasts. J. Biol. Chem. 2019, 294, 19723–19739. [Google Scholar]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Lakatta, E.G. So! What’s aging? Is cardiovascular aging a disease? J. Mol. Cell. Cardiol. 2015, 83, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rensen, S.S.; Doevendans, P.A.; van Eys, G.J. Regulation and characteristics of vascular smooth muscle cell phenotypic diversity. Neth. Heart J. 2007, 15, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heeneman, S.; Sluimer, J.; Daemen, M. Angiotensin-converting enzyme and vascular remodeling. Circ. Res. 2007, 101, 441–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.N.; Kim, M.Y.; Lim, J.H.; Kim, Y.; Shin, S.J.; Park, C.W.; Kim, Y.S.; Chang, Y.S.; Yoon, H.E.; Choi, B.S. The protective effect of resveratrol on vascular aging by modulation of the renin–angiotensin system. Atherosclerosis 2018, 270, 123–131. [Google Scholar] [CrossRef]
- Rodrigo, R.; Miranda, A.; Vergara, L. Modulation of endogenous antioxidant system by wine polyphenols in human disease. Clin. Chim. Acta 2011, 412, 410–424. [Google Scholar] [CrossRef]
- Ungvari, Z.; Bagi, Z.; Feher, A.; Recchia, F.A.; Sonntag, W.E.; Pearson, K.; de Cabo, R.; Csiszar, A. Resveratrol confers endothelial protection via activation of the antioxidant transcription factor Nrf2. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H18–H24. [Google Scholar] [CrossRef] [Green Version]
- Stangl, V.; Dreger, H.; Stangl, K.; Lorenz, M. Molecular targets of tea polyphenols in the cardiovascular system. Cardiovasc. Res. 2012, 73, 348–358. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and human health: Prevention of disease and mechanisms of action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Liu, X.; Zheng, Y.; Liu, W.; Ding, C. Aronia melanocarpa polysaccharide ameliorates inflammation and aging in mice by modulating the AMPK/SIRT1/NF-kappa B signaling pathway and gut microbiota. Sci. Rep. 2021, 11, 20558. [Google Scholar] [CrossRef]
- Kasprzak-Drozd, K.; Oniszczuk, T.; Soja, J.; Gancarz, M.; Wojtunik-Kulesza, K.; Markut-Miotła, E.; Oniszczuk, A. The Efficacy of Black Chokeberry Fruits against Cardiovascular Diseases. Int. J. Mol. Sci. 2021, 22, 6541. [Google Scholar] [CrossRef]
- Jurikova, T.; Mlcek, J.; Skrovankova, S.; Sumczynski, D.; Sochor, J.; Hlavacova, I.; Snopek, L.; Orsavova, J. Fruits of Black Chokeberry Aronia melanocarpa in the Prevention of Chronic Diseases. Molecules 2017, 22, 944. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Biswas, S.K.; Kirkham, P.A. Regulation of inflammation and redox signaling by dietary polyphenols. Biochem. Pharmacol. 2006, 72, 1439–1452. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.; Kandaswami, C.; Theoharides, T. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar] [PubMed]
- Habauzit, V.; Morand, C. Evidence for a protective effect of polyphenols-containing foods on cardiovascular health: An update for clinicians. Ther. Adv. Chronic Dis. 2012, 3, 87–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H. Polyphenols and health: Update and perspectives. Arch. Biochem. Biophys. 2006, 501, 2–5. [Google Scholar] [CrossRef]
- De Pascual-Teresa, S.; Moreno, D.A.; García-Viguera, C. Flavanols and anthocyanins in cardiovascular health: A review of current evidence. Int. J. Mol. Sci. 2010, 11, 1679–1703. [Google Scholar] [CrossRef] [Green Version]
- Wallace, T.C. Anthocyanins in cardiovascular disease. Adv. Nutr. 2011, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Khurana, S.; Venkataraman, K.; Hollingsworth, A.; Piche, M.; Tai, T. Polyphenols: Benefits to the Cardiovascular System in Health and in Aging. Nutrients 2013, 5, 3779–3827. [Google Scholar] [CrossRef]
- Samavati, L.; Uhal, B.D. ACE2, much more than just a receptor for SARS-COV-2. Front. Cell. Infect. Microbiol. 2020, 317. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Polyphenols (mg/L) | 11,237.4 ± 456.2 |
| Flavonoids (mg/L) | |
| Quercetin | 49.6 ± 3.2 |
| Quercetin-3-β-glucoside | 228.8 ± 11.0 |
| Rutin | 446.5 ± 12.5 |
| Epicatechin | 408.2 ± 25.6 |
| Anthocyanins (mg/L) | |
| Cyanidin-3-galactoside | 1498.4 ± 102.3 |
| Cyanidin-3-glucoside | 120.1 ± 8.7 |
| Cyanidin-3-arabinoside | 501.9 ± 31.8 |
| Cyanidin-3-xyloside | 4.59 ± 0.2 |
| Phenolic acids, mg/L | |
| Chlorogenic acid | 1375.6 ± 80.3 |
| Neochlorogenic acid | 1543.1 ± 111.2 |
| Body Weight (g) | p < 0.05 | BMI | p < 0.05 | Heart Weight (g) | p < 0.05 | Heart Weight Index | |
|---|---|---|---|---|---|---|---|
| CY | 158.57 | 0.52 | 0.50 | 0.31 | |||
| CO | 402.14 | * CY | 0.70 | * CY | 1.06 | * CY | 0.26 |
| A | 417.86 | # CY | 0.69 | * CY | 1.06 | # CY | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daskalova, E.; Delchev, S.; Vladimirova-Kitova, L.; Bivolarski, I.; Pencheva, M.; Denev, P. Aronia melanocarpa Fruit Juice Modulates ACE2 Immunoexpression and Diminishes Age-Related Remodeling of Coronary Arteries in Rats. Foods 2022, 11, 1220. https://doi.org/10.3390/foods11091220
Daskalova E, Delchev S, Vladimirova-Kitova L, Bivolarski I, Pencheva M, Denev P. Aronia melanocarpa Fruit Juice Modulates ACE2 Immunoexpression and Diminishes Age-Related Remodeling of Coronary Arteries in Rats. Foods. 2022; 11(9):1220. https://doi.org/10.3390/foods11091220
Chicago/Turabian StyleDaskalova, Elena, Slavi Delchev, Lyudmila Vladimirova-Kitova, Iliya Bivolarski, Mina Pencheva, and Petko Denev. 2022. "Aronia melanocarpa Fruit Juice Modulates ACE2 Immunoexpression and Diminishes Age-Related Remodeling of Coronary Arteries in Rats" Foods 11, no. 9: 1220. https://doi.org/10.3390/foods11091220