
Potential of Marine Terpenoids against SARS-CoV-2: An In Silico Drug Development Approach

 ,
,  , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Ligand and Receptor Preparation
2.2. SARS-CoV-2: Genomic−Diversity and Phylogenetic Tree Analyses
2.3. Virtual Screening by Molecular Docking Study
2.4. Physicochemical−Lipinski’s Rule of Five Profile Analyses
2.5. Toxicity and Pharmacokinetic Profile Prediction
2.6. Overall Drug Likeness and SAR Analyses
3. Results and Discussion
3.1. Virtual Screening by Molecular Docking Study
3.2. Physicochemical−Lipinski’s Rule of Five Profile Analyses
3.3. Toxicity and Pharmacokinetic Profile Prediction
3.4. Overall Drug Likeness and SAR Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO–Coronavirus Disease (COVID-19) Dashboard. Available online: https://covid19.who.int/table (accessed on 2 October 2021).
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Chellapandi, P.; Saranya, S. Genomics insights of SARS-CoV-2 (COVID-19) into target-based drug discovery. Med. Chem. Res. 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Morse, J.S.; Lalonde, T.; Xu, S.; Liu, W.R. Learning from the past: Possible urgent prevention and treatment options for severe acute respiratory infections caused by 2019-nCoV. ChemBioChem 2020, 21, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Zumla, A.; Chan, J.F.; Azhar, E.I.; Hui, D.S.; Yuen, K.Y. Coronaviruses—drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef] [Green Version]
- Bimonte, S.; Crispo, A.; Amore, A.; Celentano, E.; Cuomo, A.; Cascella, M. Potential antiviral drugs for SARS-CoV-2 treatment: Preclinical findings and ongoing clinical research. In Vivo 2020, 34, 1597–1602. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.M.; Monogue, M.L.; Jodlowski, T.Z.; Cutrell, J.B. Pharmacologic treatments for coronavirus disease-2019 (COVID-19): A review. JAMA 2020, 323, 1824–1836. [Google Scholar] [CrossRef]
- Martinez, M.A. Clinical trials of repurposed antivirals for SARS-CoV-2. Antimicrob. Agent Chemother. 2020, 64, e01101-120. [Google Scholar] [CrossRef]
- Stadler, K.; Masignani, V.; Eickmann, M.; Becker, S.; Abrignani, S.; Klenk, H.D.; Rappuoli, R. SARS−−beginning to understand a new virus. Nat. Rev. Microbiol. 2003, 1, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor−binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Elfiky, A.A. Anti-HCV, nucleotide inhibitors, repurposing against COVID-19. Life Sci. 2020, 248, 117477. [Google Scholar] [CrossRef] [PubMed]
- Gil, C.; Ginex, T.; Maestro, I.; Nozal, V.; Barrado-Gil, L.; Cuesta-Geijo, M.Á.; Urquiza, J.; Ramirez, D.; Alonso, C.; Campillo, N.E.; et al. COVID-19: Drug targets and potential treatments. J. Med. Chem. 2020, 63, 12359–12386. [Google Scholar] [CrossRef] [PubMed]
- Gentile, D.; Patamia, V.; Scala, A.; Sciortino, M.T.; Piperno, A.; Rescifina, A. Putative inhibitors of SARS-CoV-2 main protease from alibrary of marine natural products: A virtual screening and molecular modelings. Mar. Drugs 2020, 18, 225. [Google Scholar] [CrossRef] [Green Version]
- Avorn, J. The $2.6 billion pill−methodologic and policy considerations. N. Engl. J. Med. 2015, 372, 1877–1879. [Google Scholar] [CrossRef]
- Greene, J.A.; Loscalzo, J. Putting the patient back together—Social medicine, network medicine, and the limits of reductionism. N. Engl. J. Med. 2017, 377, 2493–2499. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Hou, Y.; Shen, J.; Huang, Y.; Martin, W.; Cheng, F. Network-based drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2. Cell Discov. 2020, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochem. Biophys. Acta. 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria. Biomed. Pharmacother. 2017, 90, 760–776. [Google Scholar] [CrossRef]
