
Phenolic Acid Profiling of Lactarius hatsudake Extracts, Anti-Cancer Function and Its Molecular Mechanisms


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Extraction and Purification
2.3. Total Phenolic Content
2.4. Analysis of Phenolic Profiles
2.5. Cell Culture and Polyphenol Treatments
2.6. Cell Viability Assay
2.7. Hoechst 33258 Fluorescent Staining
2.8. SDS–PAGE and Western Blot Analysis
2.9. Reverse Transcription-Quantitative PCR (RT-qPCR) Analysis
2.10. Cytometry Analysis
2.11. Apoptosis Detection
2.12. Luciferase Reporter Activity Assay
2.13. Statistical Analysis
3. Results and Discussion
3.1. The Contents and Composition of Total Phenols
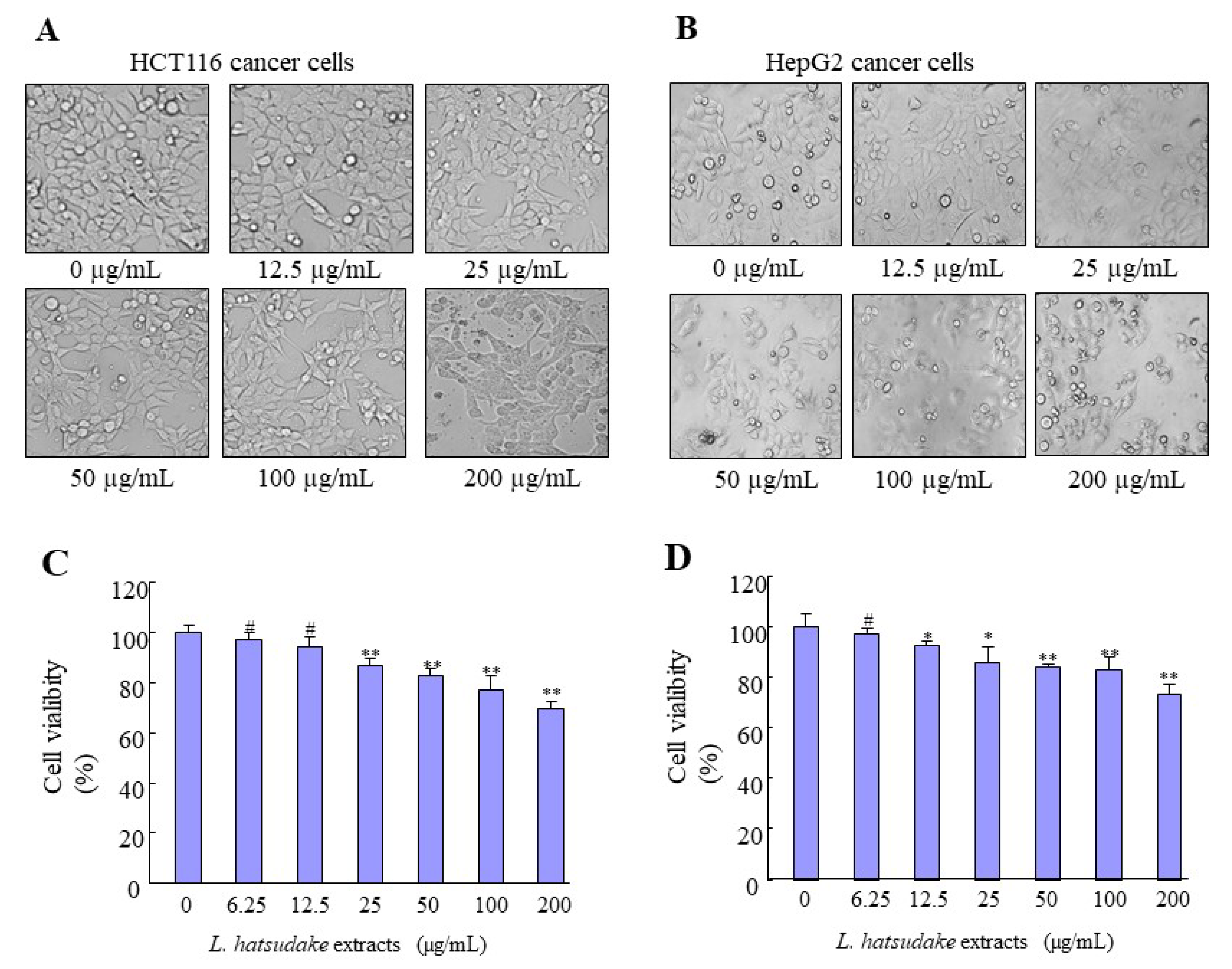
3.2. L. hatsudake Extracts Inhibited Cancer Cell Proliferation
3.3. L. hatsudake Extracts Arrest Cell Cycle of Cancer Cells
3.4. L. hatsudake Extracts Decreased CDKIs and Cyclin D1 Expressions
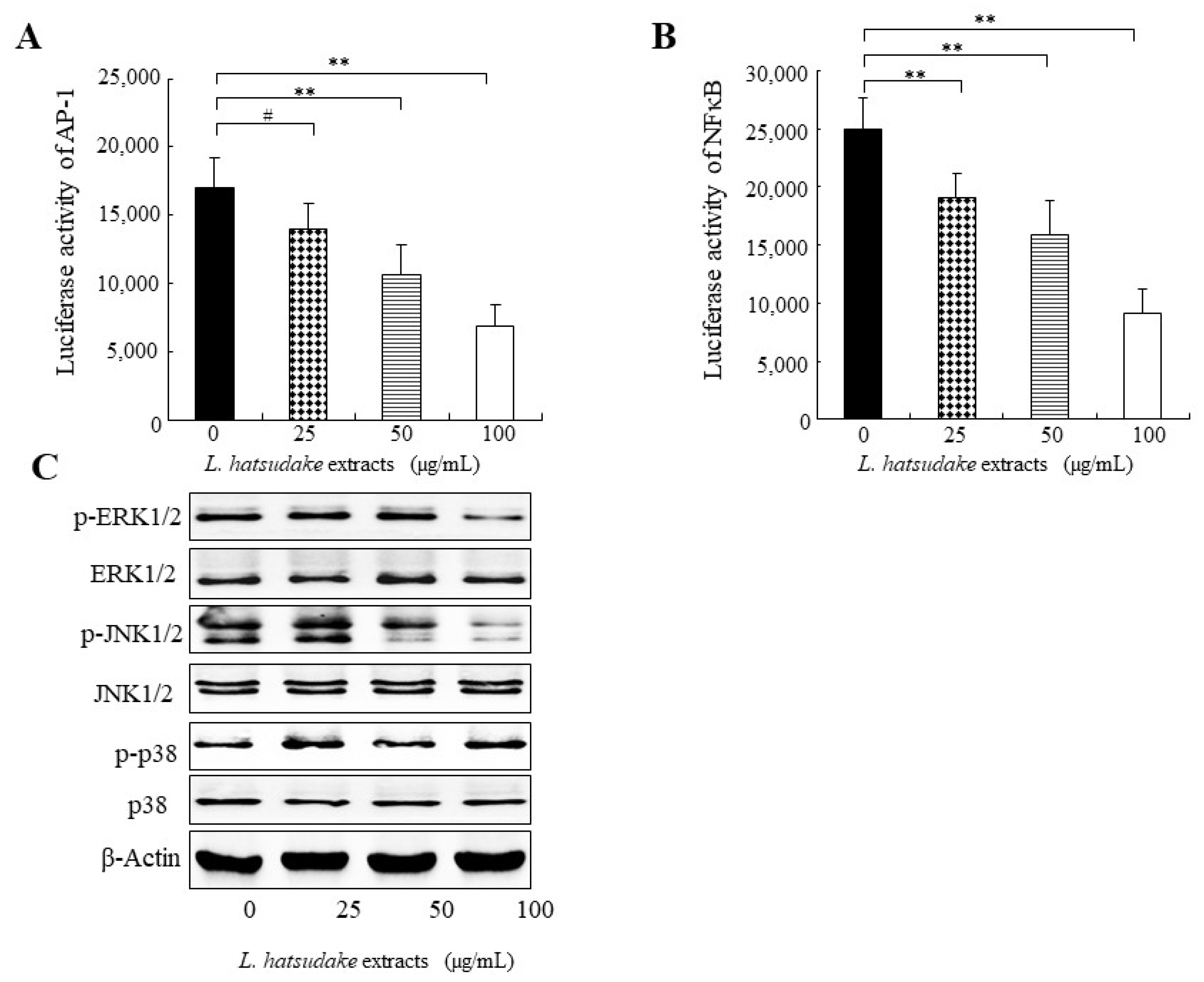
3.5. L. hatsudake Extracts Inhibited Transcript Activities of NFκB and AP-1
3.6. L. hatsudake Extracts Inhibited MAPK Signal Pathway
3.7. Pro-Apoptosis Effect and Mechanism of L. hatsudake Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [PubMed] [Green Version]
- Oueslati, S.; Ksouri, R.; Falleh, H.; Pichette, A.; Abdelly, C.; Legault, J. Phenolic content, antioxidant, anti-inflammatory and anticancer activities of the edible halophyte Suaeda fruticosa Forssk. Food Chem. 2012, 132, 943–947. [Google Scholar] [CrossRef]
- Roleira, F.M.; Tavares-da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant derived and dietary phenolic antioxidants: Anticancer properties. Food Chem. 2015, 183, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Guo, D.; Fang, L.; Sang, T.; Wu, J.; Guo, C.; Wang, Y.; Wang, Y.; Chen, C.; Chen, J.; et al. Ganoderma lucidum polysaccharide modulates gut microbiota and immune cell function to inhibit inflammation and tumorigenesis in colon. Carbohydr. Polym. 2021, 267, 118231. [Google Scholar] [CrossRef]
- Zvikas, V.; Urbanaviciute, I.; Bernotiene, R.; Kulakauskiene, D.; Morkunaite, U.; Balion, Z.; Majiene, D.; Liaudanskas, M.; Viskelis, P.; Jekabsone, A.; et al. Investigation of Phenolic Composition and Anticancer Properties of Ethanolic Extracts of Japanese Quince Leaves. Foods 2020, 10, 18. [Google Scholar] [CrossRef]
