
Ecklonia cava Extract and Its Derivative Dieckol Promote Vasodilation by Modulating Calcium Signaling and PI3K/AKT/eNOS Pathway in In Vitro and In Vivo Models


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. ECE Preparation and DK Isolation
2.3. Measurement of Cell Viability and NO Production
2.4. Measurement of Cytosolic [Ca2+] Levels
2.5. Western Blot Analysis
2.6. Molecular Docking of DK with AchM3R
2.7. Vascular Response of the Transgenic Zebrafish Tg(flk:EGFP) Model
2.8. Statistical Analysis
3. Results
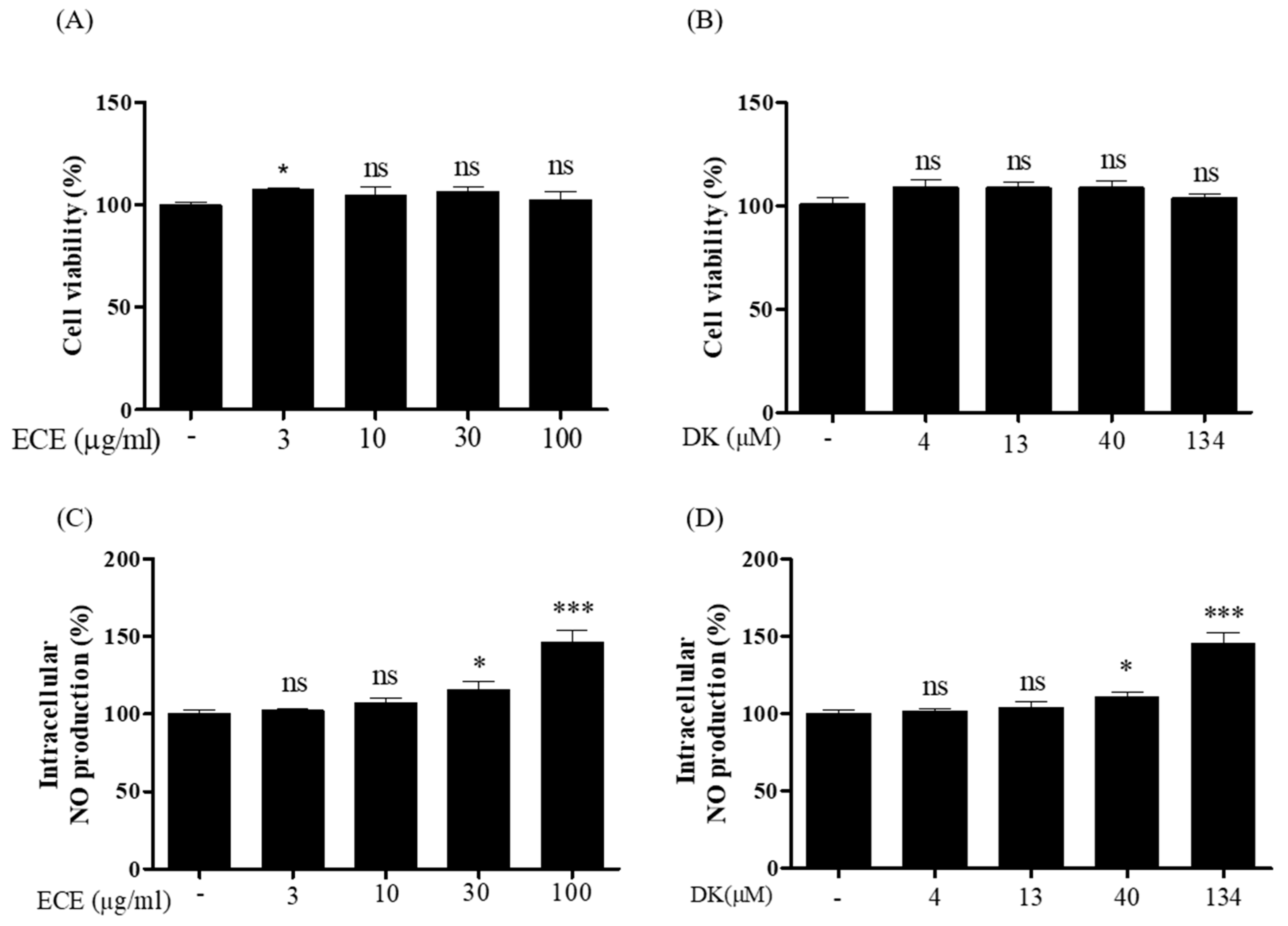
3.1. Effect of ECE and DK on Intracellular NO Production in EA.hy926 Cells
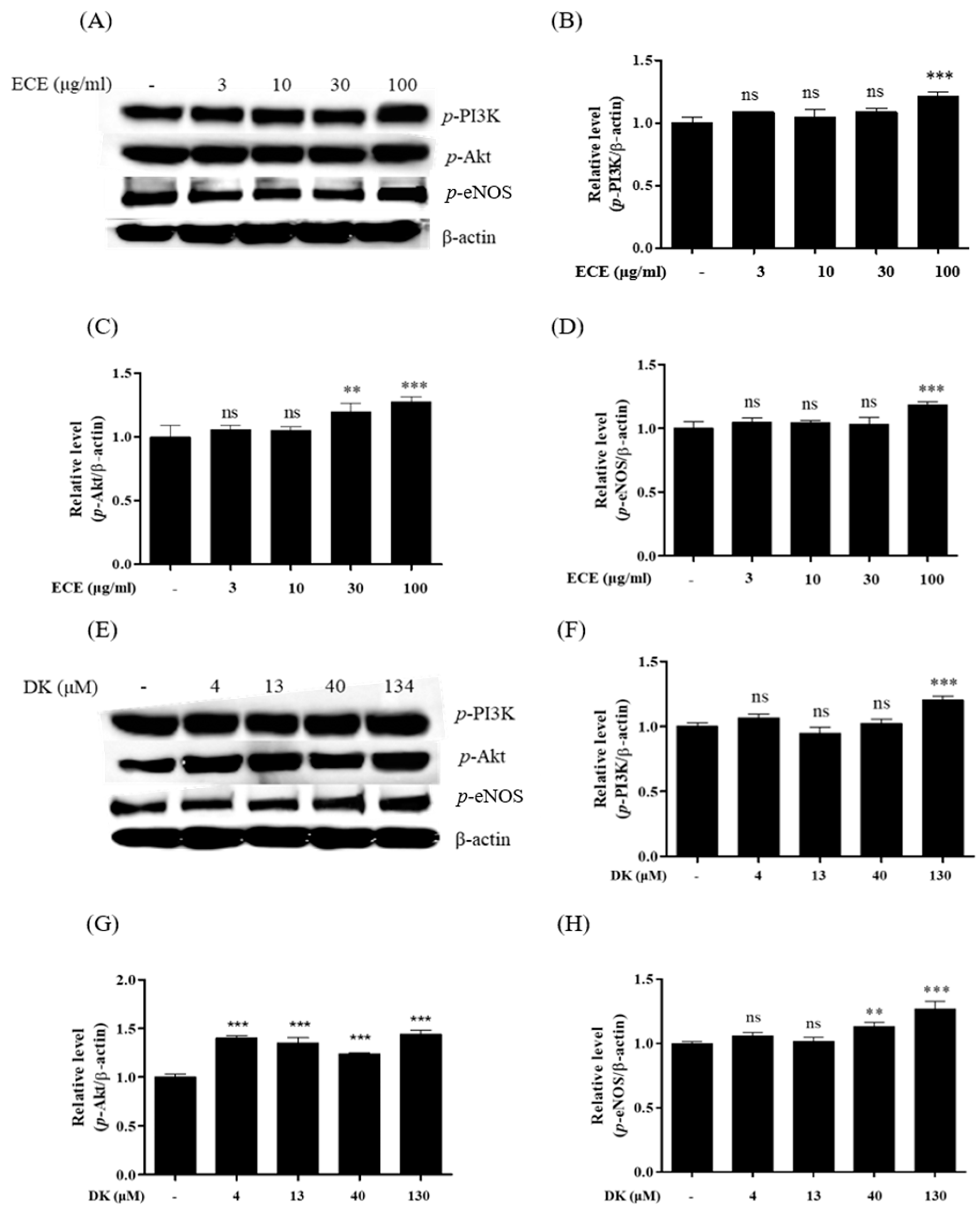
3.2. Effect of ECE and DK on the Phosphorylation of the PI3K/Akt/eNOS Pathway in EA.hy926 Cells
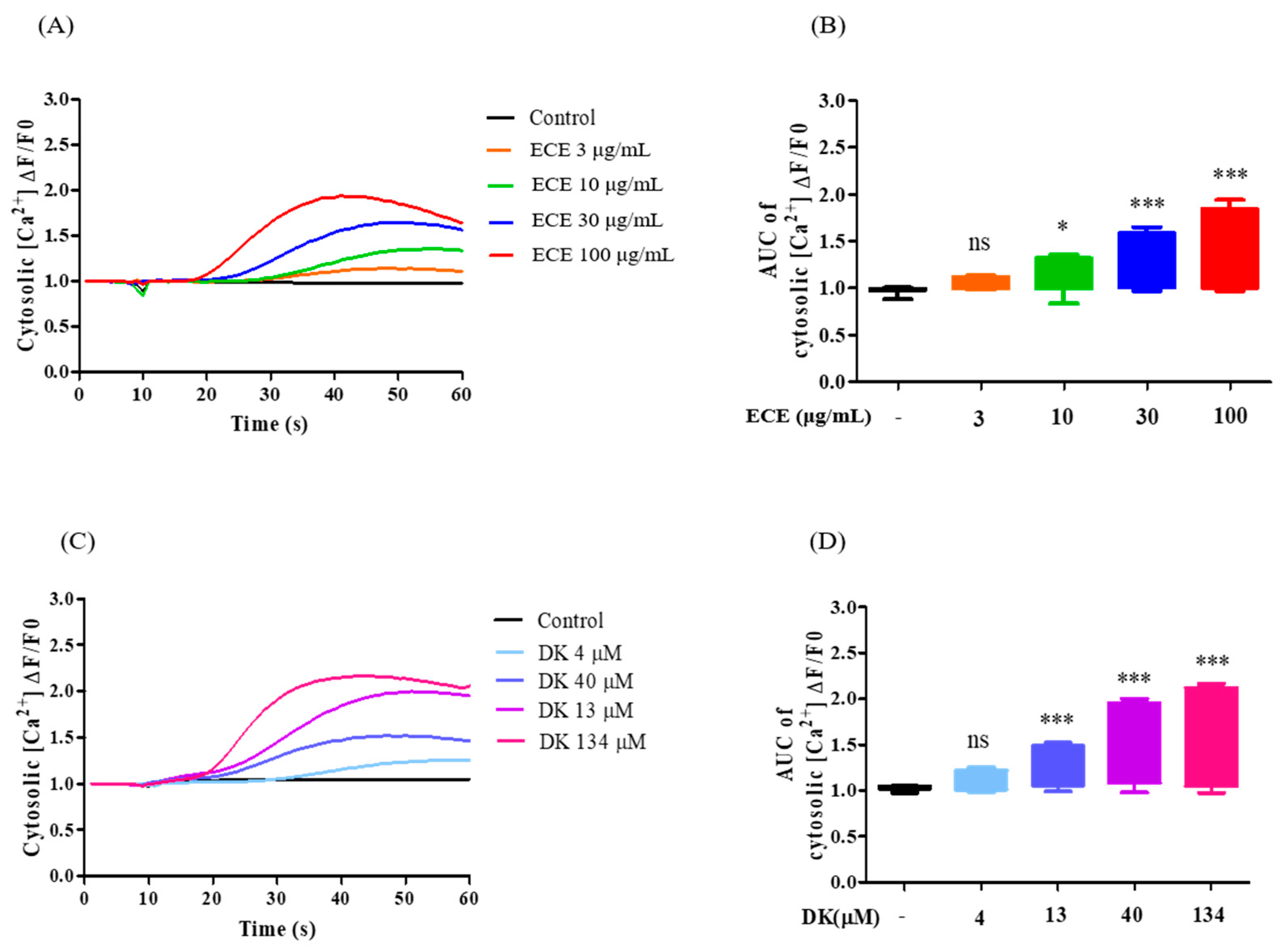
3.3. Effect of ECE and DK on [Ca2+]cytol Levels in EA.hy926 Cells
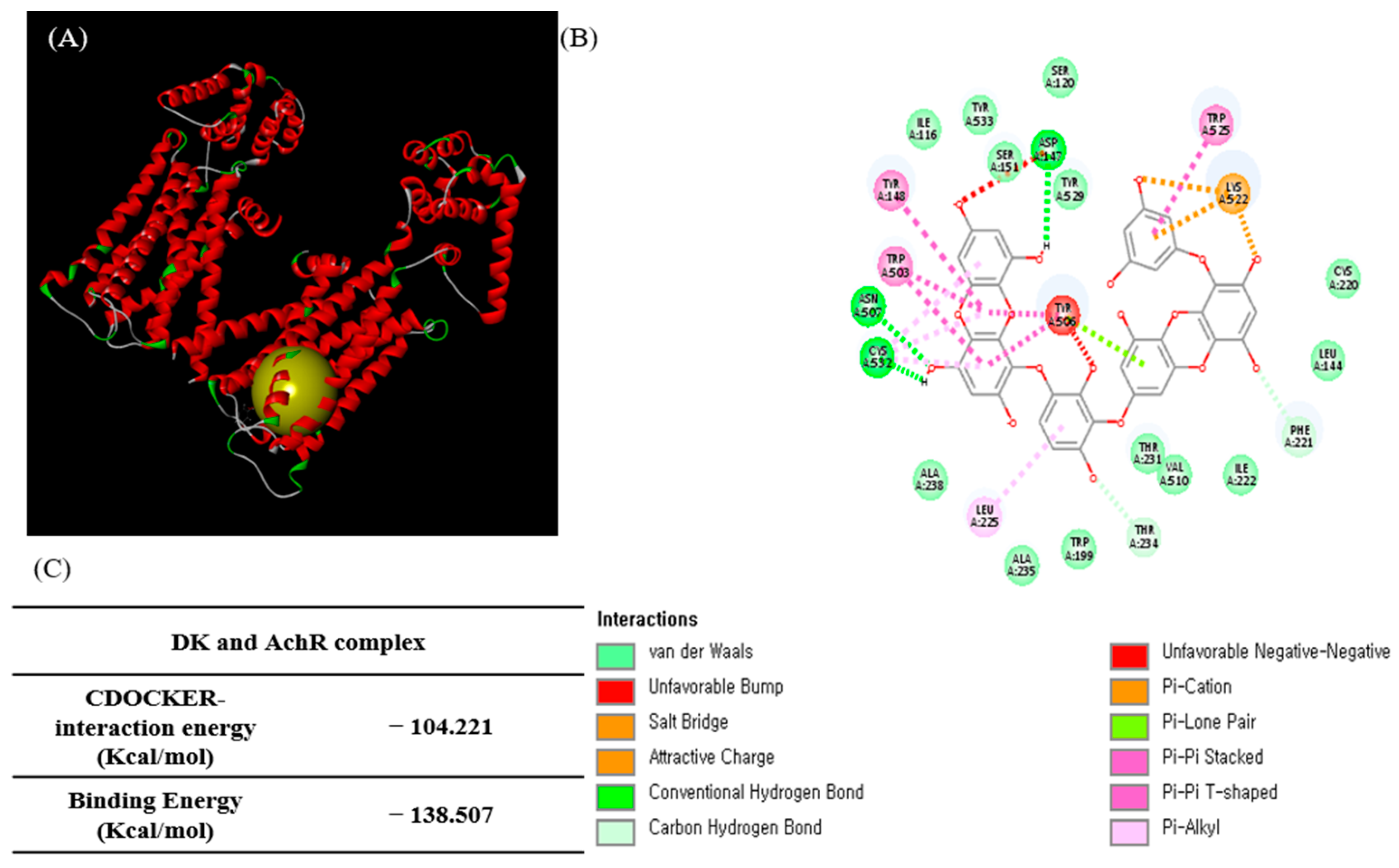
3.4. Molecular Docking Study of DK and AchM3R
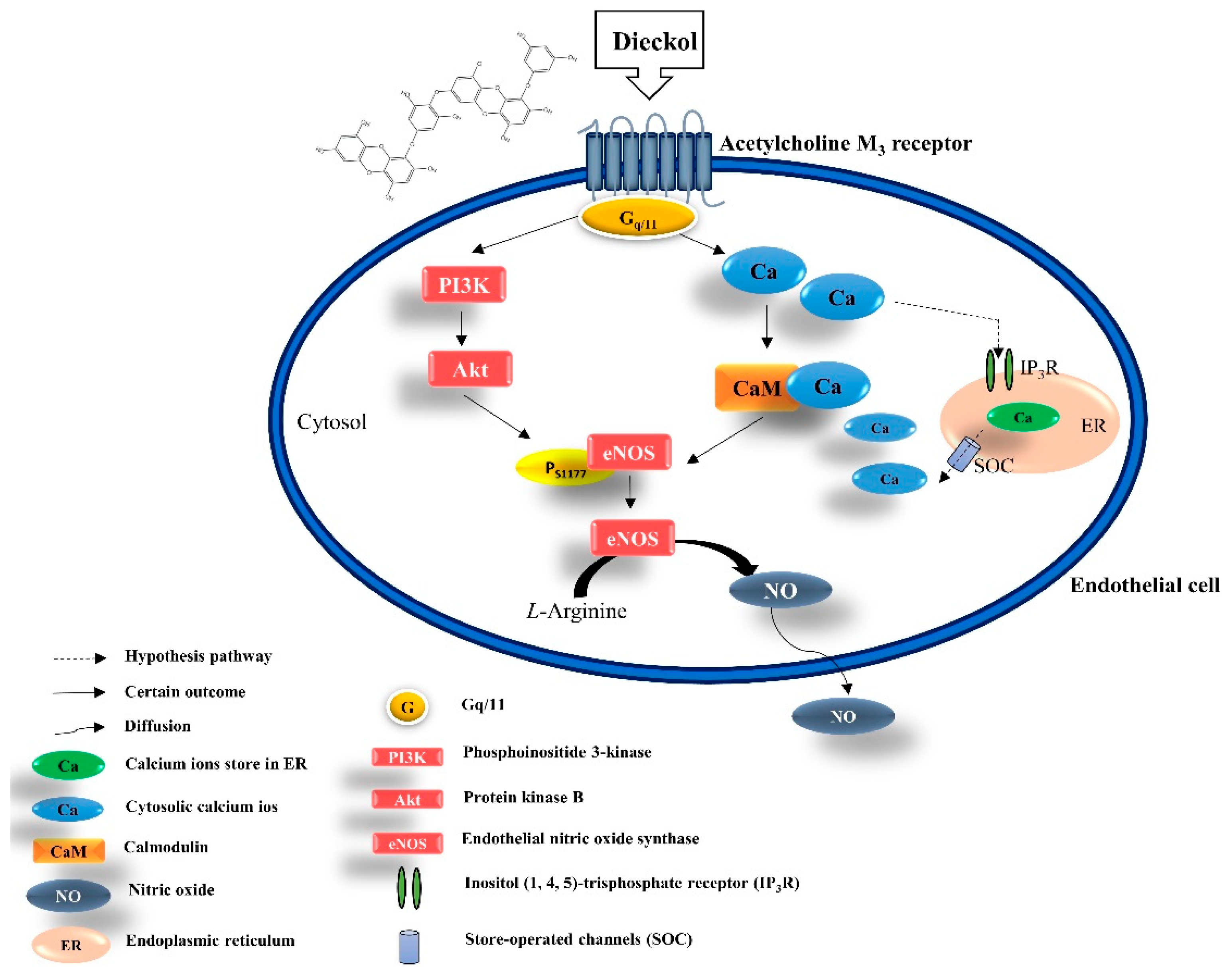
3.5. DK Induced [Ca2+]cytol Transient via AchM3R
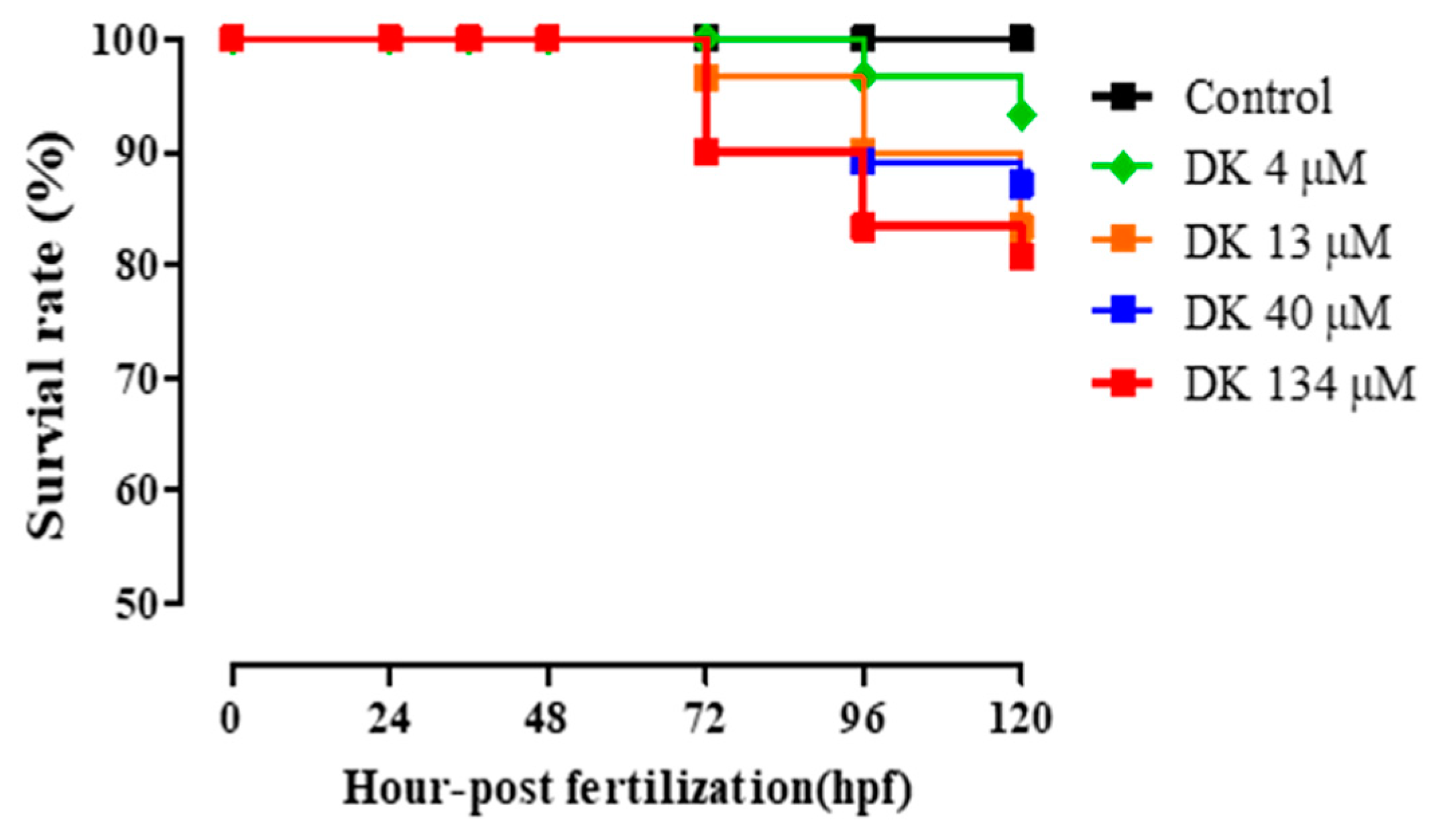
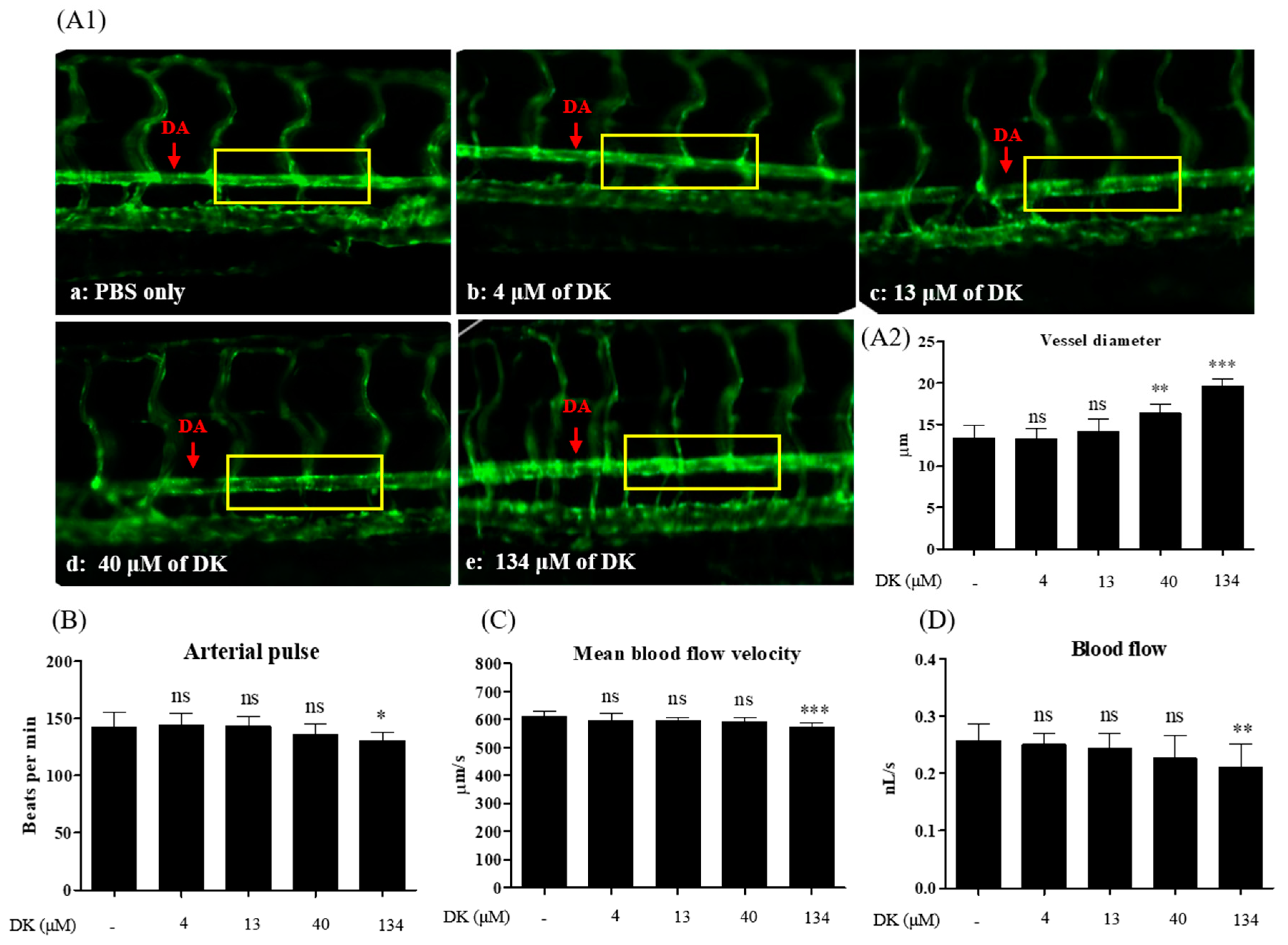
3.6. DK-Promoted Vasodilation in Zebrafish Transgenic (flk: EGFP) Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sauceda, A.E.Q.; Sáyago-Ayerdi, S.G.; Ayala-Zavala, J.F.; Wall-Medrano, A.; De La Rosa, L.A.; González-Aguilar, G.A.; Álvarez-Parrilla, E. Biological Actions of Phenolic Compounds. Fruit Veg. Phytochem. 2017, 2, 125–138. [Google Scholar] [CrossRef]
