
Promising Activities of Marine Natural Products against Hematopoietic Malignancies



Abstract
:
1. Introduction
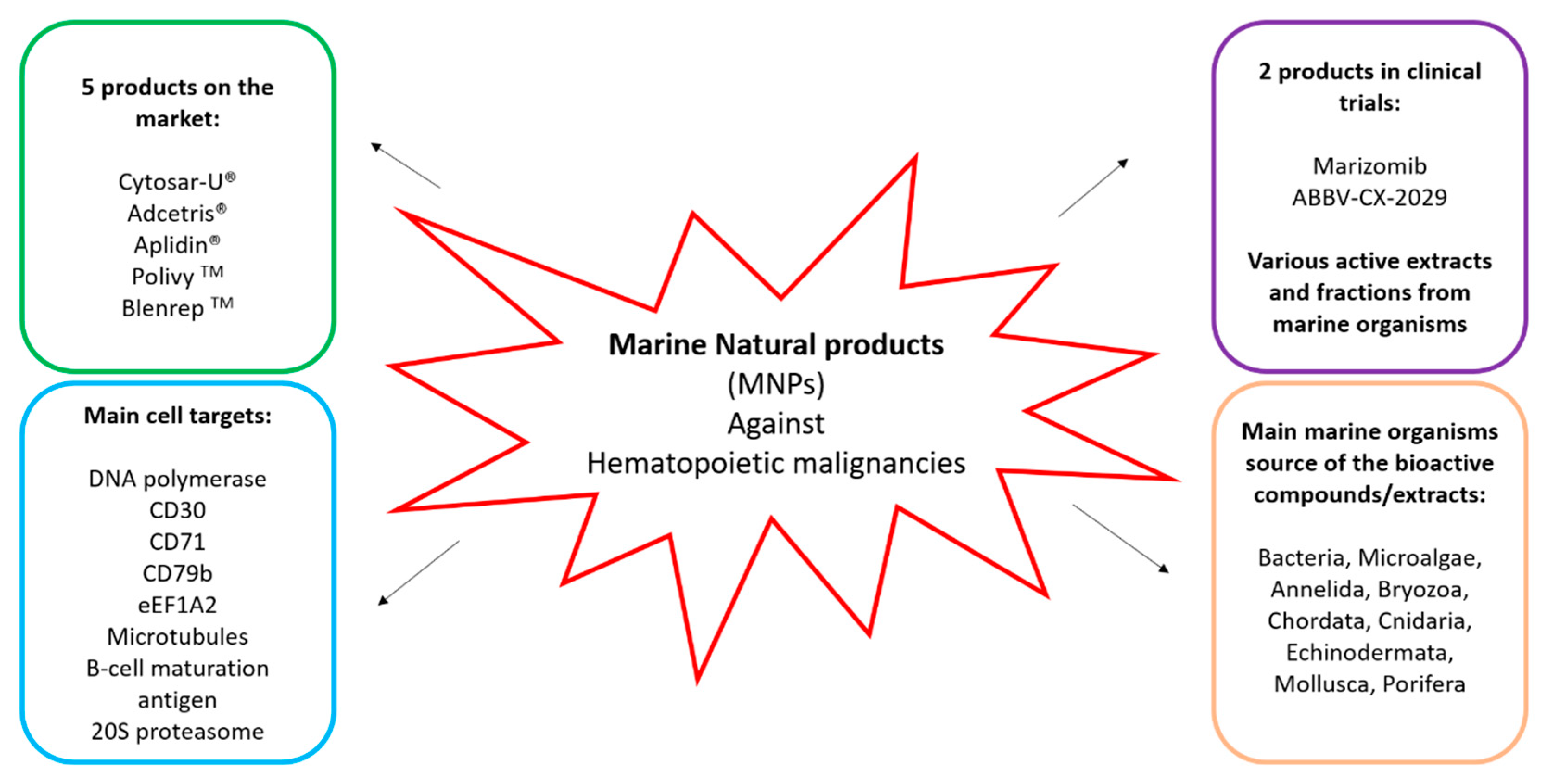
2. Compounds on the Market or in Clinical Trials
2.1. Marine Compounds Already Commercially Available and Currently in Use
2.2. Marine Compounds in Clinical Trials
3. Microorganisms
4. Macroorganisms
5. Multi-Omics Approaches for Drug Discovery
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martínez Andrade, K.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffe, E.; Harris, N.; Stein, H.; Vardiman, J. Pathology and Genetics of Tumours of Haematopoietic and Lymphoid Tissues; IARC: Lyon, France, 2001; ISBN 978-92-832-2411-2. [Google Scholar]
- Swerdlow, S.; Campo, E.; Harris, N.; Jaffe, E.; Pileri, P.; Stein, H.; Thiele, J.; Vardiman, J. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; IARC: Lyon, France, 2008; ISBN 978-92-832-2431-0. [Google Scholar]
- Vardiman, J.W. The World Health Organization (WHO) Classification of Tumors of the Hematopoietic and Lymphoid Tissues: An Overview with Emphasis on the Myeloid Neoplasms. Chem. Biol. Interact. 2010, 184, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Damiano, S.; Montagnaro, S.; Puzio, M.V.; Severino, L.; Pagnini, U.; Barbarino, M.; Cesari, D.; Giordano, A.; Florio, S.; Ciarcia, R. Effects of Antioxidants on Apoptosis Induced by Dasatinib and Nilotinib in K562 Cells. J. Cell Biochem. 2018, 119, 4845–4854. [Google Scholar] [CrossRef]
- Montagnaro, S.; Damiano, S.; Ciarcia, R.; Puzio, M.V.; Ferrara, G.; Iovane, V.; Forte, I.M.; Giordano, A.; Pagnini, U. Caprine Herpesvirus 1 (CpHV-1) as a Potential Candidate for Oncolytic Virotherapy. Cancer Biol. Ther. 2019. [Google Scholar] [CrossRef] [PubMed]
- Noble, R.L. The Discovery of the Vinca Alkaloids—Chemotherapeutic Agents against Cancer. Biochem. Cell Biol. 2011. [Google Scholar] [CrossRef]
- Agrawal, K. Doxorubicin. In xPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier: New York, NY, USA, 2007; pp. 1–5. ISBN 978-0-08-055232-3. [Google Scholar]
- Pavel, K.J.; Tomas, E.; Pavel, K. Biological Therapy of Hematologic Malignancies: Toward a Chemotherapy- Free Era. Curr. Med. Chem. 2019, 26, 1002–1018. [Google Scholar]
- Matsueda, S.; Chodon, T.; Koya, R.C. Cellular Immunotherapy in the Treatment of Hematopoietic Malignancies. In Leukemia Stem Cells in Hematologic Malignancies; Zhang, H., Li, S., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2019; pp. 217–229. ISBN 9789811373428. [Google Scholar]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine Microorganisms as a Promising and Sustainable Source of Bioactive Molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Jaspars, M.; De Pascale, D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The Marine Biodiscovery Pipeline and Ocean Medicines of Tomorrow. J. Mar. Biol. Assoc. UK 2016, 96, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [Green Version]
- Barreca, M.; Spanò, V.; Montalbano, A.; Cueto, M.; Díaz Marrero, A.R.; Deniz, I.; Erdoğan, A.; Lukić Bilela, L.; Moulin, C.; Taffin-de-Givenchy, E.; et al. Marine Anticancer Agents: An Overview with a Particular Focus on Their Chemical Classes. Mar. Drugs 2020, 18, 619. [Google Scholar] [CrossRef]
- Lindequist, U. Marine-Derived Pharmaceuticals—Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritano, C.; Rizzo, C.; Lo Giudice, A.; Saggiomo, M. Physiological and Molecular Responses to Main Environmental Stressors of Microalgae and Bacteria in Polar Marine Environments. Microorganisms 2020, 8, 1957. [Google Scholar] [CrossRef]
- Ianora, A.; Miralto, A. Toxigenic Effects of Diatoms on Grazers, Phytoplankton and Other Microbes: A Review. Ecotoxicology 2010, 19, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Martínez, K.A.; Lauritano, C.; Druka, D.; Romano, G.; Grohmann, T.; Jaspars, M.; Martín, J.; Díaz, C.; Cautain, B.; de la Cruz, M.; et al. Amphidinol 22, a New Cytotoxic and Antifungal Amphidinol from the Dinoflagellate. Amphidinium Carterae Mar. Drugs 2019, 17, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritano, C.; Romano, G.; Roncalli, V.; Amoresano, A.; Fontanarosa, C.; Bastianini, M.; Braga, F.; Carotenuto, Y.; Ianora, A. New Oxylipins Produced at the End of a Diatom Bloom and Their Effects on Copepod Reproductive Success and Gene Expression Levels. Harmful Algae 2016, 55, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Carotenuto, Y.; Procaccini, G.; Turner, J.T.; Ianora, A. Changes in Expression of Stress Genes in Copepods Feeding upon a Non-Brevetoxin-Producing Strain of the Dinoflagellate Karenia Brevis. Harmful Algae 2013, 28, 23–30. [Google Scholar] [CrossRef]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the Health Potential of Microalgae as Sustainable Sources of Bioactive Compounds. Int. J. Mol. Sci. 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
- D’Ambra, I.; Lauritano, C. A Review of Toxins from Cnidaria. Mar. Drugs 2020, 18, 507. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Martínez, K.A.; Battaglia, P.; Granata, A.; de la Cruz, M.; Cautain, B.; Martín, J.; Reyes, F.; Ianora, A.; Guglielmo, L. First Evidence of Anticancer and Antimicrobial Activity in Mediterranean Mesopelagic Species. Sci. Rep. 2020, 10, 4929. [Google Scholar] [CrossRef]
- Ruocco, N.; Costantini, S.; Zupo, V.; Lauritano, C.; Caramiello, D.; Ianora, A.; Budillon, A.; Romano, G.; Nuzzo, G.; D’Ippolito, G.; et al. Toxigenic Effects of Two Benthic Diatoms upon Grazing Activity of the Sea Urchin: Morphological, Metabolomic and de Novo Transcriptomic Analysis. Sci. Rep. 2018, 8, 5622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritano, C.; Carotenuto, Y.; Vitiello, V.; Buttino, I.; Romano, G.; Hwang, J.-S.; Ianora, A. Effects of the Oxylipin-Producing Diatom Skeletonema Marinoi on Gene Expression Levels of the Calanoid Copepod Calanus Sinicus. Mar. Genom. 2015, 24, 89–94. [Google Scholar] [CrossRef]
- Schwartsmann, G.; da Rocha, A.B.; Berlinck, R.G.; Jimeno, J. Marine Organisms as a Source of New Anticancer Agents. Lancet Oncol. 2001, 2, 221–225. [Google Scholar] [CrossRef]
- Salvador-Reyes, L.A.; Luesch, H. Biological Targets and Mechanisms of Action of Natural Products from Marine Cyanobacteria. Nat. Prod. Rep. 2015, 32, 478–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, A.M.; Devine, M.; DeRemer, D. Brentuximab Vedotin: An Anti-CD30 Antibody-Drug Conjugate. Am. J. Health Syst. Pharm. 2013, 70, 589–597. [Google Scholar] [CrossRef]
