Optical Proxies of Euxinia: Spectroscopic Studies of CDOM, Chlorophyll, and Bacteriochlorophylls in the Lagoon on Zeleny Cape (the White Sea)



Abstract
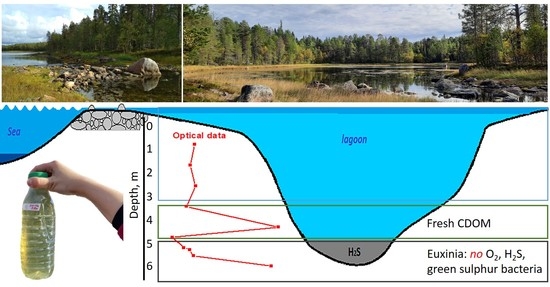
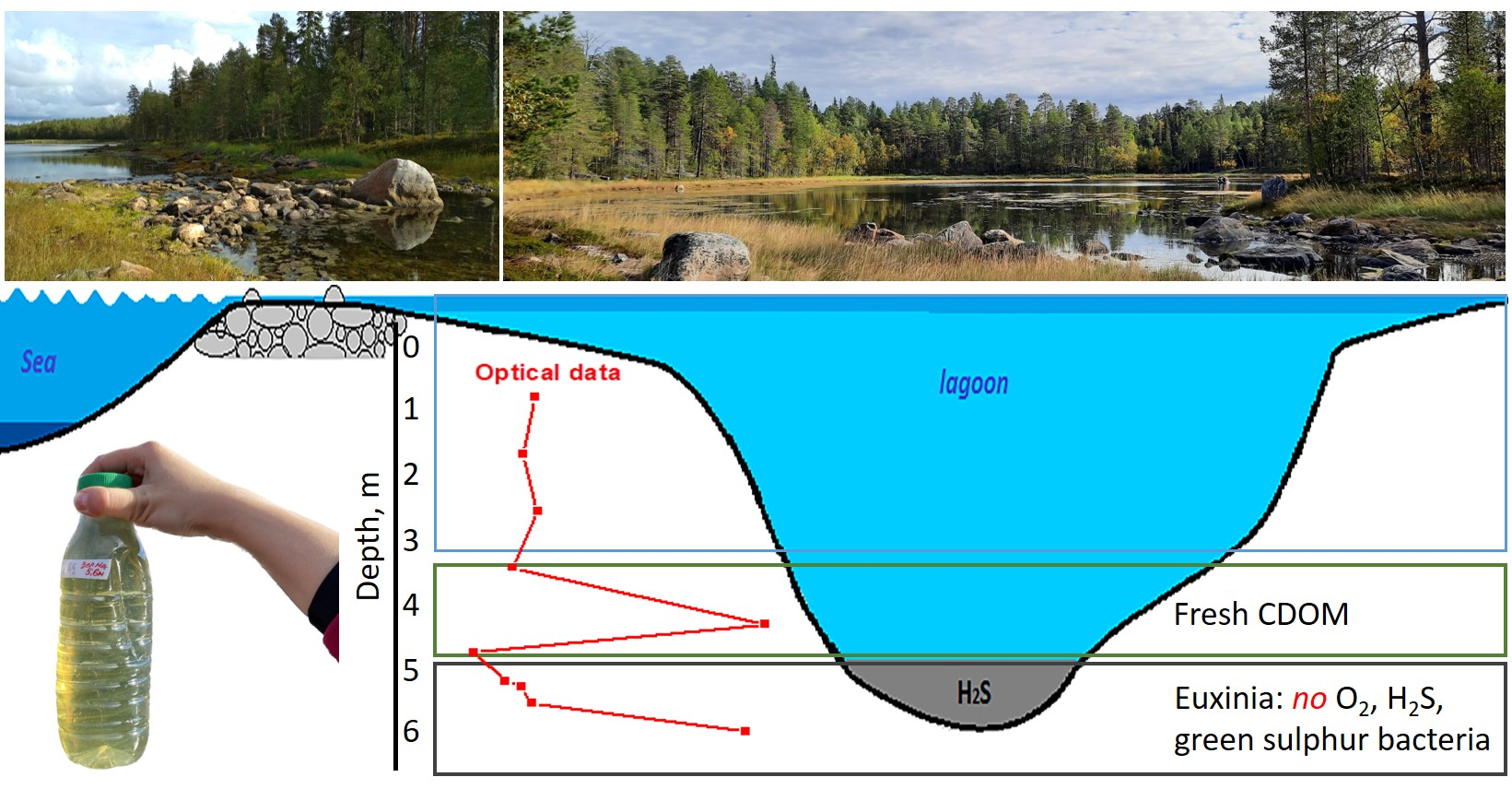
:
1. Introduction
2. Material and Methods
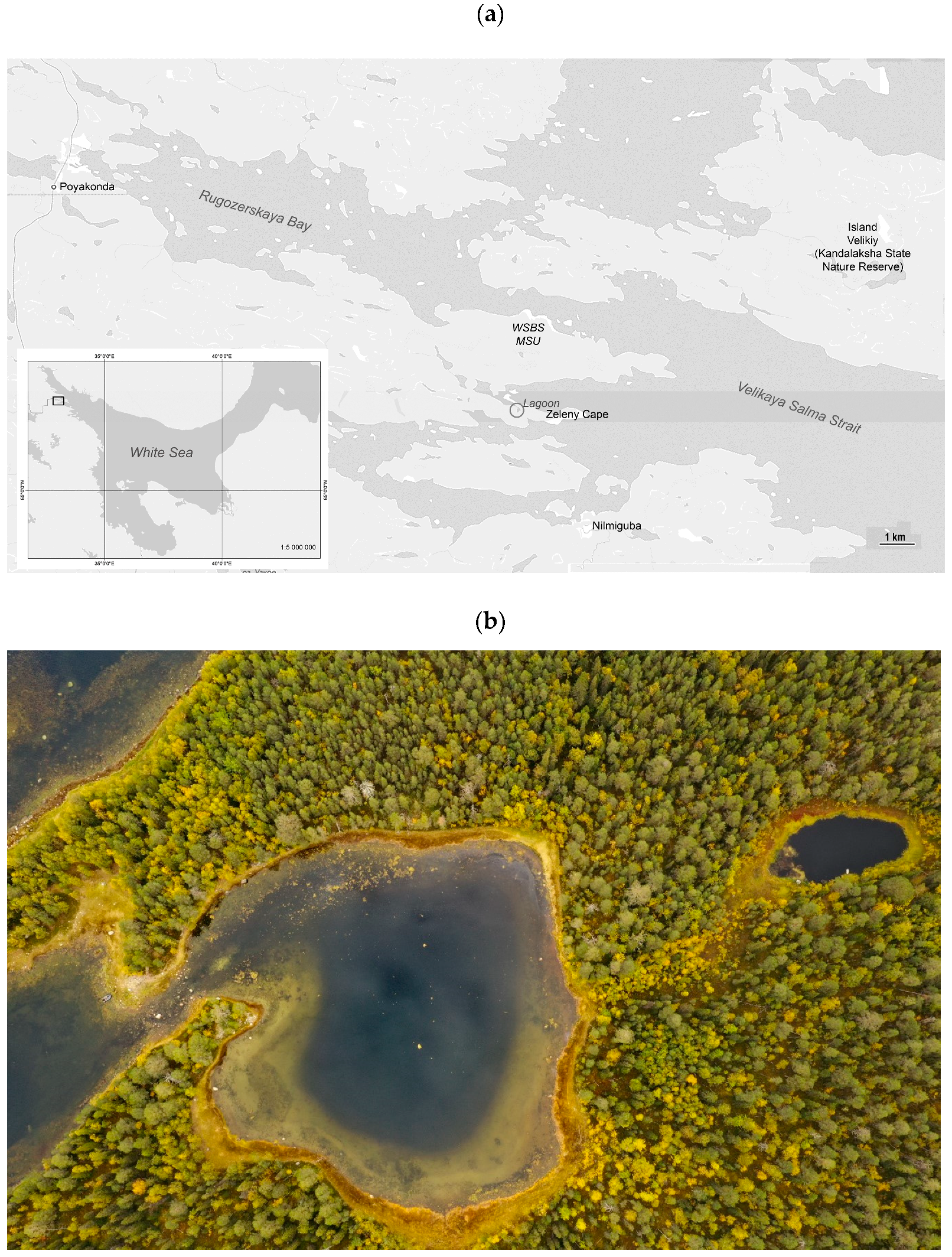
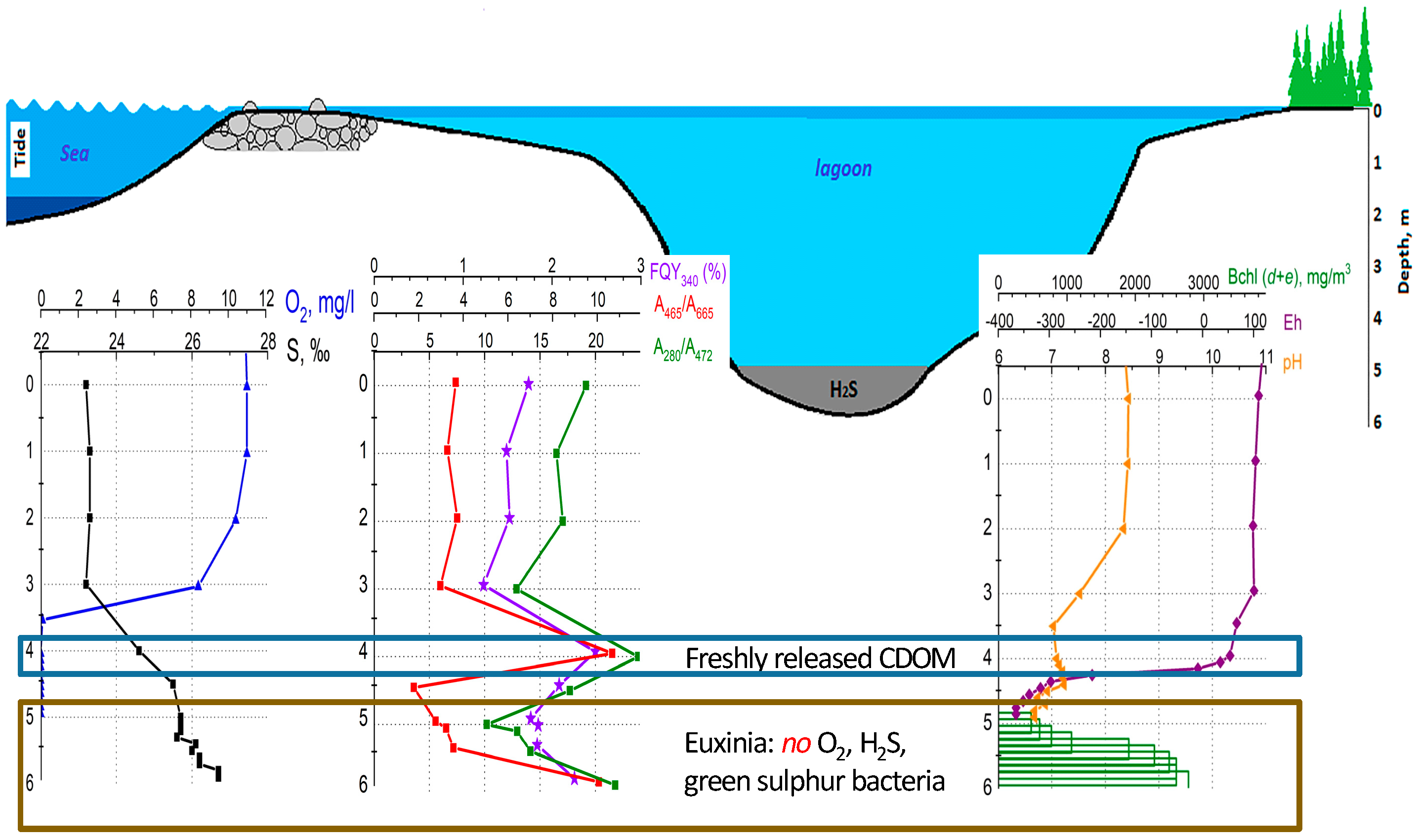
2.1. Lagoon on Zeleny Cape (Karelian Coast of the White Sea)
2.2. Water Sampling and Spectral Measurements
3. Results
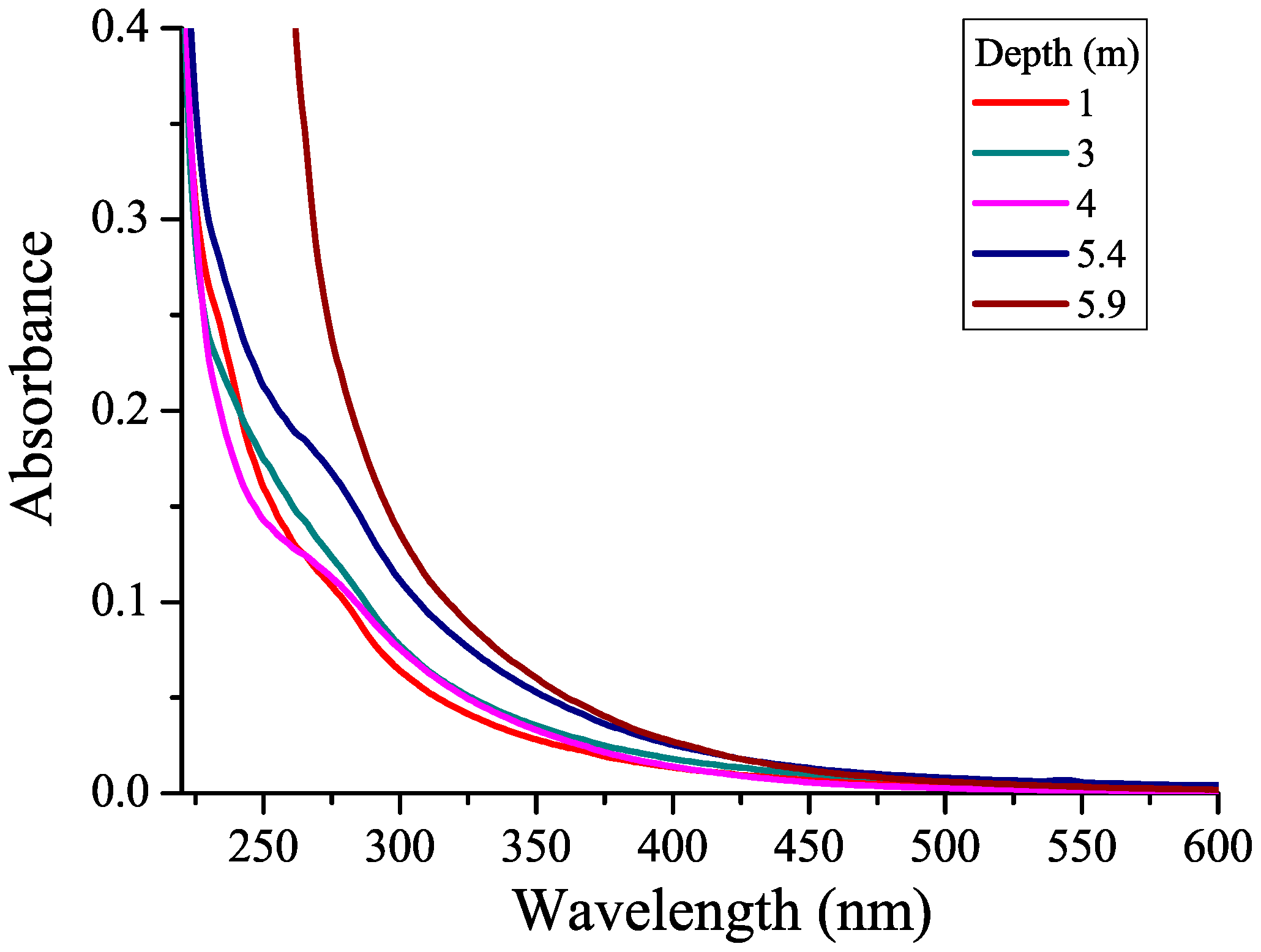
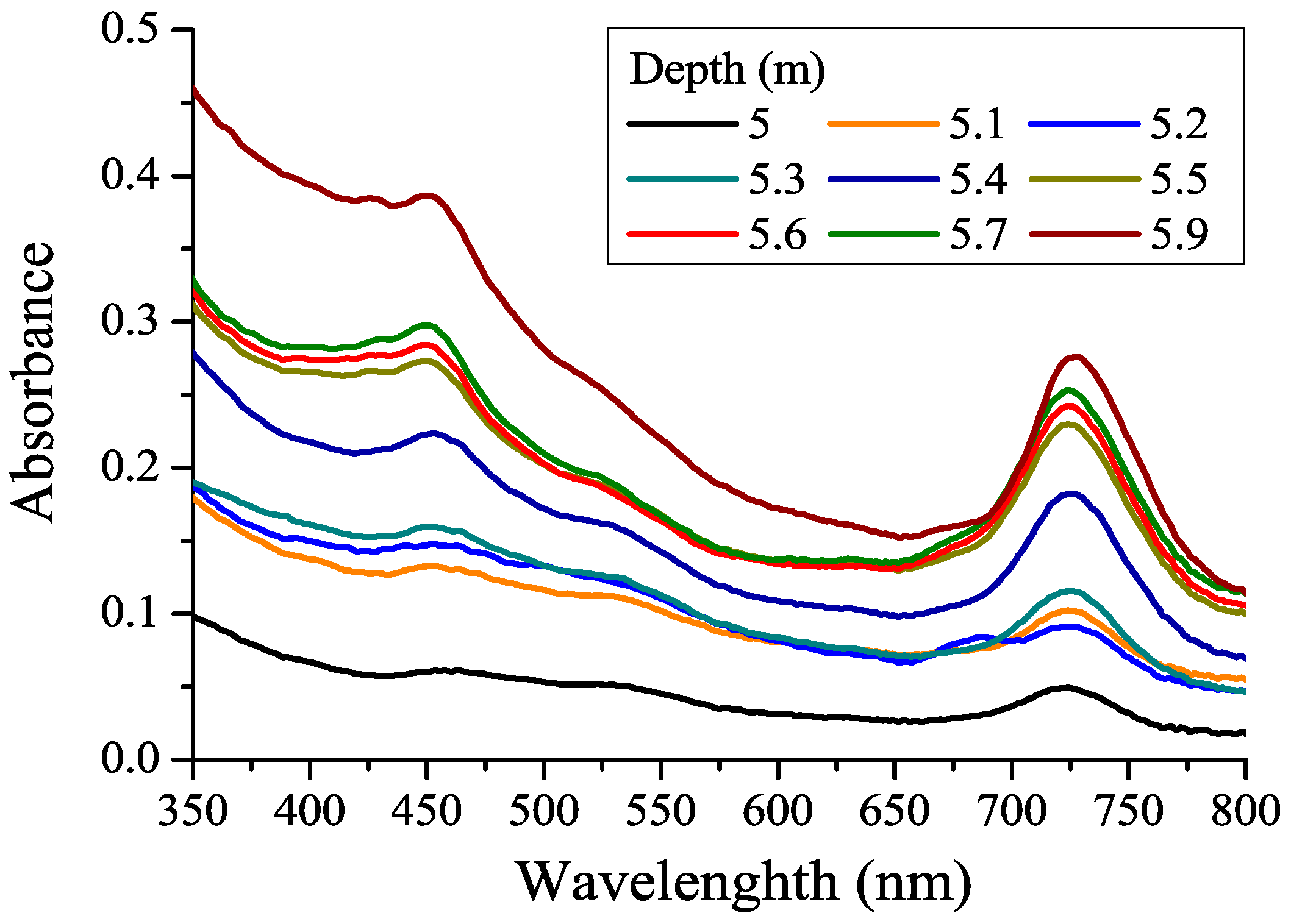
3.1. CDOM Absorption Spectra
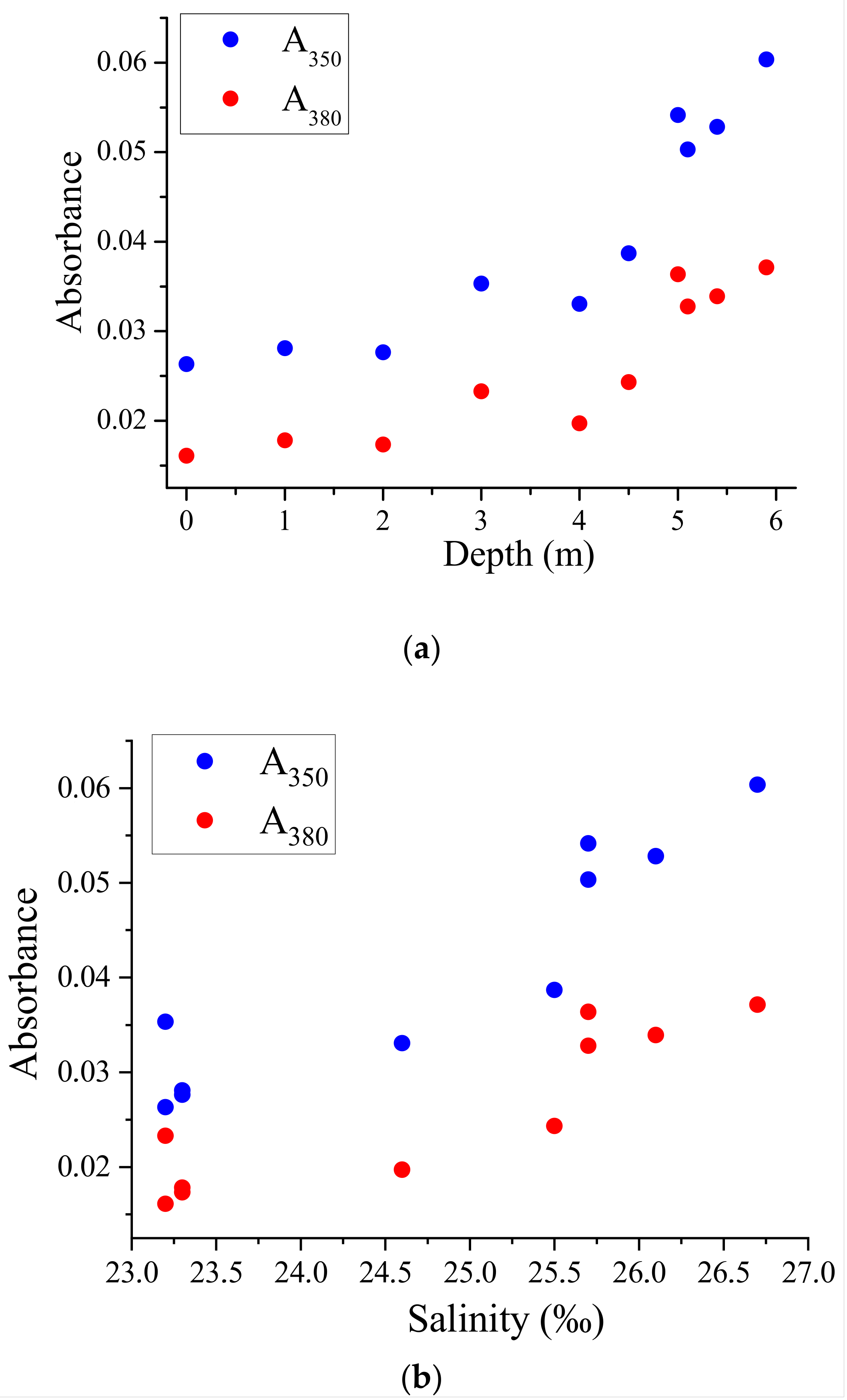
3.2. Correlation between CDOM Absorbances and Water Salinity
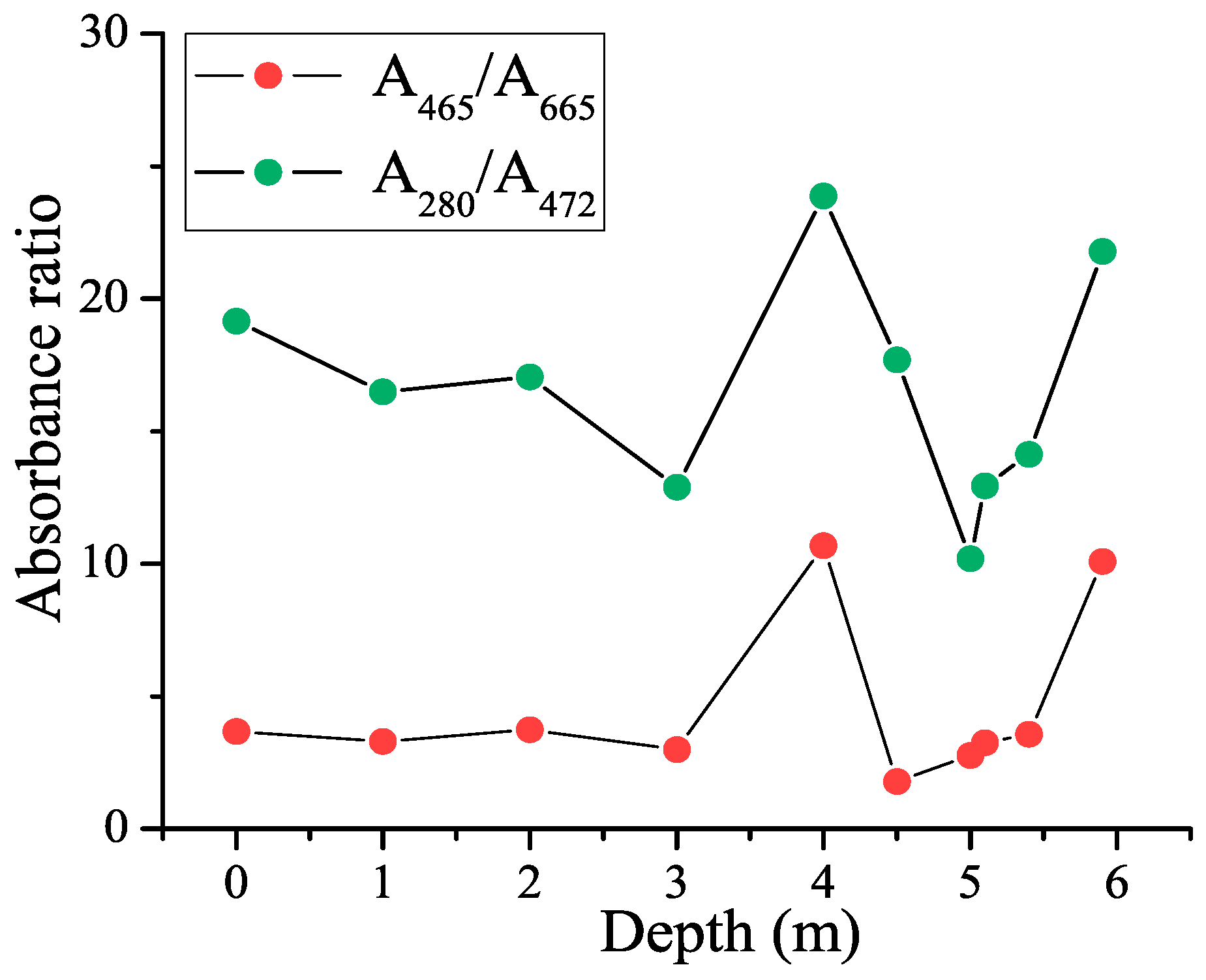
3.3. CDOM Absorbance Ratios
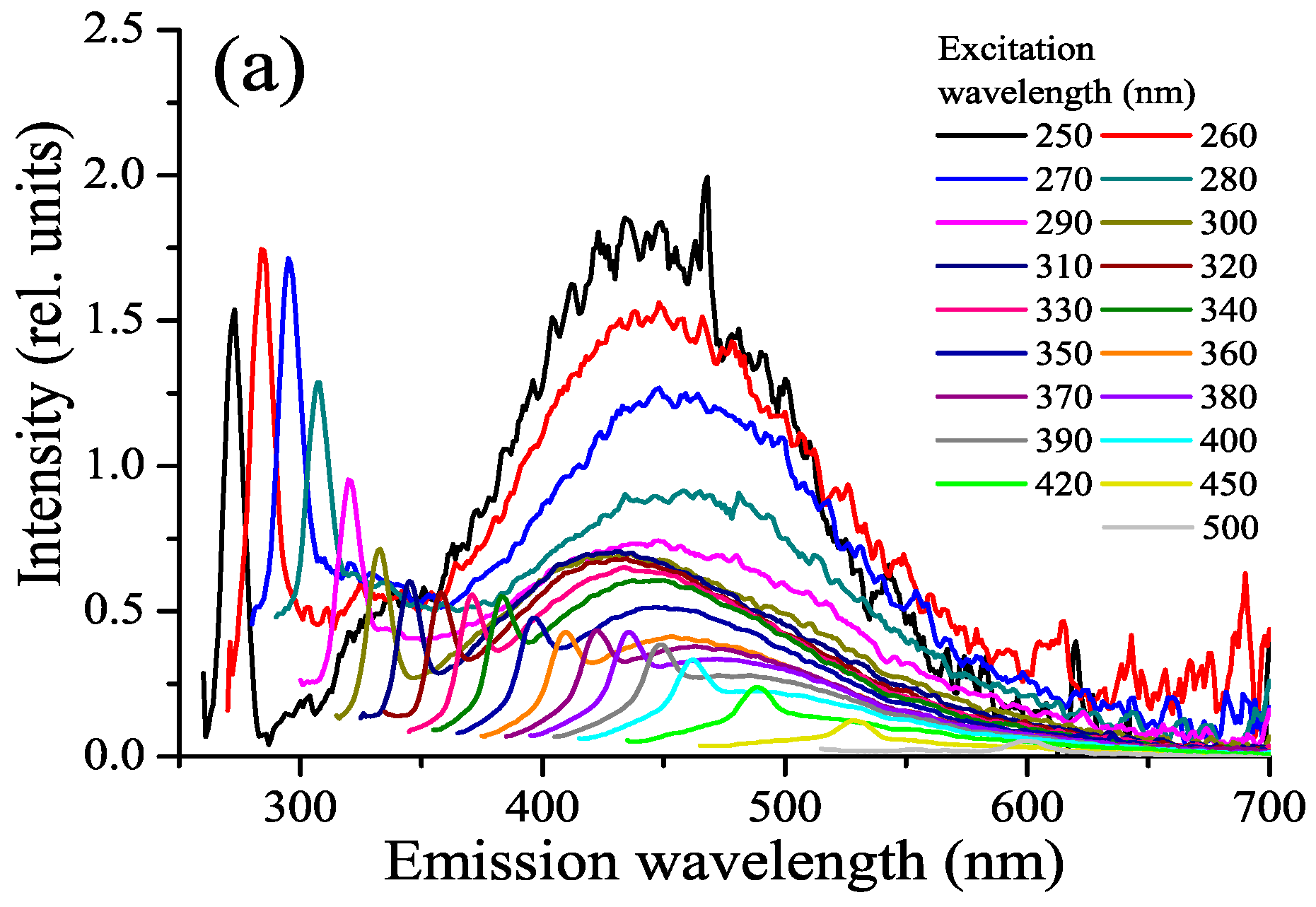
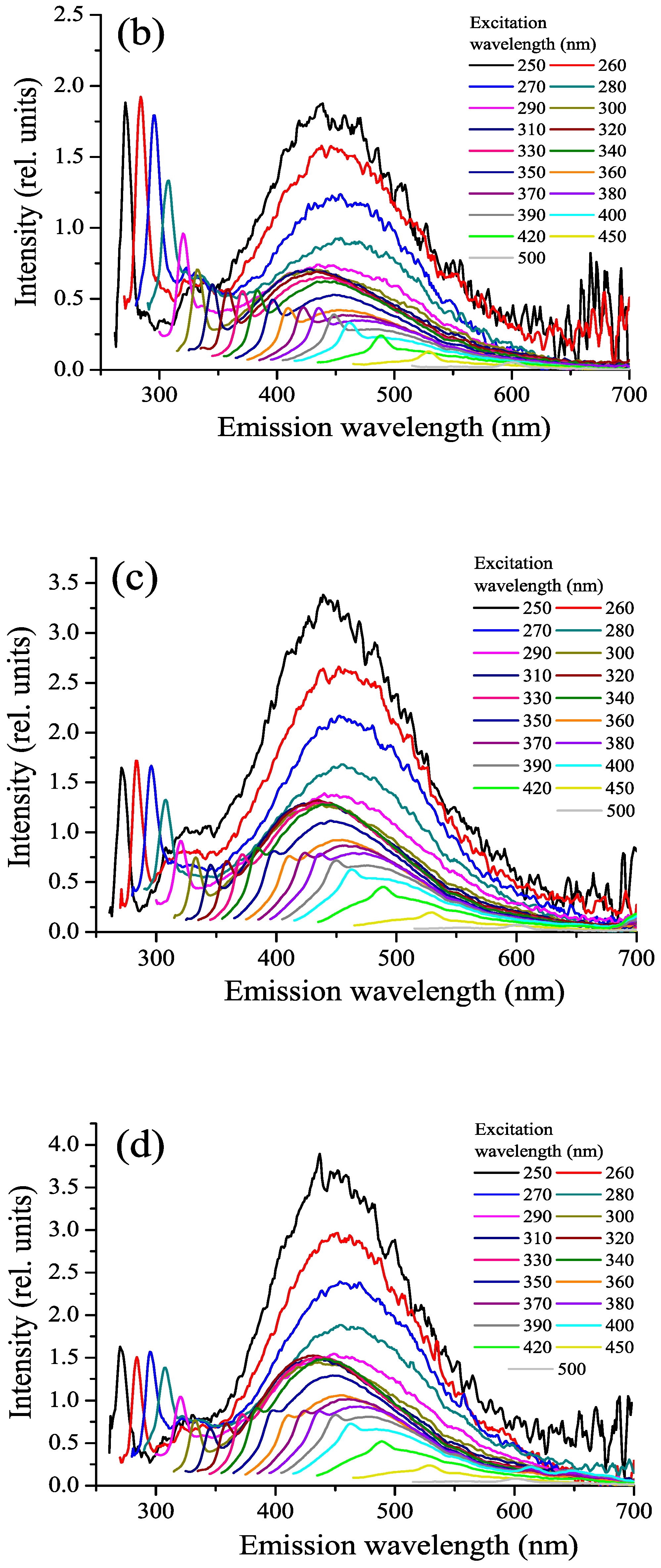
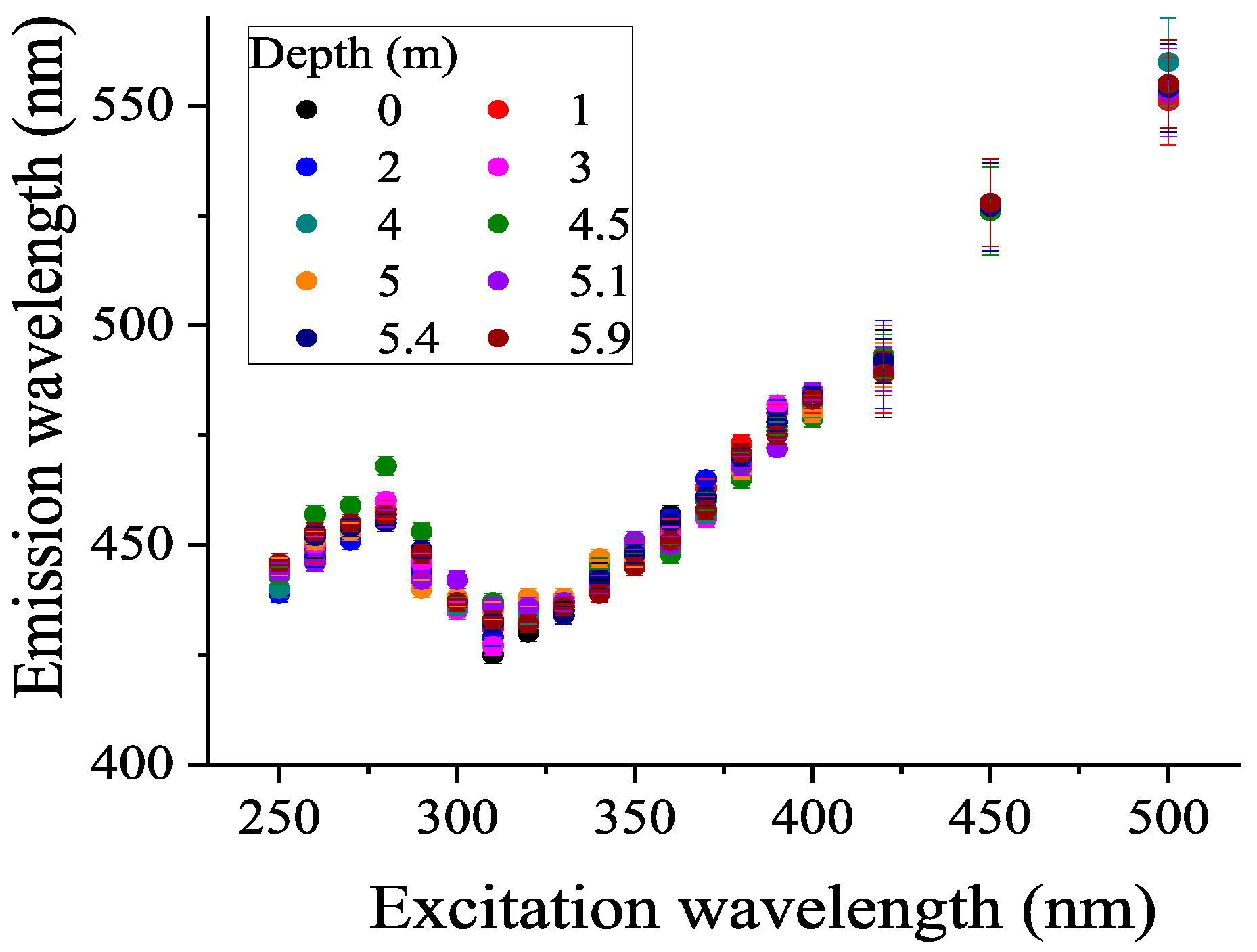
3.4. FDOM, Fluorophoric Fraction of CDOM
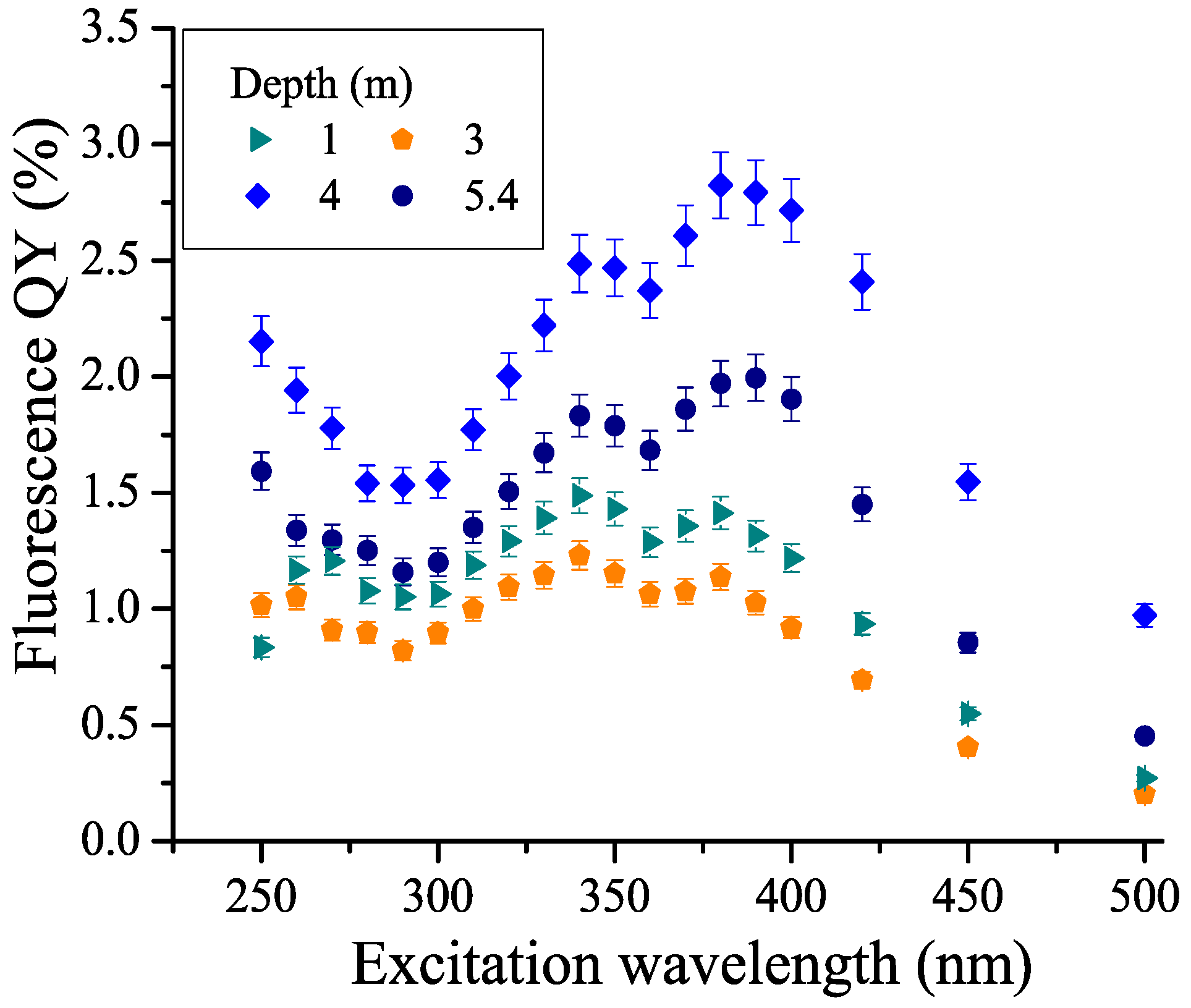
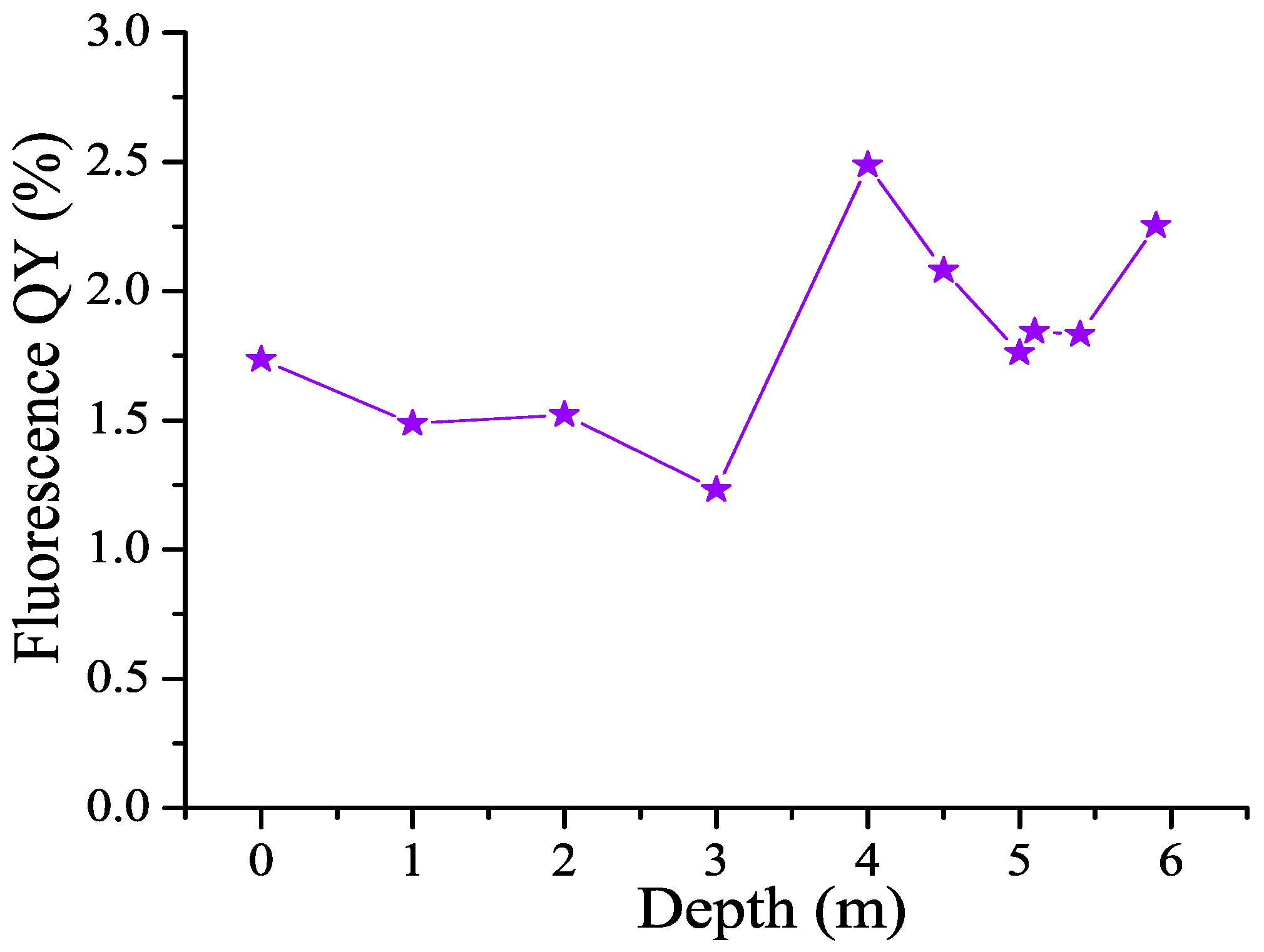
3.5. CDOM Fluorescence Quantum Yield (FDOM/CDOM) and Its Dependence on Depth
3.6. Depth Distribution of Chlorophyll and Photosynthetic Activity
3.7. Depth Distribution of Bacteriochlorophylls of Green Sulfur Bacteria
3.8. Discussion of Results: Optical Properties of CDOM, Chl, and Bchl as Proxies of Euxinia in Meromictic Water Bodies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meyer, K.M.; Kump, L.R. Oceanic Euxinia in Earth History: Causes and Consequences. Annu. Rev. Earth Planet. Sci. 2008, 36, 251–288. [Google Scholar] [CrossRef] [Green Version]
- Lyons, T.W.; Anbar, A.D.; Severmann, S.; Scott, C.; Gill, B.C. Tracking Euxinia in the Ancient Ocean: A Multiproxy Perspective and Proterozoic Case Study. Annu. Rev. Earth Planet. Sci. 2009, 37, 507–534. [Google Scholar] [CrossRef]
- Krasnova, E.D.; Pantyulin, A.N.; Belevich, T.A.; Voronov, D.A.; Demidenko, N.A.; Zhitina, L.S.; Ilyash, L.V.; Kokryatskaya, N.M.; Lunina, O.N.; Mardashova, M.V.; et al. Multidisciplinary studies of the separating lakes at different stage of isolation from the White Sea performed in March 2012. Oceanology 2013, 53, 714–717. [Google Scholar] [CrossRef]
- Krasnova, E.D.; Kharcheva, A.V.; Milyutina, I.A.; Voronov, D.A.; Patsaeva, S.V. Study of microbial communities in redox zone of meromictic lakes isolated from the White Sea using spectral and molecular methods. J. Mar. Biol. Assoc. 2015, 95, 1579–1590. [Google Scholar] [CrossRef]