- Daiber, A.; Xia, N.; Steven, S.; Oelze, M.; Hanf, A.; Kröller-Schön, S.; Münzel, T.; Li, H. New therapeutic implications of endothelial nitric oxide synthase (eNOS) function/dysfunction in car-diovascular disease. Int. J. Mol. Sci. 2019, 20, 187. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, J.; Alloubani, A.; Mari, M.; Alzaatreh, M. Cardiovascular Disease Risk Factors: Hypertension, Diabetes Mellitus and Obesity among Tabuk Citizens in Saudi Arabia. Open Cardiovasc. Med. J. 2018, 12, 41–49. [Google Scholar] [CrossRef]
- Szabo, C. Hydrogen sulfide, an enhancer of vascular nitric oxide signaling: Mechanisms and implications. Am. J. Physiol. Physiol. 2017, 312, C3–C15. [Google Scholar] [CrossRef] [PubMed]
- Gheibi, S.; Jeddi, S.; Kashfi, K.; Ghasemi, A. Regulation of vascular tone homeostasis by NO and H2S: Implications in hypertension. Biochem. Pharmacol. 2018, 149, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Zhao, L.; Jing, R.; Trexler, C.; Wang, H.; Li, Y.; Tang, H.; Huang, F.; Zhang, F.; Fang, X.; et al. Inositol 1,4,5-Trisphosphate Receptors in Endothelial Cells Play an Essential Role in Vasodilation and Blood Pressure Regulation. J. Am. Heart Assoc. 2019, 8, e011704. [Google Scholar] [CrossRef]
- Dalal, P.J.; Muller, W.A.; Sullivan, D.P. Endothelial Cell Calcium Signaling during Barrier Function and Inflammation. Am. J. Pathol. 2020, 190, 535–542. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, T.S.; de Oliveira, L.M.; de Oliveira, L.P.; da Costa, R.M.; de Cássia Tostes, R.; de Castro Georg, R.; Costa, E.A.; de Souza Lobato, N.; Filgueira, F.P.; Ghedini, P.C. Activation of PI3K/Akt pathway mediated by estrogen receptors accounts for estrone-induced vascular activation of cGMP signaling. Vascul. Pharmacol. 2018, 110, 42–48. [Google Scholar] [CrossRef]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular nitric oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulton, D.; Gratton, J.-P.; McCabe, T.J.; Fontana, J.; Fujio, Y.; Walsh, K.; Franke, T.F.; Papapetropoulos, A.; Sessa, W.C. Regulation of endothelium-derived nitric oxide production by the protein kinase Akt. Nat. Cell Biol. 1999, 399, 597–601. [Google Scholar] [CrossRef]
