
Fermented Oyster (Crassostrea gigas) Extract Cures and Prevents Prednisolone-Induced Bone Resorption by Activating Osteoblast Differentiation

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagent and Antibody
2.2. Cell Culture and Osteoblast Differentiation
2.3. Alizarin Red Staining
2.4. ALP Staining
2.5. Reverse Transcription–Polymerase Chain Reaction (RT-PCR) Using MC3T3E-1 Cells
2.6. Protein Extraction and Western Blotting
2.7. Zebrafish Maintenance and Vertebral Staining
2.8. RT-PCR Using Zebrafish Larvae
2.9. Relative Uterine Weight/Body Weight in OVX Mice
2.10. ERE-Luciferase Activity in MCF-7 Cells
2.11. Cell Proliferation Assay
2.12. Statistical Analysis
3. Results
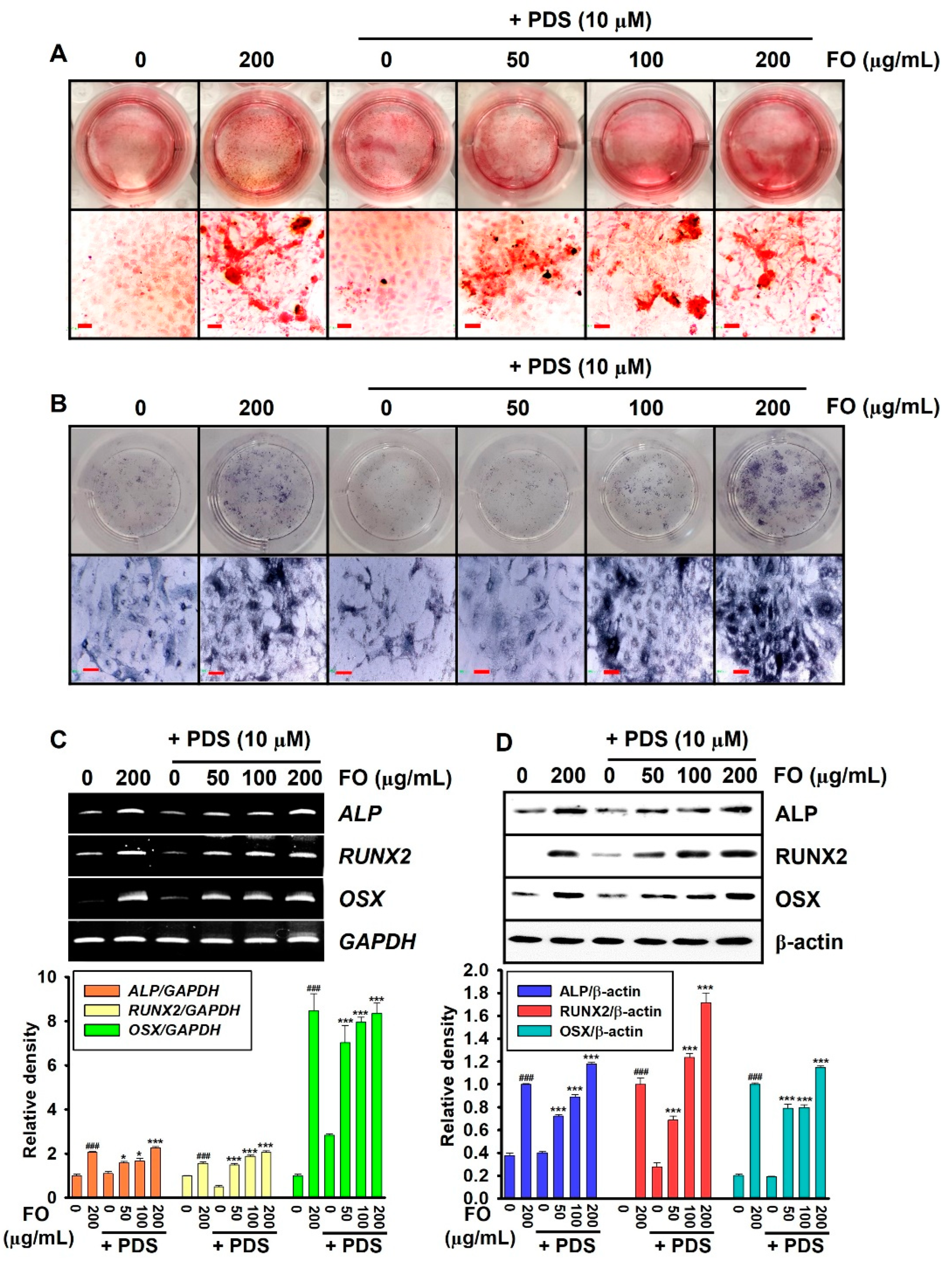
3.1. PDS-Induced Anti-Osteogenic Activity Decreases in MC3T3-E1 Cells by Post-Treatment with FO
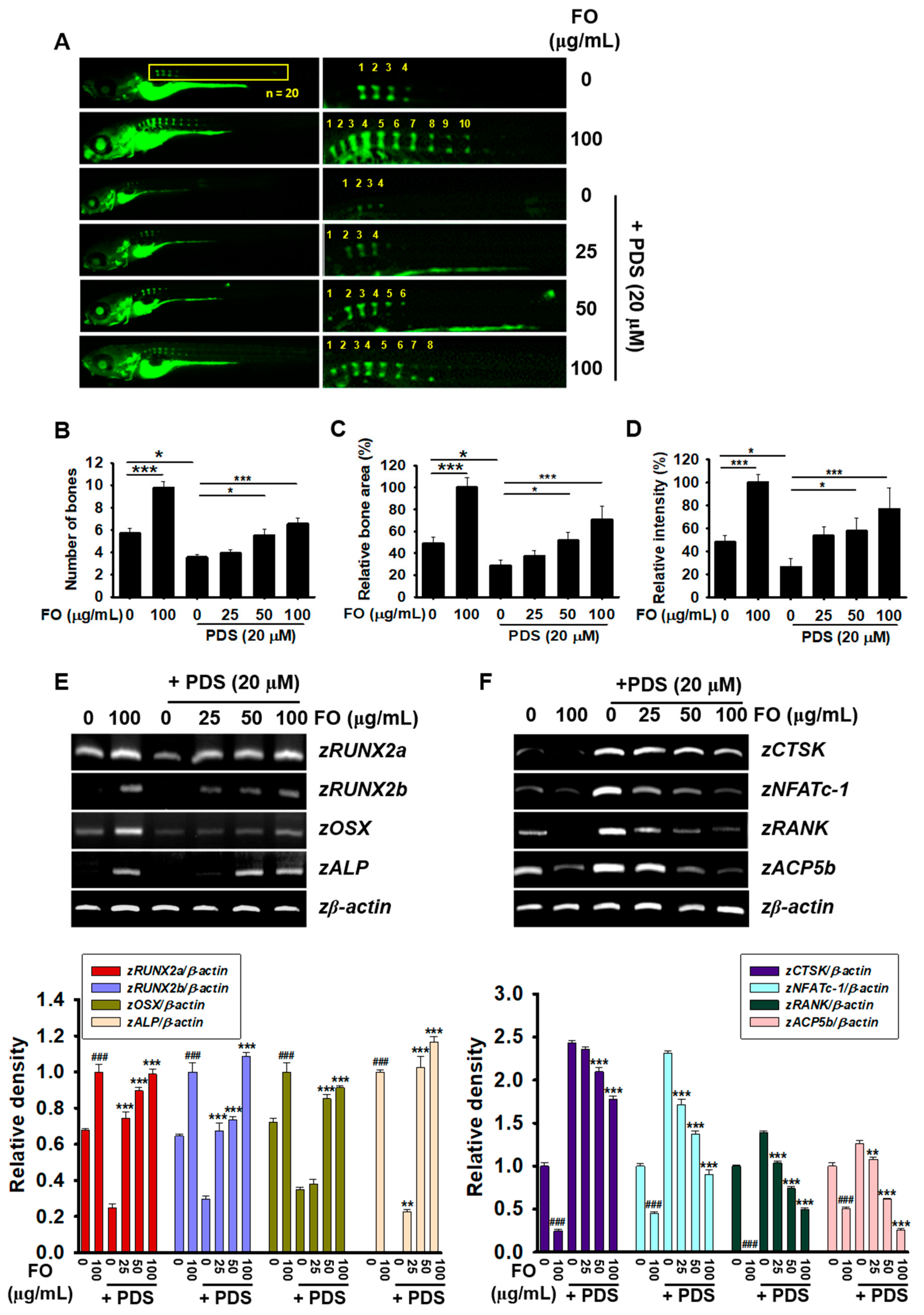
3.2. Post-Treatment with FO Overcomes PDS-Induced Delay of Vertebral Formation in Zebrafish Larvae
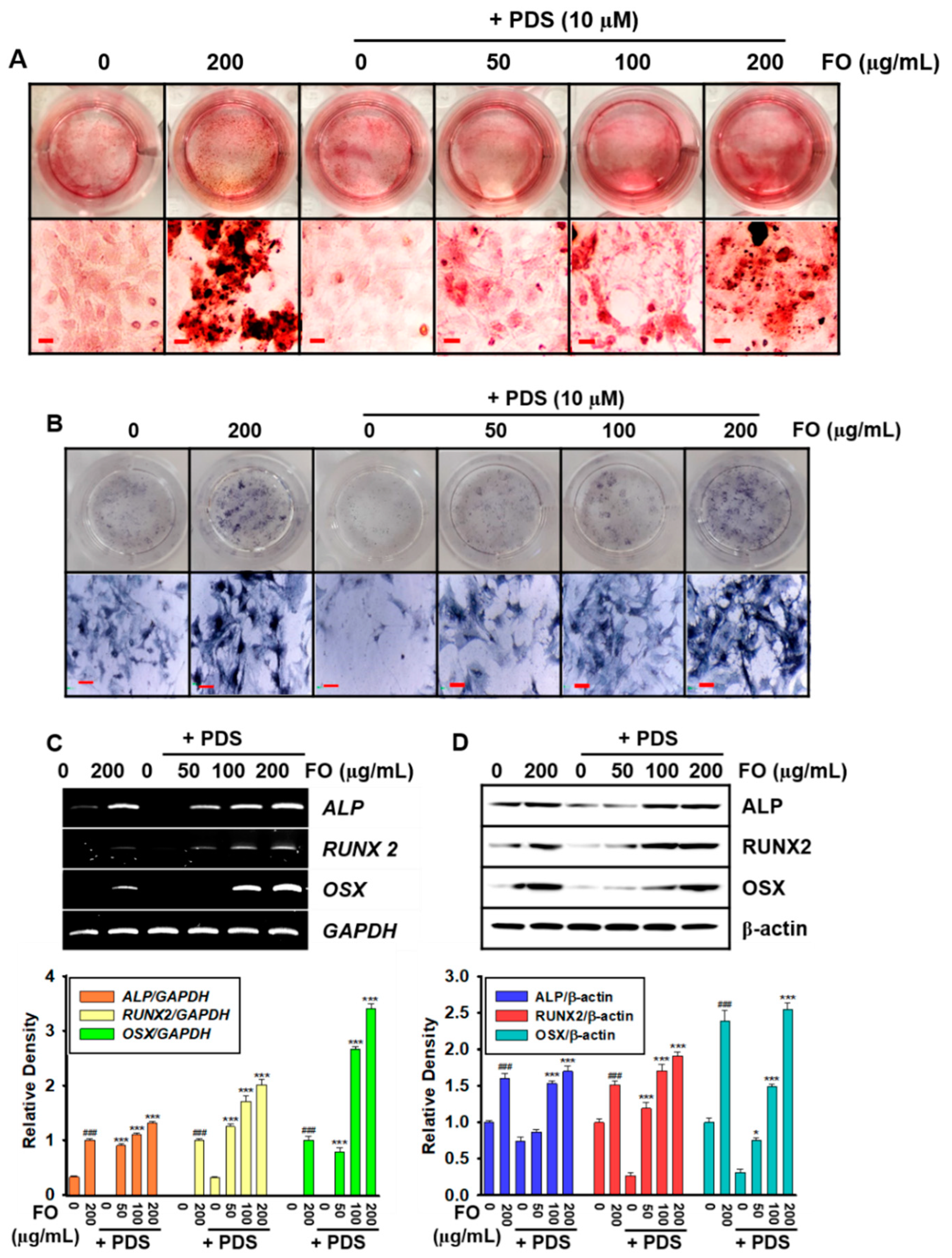
3.3. Pretreatment with FO Prevents PDS-Induced Anti-Osteogenic Activity in MC3T3-E1 Cells
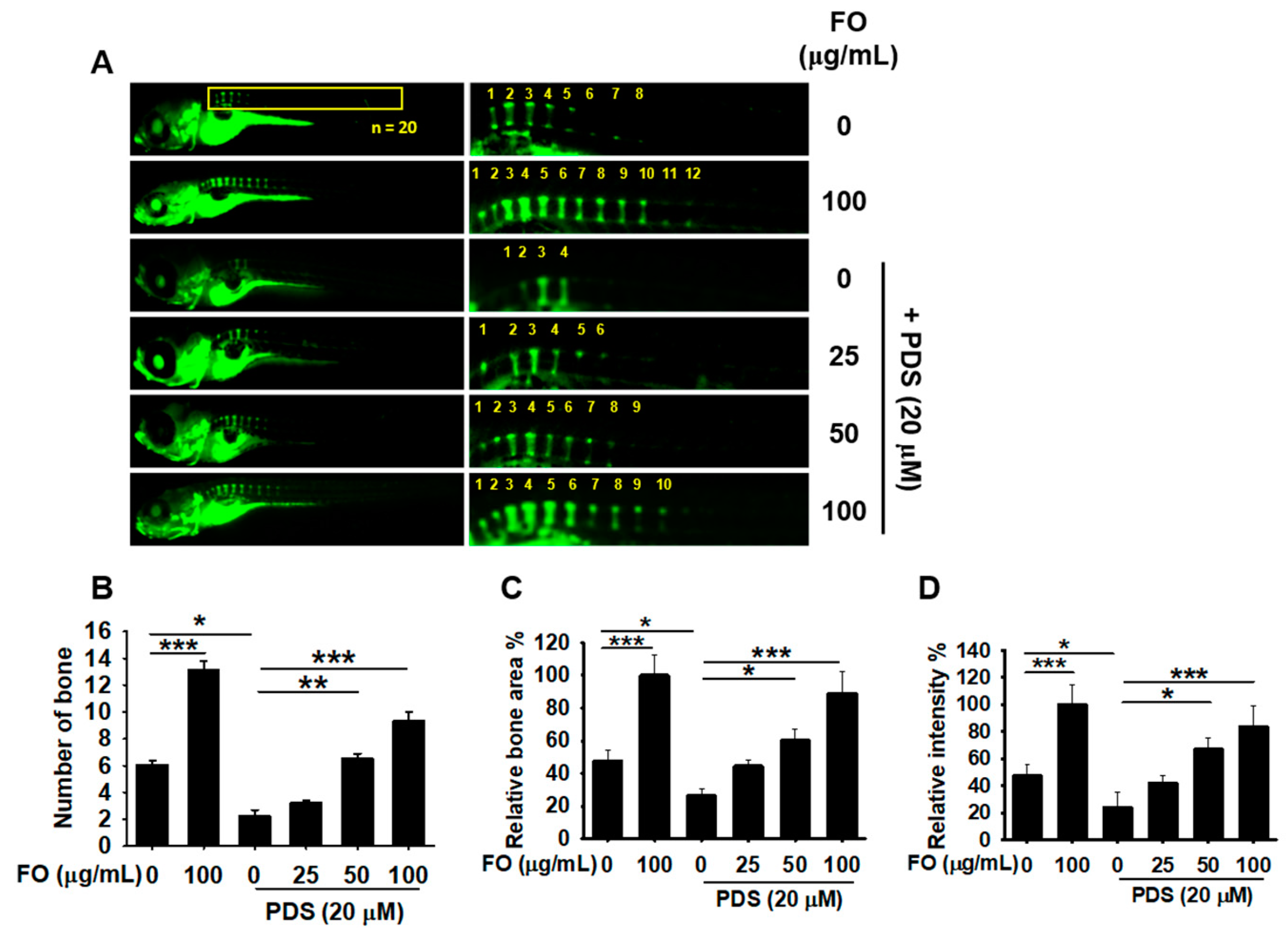
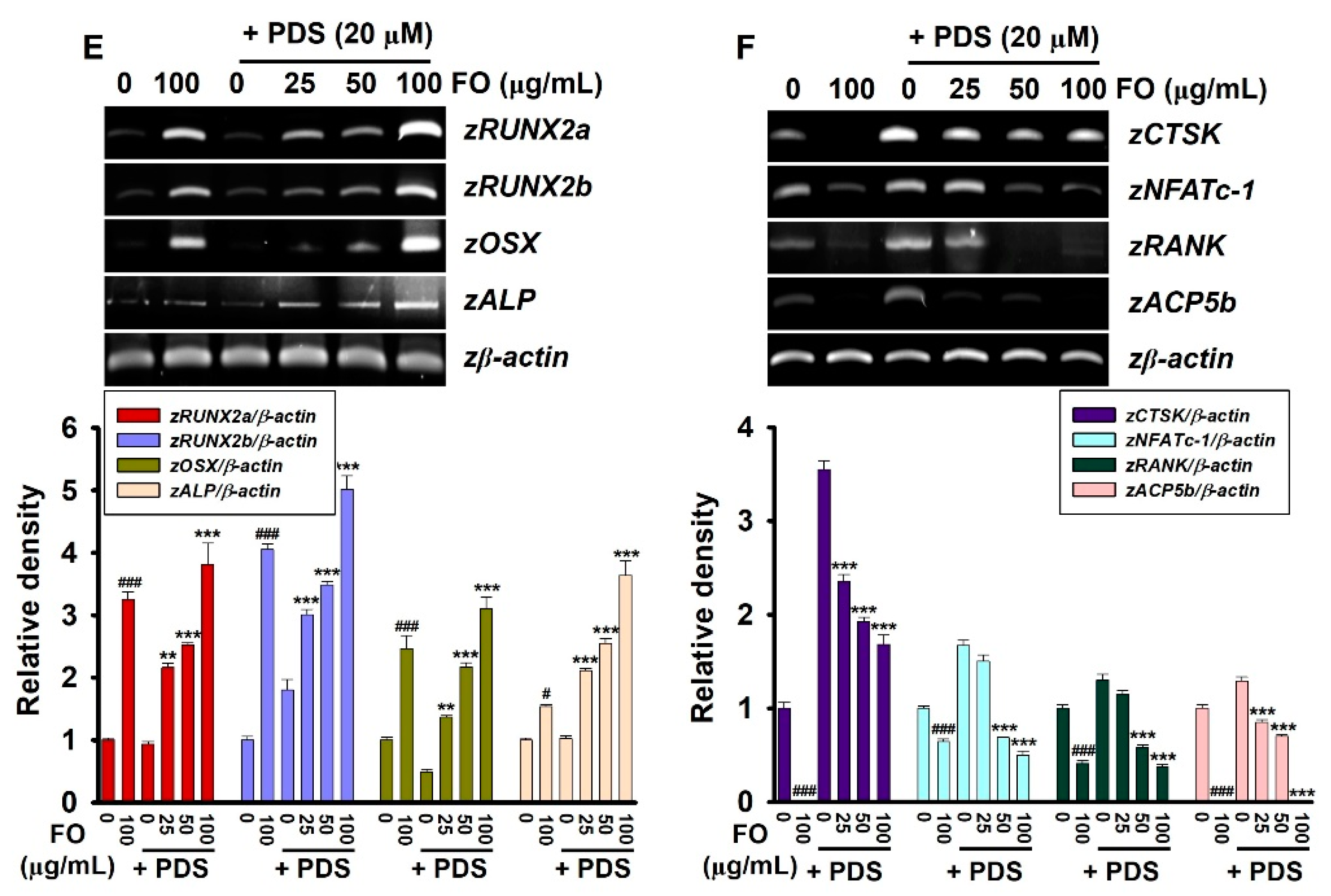
3.4. Pretreatment with FO Prevents PDS-Induced Vertebral Resorption in Zebrafish Larvae
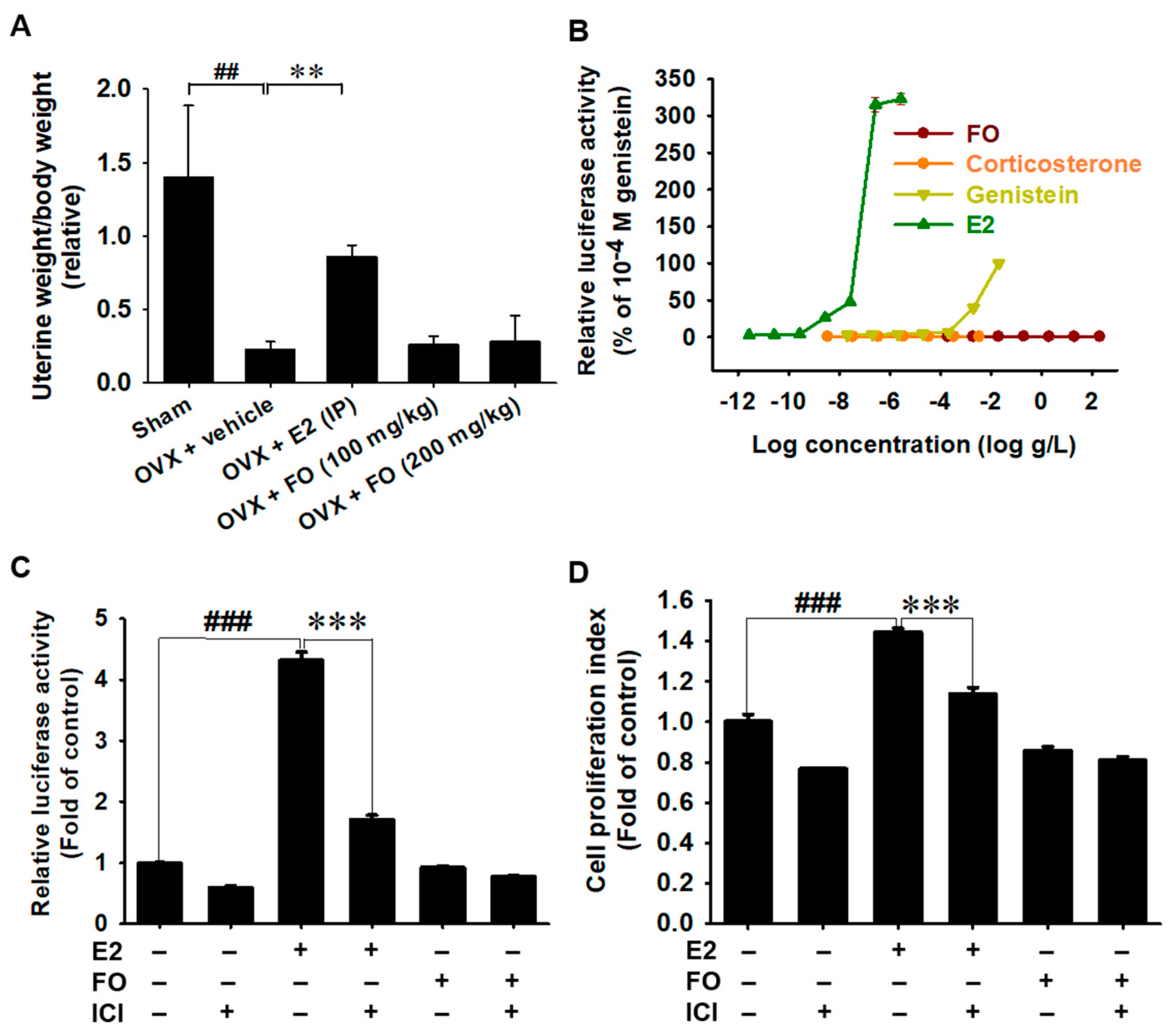
3.5. Estrogenic Activity Is Not Associated with FO
