
Effects of Sponge-Derived Alkaloids on Activities of the Bacterial α-D-Galactosidase and Human Cancer Cell α-N-Acetylgalactosaminidase

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Experimental Equipment
2.3. Collection and Identification of Sponge Material
2.4. Isolation and Purification of Compounds
2.5. Cell Cultured
2.6. Cytotoxic Activity Assays
2.7. Cell Treatment with Alkoloids
2.8. Preparation of Cell Lysate
2.9. Isolation and Purification of α-N-Acetylgalactosaminidase from Cell Lysates
2.10. Production and Purification of Recombinant α-D-galactosidase
2.11. Enzyme Assay
2.12. The Inhibitory Potency of the Compounds 1–9 for the α-PsGal and α-NaGalase
2.13. The Irreversibility of α-PsGal Inhibition by Alkaloids
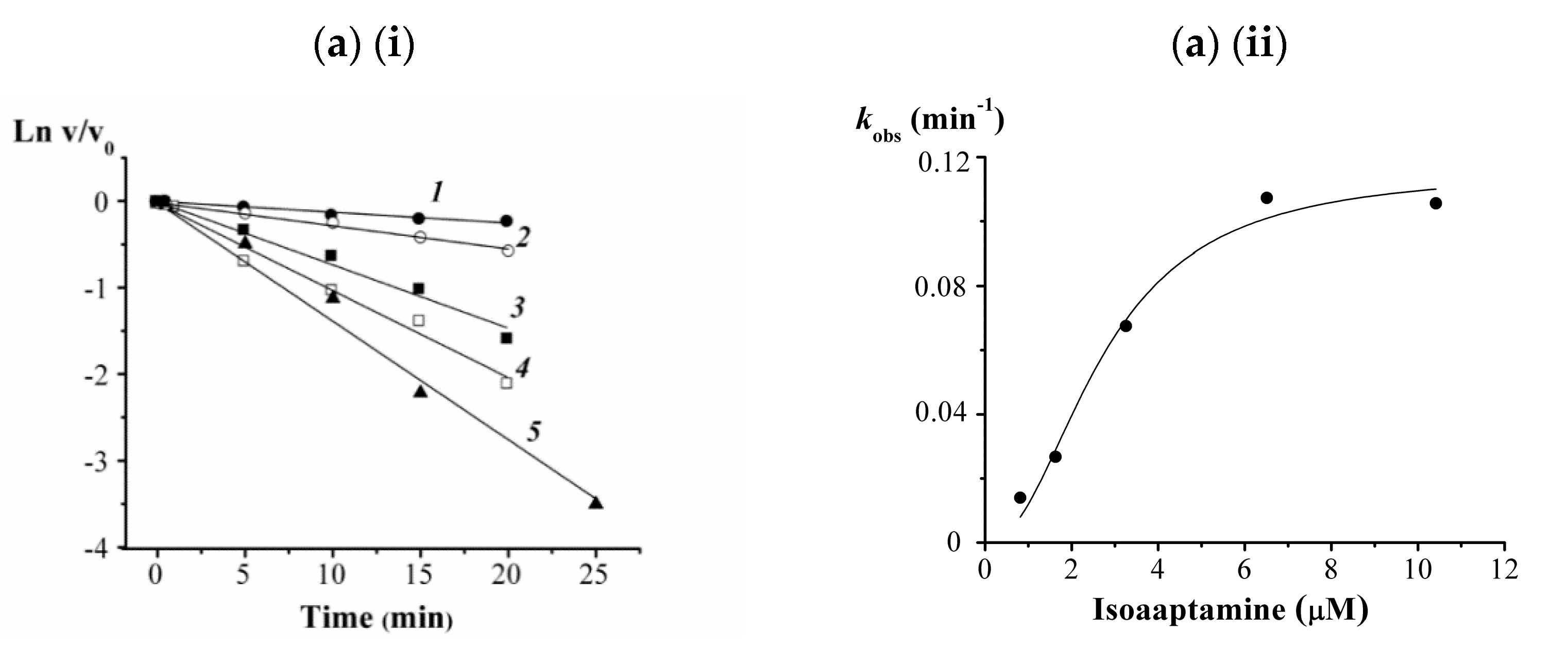
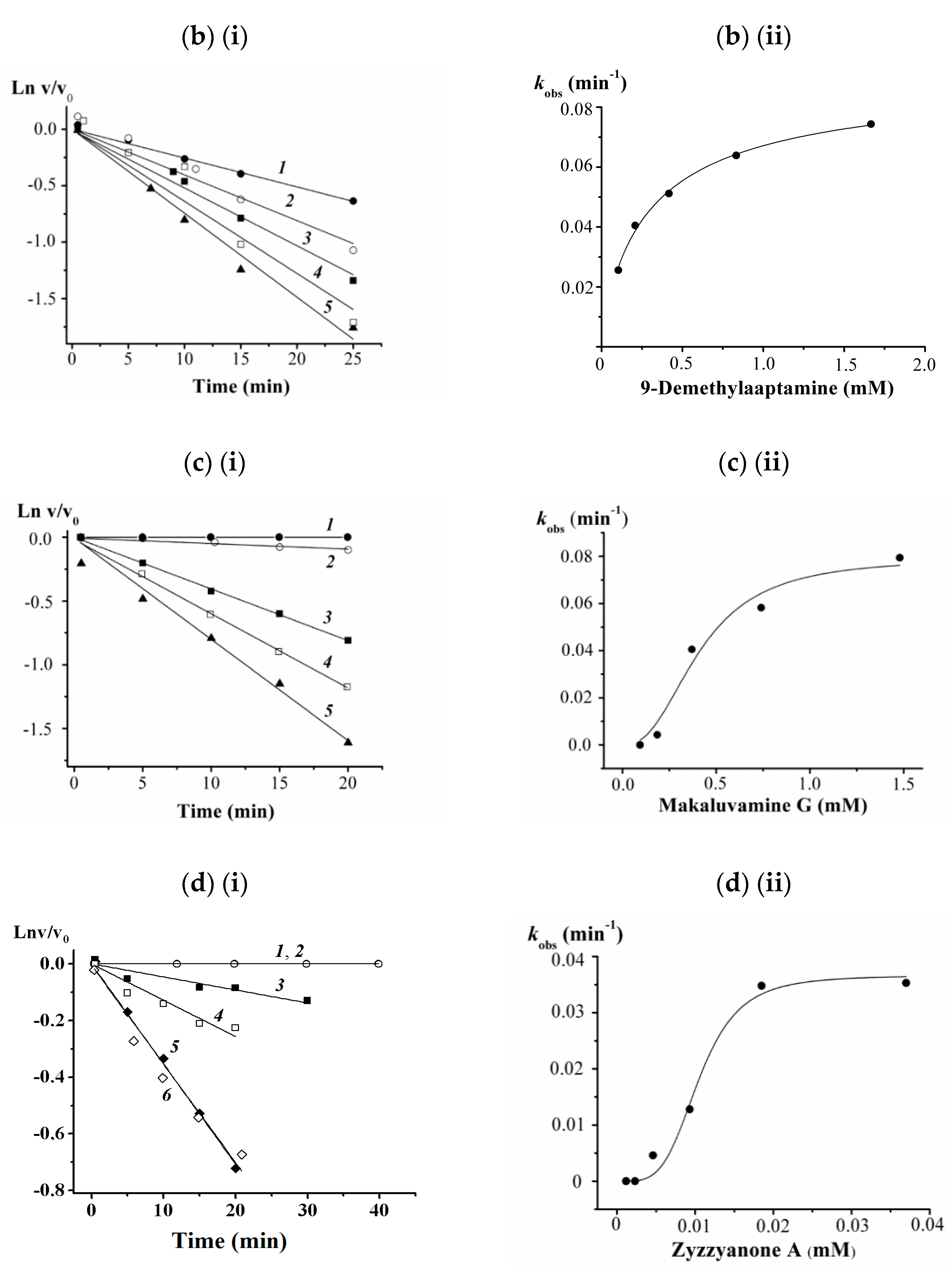
2.14. Kinetic Studies on 2, 3, 8 and 9 against the α-PsGal
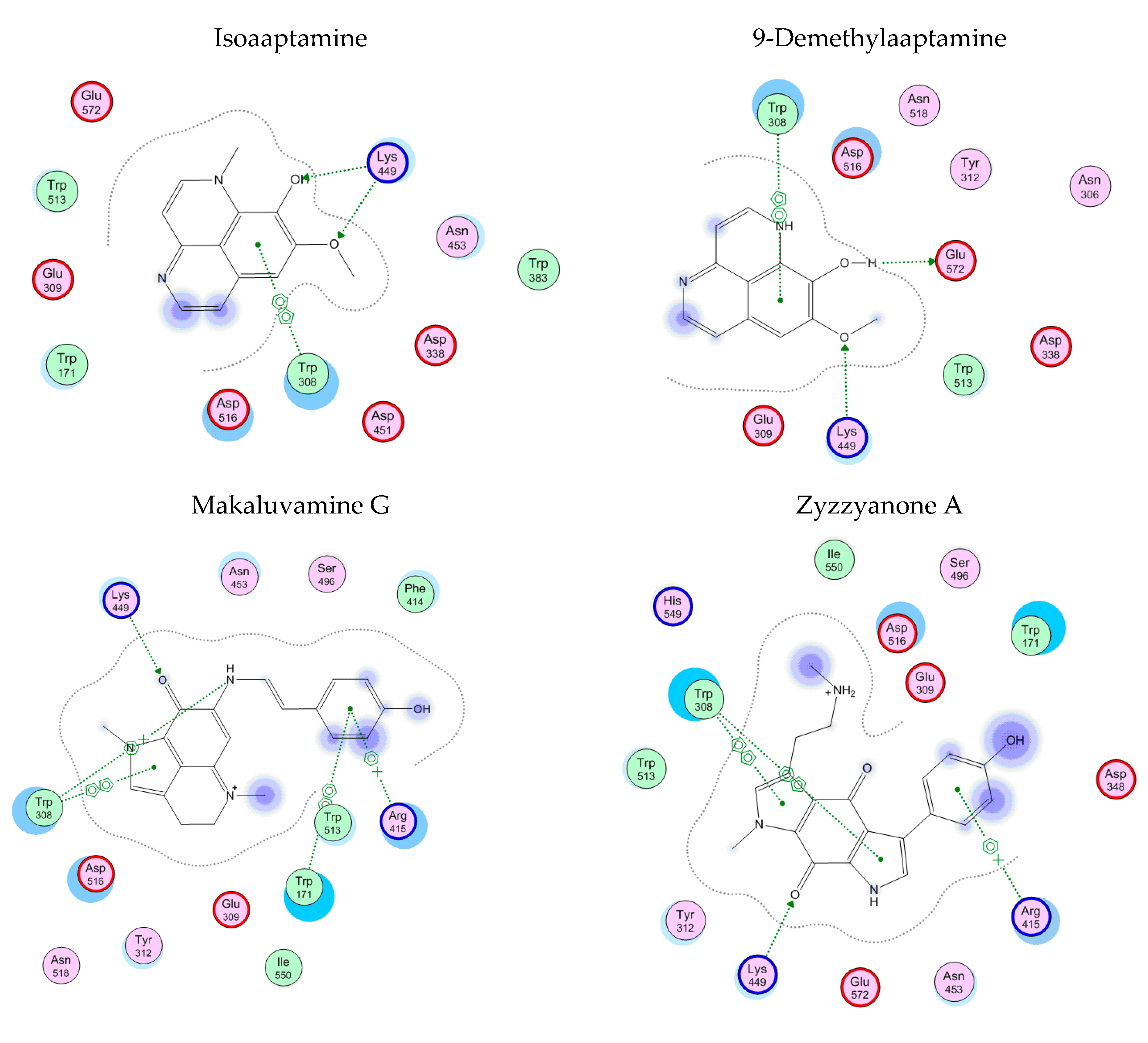
2.15. Theoretical Models of α-D-Galactosidase Complexes with Aaptamine and Makaluvamine Alkaloids
3. Results and Discussion
3.1. Identification of the Compounds
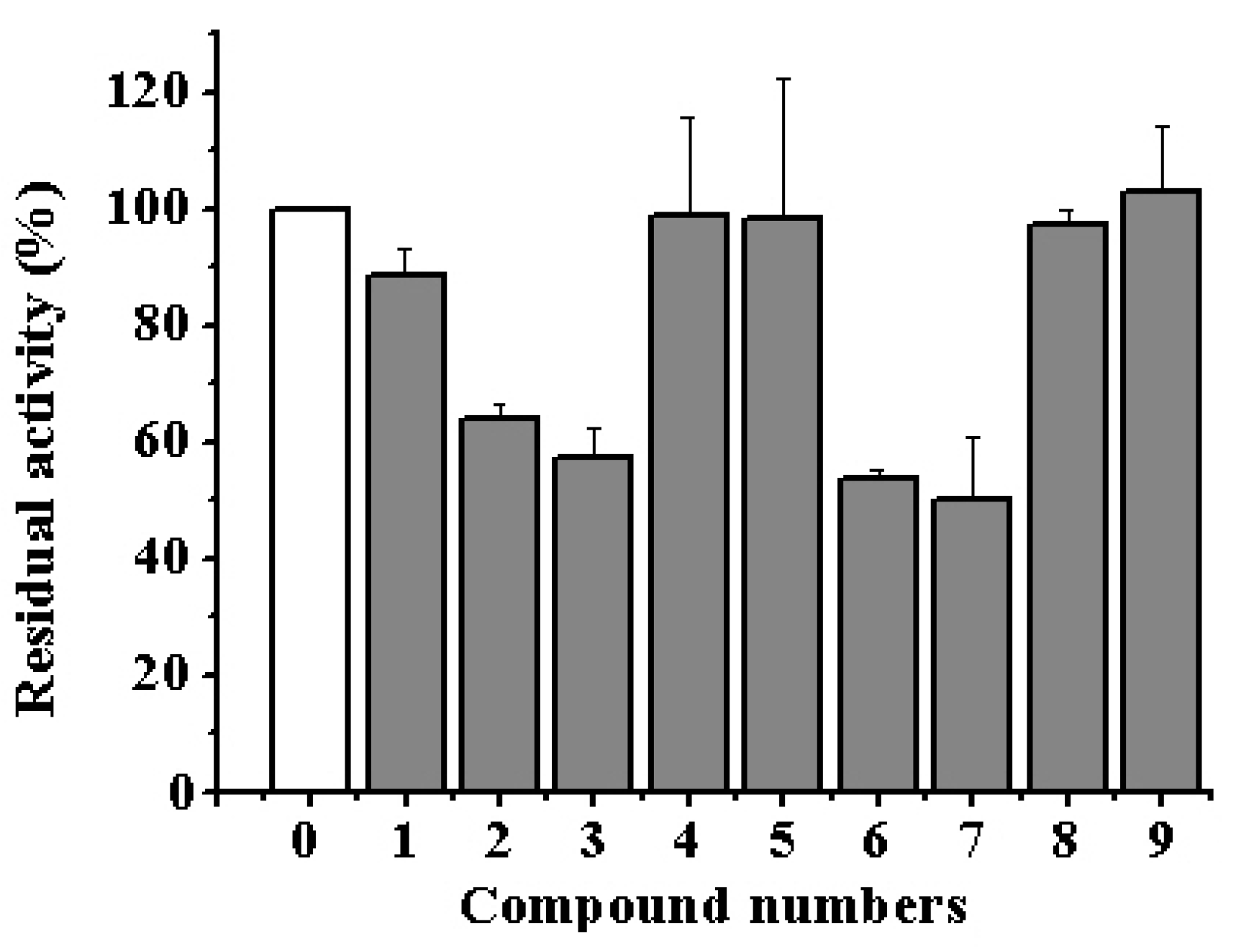
3.2. Cytotoxic Effect of Alkaloids on Human Colorectal Adenocarcinoma Cell Line DLD-1
3.3. Inhibitory Potency of Aaptamine and Makaluvamine Classes of Alkaloids on the α-NaGalase of Cancer Cells
3.4. Inhibitory Potency of Aaptamine and Makaluvamine Classes of Alkaloids on the α-PsGal of the Marine Bacterium
3.5. Kinetic Studies on 2, 3, 8, and 9 against the α-PsGal
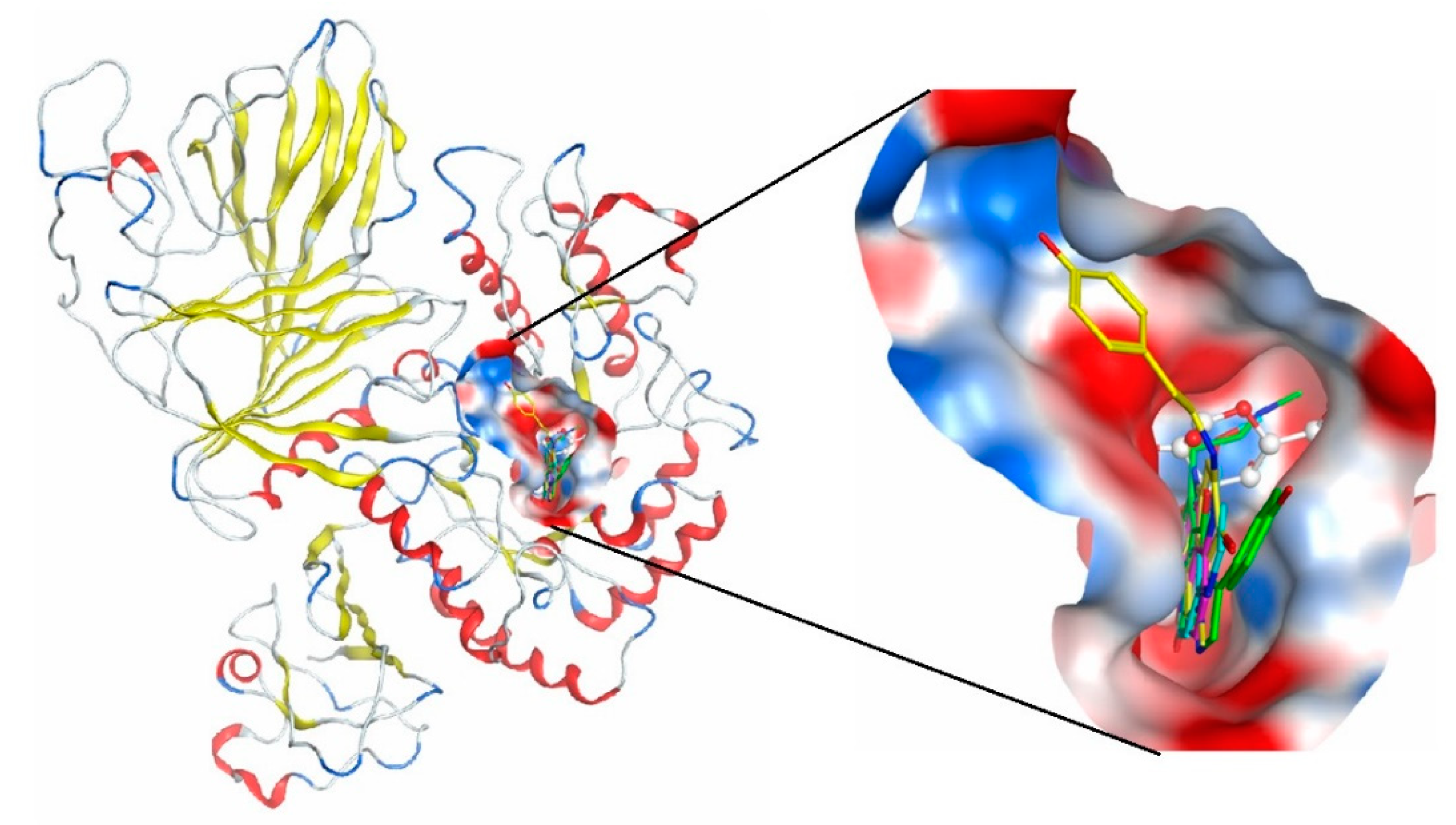
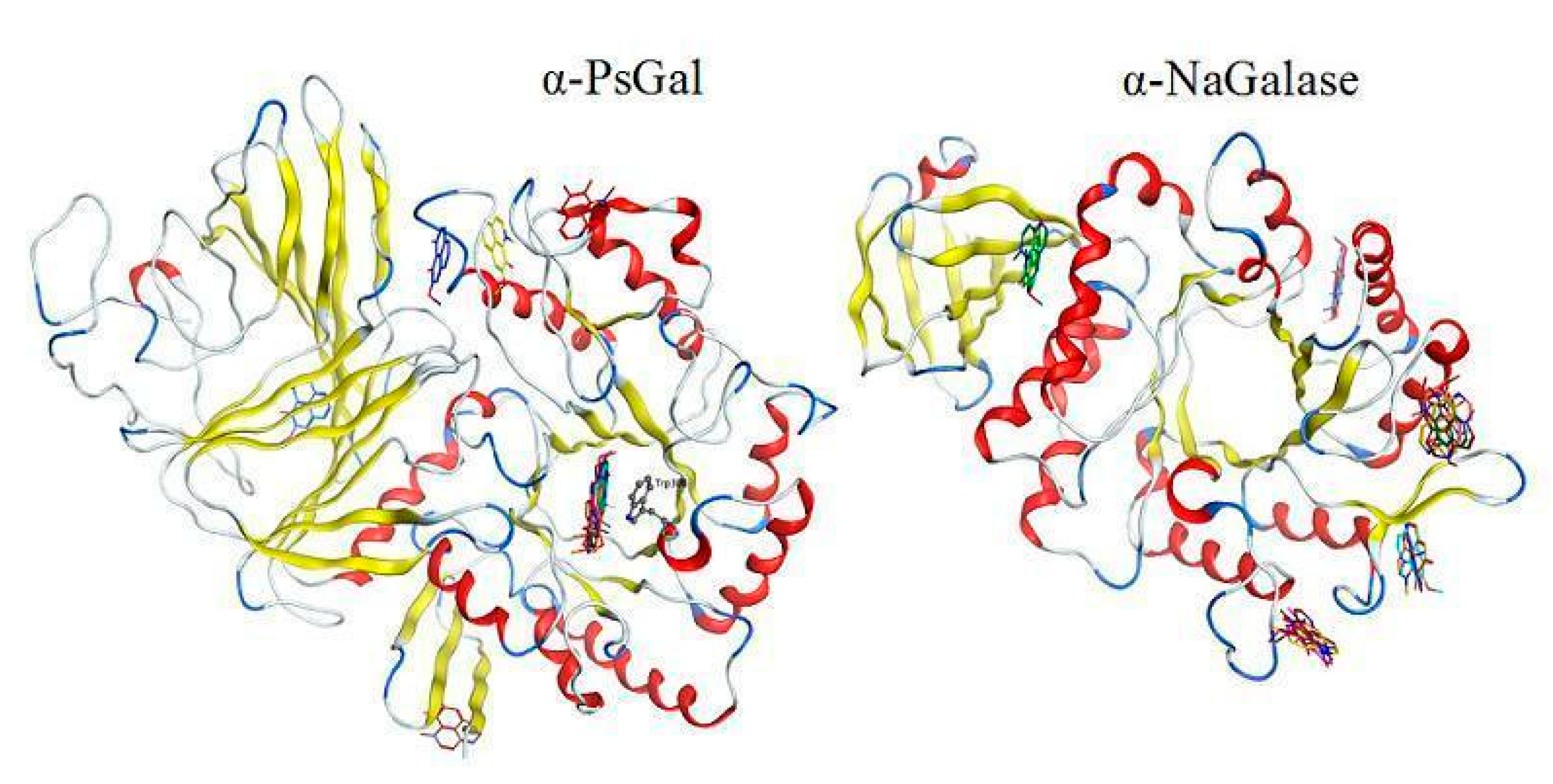
3.6. Theoretical Models of α-D-Galactosidase Complexes with Aaptamine and Makaluvamine Alkaloids
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Antunes, E.M.; Copp, B.R.; Davies-Coleman, M.T.; Samaai, T. Pyrroloiminoquinone and related metabolites from marine sponges. Nat. Prod. Rep. 2005, 22, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Nijampatnam, B.; Nadkarni, D.H.; Wu, H.; Velu, S.E. Antibacterial and antibiofilm activities of makaluvamine analogs. Microorganisms 2014, 2, 128–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, E.; Alvariño, R.; Leirós, M.; Tabudravu, J.N.; Feussner, K.; Dam, M.A.; Rateb, M.E.; Jaspars, M.; Botana, L.M. Evaluation of the Antioxidant activity of the marine pyrroloiminoquinone makaluvamines. Mar. Drugs 2016, 14, 197. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.A.; Buchanan, M.S.; Duffy, S.; Avery, V.M.; Charman, S.A.; Charman, W.N.; White, K.L.; Shackleford, D.M.; Edstein, M.D.; Andrews, K.T.; et al. Antimalarial activity of pyrroloiminoquinones from the Australian marine sponge Zyzzya sp. J. Med. Chem. 2012, 55, 5851–5858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goey, A.K.; Chau, C.H.; Sissung, T.M.; Cook, K.M.; Venzon, D.J.; Castro, A.; Ransom, T.R.; Henrich, C.J.; McKee, T.C.; McMahon, J.B.; et al. Screening and biological effects of marine pyrroloiminoquinone alkaloids: Potential inhibitors of the HIF-1α/p300 interaction. J. Nat. Prod. 2016, 79, 1267–1275. [Google Scholar] [CrossRef]
- Chen, T.; Xu, Y.; Guo, H.; Liu, Y.; Hu, P.; Yang, X.; Li, X.; Ge, S.; Velu, S.E.; Nadkarni, D.H.; et al. Experimental therapy of ovarian cancer with synthetic makaluvamine analog: In vitro and in vivo anticancer activity and molecular mechanisms of action. PLoS ONE 2011, 6, e20729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowan, J.; Shadab, M.; Nadkarni, D.H.; Kailash, K.C.; Velu, S.E.; Yusuf, N. Novel marine natural product derived pyrroloiminoquinone with potent activity against skin cancer cells. Mar. Drugs 2019, 17, 443. [Google Scholar] [CrossRef] [Green Version]
- Radisky, D.C.; Radisky, E.S.; Barrows, L.R.; Copp, B.R.; Kramer, R.A.; Ireland, C.M. Novel cytotoxic topoisomerase II inhibiting pyrroloiminoquinones from Fijian sponges of the genus Zyzzya. J. Am. Chem. Soc. 1993, 115, 1632–1638. [Google Scholar] [CrossRef]
- Lin, S.; McCauley, E.P.; Lorig-Roach, N.; Tenney, K.; Naphen, C.N.; Yang, A.-M.; Johnson, T.A.; Hernadez, T.; Rattan, R.; Valeriote, F.A.; et al. Another Look at pyrroloiminoquinone alkaloids-perspectives on their therapeutic potential from known structures and semisynthetic analogues. Mar. Drugs 2017, 15, 98. [Google Scholar] [CrossRef] [Green Version]