- Son, M.; Oh, S.; Lee, H.S.; Chung, D.-M.; Jang, J.T.; Jeon, Y.-J.; Choi, C.H.; Park, K.Y.; Son, K.H.; Byun, K. Ecklonia Cava Extract Attenuates Endothelial Cell Dysfunction by Modulation of Inflammation and Brown Adipocyte Function in Perivascular Fat Tissue. Nutrients 2019, 11, 2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.-J.; Kim, H.-S.; Je, J.-G.; Wang, L.; Jeon, Y.-J. Antioxidant Activity of Brown Seaweed, Ecklonia maxima, Collected from South Africa. J. Marine Biosci. Biotechnol. 2019, 11, 29–35. [Google Scholar]
- Kang, N.; Ko, S.-C.; Kim, H.-S.; Yang, H.-W.; Ahn, G.; Lee, S.-C.; Lee, T.-G.; Lee, J.-S.; Jeon, Y.-J. Structural Evidence for Antihypertensive Effect of an Antioxidant Peptide Purified from the Edible Marine Animal Styela clava. J. Med. Food 2020, 23, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Oh, S.; Lee, H.S.; Ryu, B.; Jiang, Y.; Jang, J.T.; Jeon, Y.-J.; Byun, K. Pyrogallol-Phloroglucinol-6,6’-Bieckol from Ecklonia cava Improved Blood Circulation in Diet-Induced Obese and Diet-Induced Hypertension Mouse Models. Mar. Drugs 2019, 17, 272. [Google Scholar] [CrossRef] [Green Version]
- Seca, A.M.L.; Pinto, D.C.G.A. Overview on the Antihypertensive and Anti-Obesity Effects of Secondary Metabolites from Seaweeds. Mar. Drugs 2018, 16, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athukorala, Y.; Jeon, Y.-J. Screening for Angiotensin 1-Converting Enzyme Inhibitory Activity of Ecklonia cava. Prev. Nutr. Food Sci. 2005, 10, 134–139. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Mohan, A. Mechanisms of food protein-derived antihypertensive peptides other than ACE inhibition. J. Funct. Foods 2014, 8, 45–52. [Google Scholar] [CrossRef]