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Briot, K.; Roux, C. Glucocorticoid-induced osteoporosis. RMD Open 2015, 1, e000014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, D.E. Basic and clinical pharmacology of glucocorticosteroids. Anesth. Prog. 2013, 60, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Raeder, J.; Dahl, V. Clinical application of glucocorticoids, antineuropathics, and other analgesic adjuvants for acute pain management. In Acute Pain Management; Ginsberg, B., Viscusi, E.R., de Leon-Cassasola, O.A., Sinatra, R.S., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 377–390. [Google Scholar] [CrossRef]
- Liberman, A.C.; Budziñski, M.L.; Sokn, C.; Gobbini, R.P.; Steininger, A.; Arzt, E. Regulatory and mechanistic actions of glucocorticoids on T and inflammatory cells. Front. Endocrinol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, A.E.; Chapman, K.E. The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights. Mol. Cell. Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef]
- Mitra, R. Adverse effects of corticosteroids on bone metabolism: A review. PM R 2011, 3, 466–471. [Google Scholar] [CrossRef]
- Payer, J.; Brazdilova, K.; Jackuliak, P. Management of glucocorticoid-induced osteoporosis: Prevalence, and emerging treatment options. Drug Healthc. Patient Saf. 2010, 2, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Kassi, E.; Nasiri-Ansari, N.; Papavassiliou, A.G. Vitamin D affects glucocorticoid action in target cells. Oncotarget 2017, 8, 7220–7221. [Google Scholar] [CrossRef] [Green Version]
- Ilias, I.; Zoumakis, E.; Ghayee, H. An overview of glucocorticoid induced osteoporosis. In Endotext; MDText.com, Inc.: South Dartmouth, MA, USA, 2018. [Google Scholar]