- Vaklavas, C.; Forero-Torres, A. Safety and Efficacy of Brentuximab Vedotin in Patients with Hodgkin Lymphoma or Systemic Anaplastic Large Cell Lymphoma. Ther. Adv. Hematol. 2012, 3, 209–225. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Alonso, M.J.; González-Santiago, L.; Martínez, T.; Losada, A.; Galmarini, C.M.; Muñoz, A. The Mechanism of Action of Plitidepsin. Curr. Opin. Investig. Drugs 2009, 10, 536–542. [Google Scholar] [PubMed]
- White, K.M.; Rosales, R.; Yildiz, S.; Kehrer, T.; Miorin, L.; Moreno, E.; Jangra, S.; Uccellini, M.B.; Rathnasinghe, R.; Coughlan, L.; et al. Plitidepsin Has Potent Preclinical Efficacy against SARS-CoV-2 by Targeting the Host Protein EEF1A. Science 2021, 371, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Bourbon, E.; Salles, G. Polatuzumab Vedotin: An Investigational Anti-CD79b Antibody Drug Conjugate for the Treatment of Diffuse Large B-Cell Lymphoma. Expert Opin. Investig. Drugs 2020, 29. [Google Scholar] [CrossRef] [PubMed]
- Markham, A. Belantamab Mafodotin: First Approval. Drugs 2020, 80, 1607–1613. [Google Scholar] [CrossRef]
- Kale, A.J.; McGlinchey, R.P.; Lechner, A.; Moore, B.S. Bacterial Self-Resistance to the Natural Proteasome Inhibitor Salinosporamide A. ACS Chem. Biol. 2011, 6, 1257–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Diao, A. Marizomib, a Potent Second Generation Proteasome Inhibitor from Natural Origin. Anticancer. Agents Med. Chem. 2015, 15, 298–306. [Google Scholar] [CrossRef]
- Richardson, P.G.; Zimmerman, T.M.; Hofmeister, C.C.; Talpaz, M.; Chanan-Khan, A.A.; Kaufman, J.L.; Laubach, J.P.; Chauhan, D.; Jakubowiak, A.J.; Reich, S.; et al. Phase 1 Study of Marizomib in Relapsed or Relapsed and Refractory Multiple Myeloma: NPI-0052-101 Part 1. Blood 2016, 127, 2693–2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badros, A.; Singh, Z.; Dhakal, B.; Kwok, Y.; MacLaren, A.; Richardson, P.; Trikha, M.; Hari, P. Marizomib for Central Nervous System-Multiple Myeloma. Br. J. Haematol. 2017, 177, 221–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.L.; El-Khoueiry, A.B.; Hafez, N.; Lakhani, N.J.; Mamdani, H.; Rodon Ahnert, J.; Sanborn, R.E.; Ho, T.; Li, R.; Waldes, J.; et al. CX-2029, a PROBODY Drug Conjugate Targeting CD71 (Transferrin Receptor): Results from a First-in-Human Study (PROCLAIM-CX-2029) in Patients (Pts) with Advanced Cancer. JCO 2020, 38, 3502. [Google Scholar] [CrossRef]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Light and Temperature Effects on Bioactivity in Diatoms. J. Appl. Phycol. 2016, 28, 939–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osvik, R.D.; Ingebrigtsen, R.A.; Norrbin, M.F.; Andersen, J.H.; Eilertsen, H.C.; Hansen, E.H. Adding Zooplankton to the OSMAC Toolkit: Effect of Grazing Stress on the Metabolic Profile and Bioactivity of a Diatom. Mar. Drugs 2021, 19, 87. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big Effects from Small Changes: Possible Ways to Explore Nature’s Chemical Diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Lauritano, C.; Martín, J.; de la Cruz, M.; Reyes, F.; Romano, G.; Ianora, A. First Identification of Marine Diatoms with Anti-Tuberculosis Activity. Sci. Rep. 2018, 8, 2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kjer, J.; Sendker, J.; Wray, V.; Guan, H.; Edrada, R.; Lin, W.; Wu, J.; Proksch, P. Chromones from the Endophytic Fungus Pestalotiopsis Sp. Isolated from the Chinese Mangrove Plant Rhizophora Mucronata. J. Nat. Prod. 2009, 72, 662–665. [Google Scholar] [CrossRef] [PubMed]
- Shigemori, H.; Bae, M.A.; Yazawa, K.; Sasaki, T.; Kobayashi, J. Alteramide A, a New Tetracyclic Alkaloid from a Bacterium Alteromonas Sp. Associated with the Marine Sponge Halichondria Okadai. J. Org. Chem. 1992, 57, 4317–4320. [Google Scholar] [CrossRef]
- Fenical, W. Chemical Studies of Marine Bacteria: Developing a New Resource. Chem. Rev. 1993, 93, 1673–1683. [Google Scholar] [CrossRef]
- Campàs, C.; Dalmau, M.; Montaner, B.; Barragán, M.; Bellosillo, B.; Colomer, D.; Pons, G.; Pérez-Tomás, R.; Gil, J. Prodigiosin Induces Apoptosis of B and T Cells from B-Cell Chronic Lymphocytic Leukemia. Leukemia 2003, 17, 746–750. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.; Jaisankar, P.; Das, S.; Sarkar, K.K.; Roy, S.; Besra, S.E.; Vedasiromani, J.R.; Ghosh, D.; Sana, B.; Mukherjee, J. Production and Purification of a Bioactive Substance Inhibiting Multiple Drug Resistant Bacteria and Human Leukemia Cells from a Salt-Tolerant Marine Actinobacterium Sp. Isolated from the Bay of Bengal. Biotechnol. Lett. 2006, 28, 1083–1088. [Google Scholar] [CrossRef]
- Singh, A.V.; Bandi, M.; Raje, N.; Richardson, P.; Palladino, M.A.; Chauhan, D.; Anderson, K.C. A Novel Vascular Disrupting Agent Plinabulin Triggers JNK-Mediated Apoptosis and Inhibits Angiogenesis in Multiple Myeloma Cells. Blood 2011, 117, 5692–5700. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Herfindal, L.; Jokela, J.; Shishido, T.; Wahlsten, M.; Døskeland, S.; Sivonen, K. Cyanobacteria from Terrestrial and Marine Sources Contain Apoptogens Able to Overcome Chemoresistance in Acute Myeloid Leukemia Cells. Mar. Drugs 2014, 12, 2036–2053. [Google Scholar] [CrossRef]
- Oftedal, L.; Selheim, F.; Wahlsten, M.; Sivonen, K.; Døskeland, S.O.; Herfindal, L. Marine Benthic Cyanobacteria Contain Apoptosis-Inducing Activity Synergizing with Daunorubicin to Kill Leukemia Cells, but Not Cardiomyocytes. Mar. Drugs 2010, 8, 2659–2672. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.A.; Goeger, D.E.; Hills, P.; Mooberry, S.L.; Huang, N.; Romero, L.I.; Ortega-Barría, E.; Gerwick, W.H.; McPhail, K.L. Coibamide A, a Potent Antiproliferative Cyclic Depsipeptide from the Panamanian Marine Cyanobacterium Leptolyngbya Sp. J. Am. Chem. Soc. 2008, 130, 6324–6325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrasidlo, W.; Mielgo, A.; Torres, V.A.; Barbero, S.; Stoletov, K.; Suyama, T.L.; Klemke, R.L.; Gerwick, W.H.; Carson, D.A.; Stupack, D.G. The Marine Lipopeptide Somocystinamide A Triggers Apoptosis via Caspase 8. Proc. Natl. Acad. Sci. USA 2008, 105, 2313–2318. [Google Scholar] [CrossRef] [Green Version]
- Foster, B.J.; Fortuna, M.; Media, J.; Wiegand, R.A.; Valeriote, F.A. Cryptophycin 1 Cellular Levels and Effects in Vitro Using L1210 Cells. Investig. New Drugs 1998, 16, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Tan, L.T. Lagunamides A and B: Cytotoxic and Antimalarial Cyclodepsipeptides from the Marine Cyanobacterium Lyngbya Majuscula. J. Nat. Prod. 2010, 73, 1810–1814. [Google Scholar] [CrossRef]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Chan, K.P.; Chen, D.Y.-K.; Tan, L.T. Lagunamide C, a Cytotoxic Cyclodepsipeptide from the Marine Cyanobacterium Lyngbya Majuscula. Phytochemistry 2011, 72, 2369–2375. [Google Scholar] [CrossRef]