- Oh, S.; Son, M.; Lee, H.S.; Kim, H.-S.; Jeon, Y.-J.; Byun, K. Protective Effect of Pyrogallol-Phloroglucinol-6,6-Bieckol from Ecklonia cava on Monocyte-Associated Vascular Dysfunction. Mar. Drugs 2018, 16, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesinghe, W.A.J.P.; Ko, S.-C.; Jeon, Y.-J. Effect of phlorotannins isolated from Ecklonia cava on angiotensin I-converting enzyme (ACE) inhibitory activity. Nutr. Res. Pract. 2011, 5, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-A.; Lee, S.-H.; Lee, J.-H.; Kang, N.; Oh, J.-Y.; Seun-Heui, S.-H.; Ahn, G.; Ko, S.C.; Fernando, S.P.; Kim, S.-Y.; et al. A marine algal polyphenol, dieckol, attenuates blood glucose levels by Akt pathway in alloxan induced hyperglycemia zebrafish model. RSC Adv. 2016, 6, 78570–78575. [Google Scholar] [CrossRef]
- Arutyunyan, I.; Fatkhudinov, T.; Kananykhina, E.; Usman, N.; Elchaninov, A.; Makarov, A.; Bolshakova, G.; Goldshtein, D.; Sukhikh, G. Role of VEGF-A in angiogenesis promoted by umbilical cord-derived mesenchymal stromal/stem cells: In vitro study. Stem Cell Res. Ther. 2016, 7, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Osborne, O.J.; Lin, S.; Ji, Z.; Damoiseux, R.; Wang, Y.; Nel, A.E.; Lin, S. Lanthanide Hydroxide Nanoparticles Induce Angiogenesis via ROS-Sensitive Signaling. Small 2016, 12, 4404–4411. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Song, W.; Xu, S.; Ma, Y.; Wei, B.; Wang, H.; Hua, S. Shenfu Injection Promotes Vasodilation by Enhancing eNOS Activity Through the PI3K/Akt Signaling Pathway In Vitro. Front. Pharmacol. 2020, 11, 121. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Wei, R.