- Weinstein, R.S.; Jia, D.; Powers, C.C.; Stewart, S.A.; Jilka, R.L.; Parfitt, A.M.; Manolagas, S.C. The Skeletal Effects of Glucocorticoid Excess Override Those of Orchidectomy in Mice. Endocrinology 2004, 145, 1980–1987. [Google Scholar] [CrossRef] [Green Version]
- Hachemi, Y.; Rapp, A.E.; Picke, A.-K.; Weidinger, G.; Ignatius, A.; Tuckermann, J. Molecular mechanisms of glucocorticoids on skeleton and bone regeneration after fracture. J. Mol. Endocrinol. 2018, 61, R75–R90. [Google Scholar] [CrossRef]
- Sousa, A.R.; Marshall, R.P.; Warnock, L.C.; Bolton, S.; Hastie, A.; Symon, F.; Hargadon, B.; Marshall, H.; Richardson, M.; Brightling, C.E.; et al. Responsiveness to oral prednisolone in severe asthma is related to the degree of eosinophilic airway inflammation. Clin. Exp. Allergy 2017, 47, 890–899. [Google Scholar] [CrossRef] [Green Version]
- Sathasivam, S. Steroids and immunosuppressant drugs in myasthenia gravis. Nat. Clin. Pract. Neurol. 2008, 4, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Flower, R.J. Prednisone. In xPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier: New York, NY, USA, 2009; pp. 1–6. [Google Scholar] [CrossRef]
- Weinstein, R.S. Glucocorticoid-Induced Bone Disease. N. Engl. J. Med. 2011, 365, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Bergström, I.; Isaksson, H.; Koskela, A.; Tuukkanen, J.; Ohlsson, C.; Andersson, G.; Windahl, S.H. Prednisolone treatment reduces the osteogenic effects of loading in mice. Bone 2018, 112, 10–18. [Google Scholar] [CrossRef]