- Nagle, D.G.; Zhou, Y.-D.; Mora, F.D.; Mohammed, K.A.; Kim, Y.-P. Mechanism Targeted Discovery of Antitumor Marine Natural Products. Curr. Med. Chem. 2004, 11, 1725–1756. [Google Scholar] [CrossRef] [PubMed]
- Patterson, G.M.; Carmeli, S. Biological Effects of Tolytoxin (6-Hydroxy-7-O-Methyl-Scytophycin b), a Potent Bioactive Metabolite from Cyanobacteria. Arch. Microbiol. 1992, 157, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Khan, Q.A.; Lu, J.; Hecht, S.M. Calothrixins, a New Class of Human DNA Topoisomerase I Poisons. J. Nat. Prod. 2009, 72, 438–442. [Google Scholar] [CrossRef]
- Prestegard, S.K. Marine Benthic Diatoms Contain Compounds Able to Induce Leukemia Cell Death and Modulate Blood Platelet Activity. Mar. Drugs 2009, 7, 605–623. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.R.; Samarakoon, K.W.; Ko, J.; Lakmal, H.H.C.; Lee, J.; An, S.; Jeon, Y.; Lee, J. Potentiality of Benthic Dinoflagellate Cultures and Screening of Their Bioactivities in Jeju Island, Korea. Afr. J. Biotechnol. 2014, 13, 792–805. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; Ko, J.-Y.; Shah, M.R.; Lee, J.-H.; Kang, M.-C.; O-Nam, K.; Lee, J.-B.; Jeon, Y.-J. In Vitro Studies of Anti-Inflammatory and Anticancer Activities of Organic Solvent Extracts from Cultured Marine Microalgae. Algae 2013, 28, 111–119. [Google Scholar] [CrossRef]
- Atasever-Arslan, B.; Yilancioglu, K.; Kalkan, Z.; Timucin, A.C.; Gür, H.; Isik, F.B.; Deniz, E.; Erman, B.; Cetiner, S. Screening of New Antileukemic Agents from Essential Oils of Algae Extracts and Computational Modeling of Their Interactions with Intracellular Signaling Nodes. Eur. J. Pharm. Sci. 2016, 83, 120–131. [Google Scholar] [CrossRef]
- Andrade, L.M. Chlorella and Spirulina Microalgae as Sources of Functional Foods, Nutraceuticals, and Food Supplements; an Overview. MOJFPT 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Ruela-de-Sousa, R.R.; Fuhler, G.M.; Blom, N.; Ferreira, C.V.; Aoyama, H.; Peppelenbosch, M.P. Cytotoxicity of Apigenin on Leukemia Cell Lines: Implications for Prevention and Therapy. Cell Death Dis. 2010, 1, e19. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, J.; Tsuda, M. Bioactive Products from Okinawan Marine Micro- and Macroorganisms. Phytochem. Rev. 2004, 3, 267–274. [Google Scholar] [CrossRef]
- Kerr, R.G.; Kerr, S.S. Marine Natural Products as Therapeutic Agents. Expert Opin. Ther. Pat. 1999, 9, 1207–1222. [Google Scholar] [CrossRef]
- Samarakoon, K.W.; Ko, J.-Y.; Lee, J.-H.; Kwon, O.-N.; Kim, S.-W.; Jeon, Y.-J. Apoptotic Anticancer Activity of a Novel Fatty Alcohol Ester Isolated from Cultured Marine Diatom, Phaeodactylum tricornutum. J. Funct. Foods 2014, 6, 231–240. [Google Scholar] [CrossRef]
- Rowland, S.J.; Belt, S.T.; Wraige, E.J.; Massé, G.; Roussakis, C.; Robert, J.M. Effects of Temperature on Polyunsaturation in Cytostatic Lipids of Haslea ostrearia. Phytochemistry 2001, 56, 597–602. [Google Scholar] [CrossRef]
- Miceli, M.; Cutignano, A.; Conte, M.; Ummarino, R.; Romanelli, A.; Ruvo, M.; Leone, M.; Mercurio, F.A.; Doti, N.; Manzo, E.; et al. Monoacylglycerides from the Diatom Skeletonema marinoi Induce Selective Cell Death in Cancer Cells. Mar. Drugs 2019, 17, 625. [Google Scholar] [CrossRef] [Green Version]
- Mondal, A.; Bose, S.; Banerjee, S.; Patra, J.K.; Malik, J.; Mandal, S.K.; Kilpatrick, K.L.; Das, G.; Kerry, R.G.; Fimognari, C.; et al. Marine Cyanobacteria and Microalgae Metabolites—A Rich Source of Potential Anticancer Drugs. Mar. Drugs 2020, 18, 476. [Google Scholar] [CrossRef] [PubMed]
- Kerksiek, K.; Mejillano, M.R.; Schwartz, R.E.; Georg, G.I.; Himes, R.H. Interaction of Cryptophycin 1 with Tubulin and Microtubules. FEBS Lett. 1995, 377, 59–61. [Google Scholar] [CrossRef] [Green Version]
- Cragg, G.M.; Newman, D.J.; Yang, S.S. Natural Product Extracts of Plant and Marine Origin Having Antileukemia Potential. The NCI Experience. J. Nat. Prod. 2006, 69, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K. Fucoxanthin and Its Deacetylated Product, Fucoxanthinol, Induce Apoptosis of Primary Effusion Lymphomas. Cancer Lett. 2011, 300, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Wanezaki, S.; Miyauchi, K.; Kurihara, H.; Kohno, H.; Kawabata, J.; Odashima, S.; Takahashi, K. Apoptosis-Inducing Effect of Fucoxanthin on Human Leukemia Cell Line HL-60. Food Sci. Technol. Res. 1999, 5, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-N.; Heo, S.-J.; Kang, S.-M.; Ahn, G.; Jeon, Y.-J. Fucoxanthin Induces Apoptosis in Human Leukemia HL-60 Cells through a ROS-Mediated Bcl-XL Pathway. Toxicol Vitro 2010, 24, 1648–1654. [Google Scholar] [CrossRef]
- Ganesan, P.; Noda, K.; Manabe, Y.; Ohkubo, T.; Tanaka, Y.; Maoka, T.; Sugawara, T.; Hirata, T. Siphonaxanthin, a Marine Carotenoid from Green Algae, Effectively Induces Apoptosis in Human Leukemia (HL-60) Cells. Biochim. Biophys. Acta Gen. Subj. 2011, 1810, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, C.; Tafuku, S.; Kadekaru, T.; Sawada, S.; Tomita, M.; Okudaira, T.; Nakazato, T.; Toda, T.; Uchihara, J.-N.; Taira, N.; et al. Anti-Adult T-Cell Leukemia Effects of Brown Algae Fucoxanthin and Its Deacetylated Product, Fucoxanthinol. Int. J. Cancer 2008, 123, 2702–2712. [Google Scholar] [CrossRef] [PubMed]
- Konishi, I.; Hosokawa, M.; Sashima, T.; Kobayashi, H.; Miyashita, K. Halocynthiaxanthin and Fucoxanthinol Isolated from Halocynthia roretzi Induce Apoptosis in Human Leukemia, Breast and Colon Cancer Cells. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 53–59. [Google Scholar] [CrossRef]
- Yamamoto, I.; Takahashi, M.; Tamura, E.; Maruyama, H.; Mori, H. Antitumor Activity of Edible Marine Algae: Effect of Crude Fucoidan Fractions Prepared from Edible Brown Seaweeds against L-1210 Leukemia. In Proceedings of the Eleventh International Seaweed Symposium; Bird, C.J., Ragan, M.A., Eds.; Springer: Dordrecht, The Netherlands, 1984; pp. 145–148. [Google Scholar]
- Li, G.; Zhang, L.; Liu, J.; Chen, S.; Xiao, T.; Liu, G.; Wang, J.; Wang, L.; Hou, M. Marine Drug Haishengsu Increases Chemosensitivity to Conventional Chemotherapy and Improves Quality of Life in Patients with Acute Leukemia. Biomed. Pharmacother. 2016, 81, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-Y.; Liu, J.-Z.; Chen, S.-G.; Zhang, B.; Wang, C.-B.; Wang, L.-X. Tegillarca granosa Extract Haishengsu Inhibits the Expression of P-Glycoprotein and Induces Apoptosis in Drug-Resistant K562/ADM Cells. Pharm. Biol. 2010, 48, 529–533. [Google Scholar] [CrossRef]
