
Transcriptomic and Metabolomic Analyses Providing Insights into the Coloring Mechanism of Docynia delavayi

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Storage
2.2. Quantification of Total Flavonoids and Anthocyanins
2.3. Metabolite Identification and Data Analysis
2.3.1. Sample Preparation and Extraction
2.3.2. UPLC Conditions
2.3.3. ESI-Q TRAP-MS/MS
2.3.4. Statistical Analysis
2.4. RNA-seq, Annotation, and Data Analysis
2.5. qRT-PCR
3. Results
3.1. Appearance Characteristic of D. delavayi Fruits, and Quantification of Total Flavonoids and Anthocyanins
3.2. Metabolome Data Quality Analysis
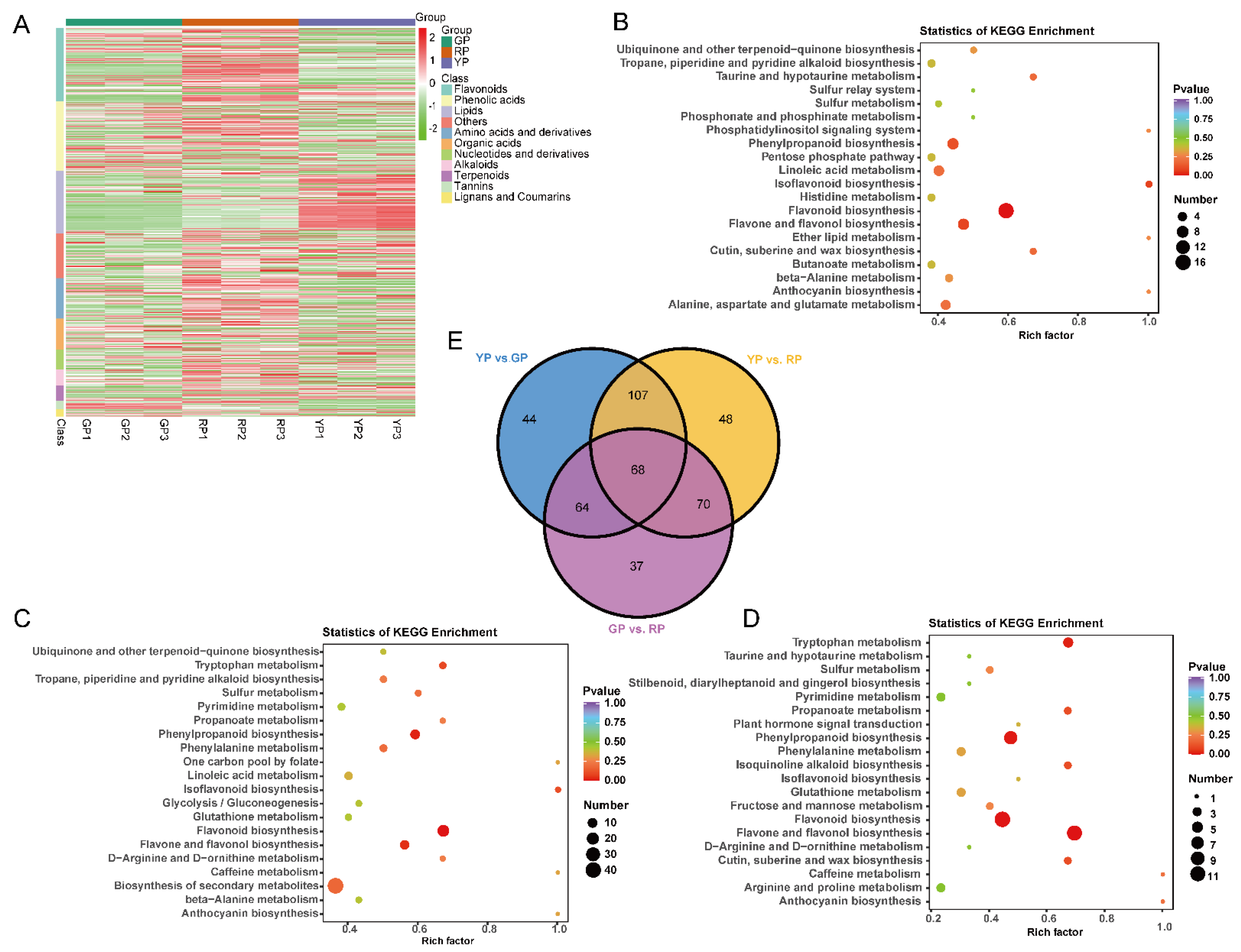
3.3. The Identification of Differentially Accumulated Metabolites (DAMs) in the Pericarp of Three Different Variations
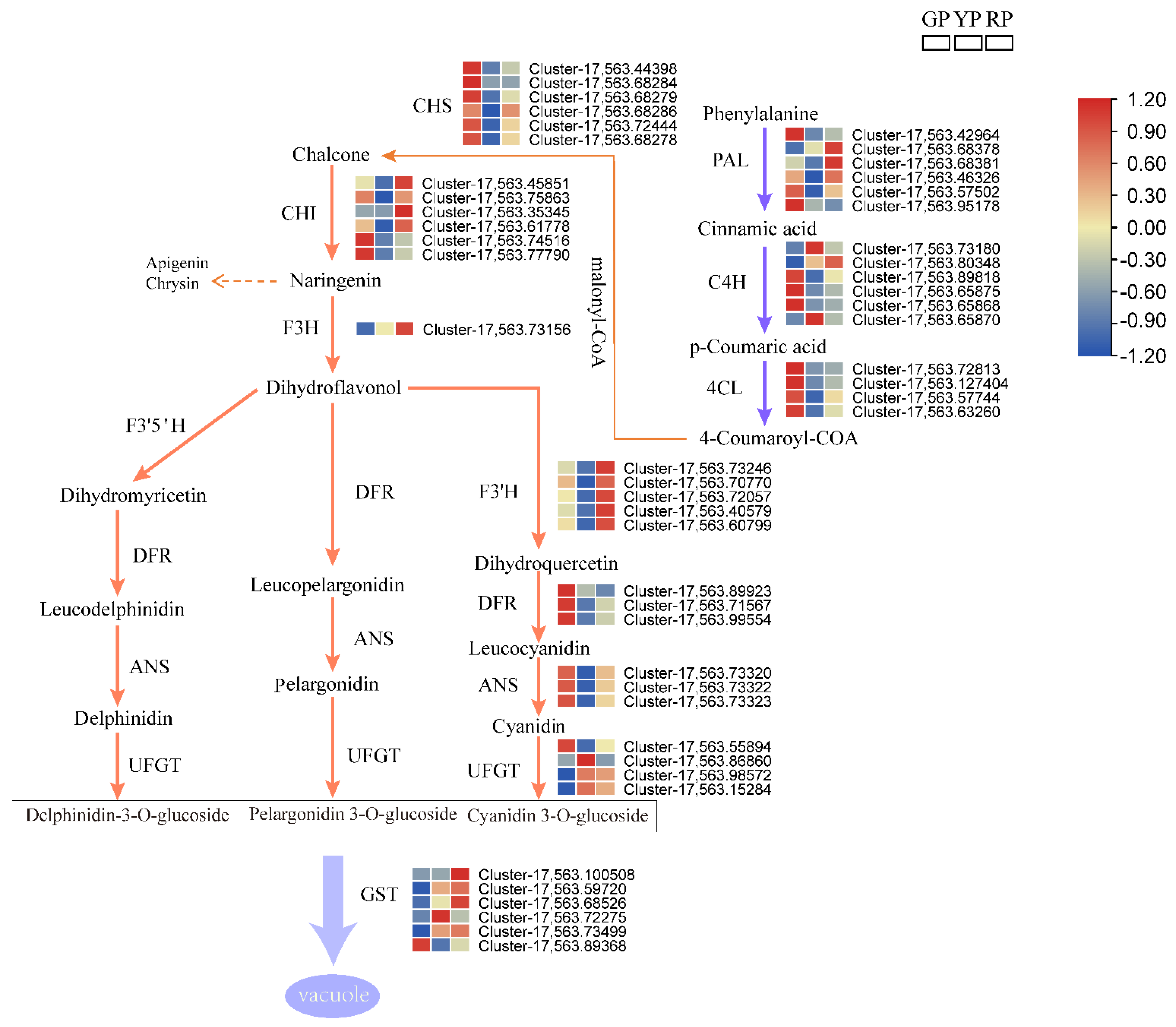
3.4. Analysis of Anthocyanins in Three Different Variations of D. delavayi
3.5. Transcriptome Sequencing and Annotation
3.6. GO Enrichment and KEGG Pathway Analyses of DEGs
3.7. Identification of TFs Related to Anthocyanin Biosynthesis
3.8. Analysis of Structural Genes Involved in Anthocyanin Biosynthesis and Gene Expression through qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Yi, K.; Chen, J.; Li, R.P.; Xie, J.; Jin, Y.; Mei, X.R.; Li, Y.J.; Liu, G.; Wang, Z.G. Purified phlorizin from DocynIa Indica (Wall.) Decne by HSCCC, compared with whole extract, phlorizin and non-phlorizin fragment ameliorate obesity, insulin resistance, and improves intestinal barrier function in high-fat-diet-fed mice. Molecules 2018, 23, 2701. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Shu, G.; Chen, L.; Mi, X.; Mei, Z.; Deng, X. A flavonoid component from Docynia delavayi (Franch.) Schneid represses transplanted H22 hepatoma growth and exhibits low toxic effect on tumor-bearing mice. Food Chem. Toxicol. 2012, 50, 3166–3173. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Reagan, K.M.; Machnicki, N.J.; Carlo, T.A.; Haak, D.C.; Peñaloza, A.L.; Levey, D.J. Evolutionary ecology of pungency in wild chilies. Proc. Natl. Acad. Sci. USA 2008, 105, 11808–11811. [Google Scholar] [CrossRef] [PubMed]
- Zamljen, T.; Jakopič, J.; Hudina, M.; Veberič, R.; Slatnar, A. Influence of intra and inter species variation in chilies (Capsicum spp.) on metabolite composition of three fruit segments. Sci. Rep. 2021, 11, 4932. [Google Scholar] [CrossRef] [PubMed]