- McClung, M.; Harris, S.T.; Miller, P.D.; Bauer, D.C.; Davison, K.S.; Dian, L.; Hanley, D.A.; Kendler, D.L.; Yuen, C.K.; Lewiecki, E.M. Bisphosphonate Therapy for Osteoporosis: Benefits, Risks, and Drug Holiday. Am. J. Med. 2013, 126, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Bilezikian, J.P.; Dempster, D.W.; Lewiecki, E.M.; Miller, P.D.; Neer, R.M.; Recker, R.R.; Shane, E.; Shoback, D.; Potts, J.T. Benefits and Risks of Bisphosphonate Therapy for Osteoporosis. J. Clin. Endocrinol. Metab. 2012, 97, 2272–2282. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Tao, L.; Shi, C.; Yang, S.; Li, D.; Sheng, J.; Tian, Y. Fermentation Improves Calcium Bioavailability in Moringa oleifera leaves and Prevents Bone Loss in Calcium-deficient Rats. Food Sci. Nutr. 2020, 8, 3692–3703. [Google Scholar] [CrossRef]
- Rizzoli, R.; Biver, E. Effects of Fermented Milk Products on Bone. Calcif. Tissue Int. 2018, 102, 489–500. [Google Scholar] [CrossRef]
- Molagoda, I.M.N.; Karunarathne, W.; Choi, Y.H.; Park, E.K.; Jeon, Y.J.; Lee, B.J.; Kang, C.H.; Kim, G.Y. Fermented Oyster Extract Promotes Osteoblast Differentiation by Activating the Wnt/β-Catenin Signaling Pathway, Leading to Bone Formation. Biomolecules 2019, 9, 711. [Google Scholar] [CrossRef] [Green Version]
- Molagoda, I.M.N.; Jayasingha, J.; Choi, Y.H.; Park, E.K.; Jeon, Y.J.; Lee, B.J.; Kim, G.Y. Fermented Oyster Extract Promotes Insulin-Like Growth Factor-1-Mediated Osteogenesis and Growth Rate. Mar. drugs 2020, 18, 472. [Google Scholar] [CrossRef]