- Miyamoto, T.; Kodama, K.; Aramaki, Y.; Higuchi, R.; Van Soest, R.W.M. Orostanal, a Novel Abeo-Sterol Inducing Apoptosis in Leukemia Cell from a Marine Sponge, Stelletta hiwasaensis. Tetrahedron Lett. 2001, 42, 6349–6351. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Lu, M.-C.; El-Shazly, M.; Lai, K.-H.; Wu, T.-Y.; Hsu, Y.-M.; Lee, Y.-L.; Liu, Y.-C. Breaking down Leukemia Walls: Heteronemin, a Sesterterpene Derivative, Induces Apoptosis in Leukemia Molt4 Cells through Oxidative Stress, Mitochondrial Dysfunction and Induction of Talin Expression. Mar. Drugs 2018, 16, 212. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xu, B.; Cui, J.; Deng, Z.; de Voogd, N.J.; Proksch, P.; Lin, W. Globostelletins A–I, Cytotoxic Isomalabaricane Derivatives from the Marine Sponge Rhabdastrella globostellata. Bioorg. Med. Chem. 2010, 18, 4639–4647. [Google Scholar] [CrossRef]
- Kong, D.; Aoki, S.; Sowa, Y.; Sakai, T.; Kobayashi, M. Smenospongine, a Sesquiterpene Aminoquinone from a Marine Sponge, Induces G1 Arrest or Apoptosis in Different Leukemia Cells. Mar. Drugs 2008, 6, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Aoki, S.; Matsui, K.; Tanaka, K.; Satari, R.; Kobayashi, M. Lembehyne A, a Novel Neuritogenic Polyacetylene, from a Marine Sponge of Haliclona sp. Tetrahedron 2000, 56, 9945–9948. [Google Scholar] [CrossRef]
- Aoki, S.; Matsui, K.; Takata, T.; Hong, W.; Kobayashi, M. Lembehyne A, a Spongean Polyacetylene, Induces Neuronal Differentiation in Neuroblastoma Cell. Biochem. Biophys. Res. Commun. 2001, 289, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Aoki, S.; Matsui, K.; Wei, H.; Murakami, N.; Kobayashi, M. Structure–Activity Relationship of Neuritogenic Spongean Acetylene Alcohols, Lembehynes. Tetrahedron 2002, 58, 5417–5422. [Google Scholar] [CrossRef]
- Song, J.; Jeon, J.-E.; Won, T.H.; Sim, C.J.; Oh, D.-C.; Oh, K.-B.; Shin, J. New Cyclic Cystine Bridged Peptides from the Sponge Suberites Waedoensis. Mar. Drugs 2014, 12, 2760–2770. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.-K.; Song, I.-H.; Park, H.Y.; Lee, Y.-J.; Lee, H.-S.; Sim, C.J.; Oh, D.-C.; Oh, K.-B.; Shin, J. Suvanine Sesterterpenes and Deacyl Irciniasulfonic Acids from a Tropical Coscinoderma Sp. Sponge. J. Nat. Prod. 2014, 77, 1396–1403. [Google Scholar] [CrossRef]
- Woo, J.-K.; Jeon, J.; Kim, C.-K.; Sim, C.J.; Oh, D.-C.; Oh, K.-B.; Shin, J. Gombamide A, a Cyclic Thiopeptide from the Sponge Clathria gombawuiensis. J. Nat. Prod. 2013, 76, 1380–1383. [Google Scholar] [CrossRef] [PubMed]
- Towle, M.J.; Salvato, K.A.; Budrow, J.; Wels, B.F.; Kuznetsov, G.; Aalfs, K.K.; Welsh, S.; Zheng, W.; Seletsky, B.M.; Palme, M.H.; et al. In Vitro and in Vivo Anticancer Activities of Synthetic Macrocyclic Ketone Analogues of Halichondrin B. Cancer Res. 2001, 61, 1013–1021. [Google Scholar] [PubMed]
- Pettit, G.R.; Cichacz, Z.; Barkoczy, J.; Dorsaz, A.-C.; Herald, D.L.; Williams, M.D.; Doubek, D.L.; Schmidt, J.M.; Tackett, L.P.; Brune, D.C.; et al. Isolation and Structure of the Marine Sponge Cell Growth Inhibitory Cyclic Peptide Phakellistatin 1. J. Nat. Prod. 1993, 56, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Tan, R.; Williams, M.D.; Tackett, L.; Schmidt, J.M.; Cerny, R.L.; Hooper, J.N.A. Isolation and Structure of Phakellistatin 2 from the Eastern Indian Ocean Marine Sponge Phakellia carteri. Bioorg. Med. Chem. Lett. 1993, 3, 2869–2874. [Google Scholar] [CrossRef]
- Meli, A.; Tedesco, C.; Della Sala, G.; Schettini, R.; Albericio, F.; De Riccardis, F.; Izzo, I. Phakellistatins: An Underwater Unsolved Puzzle. Mar. Drugs 2017, 15, 78. [Google Scholar] [CrossRef] [Green Version]
- Pettit, G.R.; Tan, R. Isolation and Structure of Phakellistatin 14 from the Western Pacific Marine Sponge Phakellia Sp.,1. J. Nat. Prod. 2005, 68, 60–63. [Google Scholar] [CrossRef]
- Coleman, J.E.; Dilip de Silva, E.; Kong, F.; Andersen, R.J.; Allen, T.M. Cytotoxic Peptides from the Marine Sponge Cymbastela sp. Tetrahedron 1995, 51, 10653–10662. [Google Scholar] [CrossRef]
- Nakao, Y.; Yeung, B.K.S.; Yoshida, W.Y.; Scheuer, P.J.; Kelly-Borges, M. Kapakahine B, a Cyclic Hexapeptide with an .Alpha.-Carboline Ring System from the Marine Sponge Cribrochalina olemda. J. Am. Chem. Soc. 1995, 117, 8271–8272. [Google Scholar] [CrossRef]
- Matsunaga, S.; Fusetani, N. Theonellamides A-E, Cytotoxic Bicyclic Peptides, from a Marine Sponge Theonella sp. J. Org. Chem. 1995, 60, 1177–1181. [Google Scholar] [CrossRef]
- Hamada, T.; Sugawara, T.; Matsunaga, S.; Fusetani, N. Polytheonamides, Unprecedented Highly Cytotoxic Polypeptides, from the Marine Sponge Theonella swinhoei: 1. Isolation and Component Amino Acids. Tetrahedron Lett. 1994, 35, 719–720. [Google Scholar] [CrossRef]
- Tabudravu, J.; Morris, L.A.; Kettenes-van den Bosch, J.J.; Jaspars, M. Wainunuamide, a Histidine-Containing Proline-Rich Cyclic Heptapeptide Isolated from the Fijian Marine Sponge Stylotella aurantium. Tetrahedron Lett. 2001, 42, 9273–9276. [Google Scholar] [CrossRef]
- Cioca, D.P.; Kitano, K. Induction of Apoptosis and CD10/Neutral Endopeptidase Expression by Jaspamide in HL-60 Line Cells. CMLS Cell. Mol. Life Sci. 2002, 59, 1377–1387. [Google Scholar] [CrossRef]
- Dassonneville, L.; Wattez, N.; Baldeyrou, B.; Mahieu, C.; Lansiaux, A.; Banaigs, B.; Bonnard, I.; Bailly, C. Inhibition of Topoisomerase II by the Marine Alkaloid Ascididemin and Induction of Apoptosis in Leukemia Cells. Biochem. Pharmacol. 2000, 60, 527–537. [Google Scholar] [CrossRef]
- Appleton, D.R.; Chuen, C.S.; Berridge, M.V.; Webb, V.L.; Copp, B.R. Rossinones A and B, Biologically Active Meroterpenoids from the Antarctic Ascidian, Aplidium Species. J. Org. Chem. 2009, 74, 9195–9198. [Google Scholar] [CrossRef]
- Sperlich, J.; Kerr, R.; Teusch, N. The Marine Natural Product Pseudopterosin Blocks Cytokine Release of Triple-Negative Breast Cancer and Monocytic Leukemia Cells by Inhibiting NF-κB Signaling. Mar. Drugs 2017, 15, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varterasian, M.L.; Mohammad, R.M.; Shurafa, M.S.; Hulburd, K.; Pemberton, P.A.; Rodriguez, D.H.; Spadoni, V.; Eilender, D.S.; Murgo, A.; Wall, N.; et al. Phase II Trial of Bryostatin 1 in Patients with Relapsed Low-Grade Non-Hodgkin’s Lymphoma and Chronic Lymphocytic Leukemia. Clin. Cancer Res. 2000, 6, 825–828. [Google Scholar] [PubMed]
- Zhang, Y.; Yi, Y. Studies on antitumor activities of triterpene glycoside colochiroside A from sea cucumber Colochirus anceps. Zhongguo Zhong Yao Za Zhi 2011, 36, 504–507. [Google Scholar] [CrossRef]
- Sun, G.-Q.; Li, L.; Yi, Y.-H.; Yuan, W.-H.; Liu, B.-S.; Weng, Y.-Y.; Zhang, S.-L.; Sun, P.; Wang, Z.-L. Two New Cytotoxic Nonsulfated Pentasaccharide Holostane (=20-Hydroxylanostan-18-Oic Acid γ-Lactone) Glycosides from the Sea Cucumber Holothuria grisea. Helv. Chim. Acta 2008, 91, 1453–1460. [Google Scholar] [CrossRef]