- Fathordoobady, F.; Mirhosseini, H.; Selamat, J.; Manap, M.Y. Effect of solvent type and ratio on betacyanins and antioxidant activity of extracts from Hylocereus polyrhizus flesh and peel by supercritical fluid extraction and solvent extraction. Food Chem. 2016, 202, 70–80. [Google Scholar] [CrossRef]
- Tuan, P.A.; Bai, S.; Yaegaki, H.; Tamura, T.; Hihara, S.; Moriguchi, T.; Oda, K. The crucial role of PpMYB10.1 in anthocyanin accumulation in peach and relationships between its allelic type and skin color phenotype. BMC Plant Biol. 2015, 15, 280. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yu, L.; He, L.Z.; Zhu, L.Y.; Xu, S.; Wan, Y.H.; Wang, H.; Wang, Y.; Zhu, W.M. Comparative transcriptome analysis of celery leaf blades identified an R2R3-MYB transcription factor that regulates apigenin metabolism. J. Agric. Food Chem. 2019, 67, 5265–5277. [Google Scholar] [CrossRef]
- Qiao, Z.Q.; Liu, S.S.; Zeng, H.J.; Li, Y.X.; Wang, X.Y.; Chen, Y.; Wang, X.M.; Cai, N. Exploring the molecular mechanism underlying the stable purple-red leaf phenotype in Lagerstroemia indica cv. Ebony embers. Int. J. Mol. Sci. 2019, 20, 5636. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Vimolmangkang, S.; SoriaGuerra, R.E.; Korban, S.S. Introduction of apple ANR genes into tobacco inhibits expression of both CHI and DFR genes in flowers, leading to loss of anthocyanin. J. Exp. Bot. 2012, 63, 2437–2447. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Lei, Y.J.; Chen, H.W.; Li, J.N.; Chen, H.Z.; Zhang, Z.W. R2R3-MYB transcription factors regulate anthocyanin biosynthesis in grapevine vegetative tissues. Front. Plant Sci. 2020, 11, 527. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Z.; Zeng, Q.W.; Wang, S.B.; Luo, Y.W.; Huang, Y.; Xin, Y.C.; He, N.J. Abnormal expression of bHLH3 disrupts a flavonoid homeostasis network, causing differences in pigment composition among mulberry fruits. Hortic. Res. 2020, 7, 83. [Google Scholar] [CrossRef]
- Yang, F.S.; Nie, S.; Liu, H.; Shi, T.L.; Tian, X.C.; Zhou, S.S.; Bao, Y.T.; Jia, K.H.; Guo, J.F.; Zhao, W.; et al. Chromosome-level genome assembly of a parent species of widely cultivated azaleas. Nat. Commun. 2020, 11, 5269. [Google Scholar] [CrossRef]
- Jiang, S.H.; Chen, M.; He, N.B.; Chen, X.L.; Wang, N.; Sun, Q.G.; Zhang, T.L.; Xu, H.F.; Fang, H.C.; Wang, Y.C.; et al. MdGSTF6, activated by MdMYB1, plays an essential role in anthocyanin accumulation in apple. Hortic. Res. 2019, 6, 40. [Google Scholar] [CrossRef]
- Yang, Y.; Cui, B.H.; Tan, Z.W.; Song, B.X.; Cao, H.N.; Zong, C.W. RNA sequencing and anthocyanin synthesis-related genes expression analyses in white-fruited Vaccinium uliginosum. BMC Genom. 2018, 19, 930. [Google Scholar] [CrossRef]