- Larghi, E.L.; Bohn, M.L.; Kaufman, T.S. Aaptamine and related products. Their isolation, chemical syntheses, and biological activity. Tetrahedron 2009, 65, 4257–4282. [Google Scholar] [CrossRef]
- Nakamura, H.; Kobayashi, J.; Ohizumi, Y.; Hirata, Y. Isolation and structure of aaptamine a novel heteroaromatic substance possessing α-blocking activity from the sea sponge Aaptos aaptos. Tetrahedron Lett. 1982, 23, 5555–5558. [Google Scholar] [CrossRef]
- Gul, W.; Hammond, N.L.; Yousaf, M.; Bowling, J.J.; Schinazi, R.F.; Wirtz, S.S.; Andrews, G.A.; Cuevas, C.; Hamann, M.T. Modification at the C9 position of the marine natural product isoaaptamine and the impact on HIV-1, mycobacterial, and tumor cell activity. Bioorg. Med. Chem. 2006, 14, 8495–8505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettit, G.R.; Hoffmann, H.; McNulty, J.; Higgs, K.C.; Murphy, A.; Molloy, D.J.; Herald, D.L.; Williams, M.D.; Pettit, R.K.; Doubek, D.L.; et al. Antineoplastic agents. 380. Isolation and X-ray crystal structure determination of isoaaptamine from the Republic of Singapore Hymeniacidon sp. and conversion to the phosphate prodrug hystatin 1. J. Nat. Prod. 2004, 67, 506–509. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.; Han, C.; Yamano, Y.; Setiawan, A.; Kobayashi, M. Aaptamines, marine spongean alkaloids, as anti-dormant mycobacterial substances. J. Nat. Med. 2014, 68, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Souza, T.M.L.; Abrantes, J.L.; Epifanio, R.A.; Fontes, C.F.L.; Frugulhetti, I.C.P.P. The alkaloid 4-methylaaptamine isolated from the sponge Aaptos aaptos impairs Herpes simplex virus type 1 penetration and immediate-early protein synthesis. Planta Med. 2007, 73, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Gong, K.; Miao, S.; Yang, L.; Wu, Y.; Guo, J.; Chen, W.; Dai, J.; Du, J.; Xi, S. Aaptamine attenuates the proliferation and progression of non-small cell lung carcinoma. Pharm Biol. 2020, 58, 1044–1054. [Google Scholar] [CrossRef]
- Jang, K.H.; Chung, S.C.; Shin, J.; Lee, S.H.; Kim, T.I.; Lee, H.S.; Oh, K.B. Aaptamines as sortase A inhibitors from the tropical sponge Aaptos aaptos. Bioorg. Med. Chem. Lett. 2007, 17, 5366–5369. [Google Scholar] [CrossRef]