- Li, F.; Wang, Y.; Li, D.; Chen, Y.Q. Dou P Are we seeing a resurgence in the use of natural products for new drug discovery? Expert Opin. Drug Discov. 2019, 14, 417–420. [Google Scholar] [CrossRef]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.S.; Paidesetty, S.K.; Dehury, B.; Sahoo, J.; Vedithi, S.C.; Mahapatra, N.; Hussain, T.; Padhy, R.N. Molecular docking and simulation study for synthesis of alternative dapsone derivative as a newer antileprosy drug in multidrug therapy. J. Cell Biochem. 2018, 119, 9838–9852. [Google Scholar] [CrossRef]
- Cheng, F. In silico oncology drug repositioning and polypharmacology. Methods Mol. Biol. 2019, 1878, 243–261. [Google Scholar] [PubMed]
- Prajapat, M.; Sarma, P.; Shekhar, N.; Avti, P.; Sinha, S.; Kaur, H.; Kumar, S.; Bhattacharyya, A.; Kumar, H.; Bansal, S.; et al. Drug targets for corona virus: A systematic review. Indian J. Pharmacol. 2020, 52, 56–65. [Google Scholar]
- Swain, S.S.; Paidesetty, S.K.; Dehury, B.; Das, M.; Chaitanya, S.V.; Padhy, R.N. Computer−aided synthesis of dapsone-phytochemical conjugates against dapsone-resistant Mycobacterium leprae. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, H.S.; Leao-Ferreira, L.R.; Moussatche, N.; Teixeira, V.L.; Cavalcanti, D.N.; Costa, L.J.; Diaz, R.; Frugulhetti, I.C.P.P. Antiviral activity of diterpenes isolated from the Brazilian marine alga Dictyotamenstrualisagainst human immunodeficiency virus type 1 (HIV−1). Antivir. Res. 2004, 64, 69–76. [Google Scholar]
- Wang, S.K.; Hsieh, M.K.; Duh, C.Y. New diterpenoids from soft coral Sarcophyton ehrenbergi. Mar. Drugs 2013, 11, 4318–4327. [Google Scholar] [CrossRef] [Green Version]
- Ellithey, M.S.; Lall, N.; Hussein, A.A.; Meyer, D. Cytotoxic, cytostatic and HIV−1 PR inhibitory activities of the soft coral Litophyton arboreum. Mar. Drugs 2013, 11, 4917–4936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucknall, R.A.; Moores, H.; Simms, R.; Hesp, B. Antiviral effects of aphidicolin, a new antibiotic produced by Cephalosporiumaphidicola. Antimicrob. Agents Chemother. 1973, 4, 294–298. [Google Scholar] [CrossRef] [Green Version]
- Mendes, G.; Soares, A.R.; Sigiliano, L.; Machado, F.; Kaiser, C.; Romeiro, N.; Gestinari, L.; Santos, N.; Romanos, M.T.V. In vitro anti−HMPV activity of meroditerpenoids from marine alga Stypopodiumzonale (dictyotales). Molecules 2011, 16, 8437–8450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarin, P.S.; Sun, D.; Thornton, A.; Müller, W.E.G. Inhibition of replication of the etiologic agent of acquired immune deficiency syndrome (human T−lymphotropic retrovirus! lymphadenopathy−associated virus) by avarol and avarone. J. Nat. Cancer Inst. 1987, 78, 663–666. [Google Scholar] [PubMed]
- Li, Y.; Ye, D.Z.; Chen, X.L.; Lu, X.H.; Shao, Z.Z.; Zhang, H.; Che, Y.S. Breviane spiroditerpenoids from an extreme-tolerant Penicillium sp. Isolated from a deep-sea sediment sample. J. Nat. Prod. 2009, 72, 912–916. [Google Scholar] [CrossRef]
- Coval, S.J.; Cross, S.S.; Bemardinelli, G.; Jefford, C.W. Brianthein V, a New cytotoxic and antiviral diterpene isolated from Briareum asbestinum. J. Nat. Prod. 1988, 51, 981–984. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Chuang, C.T.; Wen, Z.H.; Wang, S.K.; Chiou, S.F.; Hsu, C.H.; Dai, C.F.; Duh, C.Y. Bioactive norditerpenoids from the soft coral Sinularia gyrosa. Bioorg. Med. Chem. 2010, 18, 3379–3386. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Huang, K.J.; Wang, S.K.; Duh, C.Y. Capilloquinol: A novel farnesyl quinol from the Dongsha atoll soft coral Sinularia capillosa. Mar. Drugs 2011, 9, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Gul, W.; Hammond, N.L.; Yousaf, M.; Peng, J.; Holley, A.; Hamann, M.T. Chemical transformation and biological studies of marine sesquiterpene (S)-(+)-curcuphenol and its analogs. Biochim. Biophys. Acta 2007, 1770, 1513–1519. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.N.; Walsh, K.; Weedman, V.; Bergthold, J.D.; Lynch, J.; Lieu, K.L.; Braude, I.A.; Kelly, M.; Hamann, M.T. The new bioactive diterpenes cyanthiwigins E-AA from the Jamaican sponge Myrmekioderma styx. Tetrahedron 2002, 58, 7809–7819. [Google Scholar] [CrossRef]