- Zhao, Q.; Xue, Y.; Liu, Z.; Li, H.; Wang, J.; Li, Z.; Wang, Y.; Dong, P.; Xue, C. Differential Effects of Sulfated Triterpene Glycosides, Holothurin A1, and 24-Dehydroechinoside A, on Antimetastasic Activity via Regulation of the MMP-9 Signal Pathway. J. Food Sci. 2010, 75, H280–H288. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Xu, Q.-Z.; Tang, H.-F.; Yi, Y.-H.; Gong, W. Cytotoxic Holostane-Type Triterpene Glycosides from the Sea Cucumber Pentacta quadrangularis. Planta. Med. 2010, 76, 1900–1904. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.-H.; Park, E.-S.; Shin, S.-W.; Na, Y.-W.; Han, J.-Y.; Jeong, J.-S.; Shastina, V.V.; Stonik, V.A.; Park, J.-I.; Kwak, J.-Y. Stichoposide C Induces Apoptosis through the Generation of Ceramide in Leukemia and Colorectal Cancer Cells and Shows In Vivo Antitumor Activity. Clin. Cancer Res. 2012, 18, 5934–5948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayed, Y.; Sghaier, R.M.; Laouini, D.; Bacha, H. Evaluation of Anti-Proliferative and Anti-Inflammatory Activities of Pelagia noctiluca Venom in Lipopolysaccharide/Interferon-γ Stimulated RAW264.7 Macrophages. Biomed Pharm. 2016, 84, 1986–1991. [Google Scholar] [CrossRef] [PubMed]
- Moreau, D.; Tomasoni, C.; Jacquot, C.; Kaas, R.; Le Guedes, R.; Cadoret, J.-P.; Muller-Feuga, A.; Kontiza, I.; Vagias, C.; Roussis, V.; et al. Cultivated Microalgae and the Carotenoid Fucoxanthin from Odontella aurita as Potent Anti-Proliferative Agents in Bronchopulmonary and Epithelial Cell Lines. Environ. Toxicol. Pharmacol. 2006, 22, 97–103. [Google Scholar] [CrossRef]
- Rijstenbil, J.W. Effects of UVB Radiation and Salt Stress on Growth, Pigments and Antioxidative Defence of the Marine Diatom Cylindrotheca closterium. Mar. Ecol. Prog. Ser. 2003, 254, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Iio, K.; Okada, Y.; Ishikura, M. Bacterial reverse mutation test and micronucleus test of fucoxanthin oil from microalgae. Shokuhin Eiseigaku Zasshi 2011, 52, 190–193. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, E.T.; Asai, D.J.; Groweiss, A.; Lipshutz, B.H.; Fenical, W.; Jacobs, R.S.; Wilson, L. Mechanism of Action of the Marine Natural Product Stypoldione: Evidence for Reaction with Sulfhydryl Groups. J. Med. Chem. 1986, 29, 1851–1855. [Google Scholar] [CrossRef] [PubMed]
- Saikia, M.; Retnakumari, A.P.; Anwar, S.; Anto, N.P.; Mittal, R.; Shah, S.; Pillai, K.S.; Balachandran, V.S.; Thomas, R.; Anto, R.J. Heteronemin, a Marine Natural Product, Sensitizes Acute Myeloid Leukemia Cells towards Cytarabine Chemotherapy by Regulating Farnesylation of Ras. Oncotarget 2018, 9, 18115–18127. [Google Scholar] [CrossRef] [Green Version]
- Hirata, Y.; Uemura, D. Halichondrins—Antitumor Polyether Macrolides from a Marine Sponge. Pure Appl. Chem. 1986, 58, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Dzhemileva, L.U.; D’yakonov, V.A.; Makarov, A.A.; Andreev, E.N.; Yunusbaeva, M.M.; Dzhemilev, U.M. The first total synthesis of the marine acetylenic alcohol, lembehyne B—A selective inducer of early apoptosis in leukemia cancer cells. Org. Biomol. Chem. 2017, 15, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, D.; Costantini, M.; Coppola, D.; Lauritano, C.; Núñez Pons, L.; Ruocco, N.; di Prisco, G.; Ianora, A.; Verde, C. Biotechnological Applications of Bioactive Peptides From Marine Sources. In Advances in Microbial Physiology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 73, pp. 171–220. ISBN 978-0-12-815190-7. [Google Scholar]
- Bonnard, I.; Bontemps, N.; Lahmy, S.; Banaigs, B.; Combaut, G.; Francisco, C.; Colson, P.; Houssier, C.; Waring, M.J.; Bailly, C. Binding to DNA and Cytotoxic Evaluation of Ascididemin, the Major Alkaloid from the Mediterranean Ascidian Cystodytes dellechiajei. Anticancer Drug Des. 1995, 10, 333–346. [Google Scholar]
- Lindsay, B.S.; Barrows, L.; Copp, B. Structural Requirements for Biological Activity of the Marine Alkaloid Ascididemin. Bioorg. Med. Chem. Lett. 1995. [Google Scholar] [CrossRef]
- Williams, P.G. Panning for Chemical Gold: Marine Bacteria as a Source of New Therapeutics. Trends Biotechnol. 2009, 27, 45–52. [Google Scholar] [CrossRef]
- Kollár, P.; Rajchard, J.; Balounová, Z.; Pazourek, J. Marine Natural Products: Bryostatins in Preclinical and Clinical Studies. Pharm. Biol. 2014, 52, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Sudek, S.; Lopanik, N.B.; Waggoner, L.E.; Hildebrand, M.; Anderson, C.; Liu, H.; Patel, A.; Sherman, D.H.; Haygood, M.G. Identification of the Putative Bryostatin Polyketide Synthase Gene Cluster from “ Candidatus Endobugula Sertula”, the Uncultivated Microbial Symbiont of the Marine Bryozoan Bugula Neritina. J. Nat. Prod. 2007, 70, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Volpe, E.; Sambucci, M.; Battistini, L.; Borsellino, G. Fas–Fas Ligand: Checkpoint of T Cell Functions in Multiple Sclerosis. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somers, C.M.; Kwiecien, J.M.; Quinn, D.J.S. A Marine Fish Diet Reduces Spontaneous Lymphoma in Outbred Swiss-Webster Mice. Leuk. Lymphoma 2005, 46, 1797–1800. [Google Scholar] [CrossRef] [PubMed]
- Betiati, D.D.S.B.; de Oliveira, P.F.; Camargo, C.D.Q.; Nunes, E.A.; Trindade, E.B.S.D.M. Effects of Omega-3 Fatty Acids on Regulatory T Cells in Hematologic Neoplasms. Rev. Bras. Hematol. Hemoter. 2013, 35, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Ahmmed, M.K.; Ahmmed, F.; Tian, H.; Carne, A.; Bekhit, A.E.-D. Marine Omega-3 (n-3) Phospholipids: A Comprehensive Review of Their Properties, Sources, Bioavailability, and Relation to Brain Health. Compr. Rev. Food Sci. Food Saf. 2020, 19, 64–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Lancaster, G.I.; Meikle, P.J. Plasmalogens: A Potential Therapeutic Target for Neurodegenerative and Cardiometabolic Disease. Prog. Lipid. Res. 2019, 74, 186–195. [Google Scholar] [CrossRef]
- Dean, J.M.; Lodhi, I.J. Structural and Functional Roles of Ether Lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef]
- Pédrono, F.; Khan, N.A.; Legrand, A.B. Regulation of Calcium Signalling by 1-O-Alkylglycerols in Human Jurkat T Lymphocytes. Life Sci. 2004, 74, 2793–2801. [Google Scholar] [CrossRef]
- Lauritano, C.; Ferrante, M.I.; Rogato, A. Marine Natural Products from Microalgae: An -Omics Overview. Mar. Drugs 2019, 17, 269. [Google Scholar] [CrossRef] [Green Version]
- Di Dato, V.; Di Costanzo, F.; Barbarinaldi, R.; Perna, A.; Ianora, A.; Romano, G. Unveiling the Presence of Biosynthetic Pathways for Bioactive Compounds in the Thalassiosira rotula Transcriptome. Sci. Rep. 2019, 9, 9893. [Google Scholar] [CrossRef]