- Zheng, T.; Tan, W.R.; Yang, H.; Zhang, L.; Li, T.T.; Liu, B.H.; Zhang, D.W.; Lin, H.H. Regulation of anthocyanin accumulation via MYB75/HAT1/TPL-mediated transcriptional repression. PLoS Genet. 2019, 15, e1007993. [Google Scholar] [CrossRef]
- Shah, J.; Brock, G.N.; Gaskins, J. BayesMetab: Treatment of missing values in metabolomic studies using a Bayesian modeling approach. BMC Bioinform. 2019, 20, 673. [Google Scholar] [CrossRef]
- Gao, J.; Li, W.B.; Liu, H.F.; Chen, F.B. De novo transcriptome sequencing of radish (Raphanus sativus L.) fleshy roots: Analysis of major genes involved in the anthocyanin synthesis pathway. BMC Mol. Cell Biol. 2019, 20, 45. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Cheng, Y.W.; Ya, H.Y.; Xu, S.Z.; Han, J.M. Transcriptome sequencing of purple petal spot region in tree peony reveals differentially expressed anthocyanin structural genes. Front. Plant Sci. 2015, 6, 964. [Google Scholar] [CrossRef]
- Xu, H.; Zou, Q.; Yang, G.; Jiang, S.; Fang, H.; Wang, Y.; Zhang, J.; Zhang, Z.; Wang, N.; Chen, X. MdMYB6 regulates anthocyanin formation in apple both through direct inhibition of the biosynthesis pathway and through substrate removal. Hortic. Res. 2020, 7, 72. [Google Scholar] [CrossRef]
- Li, C.; Wu, J.; Hu, K.D.; Wei, S.W.; Sun, H.Y.; Hu, L.Y.; Han, Z.; Yao, G.F.; Zhang, H. PyWRKY26 and PybHLH3 cotargeted the PyMYB114 promoter to regulate anthocyanin biosynthesis and transport in red-skinned pears. Hortic. Res. 2020, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, L.; Wang, Z.; Liu, Z.; Liu, P. Variations of the nutritional composition of jujube fruit ( Ziziphus jujuba Mill.) during maturation stages. Int. J. Food Prop. 2020, 23, 1066–1081. [Google Scholar] [CrossRef]
- Li, F.; Wu, B.; Yan, L.; Qin, X.; Lai, J. Metabolome and transcriptome profiling of Theobroma cacao provides insights into the molecular basis of pod color variation. J. Plant Res. 2021, 134, 1323–1334. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.L.; Wang, W.S.; Zhang, H.Y.; Liu, X.Q.; Yu, S.B.; Xiong, L.Z.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Yi, D.B.; Zhang, H.N.; Lai, B.; Liu, L.Q.; Pan, X.L.; Ma, Z.L.; Wang, Y.C.; Xie, J.H.; Shi, S.Y.; Wei, Y.Z. Integrative analysis of the coloring mechanism of red longan pericarp through metabolome and transcriptome analyses. J. Agric. Food Chem. 2021, 69, 1806–1815. [Google Scholar] [CrossRef]