- Mardashova, M.V.; Voronov, D.A.; Krasnova, E.D. Benthic communities of coastal water bodies at different stages of isolation from the White Sea in the vicinity of the White Sea Biological Station. Biol. Bull. 2020, 47, 1133–1152. [Google Scholar] [CrossRef]
- Krasnova, E.D. Ecology of Meromictic Lakes of Russia. 1. Coastal Marine Waterbodies. Water Resour. 2021, 48, 427–438. [Google Scholar] [CrossRef]
- Repkina, T.; Shilova, O.S.; Krasnova, E.; Entin, A.; Grigoriev, V.; Vakhrameyeva, E.; Losyuk, G.; Kublitskiy, Y.; Leontie, P.; Lugovoy, N.; et al. From the sea strait to the meromictic lake: Evolution and ecosystem of a water body at the fiard coast (lake Kislo-Sladkoe at the Karelian coast of the Kandalaksha bay, the White Sea, Russia). Quat. Int. 2023, 644–645, 96–119. [Google Scholar] [CrossRef]
- Hakala, A. Meromixis as a part of lake evolution—Observations and a revised classification of true meromictic lakes in Finland. Boreal Environ. Res. 2004, 9, 37–53. [Google Scholar]
- Stewart, K.M.; Walker, K.F.; Likens, G.E. Meromictic Lakes. In Encyclopedia of Inland Waters; Academic Press: Oxford, UK, 2009; pp. 589–602. [Google Scholar]
- Krasnova, E.D.; Pantyulin, A.N.; Matorin, D.N.; Todorenko, D.A.; Belevich, T.A.; Milyutina, I.A.; Voronov, D.A. Blooming of the cryptomonad alga Rhodomonas sp. (Cryptophyta, Pyrenomonadaceae) in the redox zone of the basins separating from the White Sea. Microbiology 2014, 83, 270–277. [Google Scholar] [CrossRef]
- Krasnova, E.; Matorin, D.; Belevich, T.; Efimova, L.; Kharcheva, A.; Kokryatskaya, N.; Losyuk, G.; Todorenko, D.; Voronov, D.; Patsaeva, S. The characteristic pattern of multiple colored layers in coastal stratified lakes in the process of separation from the White Sea. Chin. J. Oceanol. Limnol. 2018, 36, 1962–1977. [Google Scholar] [CrossRef]
- Hansell, D.A. Recalcitrant dissolved organic carbon fractions. Annu. Rev. Mar. Sci. 2013, 5, 421–445. [Google Scholar] [CrossRef] [PubMed]
- Hansell, D.A.; Carlson, C.A. Biogeochemistry of Marine Dissolved Organic Matter, 2nd ed.; Academic Press: San Diego, CA, USA, 2015. [Google Scholar]
- Jansson, M.; Bergström, A.K.; Blomqvist, P.; Isaksson, A.; Jonsson, A. Impact of allochthonous organic carbon on microbial food web carbon dynamics and structure in Lake Örträsket. Arch. Hydrobiol. 1999, 144, 409–428. [Google Scholar] [CrossRef]
- Jansson, M.; Hickler, T.; Jonsson, A.; Karlsson, J. Links between terrestrial primary production and bacterial production and respiration in lakes in a climate gradient in subarctic Sweden. Ecosystems 2008, 11, 367–376. [Google Scholar] [CrossRef]