- Vingiani, G.M.; Štālberga, D.; De Luca, P.; Ianora, A.; De Luca, D.; Lauritano, C. De Novo Transcriptome of the Non-Saxitoxin Producing Alexandrium Tamutum Reveals New Insights on Harmful Dinoflagellates. Mar. Drugs 2020, 18, 386. [Google Scholar] [CrossRef]
- Elagoz, A.M.; Ambrosino, L.; Lauritano, C. De Novo Transcriptome of the Diatom Cylindrotheca closterium Identifies Genes Involved in the Metabolism of Anti-Inflammatory Compounds. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Lauritano, C.; De Luca, D.; Amoroso, M.; Benfatto, S.; Maestri, S.; Racioppi, C.; Esposito, F.; Ianora, A. New Molecular Insights on the Response of the Green Alga Tetraselmis suecica to Nitrogen Starvation. Sci. Rep. 2019, 9, 3336. [Google Scholar] [CrossRef]
- Vingiani, G.M.; De Luca, P.; Ianora, A.; Dobson, A.D.W.; Lauritano, C. Microalgal Enzymes with Biotechnological Applications. Mar. Drugs 2019, 17, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izadpanah Qeshmi, F.; Homaei, A.; Fernandes, P.; Javadpour, S. Marine Microbial l-Asparaginase: Biochemistry, Molecular Approaches and Applications in Tumor Therapy and in Food Industry. Microbiol. Res. 2018, 208, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Pieters, R.; Hunger, S.P.; Boos, J.; Rizzari, C.; Silverman, L.; Baruchel, A.; Goekbuget, N.; Schrappe, M.; Pui, C.-H. l-Asparaginase Treatment in Acute Lymphoblastic Leukemia: A Focus on Erwinia Asparaginase. Cancer 2011, 117, 238–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahiminezhad, A.; Rasoul-Amini, S.; Ghoshoon, M.B.; Ghasemi, Y. Chlorella Vulgaris, a Novel Microalgal Source for l-Asparaginase Production. Biocatal. Agric. Biotechnol. 2014, 3, 214–217. [Google Scholar] [CrossRef]
- Ali, U.; Naveed, M.; Ullah, A.; Ali, K.; Shah, S.A.; Fahad, S.; Mumtaz, A.S. l-Asparaginase as a Critical Component to Combat Acute Lymphoblastic Leukaemia (ALL): A Novel Approach to Target ALL. Eur. J. Pharmacol. 2016, 771, 199–210. [Google Scholar] [CrossRef]
- Abd El Baky, H.; El baroty, G. Optimization of Growth Conditions for Purification and Production of l-Asparaginase by Spirulina maxima. Evid. Based Complementary Altern. Med. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Prihanto, A.A.; Wakayama, M. Combination of Environmental Stress and Localization of l-Asparaginase in Arthrospira platensis for Production Improvement. 3 Biotech 2014, 4, 647–653. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; De Luca, D.; Ferrarini, A.; Avanzato, C.; Minio, A.; Esposito, F.; Ianora, A. De Novo Transcriptome of the Cosmopolitan Dinoflagellate Amphidinium carterae to Identify Enzymes with Biotechnological Potential. Sci. Rep. 2017, 7, 11701. [Google Scholar] [CrossRef] [Green Version]
- Tangerina, M.M.P.; Furtado, L.C.; Leite, V.M.B.; Bauermeister, A.; Velasco-Alzate, K.; Jimenez, P.C.; Garrido, L.M.; Padilla, G.; Lopes, N.P.; Costa-Lotufo, L.V.; et al. Metabolomic Study of Marine Streptomyces Sp.: Secondary Metabolites and the Production of Potential Anticancer Compounds. PLoS ONE 2020, 15, e0244385. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, J.; Lin, Y. Snake Venoms in Cancer Therapy: Past, Present and Future. Toxins 2018, 10, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, N.L.; Thakur, A.N.; Müller, W.E.G. Marine Natural Products in Drug Discovery. Nat. Product. Radiance 2005, 4, 471–477. [Google Scholar]
- Lebar, M.D.; Heimbegner, J.L.; Baker, B.J. Cold-Water Marine Natural Products. Nat. Prod. Rep. 2007, 24, 774–797. [Google Scholar] [CrossRef]
- Skropeta, D. Deep-Sea Natural Products. Nat. Prod. Rep. 2008, 25, 1131–1166. [Google Scholar] [CrossRef] [Green Version]
- Ruocco, N.; Esposito, R.; Bertolino, M.; Zazo, G.; Sonnessa, M.; Andreani, F.; Coppola, D.; Giordano, D.; Nuzzo, G.; Lauritano, C.; et al. A Metataxonomic Approach Reveals Diversified Bacterial Communities in Antarctic Sponges. Mar. Drugs 2021, 19, 173. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Ianora, A. Grand Challenges in Marine Biotechnology: Overview of Recent EU-Funded Projects. In Grand Challenges in Marine Biotechnology; Rampelotto, P.H., Trincone, A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 425–449. ISBN 978-3-319-69074-2. [Google Scholar]
- Naumann, T.; Çebi, Z.; Podola, B.; Melkonian, M. Growing Microalgae as Aquaculture Feeds on Twin-Layers: A Novel Solid-State Photobioreactor. J. Appl. Phycol. 2013, 25, 1413–1420. [Google Scholar] [CrossRef]
- Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; van der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers, J.M.; et al. Mass Spectral Molecular Networking of Living Microbial Colonies. Proc. Natl. Acad. Sci. USA 2012, 109, E1743–E1752. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.D.; Wu, C.-H.; Moree, W.J.; Lamsa, A.; Medema, M.H.; Zhao, X.; Gavilan, R.G.; Aparicio, M.; Atencio, L.; Jackson, C.; et al. MS/MS Networking Guided Analysis of Molecule and Gene Cluster Families. Proc. Natl. Acad. Sci. USA 2013, 110, E2611–E2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Compound Name | Trademark | Marine Organism | Chemical Class | Molecular Target | Disease Area | Company/Institution |
|---|---|---|---|---|---|---|
| Cytarabine (Ara-C) | Cytosar-U® (1969) * | Sponge | Nucleoside | DNA polymerase | Leukemia | Pfizer |
| Brentuximab vedotin (SGN-35) | Adcetris® (2011) * | Mollusk/cyanobacterium | ADC (MMAE) | CD30 & microtubules | Anaplastic large T-cell systemic malignant lymphoma, Hodgkin’s disease | Seattle Genetics |
| Plitidepsin ** | Aplidin® | Tunicate | Depsipeptide | eEF1A2 | Multiple myeloma, leukemia, lymphoma | Pharmamar |
| Polatuzumab vedotin (DCDS-4501A) | Polivy TM (2019) * | Mollusk/cyanobacterium | ADC (MMAE) | CD79b & microtubules | Non-Hodgkin’s lymphoma, chronic lymphocytic leukemia, lymphoma, B-Cell lymphoma | Genetech/Roche |
| Belantamab mafodotin-blmf | Blenrep TM (2020) * | Mollusk/cyanobacterium | ADC (MMAF) | BCMA | Relapsed/refractory multiple myeloma | GlaxoSmith |
| Compound Name | Marine Organism | Chemical Class | Molecular Target | Disease Area | Company/Institution |
|---|---|---|---|---|---|
| Phase III | |||||
| Marizomib (Salinosporamide A; NPI-0052) | Bacterium | Beta-lactone-gamma lactam | 20S proteasome | Cancer: Non-small cell lung cancer, pancreatic cancer, melanoma, lymphoma, multiple myeloma | Triphase |
| Phase II | |||||
| ABBV-2029 | Mollusk/cyanobacterium | ADC (MMAE) | CD71 | Cancer: Solid tumor, head and neck cancer, non-small cell lung cancer, pancreatic cancer, diffuse large B-cell lymphoma | AbbVie & CytomX Therapeutics |