-B.; Hong, Q.; Cui, S.-Y.; Chen, X.-M. Grape seed extract enhances eNOS expression and NO production through regulating calcium-mediated AKT phosphorylation in H2O2-treated endothelium. Cell Biol. Int. 2010, 34, 1055–1061. [Google Scholar] [CrossRef]
- Abdulfatai, U.; Uzairu, A.; Uba, S. Quantitative structure-activity relationship and molecular docking studies of a series of quinazolinonyl analogues as inhibitors of gamma amino butyric acid aminotransferase. J. Adv. Res. 2017, 8, 33–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinney, M.C.; Weinstein, B.M. Chapter 4 Using the Zebrafish to Study Vessel Formation. Methods Enzymol. 2008, 444, 65–97. [Google Scholar] [CrossRef]
- Lu, Y.A.; Jiang, Y.; Yang, H.-W.; Hwang, J.; Jeon, Y.-J.; Ryu, B. Diphlorethohydroxycarmalol Isolated from Ishige okamurae Exerts Vasodilatory Effects via Calcium Sig-naling and PI3K/Akt/eNOS Pathway. Int. J. Mol. Sci. 2021, 22, 1610. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.V.; Monzo, K.; Cha, Y.R.; Pan, W.; Weinstein, B.M. Vascular Development in the Zebrafish. Cold Spring Harb. Perspect. Med. 2012, 2, a006684. [Google Scholar] [CrossRef] [Green Version]
- Fernando, K.H.N.; Yang, H.-W.; Jiang, Y.; Jeon, Y.-J.; Ryu, B. Diphlorethohydroxycarmalol Isolated from Ishige okamurae Represses High Glucose-Induced Angiogenesis In Vitro and In Vivo. Mar. Drugs 2018, 16, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minozzo, B.; Fernandes, D.; Beltrame, F. Phenolic Compounds as Arginase Inhibitors: New Insights Regarding Endothelial Dysfunction Treatment. Planta Med. 2018, 84, 277–295. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, J.; Wier, W.; Chen, L.; Blaustein, M.P. NO-induced vasodilation correlates directly with BP in smooth muscle-Na/Ca exchanger-1-engineered mice: Elevated BP does not attenuate endothelial function. Am. J. Physiol. Circ. Physiol. 2021, 320, H221–H237. [Google Scholar] [CrossRef]