- Khundzhua, D.A.; Patsaeva, S.V.; Terekhova, V.A.; Yuzhakov, V.I. Spectral characterization of fungal metabolites in aqueous medium with humus substances. J. Spectrosc. 2013, 2013, 538608. [Google Scholar] [CrossRef] [Green Version]
- Fedoseeva, E.; Stepanov, A.; Yakimenko, O.; Patsaeva, S.; Freidkin, M.; Khundzhua, D.; Terekhova, V. Biodegradation of humic substances by microscopic filamentous fungi: Chromatographic and spectroscopic proxies. J. Soils Sediments 2019, 19, 2676–2687. [Google Scholar] [CrossRef]
- Fedoseeva, E.V.; Patsaeva, S.V.; Khundzhua, D.A.; Pukalchik, M.A.; Terekhova, V.A. Effect of exogenic humic substances on various growth endpoints of Alternaria Alternata and Trichoderma Harzianum in the experimental conditions. Waste Biomass Valorization 2021, 12, 211–222. [Google Scholar] [CrossRef]
- Tranvik, L.J. Dystrophy in Freshwater Systems. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Catrouillet, C.; Davranche, M.; Dia, A.; Coz, M.; Marsac, R.; Pourret, O.; Gruau, G. Geochemical modeling of Fe(II) binding to humic and fulvic acids. Chem. Geol. 2014, 372, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Terekhova, V.A.; Fedoseeva, E.V.; Panova, M.I.; Chukov, S.N. Bioassay of humic products as potential remedies: A review. Eurasian Soil Sci. 2022, 55, 868–878. [Google Scholar] [CrossRef]
- Carlsson, P.; Segatto, A.; Granéli, E. Nitrogen bound to humic matter of terrestrial origin-a nitrogen pool for coastal phytoplankton? Mar Ecol Prog Ser. 1993, 97, 105–116. [Google Scholar] [CrossRef]
- Bricaud, A.; Morel, A.; Prieur, L. Absorption by dissolved organic matter of the sea (yellow substance) in the UV and visible domain. Limnol. Oceanogr. 1981, 26, 43–53. [Google Scholar] [CrossRef]
- McKnight, D.M.; Boyer, E.W.; Westerhoff, P.K.; Doran, P.T.; Kulbe, T.; Andersen, D.T. Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnol. Oceanogr. 2001, 46, 38–48. [Google Scholar] [CrossRef]
- Jaffe, R.; McKnight, D.; Maie, N.; Cory, R.; McDowell, W.H.; Campbell, J.L. Spatial and temporal variations in DOM composition in ecosystems: The importance of long-term monitoring of optical properties. J. Geophys. Res. 2008, 113, 04032. [Google Scholar] [CrossRef]
- Helms, J.R.; Stubbins, A.; Ritchie, J.C.; Minor, E.C.; Kieber, D.J.; Mopper, K. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol. Oceanogr. 2008, 53, 955–969. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, L.; Stedmon, C.A.; Tragh, T.; Markager, S.; Middleboe, M.; Sondergaard, M. Global trends in the fluorescence characteristics and distribution of marine dissolved organic matter. Mar. Chem. 2011, 126, 139–148. [Google Scholar] [CrossRef]