- Damazo-Lima, M.; Rosas-Perez, G.; Reynoso-Camacho, R.; Perez-Ramirez, I.F.; Rocha-Guzman, N.E.; de Los Rios, E.A.; Ramos-Gomez, M. Chemopreventive Effect of the Germinated Oat and its Phenolic-AVA Extract in Azoxymethane/Dextran Sulfate Sodium (AOM/DSS) Model of Colon Carcinogenesis in Mice. Foods 2020, 9, 169. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.-L.; Liu, L.-P.; Wang, M.; Gao, J.-M. Bioactive ergosterol derivatives isolated from the fungus Lactarius hatsudake. Chem. Nat. Compd. 2007, 43, 637–638. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, H.; Hao, S.; Pan, D.; Wang, G.; Yu, W. Evaluation of phenolic composition and antioxidant properties of different varieties of Chinese citrus. Food Chem. 2021, 364, 130413. [Google Scholar] [CrossRef]
- Bvenura, C.; Kambizi, L. Composition of Phenolic Compounds in South African Schinus molle L. Berries. Foods 2022, 11, 1376. [Google Scholar] [CrossRef]
- Lu, C.; Li, C.; Chen, B.; Shen, Y. Composition and antioxidant, antibacterial, and anti-HepG2 cell activities of polyphenols from seed coat of Amygdalus pedunculata Pall. Food Chem. 2018, 265, 111–119. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, J.; Herald, T.; Cox, S.; Noronha, L.; Perumal, R.; Lee, H.S.; Smolensky, D. Anticancer Activity of a Novel High Phenolic Sorghum Bran in Human Colon Cancer Cells. Oxidative Med. Cell. Longev. 2020, 2020, 2890536. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, M.; Wu, K.; Chai, X.; Yu, H.; Tao, Z.; Wang, J. Immunomodulatory and anticancer activities of phenolics from emblica fruit (Phyllanthus emblica L.). Food Chem. 2012, 131, 685–690. [Google Scholar] [CrossRef]
- Hamid, H.A.; Ramli, A.N.M.; Zamri, N.; Yusoff, M.M. UPLC-QTOF/MS-based phenolic profiling of Melastomaceae, their antioxidant activity and cytotoxic effects against human breast cancer cell MDA-MB-231. Food Chem. 2018, 265, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Stanikunaite, R.; Khan, S.I.; Trappe, J.M.; Ross, S.A. Cyclooxygenase-2 inhibitory and antioxidant compounds from the truffle Elaphomyces granulatus. Phytother. Res. 2009, 23, 575–578. [Google Scholar] [CrossRef]
- Cheung, L.M.; Cheung, P.C.K.; Ooi, V.E.C. Antioxidant activity and total phenolics of edible mushroom extracts. Food Chem. 2003, 81, 249–255. [Google Scholar] [CrossRef]
- Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant properties of phenolic compounds occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Taofiq, O.; Calhelha, R.C.; Heleno, S.; Barros, L.; Martins, A.; Santos-Buelga, C.; Queiroz, M.; Ferreira, I. The contribution of phenolic acids to the anti-inflammatory activity of mushrooms: Screening in phenolic extracts, individual parent molecules and synthesized glucuronated and methylated derivatives. Food Res. Int. 2015, 76, 821–827. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.M.; Angeloni, S.; Abouelenein, D.; Acquaticci, L.; Xiao, J.; Sagratini, G.; Maggi, F.; Vittori, S.; Caprioli, G. A new HPLC-MS/MS method for the simultaneous determination of 36 polyphenols in blueberry, strawberry and their commercial products and determination of antioxidant activity. Food Chem. 2022, 367, 130743. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Antioxidant constituents of three selected red and green color Amaranthus leafy vegetable. Sci. Rep. 2019, 9, 18233. [Google Scholar] [CrossRef] [Green Version]
- Pinto, D.; Vieira, E.F.; Peixoto, A.F.; Freire, C.; Freitas, V.; Costa, P.; Delerue-Matos, C.; Rodrigues, F. Optimizing the extraction of phenolic antioxidants from chestnut shells by subcritical water extraction using response surface methodology. Food Chem. 2021, 334, 127521. [Google Scholar] [CrossRef]
- Yahia, E.M.; Gutierrez-Orozco, F.; Moreno-Perez, M.A. Identification of phenolic compounds by liquid chromatography-mass spectrometry in seventeen species of wild mushrooms in Central Mexico and determination of their antioxidant activity and bioactive compounds. Food Chem. 2017, 226, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi Ekbatan, S.; Li, X.Q.; Ghorbani, M.; Azadi, B.; Kubow, S. Chlorogenic Acid and Its Microbial Metabolites Exert Anti-Proliferative Effects, S-Phase Cell-Cycle Arrest and Apoptosis in Human Colon Cancer Caco-2 Cells. Int. J. Mol. Sci. 2018, 19, 723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabiani, R.; Rosignoli, P.; De Bartolomeo, A.; Fuccelli, R.; Servili, M.; Morozzi, G. The production of hydrogen peroxide is not a common mechanism by which olive oil phenols induce apoptosis on HL60 cells. Food Chem. 2011, 125, 1249–1255. [Google Scholar] [CrossRef]