- Ma, Y.R.; Yang, M.N.; Wang, J.J.; Jiang, C.Z.; Wang, Q.G. Application of exogenous ethylene inhibits postharvest peel browning of ‘Huangguan’ pear. Front. Plant Sci. 2017, 7, 2029. [Google Scholar] [CrossRef]
- Zhou, X.Z.; Xue, Y.B.; Mao, M.Q.; He, Y.H.; Adjei, M.O.; Yang, W.; Hu, H.; Liu, J.W.; Feng, L.J.; Zhang, H.L.; et al. Metabolome and transcriptome profiling reveals anthocyanin contents and anthocyanin-related genes of chimeric leaves in Ananas comosus var. bracteatus. BMC Genom. 2021, 22, 331. [Google Scholar] [CrossRef]
- Lang, X.; Li, N.; Li, L.; Zhang, S. Integrated metabolome and transcriptome analysis uncovers the role of anthocyanin metabolism in Michelia maudiae. Int. J. Genom. 2019, 2019, 4393905. [Google Scholar] [CrossRef]
- Tong, Y.Q.; Deng, H.T.; Kong, Y.W.; Tan, C.; Chen, J.; Wan, M.Z.; Wang, M.Y.; Yan, T.C.; Meng, X.J.; Li, L. Stability and structural characteristics of amylopectin nanoparticle-binding anthocyanins in Aronia melanocarpa. Food Chem. 2020, 311, 125687. [Google Scholar] [CrossRef]
- Strygina, K.V.; Kochetov, A.V.; Khlestkina, E.K. Genetic control of anthocyanin pigmentation of potato tissues. BMC Genet. 2019, 20 (Suppl. 1), 27. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Huhman, D.; Shadle, G.; He, X.Z.; Sumner, L.W.; Tang, Y.; Dixon, R.A. MATE2 mediates vacuolar sequestration of flavonoid glycosides and glycoside malonates in Medicago truncatula. Plant Cell 2011, 23, 1536–1555. [Google Scholar] [CrossRef]
- Zhao, J.R.; Shen, F.; Gao, Y.; Wang, D.J.; Wang, K. Parallel bud mutation sequencing reveals that fruit sugar and acid metabolism potentially influence stress in malus. Int J. Mol. Sci. 2019, 20, 5988. [Google Scholar] [CrossRef]
- Li, T.T.; Fan, Y.F.; Qin, H.; Dai, G.L.; Li, G.X.; Li, Y.L.; Wang, J.J.; Yin, Y.; Chen, F.; Qin, X.Y.; et al. Transcriptome and flavonoids metabolomic analysis identifies regulatory networks and hub genes in black and white fruits of Lycium ruthenicum murray. Front. Plant Sci. 2020, 11, 1256. [Google Scholar] [CrossRef]
- Karppinen, K.; Tegelberg, P.; Häggman, H.; Jaakola, L. Abscisic acid regulates anthocyanin biosynthesis and gene expression associated with cell wall modification in ripening bilberry (Vaccinium myrtillus L.) fruits. Front. Plant Sci. 2018, 9, 1259. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, J.B.; Yoon, Y.H.; Kim, S.H.; Ahn, J.W.; Jeong, I.Y.; Kang, S.Y.; Seo, Y.W.; Kim, D.S. The effects of chronic gamma irradiation on oxidative stress response and the expression of anthocyanin biosynthesis-related genes in wheat (Triticum aestivum). Int. J. Radiat. Biol. 2014, 90, 1218–1228. [Google Scholar] [CrossRef]