- Breves, W.; Reuter, R. Bio-optical properties of gelbstoff in the Arabian Sea at the onset of the southwest monsoon. Earth Planet. Sci. Proc. Indian Acad. Sci. 2012, 109, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Lonborg, C.; Yokokawa, T.; Herndl, G.J.; Alvarez-Salgado, X.A. Production and degradation of fluorescent dissolved organic matter in surface waters of the eastern north Atlantic ocean. Deep Sea Res. I 2015, 96, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Jian, Z.; Xu, J.; Huang, X.; Yang, W.; Hu, Q. Optical Absorption Characteristics, Spatial Distribution, and Source Analysis of Colored Dissolved Organic Matter in Wetland Water around Poyang Lake. Water 2021, 13, 274. [Google Scholar] [CrossRef]
- Reuter, R.; Willkomm, R.; Krieger, J.; Milchers, W.; Patsayeva, S.; Hengstermann, T. Measurement of gelbstoff in seawater from aircraft and on board research vessels using fluorescence techniques. In Lidar Techniques for Remote Sensing II; Werner, C., Ed.; 1995; Volume 2581, pp. 48–59. Available online: https://spie.org/Publications/Proceedings/Paper/10.1117/12.228520?SSO=1 (accessed on 28 May 2023).
- Harsdorf, S.; Reuter, R. Laser remote sensing in highly turbid waters: Validity of the lidar equation. In Environmental Sensing and Applications; SPIE: Washington, DC, USA, 1999; Volume 3821. [Google Scholar] [CrossRef]
- Barbini, R.; Colao, F.; Fantoni, R.; Fiorani, L.; Kolodnikova, N.; Palucci, A. Laser remote sensing calibration of ocean color satellite data. Ann. Geophys. 2009, 49. [Google Scholar] [CrossRef]
- Fiorani, L.; Angelini, F.; Colao, F.; Nuvoli, M.; Palucci, A. Laser radars for marine monitoring. In Proceedings of the XXII International Symposium on High Power Laser Systems and Applications, Frascati, Italy, 9–12 October 2019; p. 1104212. [Google Scholar] [CrossRef]
- Aruffo, E.; Chiuri, A.; Angelini, F.; Artuso, F.; Cataldi, D.; Colao, F.; Fiorani, L.; Menicucci, I.; Nuvoli, M.; Pistilli, M.; et al. Hyperspectral Fluorescence LIDAR Based on a Liquid Crystal Tunable Filter for Marine Environment Monitoring. Sensors 2020, 20, 410. [Google Scholar] [CrossRef] [Green Version]
- Kahru, M.; Mitchell, B.G. Seasonal and nonseasonal variability of satellite-derived chlorophyll and colored dissolved organic matter concentration in the California Current. J. Geophys. Res. 2001, 106, 2517–2529. [Google Scholar] [CrossRef]