- Ioffina, D.I.; Volkovitskaya, O.E.; Gorkin, V.Z.; Rebachuk, N.M.; Utkina, N.K.; Fedoreev, S.A. Aaptamine—New selective type A monoamine-oxidase inhibitor. Pharm. Chem. J. 1990, 24, 456–458. [Google Scholar] [CrossRef]
- Sova, V.V.; Fedoreev, S.A. Metabolites from sponges as inhibitors of β-1,3-glucanase. Chem. Nat. Compd. 1990, 26, 420–422. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Yamanokuchi, R.; Yoshitomi, M.; Sato, K.; Ikeda, T.; Rotinsulu, H.; Mangindaan, R.E.P.; de Voogd, N.J.; van Soest, R.W.M.; Yokosawa, H. Aaptamine, an alkaloid from the sponge Aaptos suberitoides, functions as a proteasome inhibitor. Bioorg. Med. Chem. Lett. 2010, 20, 3341–3343. [Google Scholar] [CrossRef]
- Tischler, D.A. Perspective on enzyme inhibitors from marine organisms. Mar. Drugs 2020, 18, 431. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, N.; Costantini, S.; Palumbo, F.; Costantini, M. Marine sponges and bacteria as challenging sources of enzyme inhibitors for pharmacological applications. Mar. Drugs 2017, 15, 173. [Google Scholar] [CrossRef] [Green Version]
- Asano, N. Glycosidase inhibitors: Update and perspectives on practical use. Glycobiology 2003, 13, 93R–104R. [Google Scholar] [CrossRef]
- Yamamoto, N.; Urade, M. Pathogenic significance of α-N-acetylgalactosaminidase activity found in the hemagglutinin of influenza virus. Microbes Infect. 2005, 7, 674–681. [Google Scholar] [CrossRef]
- Yamamoto, N. Pathogenic significance of α-N-acetylgalactosaminidase activity found in the envelope glycoprotein gp160 of human immunodeficiency virus Type, I. AIDS Res. Hum. Retroviruses 2006, 22, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Van Diggelen, O.P.; Schindler, D.; Kleijer, W.J.; Huijmans, J.M.G.; Galjaard, H.; Linden, H.-U.; Peter-Katalinic, J.; Egge, H.; Cantz, M. Lysosomal alpha-NaGalase deficiency: A new inherited metabolic disease. Lancet 1987, 2, 804. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Balabanova, L.A.; Pennacchio, A.; Trincone, A. Hooked on α-D-galactosidases: From biomedicine to enzymatic synthesis. Crit. Rev. Biotechnol. 