- Han, Y.H.; Huang, K.Y.; Liu, Y.J.; Jiao, T.M.; Ma, G.L.; Qian, Y.M.; Wang, P.Q.; Dai, X.L.; Gao, L.P.; Xia, T. Functional analysis of two Flavanone-3-Hydroxylase genes from camellia sinensis: A critical role in flavonoid accumulation. Genes 2017, 8, 300. [Google Scholar] [CrossRef]
- Jin, X.H.; Huang, H.; Wang, L.; Sun, Y.; Dai, S.L. Transcriptomics and metabolite analysis reveals the molecular mechanism of anthocyanin biosynthesis branch pathway in different Senecio cruentus cultivars. Front. Plant Sci. 2016, 7, 1307. [Google Scholar] [CrossRef]
- Zhou, C.B.; Mei, X.; Rothenberg, D.O.N.; Yang, Z.B.; Zhang, W.T.; Wan, S.H.; Yang, H.J.; Zhang, L.Y. Metabolome and transcriptome analysis reveals putative genes involved in anthocyanin accumulation and coloration in white and pink tea (Camellia sinensis) flower. Molecules 2020, 25, 190. [Google Scholar] [CrossRef]
- Muñoz, C.; Gomez-Talquenca, S.; Chialva, C.; Ibáñez, J.; Martinez-Zapater, J.M.; Peña-Neira, Á.; Lijavetzky, D. Relationships among gene expression and anthocyanin composition of Malbec grapevine clones. J. Agric. Food Chem. 2014, 62, 6716–6725. [Google Scholar] [CrossRef]
- Lotkowska, M.E.; Tohge, T.; Fernie, A.R.; Xue, G.P.; Balazadeh, S.; Mueller-Roeber, B. The Arabidopsis transcription factor MYB112 promotes anthocyanin formation during salinity and under high light stress. Plant Physiol. 2015, 169, 1862–1880. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.F.; Zhang, M.X.; Zhang, G.J.; Li, P.M.; Ma, F.W. Differential regulation of anthocyanin synthesis in apple peel under different sunlight intensities. Int. J. Mol. Sci. 2019, 20, 6060. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Cheng, Y.W.; Xu, S.Z.; Ma, H.P.; Han, J.M.; Zhang, Y. Tree peony variegated flowers show a small insertion in the F3′H gene of the acyanic flower parts. BMC Plant Biol. 2020, 20, 211. [Google Scholar] [CrossRef]
- Liu, H.L.; Lou, Q.; Ma, J.R.; Su, B.B.; Gao, Z.Z.; Liu, Y.L. Cloning and functional characterization of dihydroflavonol 4-reductase gene involved in anthocyanidin biosynthesis of Grape Hyacinth. Int. J. Mol. Sci. 2019, 20, 4743. [Google Scholar] [CrossRef]
- Katsu, K.; Suzuki, R.; Tsuchiya, W.; Inagaki, N.; Yamazaki, T.; Hisano, T.; Yasui, Y.; Komori, T.; Koshio, M.; Kubota, S.; et al. A new buckwheat dihydroflavonol 4-reductase (DFR), with a unique substrate binding structure, has altered substrate specificity. BMC Plant Biol. 2017, 17, 239. [Google Scholar] [CrossRef]