- Fantoni, R.; Fiorani, L.; Palucci, A.; Okladnikov, I.G. New algorithm for CDOM retrieval from satellite imagery. In Technical Reports of the Italian Agency for New Technologies, Energy and the Environment; Renieri, A., Ed.; ENEA: Frascati, Italy, 2005. [Google Scholar]
- Aurin, D.; Mannino, A.; Lary, D.J. Remote Sensing of CDOM, CDOM Spectral Slope, and Dissolved Organic Carbon in the Global Ocean. Appl. Sci. 2018, 8, 2687. [Google Scholar] [CrossRef] [Green Version]
- Groom, S.; Sathyendranath, S.; Ban, Y.; Bernard, S.; Brewin, R.; Brotas, V.; Brockmann, C.; Chauhan, P.; Choi, J.-K.; Chuprin, A.; et al. Satellite Ocean Colour: Current Status and Future Perspective. Front. Mar. Sci. 2019, 6, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patsaeva, S.; Khundzhua, D.; Trubetskoj, O.A.; Trubetskaya, O.E. Excitation-dependent fluorescence quantum yield for freshwater chromophoric dissolved organic matter from northern Russian lakes. J. Spectrosc. 2018, 2018, 3168320. [Google Scholar] [CrossRef] [Green Version]
- Matorin, D.; Antal, T.; Ostrowska, M.; Rubin, A.; Ficek, D.; Majchrowski, R. Chlorophyll fluorimetry as a method for studying light absorption by photosynthetic pigments in marine algae. Oceanologia 2004, 46, 519–531. [Google Scholar]
- Emeliantsev, P.S.; Zhiltsova, A.A.; Krasnova, E.D.; Voronov, D.A.; Rymar, V.V.; Patsaeva, S.V. Quantification of chlorosomal bacteriochlorophylls using absorption spectra of green sulfur bacteria in natural water. Mosc. Univ. Phys. Bull. 2020, 75, 137–142. [Google Scholar] [CrossRef]
- Zhiltsova, A.A.; Filippova, O.A.; Krasnova, E.D.; Voronov, D.A.; Patsaeva, S.V. Comparative analysis of spectral methods for determining bacteriochlorophyll d concentration in green sulfur bacteria in water. Atmos. Ocean. Opt. 2022, 35, 562–568. [Google Scholar] [CrossRef]
- Yakimenko, O.; Khundzhua, D.; Izosimov, A.; Yuzhakov, V.; Patsaeva, S. Source indicator of commercial humic products: UV-vis and fluorescence proxies. J. Soils Sediments 2018, 18, 1279–1291. [Google Scholar] [CrossRef]
- Trubetskaya, O.E.; Richard, C.; Patsaeva, S.V.; Trubetskoj, O.A. Evaluation of aliphatic/aromatic compounds and fluorophores in dissolved organic matter of contrasting natural waters by SEC-HPLC with multi-wavelength absorbance and fluorescence detections. Spectrochim. Acta—Part A 2020, 238, 118450. [Google Scholar] [CrossRef]