2016, 36, 233–245. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Sova, V.V.; Nedashkovskaya, O.I.; Kuhlmann, R.A.; Likhosherstov, L.M.; Martynova, M.I.; Mihailov, V.V.; Elyakova, L.A. α-D-galactosidase of the marine bacterium Pseudoalteromonas sp. KMM 701. Biochemisrty 1998, 63, 1209–1215. [Google Scholar]
- Balabanova, L.A.; Bakunina, I.Y.; Nedashkovskaya, O.I.; Makarenkova, I.D.; Zaporozhets, T.S.; Besednova, N.N.; Zvyagintseva, T.N.; Rasskazov, V.A. Molecular characterization and therapeutic potential of a marine bacterium Pseudoalteromonas sp. KMM 701 α-D-galactosidase. Mar. Biotechnol. 2010, 12, 111–120. [Google Scholar] [CrossRef]
- Balabanova, L.; Shkryl, Y.; Slepchenko, L.; Cheraneva, D.; Podvolotskaya, A.; Bakunina, I.; Nedashkovskaya, O.; Son, O.; Tekutyeva, L. Genomic features of a food-derived Pseudomonas aeruginosa strain PAEM and biofilm-associated gene expression under a marine bacterial α-galactosidase. Int. J. Mol. Sci. 2020, 21, 7666. [Google Scholar] [CrossRef]
- Yamamoto, N.; Naraparaju, V.R.; Asbell, S.O. Deglycosylation of serum vitamin D3-binding protein leads to immunosuppression in cancer patients. Cancer Res. 1996, 56, 2827–2831. [Google Scholar] [CrossRef] [PubMed]
- Saburi, E.; Tavakol-Afshari, J.; Biglari, S.; Mortazavi, Y. Is α-N-acetylgalactosaminidase the key to curing cancer? A mini-review and hypothesis. JBUON 2017, 22, 1372–1377. [Google Scholar] [PubMed]
- Albracht, S.P.J.; van Pelt, J. Multiple exo-glycosidases in human serum as detected with the substrate DNP-α-GalNAc. II. Three α-N-acetylgalactosaminidase-like activities in the pH 5 to 8 region. BBA Clin. 2017, 8, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Ayers, B.J.; Hollinshead, J.; Saville, A.W.; Nakagawa, S.; Adachi, I.; Kato, A.; Izumori, K.; Bartholomew, B.; Fleet, G.W.J.; Nash, R.J. Iteamine, the first alkaloid isolated from Itea virginica L. inflorescence. Phytochemistry 2014, 100, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Clark, N.E.; Metcalf, M.C.; Best, D.; Fleet, G.W.J.; Garman, S.C. Pharmacological chaperones for human α-N-acetylgalactosaminidase. Proc. Nalt. Acad. Sci. USA 2012, 109, 17400–17405. [Google Scholar] [CrossRef] [Green Version]