- Hartmann, A.; Patz, C.D.; Andlauer, W.; Dietrich, H.; Ludwig, M. Influence of processing on quality parameters of strawberries. J. Agric. Food Chem. 2008, 56, 9484–9489. [Google Scholar] [CrossRef]
- Zhang, X.D.; Allan, A.C.; Chen, X.Q.; Fan, L.; Chen, L.M.; Shu, Q.; Su, J.; Li, K.Z. Coloration, anthocyanin profile and metal element content of Yunnan Red Pear (Pyrus pyrifolia). Hortic. Sci. 2012, 39, 164–171. [Google Scholar] [CrossRef]
- Ban, Y.; Kondo, S.; Ubi, B.E.; Honda, C.; Bessho, H.; Moriguchi, T. UDP-sugar biosynthetic pathway: Contribution to cyanidin 3-galactoside biosynthesis in apple skin. Planta 2009, 230, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yuan, Y.; Tang, Z.Z.; Huang, Y.; Kang, C.Y.; Deng, X.X.; Xu, Q. Retrotransposon promoter of Ruby1 controls both light- and cold-induced accumulation of anthocyanins in blood orange. Plant Cell Environ. 2019, 42, 3092–3104. [Google Scholar] [CrossRef]
- Zhang, D.L.; Quantick, P.C.; Grigor, J.M. Changes in phenolic compounds in Litchi (Litchi chinensis Sonn.) fruit during postharvest storage. Postharvest Biol. Technol. 2000, 19, 165–172. [Google Scholar] [CrossRef]
- Zhou, H.; Peng, Q.; Zhao, J.B.; Owiti, A.; Ren, F.; Liao, L.; Wang, L.; Deng, X.B.; Jiang, Q.; Han, Y.P. Multiple R2R3-MYB transcription factors involved in the regulation of anthocyanin accumulation in peach flower. Front. Plant Sci. 2016, 7, 1557. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Ion Abundance | Fold Change | ||||
|---|---|---|---|---|---|---|
| GP | YP | RP | GP vs. RP | YP vs. GP | YP vs. RP | |
| Cyanidin-3-O-galactoside | 4.55 × 105 | 1.96 × 107 | 1.80 × 108 | 396.071 | 0.023 | 9.188 |
| Cyanidin-3-O-glucoside | 3.89 × 105 | 2.17 × 107 | 1.74 × 108 | 445.847 | 0.018 | 7.977 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Song, Y.; Wang, D. Transcriptomic and Metabolomic Analyses Providing Insights into the Coloring Mechanism of Docynia delavayi. Foods 2022, 11, 2899. https://doi.org/10.3390/foods11182899
Wang Y, Song Y, Wang D. Transcriptomic and Metabolomic Analyses Providing Insights into the Coloring Mechanism of Docynia delavayi. Foods. 2022; 11(18):2899. https://doi.org/10.3390/foods11182899
Chicago/Turabian StyleWang, Yuchang, Yuyang Song, and Dawei Wang. 2022. "Transcriptomic and Metabolomic Analyses Providing Insights into the Coloring Mechanism of Docynia delavayi" Foods 11, no. 18: 2899. https://doi.org/10.3390/foods11182899