- Drozdova, A.N.; Kravchishina, M.D.; Khundzhua, D.A.; Freidkin, M.P.; Patsaeva, S.V. Fluorescence quantum yield of CDOM in coastal zones of the Arctic seas. Intern. J. Remote Sens. 2018, 39, 9356–9379. [Google Scholar] [CrossRef]
- Romankevich, E.A. Geochemistry of Organic Matter in Ocean; Springer: Berlin/Heidelberg, Germany, 1986; p. 336. [Google Scholar] [CrossRef]
- Zaitseva, A.F.; Konyukhov, I.V.; Kazimirko, Y.V.; Pogosyan, S.I. Optical Characteristics and Distribution of Chromophoric Dissolved Organic Matter in Onega Bay (White Sea) during the Summer Season (Findings from an Expedition from June 22 to 26, 2015). Oceanology 2018, 58, 233–239. [Google Scholar] [CrossRef]
- Drozdova, A.N.; Nedospasov, A.A.; Lobus, N.V.; Patsaeva, S.V.; Shchuka, S.A. CDOM optical properties and DOC content in the largest mixing zones of the Siberian shelf seas. Intern. J. Remote Sens. 2021, 13, 1145. [Google Scholar] [CrossRef]
- Glukhovets, D.; Goldin, Y. Research of the relationship between salinity and yellow substance fluorescence in the Kara Sea. Fundam. I Prikl. Gidrofiz. 2018, 11, 34–39. [Google Scholar] [CrossRef]
- Pavlov, A.K.; Stedmon, C.A.; Semushin, A.V.; Martma, T.; Ivanov, B.V.; Kowalczuk, P.; Granskog, M.A. Linkages between the circulation and distribution of dissolved organic matter in the White Sea, Arctic Ocean. Cont. Shelf Res. 2016, 119, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Patsayeva, S.; Reuter, R. Spectroscopic study of major components of dissolved organic matter naturally occurring in water. In Global Process Monitoring and Remote Sensing of the Ocean and Sea Ice; Deering, D.W., Gudmandsen, P., Eds.; SPIE: Washington, DC, USA, 1995; Volume 2586, pp. 151–160. [Google Scholar] [CrossRef]
- Domeizel, M.; Khalil, A.; Prudent, P. UV spectroscopy: A tool for monitoring humification and for proposing an index of the maturity of compost. Bioresour. Technol. 2004, 94, 177–184. [Google Scholar] [CrossRef]
- Albrecht, R.; Petit, J.L.; Terrom, G.; Périssol, C. Comparison between UV spectroscopy and NIRS to assess humification process during sewage sludge and green wastes co-composting. Bioresour. Technol. 2011, 102, 4495–4500. [Google Scholar] [CrossRef]
- Kononova, M.M. Soil Organic Matter, Its Nature, Its Role in Soil Formation and in Soil Fertility; Pergamon: Oxford, UK, 1966. [Google Scholar]
- Stevenson, F.J. Humus Chemistry: Genesis, Composition, Reactions; John Wiley and Sons: New York, NY, USA, 1994. [Google Scholar]
- Coble, P.G. Characterization of marine and terrestrial DOM in seawater using excitation—emission matrix spectroscopy. Mar. Chem. 1996, 51, 325–346. [Google Scholar] [CrossRef]