| Compound | Activity on Cells/Tissues | Organism | Mechanism of Action | Concentration | Reference |
|---|---|---|---|---|---|
| Pestalotiopsone F | L5178Y lymphoma cells | Fungus Pestalatiopsis sp. endophytic to the mangrove plant Rhizophora mucronata | NA | EC50 value of 9 µg/mL | [44] |
| Alteramide | P388 lymphocytic leukemia, L1210 murine lymphoma | Alteromonas sp. (bacteria associated with the pacific sponge Halichondria okadai) | NA | IC50 values of 0.1 and 1.7 µg/mL, respectively | [45] |
| Altemicidin | L1210 murine leukemia | Bacteria Streptomyces sioyaensis SA-1758 | NA | IC50 value of 0.84 µg/mL | [46] |
| Prodigiosin (2-methyl-3-pentyl-6-methoxyprodigiosene) | B-cell chronic lymphocytic leukemia primary cells (n = 32 patients) | Bacterium Serratia marcescens | Apoptosis, induced PARP cleavage, and increased caspase 9 | IC50 value of 116 ± 25 nM. | [47] |
| NA | Acute myeloid leukemia HL-60 cells | Actinobacterium sp. | NA | 0.05 µg/mL | [48] |
| Plinabulin | Multiple myeloma cells, (MM.1S, MM.1R-dex resistant), RPMI-8226, and INA-6 (IL-6-dependent) | Aspergillus sp. | Apoptosis | IC50 value of 8–10 nM | [49] |
| Aqueous and organic extracts | Rat acute myeloid leukemia cells (IPC-81) and human leukemia patient-derived cell line, Molm13 | 40 cyanobacterial isolates | Apoptosis | NA | [50] |
| Extracts | Acute myeloid leukemia HL-60 cells | 41 marine cyanobacterial strains | Apoptosis | 4 mg dry weight/biomass/mL | [51] |
| Colbamide A | HL-60 cells | Cyanobacterium Leptolyngbya sp. | NA | IC50 value of 7.4 nM | [52] |
| Somocystinamide A | CEM (Leukemia) U266 (myeloma) | Cyanobacterium Lyngbya majuscula | Apoptosis via caspase 8 | IC50 values of 14 and 5.8 µM | [53] |
| Cryptophycim 1 | L1210 (murine leukemia cells) | Cyanobacterium Nostoc sp. | Disruption of microtubule assembly | NS | [54] |
| Lagunamidase A,B,C | P388 (murine leukemia cells) | Cyanobacterium Lyngbya majuscule | NA | IC50 values of 6.4, 20.5 and 2.1 nM | [55,56] |
| Microcolin A | P388 cells | Cyanobacterium Lyngbya majuscula | NA | IC50 value of 380 nM | [57] |
| Tolitoxin | L1210 (murine leukemia cells) | Cyanobacterium Seytonema ocellatum | NA | IC50 value of 3.9 nM | [58] |
| Calothrixins A and B | CEM (leukemia) | Cyanobacterium Colothrix sp. | G1 arrest and cell accumulation in S and G2/M phases | IC50 values of 0.20–5.13 µM | [59] |
| NA | IPC-81 rat leukemia cells | Ten microalgae | Classical apoptotic or autophagic features | 1.5 mg/mL | [60] |
| NA | human promyelocytic leukemia cell line (HL-60) | Microalgae Amphidinium operculatum and Ostreopsis ovata | NA | 50 µg/mL | [61] |
| NA | human promyelocytic leukemia cell line (HL-60) | Microalgae Amphidinium carterae and Chlorella ovalis | NA | 25 and 50 µg/mL | [62] |
| NA | human promyelocytic leukemia cell line (HL-60) and human chronic myeloid leukemia cell line (K562) | Various microalgae | Apoptosis (see text for details) | Tested concentrations of 1–500 µg/mL | [63] |
| Apigenin | Human myeloid HL-60 and erythroid TF1 cancer cells. | Microalga Chlorella sp., etc. | Inhibited PI3K/PKB pathway/induced apoptosis in HL-60 cells; Autophagy in TF1 cells | 30–100 µM | [64,65] |
| Amphidinolide H | L1210 murine lymphoma cells and in mice implanted with P388 leukemia | Amphidinium sp. GA3P. | NA | In vitro: IC50 value of 0.5 ng/mL In vivo: IC100 value of 0.2 mg/kg | [66] |
| Carbenolide | K562 leukemia cells | Amphidinium sp. GA3P. | Inhibited DNA topoisomerase I and II; increased lifespan in mice implanted with P388 leukemia cells | IC50 30 ng/mL | [67] |
| 8-acetoxy-6-methyloctanoate (NAMO) | human promyelocytic leukemia cell line (HL-60) | Microalga Phaeodactylum tricornutum | DNA damage, increased apoptotic body formation with the activation of the pro-apoptotic protein Bax, the suppression of the anti-apoptotic protein Bcl-xL, and an increase of caspase 3 and p53 proteins | 50 µg/mL | [68] |
| Hasla-6(17),9,13,23- tetraene | P388 leukemia cells | Microalga Haslea ostreria | NA | IC50 value of 3.3 µg/mL | [69] |
| Monoacylglycerides (MAGs) | U937 (human histiocytic lymphoma) | Microalga S. marinoi | Apoptosis; activation caspase 3/7 | 5–50 µg/mL | [70] |
| Extracts | U937 (human histiocytic lymphoma) | Microalgae D. salina, T. suecica, S. marinoi, C. affinis and T. rotula | NA | 100–500 µg/mL | [70] |
| Compound | Activity on Cells/Tissues | Organism | Mechanism of Action | Concentration | Reference |
|---|---|---|---|---|---|
| NA | K562 and HL-60 leukemia cell lines | Annelida, Bryozoa, Chlorophyta, Chordata, Cnidaria, Cyanophyta, Echinodermata, Mollusca, Phaeophyta, Porifera, Rhodophyta and Tracheophyta | NA | 100 µg/mL | [73] |
| Fucoxanthin and fucoxanthinol | Human primary effusion lymphoma BCBL-1 and TY-1 cells | Brown seaweed Cladosiphon okamuranus Tokida | Cell cycle arrest during the G1 phase and caspase-dependent apoptosis, inhibited the activation of nuclear factor-κB, activator protein-1, and phosphatidylinositol 3-kinase/Akt pathways, and downregulated anti-apoptotic proteins and cell cycle | IC50 values of 2.4–3.3 and 1.1 µM, respectively | [74] |
| Fucoxanthin | Human leukemic HL-60 cells | Brown alga Undaria pinnatifida | NA | 11.3–45.2 μM | [75] |
| Fucoxanthin | Human leukemic HL-60 cells | Brown alga Ishige okamurae | Increase in reactive oxygen species (ROS), cleavage of caspases 3 and 7, and poly-ADP-ribose polymerase (PARP), and decrease in Bcl-xL levels | 12.1 μM | [76] |
| Siphonaxanthin | Human leukemic HL-60 cells | Green macroalga Codium fragile | Decreased Bcl-2 expression, upregulated the expression of GADD45α and DR5, and activated caspase 3 | 20 µM | [77] |
| Fucoxanthin and fucoxanthinol | Adult T-cell leukemia (ATL), human T-cell leukemia virus type 1 (HTLV-1) | Brown algae Undaria pinnatifida | In vitro: Cell cycle arrest during G(1) phase (by reducing the expression of cyclin D1, cyclin D2, CDK4, and CDK6), induced the expression of the apoptosis regulator protein GADD45α, and reduced the expression of Bcl-2, XIAP, cIAP2, and survivin. In addition, they induced the activation of caspases 3, 8, and 9, suppressed IkappaBalpha phosphoryla-tion and JunD expression; In vivo: Suppression of tumor growth | IC50 values of 1.20–4.46 μM and 0.86–1.83 μM, respectively. | [78] |
| Halocynthiaxanthin and fucoxanthinol | Human leukemic HL-60 cells | Sea squirt Halocynthia roretzi | DNA fragmentations and apoptosis (reduction of expression levels of apoptosis-suppressing protein Bcl-2) | 12.5 µM | [79] |