- Bakunina, I.Y.; Chadova, O.A.; Malyarenko, O.S.; Ermakova, S.P. The Effect of fucoidan from the brown alga Fucus evanescence on the activity of α-N-acetylgalactosaminidase of human colon carcinoma cells. Mar. Drugs 2018, 16, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakunina, I.Y.; Kol’tsova, E.A.; Pokhilo, N.D.; Shestak, O.P.; Yakubovskaya, A.Y.; Zvyagintseva, T.N.; Anufriev, V.F. Effect of 5-hydroxy- and 5,8-dihydroxy-1,4-naphthoquinones on the hydrolytic activity of alpha-galactosidase. Chem. Nat. Comp. 2009, 45, 69–73. [Google Scholar] [CrossRef]
- Utkina, N.K.; Likhatskaya, G.N.; Balabanova, L.A.; Bakunina, I.Y. Sponge-derived polybrominated diphenyl ethers and dibenzo-p-dioxins, irreversible inhibitors of the bacterial α-D-galactosidase. Environ. Sci. Processes Impacts 2019, 21, 1754–1763. [Google Scholar] [CrossRef]
- Bakunina, I.; Likhatskaya, G.; Slepchenko, L.; Balabanova, L.; Tekutyeva, L.; Son, O.; Shubina, L.; Makarieva, T. Effect of pentacyclic guanidine alkaloids from the sponge Monanchora pulchra on activity of α-glycosidases from marine bacteria. Mar. Drugs 2019, 17, 22. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, H.; Kobayashi, J.; Ohizumi, Y.; Hirata, Y. Aaptamines. Novel benzo[de] [I,6] naphthyridines from the Okinawan marine sponge Aaptos aaptos. J. Chem. Soc. Perkin Trans. I 1987, 173–176. [Google Scholar] [CrossRef]
- Utkina, N.K.; Denisenko, V.A.; Pushilin, M.A. Aaptanone, a novel zwitterionic metabolite of the aaptamine class with an oxygenated 1,6-naphthyridine core from the Vietnamese marine sponge Aaptos aaptos. Tetrahedron Lett. 2009, 50, 2580–2582. [Google Scholar] [CrossRef]
- Stierle, D.B.; Faulkner, D.J. Two new pyrroloquinoline alkaloids from the sponge Damiria sp. J. Nat. Prod. 1991, 54, 1131–1133. [Google Scholar] [CrossRef]
- Schmidt, E.W.; Harper, M.K.; Faulkner, D.J. Makaluvamines H-M and damirone C from the pohnpeian sponge Zyzzya fuliginosa. J. Nat. Prod. 1995, 58, 1861–1867. [Google Scholar] [CrossRef] [PubMed]