- Jeong, J.-W.; Choi, S.H.; Han, M.H.; Kim, G.-Y.; Park, C.; Hong, S.H.; Lee, B.-J.; Park, E.K.; Kim, S.O.; Leem, S.-H.; et al. Protective Effects of Fermented Oyster Extract against RANKL-Induced Osteoclastogenesis through Scavenging ROS Generation in RAW 264.7 Cells. Int. J. Mol. Sci. 2019, 20, 1439. [Google Scholar] [CrossRef] [Green Version]
- Ihn, H.J.; Kim, J.A.; Lim, S.; Nam, S.-H.; Hwang, S.H.; Lim, J.; Kim, G.-Y.; Choi, Y.H.; Jeon, Y.-J.; Lee, B.-J.; et al. Fermented Oyster Extract Prevents Ovariectomy-Induced Bone Loss and Suppresses Osteoclastogenesis. Nutrients 2019, 11, 1392. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.R.; Lai, Y.H.; Tsai, J.J.; Hsiao, C.D. Live fluorescent staining platform for drug-screening and mechanism-analysis in zebrafish for bone mineralization. Molecules 2017, 22, 2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, K.; Takahira, K.; Inari, M.; Satoh, Y.; Hayakawa, K.; Tabuchi, Y.; Ogai, K.; Nishiuchi, T.; Kondo, T.; Mikuni-Takagaki, Y.; et al. Zebrafish scales respond differently to in vitro dynamic and static acceleration: Analysis of interaction between osteoblasts and osteoclasts. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 166, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Athapaththu, A.M.G.K.; Molagoda, I.M.N.; Jayasooriya, R.G.P.T.; Choi, Y.H.; Jeon, Y.-J.; Park, J.-H.; Lee, B.-J.; Kim, G.-Y. Gamma-Aminobutyric Acid (GABA) Promotes Growth in Zebrafish Larvae by Inducing IGF-1 Expression via GABAA and GABAB Receptors. Int. J. Mol. Sci. 2021, 22, 11254. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.M.; Lee, E.H. Transcriptional regulatory cascades in Runx2-dependent bone development. Tissue Eng. Part B Rev. 2013, 19, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C. Transcriptional regulation of bone formation by the osteoblast-specific transcription factor Osx. J. Orthop. Surg. Res. 2010, 5, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; Wang, C.; Tang, Q.; Yang, F.; Xu, Y. Possible mechanisms of prednisolone-induced osteoporosis in zebrafish larva. Biomed. Pharmacother. 2018, 101, 981–987. [Google Scholar] [CrossRef]
- Molagoda, I.M.N.; Kang, C.-H.; Lee, M.-H.; Choi, Y.H.; Lee, C.-M.; Lee, S.; Kim, G.-Y. Fisetin promotes osteoblast differentiation and osteogenesis through GSK-3β phosphorylation at Ser9 and consequent β-catenin activation, inhibiting osteoporosis. Biochem. Pharmacol. 2021, 192, 114676. [Google Scholar] [CrossRef]
- Sun, B.; Sun, J.; Han, X.; Liu, H.; Li, J.; Du, J.; Feng, W.; Liu, B.; Cui, J.; Guo, J.; et al. Immunolocalization of MMP 2, 9 and 13 in prednisolone induced osteoporosis in mice. Histol. Histopathol. 2015, 31, 11702. [Google Scholar] [CrossRef]
- Di Cosmo, A.; Di Cristo, C.; Paolucci, M. A estradiol-17b receptor in the reproductive system of the female of Octopus vulgaris: Characterization and immunolocalization. Mol. Reprod. Dev. 2002, 61, 367–375. [Google Scholar] [CrossRef]
- Fan, J.J.; Wang, S.; Tang, J.P.; Zhao, J.L.; Wang, L.; Wang, J.X.; Liu, S.L.; Li, F.; Long, S.X.; Yang, Y. Bioaccumulation of endocrine disrupting compounds in fish with different feeding habits along the largest subtropical river, China. Environ. Pollut. 