- Del Vecchio, R.; Blough, N.V. On the origin of the optical properties of humic substances. Environ. Sci. Technol. 2004, 38, 3885–3891. [Google Scholar] [CrossRef]
- Gosteva, O.Y.; Izosimov, A.A.; Patsaeva, S.V.; Yakimenko, O.S.; Yuzhakov, V.I. Fluorescence of aqueous solutions of commercial humic products. J. Appl. Spectrosc. 2012, 78, 884–891. [Google Scholar] [CrossRef]
- Wünsch, U.; Murphy, K.; Stedmon, C. Fluorescence Quantum Yields of Natural Organic Matter and Organic Compounds: Implications for the Fluorescence-based Interpretation of Organic Matter Composition. Front. Mar. Sci. 2015, 2, 98. [Google Scholar] [CrossRef] [Green Version]
- Donard, O.F.X.; Lamotte, M.; Belin, C.; Ewald, M. High-sensitivity fluorescence spectroscopy of Mediterranean waters using a conventional or a pulsed laser excitation source. Mar. Chem. 1989, 27, 117–136. [Google Scholar] [CrossRef]
- Fedoseeva, E.; Patsaeva, S.; Stom, D.; Terekhova, V. Excitation-dependent fluorescence helps to indicate fungal contamination of aquatic environments and to differentiate filamentous fungi. Photonics 2022, 9, 692. [Google Scholar] [CrossRef]
- Matorin, D.N.; Todorenko, D.A.; Voronov, D.A.; Goryachev, S.N.; Bratkovskaya, L.B.; Krasnova, E.D. Characteristics of the distribution and state of phytoplankton at various depths in Kislo-Sladkoe lake (White Sea). Mosc. Univ. Biol. Sci. Bull. 2022, 77, 165–171. [Google Scholar] [CrossRef]
- Mindolina, Y.; Selivanova, E.; Ignatenko, M.; Krasnova, E.; Voronov, D.; Plotnikov, A. Taxonomic composition of protist communities in the coastal stratified lake Kislo-Sladkoe (Kandalaksha Bay, White Sea) revealed by microscopy. Diversity 2023, 15, 44. [Google Scholar] [CrossRef]
- Grouzdev, D.; Gaisin, V.; Lunina, O.; Krutkina, M.; Krasnova, E.; Voronov, D.; Baslerov, R.; Sigalevich, P.; Savvichev, A.; Gorlenko, V. Microbial communities of stratified aquatic ecosystems of Kandalaksha bay (White Sea) shed light on the evolutionary history of green and brown morphotypes of Chlorobiota. FEMS Micriobiol. Ecol. 2022, 98, 103. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.S.; Manni, M.; Hassler, C.; Cable, R.N.; Duhaime, M.B.; Zdobnov, E.M. Genomic insights into the coupling of a Chlorella-like microeukaryote and sulfur bacteria in the chemocline of permanently stratified Lake Cadagno. ISME J. 2023, 17, 903–915. [Google Scholar] [CrossRef]
- Chen, R.F.; Bada, J.L. The fluorescence of dissolved organic matter in seawater. Mar. Chem. 1992, 37, 191–221. [Google Scholar] [CrossRef]