- Carney, J.R.; Scheuer, P.J.; Kelly-Borges, M. Makaluvamine G, a cytotoxic pigment from an Indonesian sponge Histodermella sp. Tetrahedron 1993, 49, 8483–8486. [Google Scholar] [CrossRef]
- Utkina, N.R.; Makarchenko, A.E.; Denisenko, V.A.; Dmitrenok, P.S. Zyzzyanone A, a novel pyrrolo[3,2-f]indole alkaloid from the Australian marine sponge Zyzzya fuliginosa. Tetrahedron Lett. 2004, 45, 7491–7494. [Google Scholar] [CrossRef]
- Laemmli, V.K. Cleavage of structural proteins during of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Golotin, V.A.; Balabanova, L.A.; Noskova, Y.A.; Slepchenko, L.A.; Bakunina, I.Y.; Vorobieva, N.S.; Terentieva, N.A.; Rasskazov, V.A. Optimization of cold-adapted α-D-galactosidase expression in Escherichia coli. Protein Expr. Purif. 2016, 123, 14–18. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Balabanova, L.A.; Golotin, V.A.; Slepchenko, L.A.; Isakov, V.V.; Rasskazov, V.V. Stereochemical course of hydrolytic reaction catalyzed by alpha-galactosidase from cold adaptable marine bacterium of genus Pseudoalteromonas. Front. Chem./Chem. Biol. 2014, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Varfolomeev, S.D. Khimicheskaya Enzimologiya (Chemical Enzymology); Akademiya: Moscow, Russia, 2005; p. 472. [Google Scholar]
- Dubrovskaya, Y.V.; Makarieva, T.N.; Shubina, L.K.; Bakunina, I.Y. Effect of pentacyclic guanidine alkaloids from the marine sponge Monanchora pulchra Lambe, 1894 on activity of natural β-1,3-D-glucanase from the marine fungus Chaetomium indicum Corda, 1840 and the marine bivalve mollusk Spisula sachalinensis, Schrenck, 1861. Rus. J. Mar. Biol. 2018, 44, 127–134. [Google Scholar] [CrossRef]
- Parsons, Z.D.; Gates, K.S. Redox regulation of protein tyrosine phosphatases: Methods for kinetic analysis of covalent enzyme inactivation. Methods Enzymol. 2013, 528, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Bakunina, I.; Slepchenko, L.; Anastyuk, S.; Isakov, V.; Likhatskaya, G.; Kim, N.; Tekutyeva, L.; Son, O.; Balabanova, L. Characterization of properties and transglycosylation abilities of recombinant α–galactosidase from cold adapted marine bacterium Pseudoalteromonas KMM 701 and