2019, 247, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Le Curieux-Belfond, O.; Fiévet, B.; Séralini, G.; Mathieu, M. Short-term bioaccumulation, circulation and metabolism of estradiol-17b in the oyster Crassostrea gigas. J. Exp. Mar. Biol. Ecol. 2005, 325, 125–133. [Google Scholar] [CrossRef]
- Bateman, M.E.; Strong, A.L.; McLachlan, J.A.; Burow, M.E.; Bunnell, B.A. The Effects of Endocrine Disruptors on Adipogenesis and Osteogenesis in Mesenchymal Stem Cells: A Review. Front. Endocrinol. 2017, 7, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daston, G.P.; Cook, J.C.; Kavlock, R.J. Uncertainties for Endocrine Disrupters: Our View on Progress. Toxicol. Sci. 2003, 74, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene * | Primer Sequence (5′-3′) | Size | Tm |
|---|---|---|---|
| ALP | F: 5′-TTGTGGCCCTCTCCAAGACA-3′ | 198 bp | 60 °C |
| R: 5′-GACTTCCCAGCATCCTTGGC-3′ | |||
| RUNX2 | F: 5′-CATGGTGGAGATCATCGCGG-3′ R: 5′-GGCCATGACGGTAACCACAG-3′ | 171 bp | 60 °C |
| OSX | F: 5′-AAGGCGGTTGGCAATAGTGG-3′ R: 5′-GCAGCTGTGAATGGGCTTCT-3′ | 194 bp | 60 °C |
| GAPDH | F: 5′-ACCACAGTCCATGCCATCAC-3′ | 480 bp | 63 °C |
| R: 5′-CACCACCCTGTTGCTGTAGC-3′ |
| Gene * | Primer Sequence (5′-3′) | Size | Tm |
|---|---|---|---|
| zRUNX2a | F: 5′-GACGGTGGTGACGGTAATGG-3′ | 173 bp | 58 °C |
| R: 5′-TGCGGTGGGTTCGTGAATA-3′ | |||
| zRUNX2b | F: 5′-CGGCTCCTACCAGTTCTCCA-3′ | 149 bp | 59 °C |
| R: 5′-CCATCTCCCTCCACTCCTCC-3′ | |||
| zOSX | F: 5′-GGCTATGCTAACTGCGACCTG-3′ | 153 bp | 56 °C |
| R: 5′-GCTTTCATTGCGTCCGTTTT-3′ | |||