- Annuk, M.; Zilmer, M.; Fellström, B. Endothelium-dependent vasodilation and oxidative stress in chronic renal failure: Impact on cardiovascular disease. Kidney Int. 2003, 63, S50–S53. [Google Scholar] [CrossRef] [Green Version]
- Taddei, S.; Virdis, A.; Mattei, P.; Ghiadoni, L.; Sudano, I.; Salvetti, A. Defective l -Arginine–Nitric Oxide Pathway in Offspring of Essential Hypertensive Patients. Circulation 1996, 94, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Pelster, B.; Grillitsch, S.; Schwerte, T. NO as a mediator during the early development of the cardiovascular system in the zebrafish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 142, 215–220. [Google Scholar] [CrossRef]
- Appeldoorn, M.M.; Venema, D.P.; Peters, T.H.F.; Koenen, M.E.; Arts, I.C.W.; Vincken, J.-P.; Gruppen, H.; Keijer, J.; Hollman, P.C.H. Some Phenolic Compounds Increase the Nitric Oxide Level in Endothelial Cells in Vitro. J. Agric. Food Chem. 2009, 57, 7693–7699. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, M.; Wang, M.; Li, X. Flow-mediated vasodilation through mechanosensitive G protein-coupled receptors in endothelial cells. Trends Cardiovasc. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Devika, N.T.; Ali, B.M.J. Analysing calcium dependent and independent regulation of eNOS in endothelium triggered by extracellular signalling events. Mol. BioSyst. 2013, 9, 2653–2664. [Google Scholar] [CrossRef]