its C494N, D451A mutants. Mar. Drugs 2018, 16, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredslund, F.; Hachem, M.A.; Larsen, R.J.; Sørensen, P.G.; Coutinho, P.M.; Lo Leggio, L.; Svensson, B. Crystal structure of α-galactosidase from Lactobacillus acidophilus NCFM: Insight into tetramer formation and substrate binding. J. Mol Biol. 2011, 412, 466–480. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | v/v0 (%) | Compound | v/v0 (%) |
|---|---|---|---|
| 1 | 79 1 | 6 | 87 2 |
| 2 | 0 1 | 7 | 67 2 |
| 3 | 0 1 | 8 | 28 2 |
| 4 | 100 1 | 9 | 0 2 |
| 5 | 87 2 | 6.4% EtOH | 100 |
| Compound | Structure | Ki (µM) 1 | kinact (min−1) 2 | n 3 |
|---|---|---|---|---|
| aaptamine (1) |  | nd 4 | ||
| isoaaptamine (2) |  | 2.70 ± 0.42 | 0.12 ± 0.01 | 2.17 ± 0.55 |
| 9-demethylaaptamine(3) |  | 300 ± 77 | 0.092 ± 0.008 | 0.84 ± 0.12 |
| aaptanone (4) |  | nd 4 | ||
| damirone A (5) |  | nd 4 | ||
| ;damirone B (6) |  | nd 4 | ||
| makaluvamine H (7) |  | nd 4 | ||
| makaluvamine G (8) |  | 411 ± 86 | 0.079 ± 0.011 | 2.5 ± 0.97 |
| zyzzyanone A (9) |  | 105 ± 9 | 0.037 ± 0.003 | 4.10 ± 1.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Utkina, N.; Likhatskaya, G.; Malyarenko, O.; Ermakova, S.; Balabanova, L.; Slepchenko, L.; Bakunina, I. Effects of Sponge-Derived Alkaloids on Activities of the Bacterial α-D-Galactosidase and Human Cancer Cell α-N-Acetylgalactosaminidase. Biomedicines 2021, 9, 510. https://doi.org/10.3390/biomedicines9050510
Utkina N, Likhatskaya G, Malyarenko O, Ermakova S, Balabanova L, Slepchenko L, Bakunina I. Effects of Sponge-Derived Alkaloids on Activities of the Bacterial α-D-Galactosidase and Human Cancer Cell α-N-Acetylgalactosaminidase. Biomedicines. 2021; 9(5):510. https://doi.org/10.3390/biomedicines9050510
Chicago/Turabian StyleUtkina, Natalia, Galina Likhatskaya, Olesya Malyarenko, Svetlana Ermakova, Larisa Balabanova, Lubov Slepchenko, and Irina Bakunina. 2021. "Effects of Sponge-Derived Alkaloids on Activities of the Bacterial α-D-Galactosidase and Human Cancer Cell α-N-Acetylgalactosaminidase" Biomedicines 9, no. 5: 510. https://doi.org/10.3390/biomedicines9050510