- Kohno, K.; Miyake, M.; Sano, O.; Tanaka-Kataoka, M.; Yamamoto, S.; Koya-Miyata, S.; Arai, N.; Fujii, M.; Watanabe, H.; Ushio, S.; et al. Anti-inflammatory and immunomodulatory properties of 2-amino-3H-phenoxazin-3-one. Biol. Pharm. Bull. 2008, 31, 1938–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnero, A.; Hannon, G.J. The INK4 family of CDK Inhibitors. Curr. Top. Microbiol. Immunol. 1998, 227, 43–55. [Google Scholar] [PubMed]
- Sherr, C.J.; Roberts, J.M. CDK inhibitor: Positive and negative regulators of G1-phase progression. Genes Dev. 1999, 13, 1501–1512. [Google Scholar] [CrossRef] [Green Version]
- Lowe, S.W. Cancer therapy and p53. Curr. Opin. Oncol. 1995, 7, 547–553. [Google Scholar] [CrossRef]
- Sellers, W.R.; Kaelin, W.G., Jr. Role of the retinoblastoma protein in the pathogenesis of human cancer. J. Clin. Oncol. 1997, 15, 3301–3312. [Google Scholar] [CrossRef]
- Santarpia, L.; Lippman, S.M.; El-Naggar, A.K. Targeting the MAPK-RAS-RAF signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 103–119. [Google Scholar]
- Lee, K.C.; Chen, W.T.; Liu, Y.C.; Lin, S.S.; Hsu, F.T. Amentoflavone Inhibits Hepatocellular Carcinoma Progression Through Blockage of ERK/NF-kB Activation. In Vivo 2018, 32, 1097–1103. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.H.; Liu, X.; Shen, M.Y.; Nie, S.P.; Zhang, H.; Li, C.; Gong, D.M.; Xie, M.Y. Purification, physicochemical characterization and anticancer activity of a polysaccharide from Cyclocarya paliurus leaves. Food Chem. 2013, 136, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, U.; Szewczyk, K.; Owczarek, K.; Hrabec, Z.; Podsedek, A.; Koziolkiewicz, M.; Hrabec, E. Flavanols from Japanese quince (Chaenomeles japonica) fruit inhibit human prostate and breast cancer cell line invasiveness and cause favorable changes in Bax/Bcl-2 mRNA ratio. Nutr. Cancer 2013, 65, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Cincin, Z.B.; Unlu, M.; Kiran, B.; Bireller, E.S.; Baran, Y.; Cakmakoglu, B. Apoptotic Effects of Quercitrin on DLD-1 Colon Cancer Cell Line. Pathol. Oncol. Res. 2015, 21, 333–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, G.J.; Jin, X.L.; Qian, Y.P.; Wang, Q.; Yang, R.T.; Dai, F.; Tang, J.J.; Shang, Y.J.; Cheng, L.X.; Yang, J.; et al. Hydroxycinnamic acids as DNA-cleaving agents in the presence of Cu(II) ions: Mechanism, structure-activity relationship, and biological implications. Chemistry 2009, 15, 12889–12899. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Zhao, M.; Yang, B.; Ren, J.; Shen, G.; Rao, G. Antioxidant and antiproliferative capacities of phenolics purified from Phyllanthus emblica L. fruit. Food Chem. 2011, 126, 277–282. [Google Scholar] [CrossRef]