| Fucoidan | Male CDF1 mice inoculated intraperitoneally with a suspension of 105 leukemia L1210 cells | Seaweeds Laminaria angustata var. longissima, Laminaria japonica, Laminaria japonica var. ochotensis, Ecklonia cava and Eisenia bicyclis | NA | 100–200 mg/kg/d for 6 days | [80] |
| Haishengsu | In patients and drug-resistant leukemia cell line (K562/ADM cells) | Seashell Tegillarca granosa L. | Reduction in the expression of P-gp and sorcin | In vivo: 2.4 mg in 250 mL normal saline and given daily over 4 h for 28 days; In vitro: 10–40 µg/mL | [81,82] |
| Orostal and KPN-2001 | Human leukemic HL-60 cells | Sponge Stelletta hiwasaensis | Apoptosis | IC50 value of 1.7 and 2.2 µM | [83] |
| Heteronemin | Chronic myelogenous leukemia K562 cells, human promyelocytic leukemia HL-60 cells, human acute lymphoblastic leukemia Molt4 cells; in vivo mice model inoculated with Molt4 cells | Sponge Hippospongia sp | Apoptosis in leukemia Molt4 cells by inducing an increase in oxidative stress (increase in reactive oxygen species production), mitochondrial dysfunction, upregulation of talin and phosphorylated talin expression | In vitro: EC50 0.41 ± 0.08 mg/mL for K562 cells, 0.16 ± 0.05 mg/mL for HL-60 cells and 0.10 ± 0.004 mg/mL for Molt4 cells; In vivo: 0.31 µg/g | [84] |
| Rhabdastrellic acid A | Human leukemic HL-60 cells | Sponge Rhabdastrella globostellata | Damages the ubiquitin-proteasome pathway (UPP), induced caspase 3 | 5 µM | [85] |
| Smenospongine | Chronic myelogenous leukemia cells K562, HL-60 and human histiocytic lymphoma cells U937 | Sponge Dactylospongia elegans | Induced G1 phase arrest and increased expression of p21 and inhibited phosphorylation of Rb in K562 cells | 5–15 µM | [86] |
| Lembehynes A, B and C | Immortalized line of human T lymphocyte cells Jurkat, and human leukemia cells HL-60 and K562 | Indonesian marine sponge Haliclona sp. | Apoptosis | IC50 values of 2, 2.2, and 3 µM, respectively | [87,88,89] |
| Chujamides A and B | Chronic myelogenous leukemia cells K562 | Sponge Suberites waedoensis | NA | LC50 values of 37 and 55.6 µM, respectively | [90] |
| Compounds 1–8 | Chronic myelogenous leukemia cells K562 | Sponge Coscinoderma sp. | NA | LC50 values of 0.9–5.5 µM | [91] |
| Gombamide A | Chronic myelogenous leukemia cells K562 | Sponge Clathria gombawuiensis | NA | LC50 value of 6.9 µM | [92] |
| ER-076349 and ER08652 | Human leukemic HL-60 cells and U937 histiocytic lymphoma | Analogs of Halichondrin B originally found in a variety of marine sponges | Microtubule destabilized effects | IC50 values of 0.15 and 0.07 nM | [93] |
| Phakellistatins 1, 2 and 3 | P388 murine leukemia cell-line | Sponge Phakellia costata | NA | ED50 values of 7.5, 0.34, and 0.33 µg/mL, respectively | [94,95,96] |
| Phakellistatin 14 | P388 murine leukemia cell line | Sponge Phakellia sp. | NA | ED50 value of 5 µg/mL | [97] |
| Criamides A and B | P388 murine leukemia cell line | Sponge Cymbastela sp. | NA | ED50 value of 0.007 µg/mL | [98] |
| Kapakahine B | P388 murine leukemia cell line | Sponge Cribrochalina olemda | NA | IC50 value of 5 µg/mL | [99] |
| Theonellamides A– F | P388 and L1210 murine leukemia cells | Sponge Theonella sp. | NA | IC50 values of 0.9–5 µg/mL | [100] |
| Polytheonamides A-C | Mouse lymphocytic leukemia L1210 cells | Sponge Theonella swinhoei | NA | EC50 value of <4 ng/mL | [101] |
| Wainunuamide | K562 leukemia cancer cells | Sponge Stylotella aurantium | NA | 18.36 µg/mL | [102] |
| Jaspamide | Human promyelocytic leukemia HL-60 | Sponge Jaspis sp. and Hemiastrella sp. | Apoptosis, neutral endopeptidase (NEP)/CD10 expression on the surface of the apoptotic cells | 100 nM | [103] |
| Ascididemin | HL-60 and the murine P388 and P388CPT5 (sensitive and resistant to camptothecin, respectively) leukemia cell lines | Ascidia Cystodytes dellechiajei/Okinawan tunicate Didemnum species | Double-stranded cleavage of DNA by topoisomerase II, apoptosis mediated by caspase 3 | In vitro: EC50 values of 0.48, 2.4, and 0.05 µM, respectively | [104] |
| Rossinones A and B | P388 murine leukemia cell-line | Aplidium ascidian species | NA | IC50 values of 0.39 and 0.084 µM, respectively | [105] |
| Pseudopterosins, | THP-1 acute monocytic leukemia cells | Soft coral Antillogorgia elisabethae (formerly Pseudopterogorgia elisabethae) | Block of nuclear factor-κB (NF- κB), inhibiting the phosphorylation of p65, inducing a nuclear translocation of the glucocorticoid receptor and reducing the production of the pro-inflammatory cytokines, such as interleukin-6 (IL-6), tumor necrosis factor alpha (TNF-α), and monocyte chemotactic protein 1 (MCP-1) | IC50 value of 24.4 µM | [106] |
| Bryostatin 1 | Patients with low grade non-Hodgkin’s lymphoma or chronic lymphocytic leukemia | Bryozoan Bugula neritina | Upregulation in the co-expression of CD11c/CD22 on CD20+ B cells. | 120 µg/m2 | [107] |
| Colochiroside A | P388 and HL-60 leukemia cell-line | Sea cucumber Cercodemas anceps | Inhibited cell growth | 3.61 ± 0.55 µg/mL | [108] |
| 17-dehydroxyholothurinoside A and griseaside A | Human acute lymphoblastic leukemia cell line Molt-4 | Sea cucumber Holoturia grisea | Cytotoxicity | 0.34 and 0.521 µM | [109] |
| Fuscocinerosides A–C, Pervicoside C, and holoturin A | Human leukemia HL-60 | Sea cucumber Holoturia fuscocinerea | Cytotoxicity | IC50 value of 0.88 µg/mL | [109] |
| Philinopsides A and B | P388 murine leukemia cell-line | Sea cucumber Colochirus quadrangularis | Cytotoxicity | IC50 values of 0.60–3.95 µM | [110] |
| Pentactasides I–III | P388 murine leukemia cell-line | Sea cucumber Colochirus quadrangularis | Cytotoxicity | IC50 0.60 to 3.65 µM | [111] |
| Stichoposide C | Human leukemia HL-60 | Sea cucumber Thelenota anax | Activation of FAS and caspase 8, cleavage of Bid, mitochondrial damage, and caspase 3 activation | Dose-dependent, 0.3–1.5 µmol/L | [112] |
| Raw venom, fractions F1–F3 | Human myelogenous leukemia cells (K562) | Jellyfish Pelagia noctiluca | Cytotoxicity | 5–100 µg/mL | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saide, A.; Damiano, S.; Ciarcia, R.; Lauritano, C. Promising Activities of Marine Natural Products against Hematopoietic Malignancies. Biomedicines 2021, 9, 645. https://doi.org/10.3390/biomedicines9060645
Saide A, Damiano S, Ciarcia R, Lauritano C. Promising Activities of Marine Natural Products against Hematopoietic Malignancies. Biomedicines. 2021; 9(6):645. https://doi.org/10.3390/biomedicines9060645
Chicago/Turabian StyleSaide, Assunta, Sara Damiano, Roberto Ciarcia, and Chiara Lauritano. 2021. "Promising Activities of Marine Natural Products against Hematopoietic Malignancies" Biomedicines 9, no. 6: 645. https://doi.org/10.3390/biomedicines9060645