- Determann, S.; Reuter, R.; Wilkomm, R. Fluorescent matter in the eastern Atlantic Ocean.2. Vertical profiles and relation to water masses. Deep Sea Res. I 1996, 43, 345–360. [Google Scholar] [CrossRef]
- Xiao, M.; Chen, Z.; Zhang, Y.; Wen, Y.; Shang, L.; Zhong, J. The Optical Characterization and Distribution of Dissolved Organic Matter in Water Regimes of Qilian Mountains Watershed. Int. J. Environ. Res. Public Health 2022, 19, 59. [Google Scholar] [CrossRef]
- Ma, S.; Zhang, X.; Xiong, Y.; Huang, G.; Han, Y.; Funari, V. Assessment of Eutrophication and DOC Sources Tracing in the Sea Area around Dajin Island Using CASI and MODIS Images Coupled with CDOM Optical Properties. Sensors 2021, 21, 4765. [Google Scholar] [CrossRef]
- Zhang, X.; Du, Y.; Mao, Z.; Bi, L.; Chen, J.; Jin, H.; Ma, S. Dissolved Organic Carbon Source Attribution in the Changjiang Outflow Region of the East China Sea. Sensors 2021, 21, 8450. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth, m | 3.0 | 4.0 | 4.5 | 5.0 | 5.1 | 5.2 | 5.3 | 5.4 | 5.5 | 5.6 | 5.7 | 5.9 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chl F0 | 49 | 163 | 174 | 415 | 2175 | 3202 | -- | -- | ||||
| Chl Fv/Fm | 0.52 | 0.63 | 0.33 | 0.22 | 0.76 | 0.77 | 0 | 0 | ||||
| Eh, mV | 99 | 101 | 67 | 54 | 35 | −9 | −216 | −297 | −317 | −339 | −351 | −365 |
| O2, % | 94 | 82 | 0.9 | 0 | 0 | 0 | ||||||
| Bchl (d+e) mg/m3 | 0 | 0 | 0 | 477 | 690 | 600 * | 1067 | 1905 | 2278 | 2495 | 2598 | 2775 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokolovskaya, Y.G.; Krasnova, E.D.; Voronov, D.A.; Matorin, D.N.; Zhiltsova, A.A.; Patsaeva, S.V. Optical Proxies of Euxinia: Spectroscopic Studies of CDOM, Chlorophyll, and Bacteriochlorophylls in the Lagoon on Zeleny Cape (the White Sea). Photonics 2023, 10, 672. https://doi.org/10.3390/photonics10060672
Sokolovskaya YG, Krasnova ED, Voronov DA, Matorin DN, Zhiltsova AA, Patsaeva SV. Optical Proxies of Euxinia: Spectroscopic Studies of CDOM, Chlorophyll, and Bacteriochlorophylls in the Lagoon on Zeleny Cape (the White Sea). Photonics. 2023; 10(6):672. https://doi.org/10.3390/photonics10060672
Chicago/Turabian StyleSokolovskaya, Yu. G., E. D. Krasnova, D. A. Voronov, D. N. Matorin, A. A. Zhiltsova, and S. V. Patsaeva. 2023. "Optical Proxies of Euxinia: Spectroscopic Studies of CDOM, Chlorophyll, and Bacteriochlorophylls in the Lagoon on Zeleny Cape (the White Sea)" Photonics 10, no. 6: 672. https://doi.org/10.3390/photonics10060672