- Tang, D.; Wu, D.; Hirao, A.; Lahti, J.M.; Liu, L.; Mazza, B.; Kidd, V.J.; Mak, T.W.; Ingram, A.J. ERK activation mediates cell cycle arrest and apoptosis after DNA damage independently of p53. J. Biol. Chem. 2002, 277, 12710–12717. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward | Reverse |
|---|---|---|
| p15 | 5′ GTT GTT TGG TTA TTG TAT GGG 3′ | 5′ CCC TTA TTC TCC TCA CAC AT 3′ |
| p16 | 5′ CCC AAC GCA CCG AAT AGT TAC 3′ | 5′ GTT CTT TCA ATC GGG GAT GTC 3′ |
| p18 | 5′ GGG GAC CTA GAG CAA CTT ACT AGT TT 3′ | 5′ AAA TCG GGA TTA GCA CCT CTA AGT A 3′ |
| p21 | 5′ ATG TGG ACC TGT CAC TGT CTT GTA 3′ | 5′ GTT GGA GTG GTA GAA ATC TGT CAT 3′ |
| p27 | 5′ AGT GGA TGA TGA GAT TGT GGA GTT 3′ | 5′ AAC AAG TCT AAG CTG GTG TTT TTC C 3′ |
| p53 | 5′ CCC AAG CAA TGG ATG ATT TGA 3′ | 5′ GGC ATT CTG GGA GCT TCA TCT 3′ |
| β-actin | 5′ CAT GTA CGT TGC TAT CCA GGC 3′ | 5′ CTC CTT AAT GTC ACG CAC GAT 3′ |
| Parent Ion | Sub-Ion Ion | |
|---|---|---|
| Chlorogenic acid | 353.15 | 96.70 |
| Ferulic acid | 193.04 | 148.83 |
| Caffeic acid | No detected | No detected |
| Gallic acid | 187.08 | 124.82 |
| Pyrogallol | 125.08 | 97.62 |
| Cinnamic acid | 147.11 | 120.78 |
| Myricetin | 317.08 | 272.76 |
| Substance-1 | 264.97 | 96.74, 79.72 |
| Substance-2 | 187.04 | 124.69 |
| Substance-3 | 278.77 | 261.09 |
| Substance-4 | 309.13 | 96.89, 290.78, 208.70 |
| Substance-5 | 433.18 | 152.77, 78.77, 170.47 |
| Substance-6 | 295.02 | 276.85, 156.70, 182.72 |
| Substance-7 | 149.87 | 95.69, 107.38, 122.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Zhang, X.; Qin, H.; Luo, F.; Ren, J. Phenolic Acid Profiling of Lactarius hatsudake Extracts, Anti-Cancer Function and Its Molecular Mechanisms. Foods 2022, 11, 1839. https://doi.org/10.3390/foods11131839
Yang Q, Zhang X, Qin H, Luo F, Ren J. Phenolic Acid Profiling of Lactarius hatsudake Extracts, Anti-Cancer Function and Its Molecular Mechanisms. Foods. 2022; 11(13):1839. https://doi.org/10.3390/foods11131839
Chicago/Turabian StyleYang, Qiao, Xiaoyi Zhang, Huini Qin, Feijun Luo, and Jiali Ren. 2022. "Phenolic Acid Profiling of Lactarius hatsudake Extracts, Anti-Cancer Function and Its Molecular Mechanisms" Foods 11, no. 13: 1839. https://doi.org/10.3390/foods11131839