| zALP | F: 5′-CAAGAACTCAACAAGAAC-3′ | 170 bp | 48 °C |
| R: 5′-TGAGCATTGGTGTTATAC-3′ | |||
| zCTSK | F: 5′-GGACTCAATCACTATCACT-3′ | 117 bp | 56 °C |
| R: 5′-AGAACAAGACATCTAAGACA-3′ | |||
| zNFATc-1 | F: 5′-AACCTTCCTCGTTCCCTCAA-3′ | 152 bp | 57 °C |
| R: 5′-CGCTGTTATCCTCCACCTCA-3′ | |||
| zRANK | F: 5′-GCACGGTTATTGTTGTTA-3′ | 109 bp | 49 °C |
| R: 5′-TATTCAGAGGTGGTGTTAT-3′ | |||
| zACP5b | F: 5′-GCTGCTGCTAACAAACAAT-3′ | 76 bp | 52 °C |
| R: 5′-GACCAACCACGATGACAA-3′ | |||
| zβ-actin | F: 5′-CGAGCGTGGCTACAGCTTCA-3′ | 155 bp | 60 °C |
| R: 5′-GACCGTCAGGCAGCTCATAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molagoda, I.M.N.; Athapaththu, A.M.G.K.; Park, E.K.; Choi, Y.H.; Jeon, Y.-J.; Kim, G.-Y. Fermented Oyster (Crassostrea gigas) Extract Cures and Prevents Prednisolone-Induced Bone Resorption by Activating Osteoblast Differentiation. Foods 2022, 11, 678. https://doi.org/10.3390/foods11050678
Molagoda IMN, Athapaththu AMGK, Park EK, Choi YH, Jeon Y-J, Kim G-Y. Fermented Oyster (Crassostrea gigas) Extract Cures and Prevents Prednisolone-Induced Bone Resorption by Activating Osteoblast Differentiation. Foods. 2022; 11(5):678. https://doi.org/10.3390/foods11050678
Chicago/Turabian StyleMolagoda, Ilandarage Menu Neelaka, Athapaththu Mudiyanselage Gihan Kavinda Athapaththu, Eui Kyun Park, Yung Hyun Choi, You-Jin Jeon, and Gi-Young Kim. 2022. "Fermented Oyster (Crassostrea gigas) Extract Cures and Prevents Prednisolone-Induced Bone Resorption by Activating Osteoblast Differentiation" Foods 11, no. 5: 678. https://doi.org/10.3390/foods11050678