- Bełtowski, J.; Jamroz-Wiśniewska, A. Hydrogen Sulfide and Endothelium-Dependent Vasorelaxation. Molecules 2014, 19, 21183–21199. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Mendelsohn, M.E. Synergistic activation of endothelial nitric-oxide synthase (eNOS) by HSP90 and Akt: Calcium-independent eNOS activation involves formation of an HSP90-Akt-CaM-bound eNOS complex. J. Biol. Chem. 2003, 278, 30821–30827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiss, E.H.; Dirsch, V.M. Regulation of eNOS enzyme activity by posttranslational modification. Curr. Pharm. Des. 2014, 20, 3503–3513. [Google Scholar] [CrossRef] [Green Version]
- Martinotti, S.; Patrone, M.; Balbo, V.; Mazzucco, L.; Ranzato, E. Endothelial Response Boosted by Platelet Lysate: The Involvement of Calcium Toolkit. Int. J. Mol. Sci. 2020, 21, 808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Caliman, A.D.; McCammon, J.A. Allosteric Effects of Sodium Ion Binding on Activation of the M3 Muscarinic G-Protein-Coupled Receptor. Biophys. J. 2015, 108, 1796–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Archundia, M.; Cordomí, A.; Garriga, P.; Perez, J.J. Molecular Modeling of the M3 Acetylcholine Muscarinic Receptor and Its Binding Site. J. Biomed. Biotechnol. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Weston-Green, K.; Huang, X.-F.; Lian, J.; Deng, C. Effects of olanzapine on muscarinic M3 receptor binding density in the brain relates to weight gain, plasma insulin and metabolic hormone levels. Eur. Neuropsychopharmacol. 2012, 22, 364–373. [Google Scholar] [CrossRef]
- Kato, M.; Komamura, K.; Kitakaze, M. Tiotropium, a Novel Muscarinic M3 Receptor Antagonist, Improved Symptoms of Chronic Obstructive Pulmonary Disease Complicated by Chronic Heart Failure. Circ. J. 2006, 70, 1658–1660. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.; Vanpatten, S.; Lakshminrusimha, S.; Patel, H.; Coleman, T.R.; Al-Abed, Y. Effects of novel muscarinic M3 receptor ligand C1213 in pulmonary arterial hypertension models. Physiol. Rep. 2016, 4, e13069. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Lindgren, A.; Wu, X.; Liu, N.-A.; Lin, S. High-Throughput Screening for Bioactive Molecules Using Primary Cell Culture of Transgenic Zebrafish Embryos. Cell Rep. 2012, 2, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahrami, N.; Childs, S.J. Development of vascular regulation in the zebrafish embryo. Development 2020, 147, dev183061. [Google Scholar] [CrossRef] [PubMed]
- Margiotta-Casaluci, L.; Owen, S.F.; Rand-Weaver, M.; Winter, M.J. Testing the Translational Power of the Zebrafish: An Interspecies Analysis of Responses to Cardiovascular Drugs. Front. Pharmacol. 2019, 10, 893. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.-A.; Je, J.-G.; Hwang, J.; Jeon, Y.-J.; Ryu, B. Ecklonia cava Extract and Its Derivative Dieckol Promote Vasodilation by Modulating Calcium Signaling and PI3K/AKT/eNOS Pathway in In Vitro and In Vivo Models. Biomedicines 2021, 9, 438. https://doi.org/10.3390/biomedicines9040438
Lu Y-A, Je J-G, Hwang J, Jeon Y-J, Ryu B. Ecklonia cava Extract and Its Derivative Dieckol Promote Vasodilation by Modulating Calcium Signaling and PI3K/AKT/eNOS Pathway in In Vitro and In Vivo Models. Biomedicines. 2021; 9(4):438. https://doi.org/10.3390/biomedicines9040438
Chicago/Turabian StyleLu, Yu-An, Jun-Geon Je, Jin Hwang, You-Jin Jeon, and BoMi Ryu. 2021. "Ecklonia cava Extract and Its Derivative Dieckol Promote Vasodilation by Modulating Calcium Signaling and PI3K/AKT/eNOS Pathway in In Vitro and In Vivo Models" Biomedicines 9, no. 4: 438. https://doi.org/10.3390/biomedicines9040438