- Chill, L.; Rudi, A.; Aknin, M.; Loya, S.; Hizi, A.; Kashman, Y. New sesterterpenes from Madagascan Lendenfeldia sponges. Tetrahedron 2004, 60, 10619–10626. [Google Scholar] [CrossRef]
- Pardo-Vargas, A.; Oliveira, I.B.; Stephens, P.R.S.; Cirne-Santos, C.C.; de Palmer Paixão, I.C.N.; Ramos, F.A.; Jimenez, C.; Rodriguez, J.; Resende, J.A.A.C.; Teixeira, V.L.; et al. Dolabelladienols A-C, new diterpenes isolated from Brazilian brown alga Dictyota pfaffii. Mar. Drugs 2014, 12, 4247–4259. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.K.; Hsieh, M.K.; Duh, C.Y. Three new cembranoids from the Taiwanese soft coral Sarcophyton ehrenbergi. Mar. Drugs 2012, 10, 1433–1444. [Google Scholar] [CrossRef] [Green Version]
- Kohmoto, S.; McConnell, O.J.; Wright, A.E.; Cross, S.S. Isospongiadiol, a cytotoxic and antiviral diterpene from a caribbean deep−water marine sponge Spongia sp. Chem. Lett. 1987, 16, 1687–1690. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Wang, S.K.; Duh, C.Y. Secocrassumol, a seco-cembranoid from the Dongsha Atoll soft coral Lobophytum crassum. Mar. Drugs 2014, 12, 6028–6037. [Google Scholar] [CrossRef] [Green Version]
- Koehn, F.E.; Gunasekera, S.P.; Niel, D.N.; Cross, S.S. Halitunal, an Unusual Diterpene Aldehyde from The Marine Alga Halimeda tuna. Tetrahedron Lett. 1991, 32, 169–172. [Google Scholar] [CrossRef]
- Tanaka, J.I.; Ogawa, N.; Liang, J.; Higa, T.; Garcia-Gravalos, D. Natural marine antiviral products. Tetrahedron 1993, 49, 811–822. [Google Scholar] [CrossRef]
- Du, L.; Shen, L.L.; Yu, Z.G.; Chen, J.; Guo, Y.W.; Tang, Y.; Shen, X.; Jiang, H.L. Hyrtiosal, from the marine sponge Hyrtios erectus, inhibits HIV-1 integrase binding to viral DNA by a new inhibitor binding site. Chem. Med. Chem. 2008, 3, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Wang, X.M. A new sesquiterpenoid hydroquinone from the marine sponge Dysidea arenaria. Molecules 2008, 13, 1275–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.B.; Zink, D.; Polishook, J.; Valentino, D.; Shafiee, A.; Silverman, K.C.; Felock, P.; Vilella, D.; Hazuda, D.; Lingham, R.B. Structure and absolute stereochemistry of HIV-1 integrase inhibitor integric acid. A novel eremophilane sesquiterpenoid produced by a Xylaria sp. Tetrahedron Lett. 1999, 40, 8775–8779. [Google Scholar] [CrossRef]
- Maier, M.S.; Roccatagliata, A.J.; Kuriss, A.; Chludil, H.; Seldes, A.M.; Pujol, C.A.; Damonte, E.B. Two new cytotoxic and virucidal trisulfated triterpene glycosides from the Antarctic Sea cucumber Staurocucumis liouvillei. J. Nat. Prod. 2001, 64, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.A.; Gustafson, K.R.; Boyd, M.R. HIV-inhibitory cembrane derivatives from a Philippines collection of the soft coral Lobophytum species. J. Nat. Prod. 2000, 63, 531–533. [Google Scholar] [CrossRef]
- Tanaka, J.; Higa, T.; Suwanborirux, K.; Kopol, U.; Bernardinelli, G.; Jefford, C.W. Bioactive norsesterterpene 1,2-Dioxanes from a Thia Sponge, Mycale sp. J. Org. Chem. 1993, 58, 2999–3002. [Google Scholar] [CrossRef]
- Loy, S.; Bakhanashvili, M.; Kashman, Y.; Hizi, A. Peyssonols A and B, two novel inhibitors of the reverse transcriptases of human immunodeficiency virus types 1 and 2. Arch. Biochem. Biophys. 1995, 316, 789–796. [Google Scholar] [CrossRef]
- Rodriguez, I.L.; Shi, Y.P.; Garcia, O.J.; Rodriguez, A.D.; Mayer, A.M.; Sanchez, J.A.; Ortega−Barrio, E.; Gonzalez, J. New pseudopterosin and seco-pseudopterosin diterpene glycosides from two colombian isolates of Pseudopterogorgia elisabethae and their diverse biological activities. J. Nat. Prod. 2004, 67, 1672–1680. [Google Scholar] [CrossRef] [PubMed]
- Zjawiony, J.K.; Bartyzel, P.; Hamann, M.T. Chemistry of puupehenone: 1,6-conjugate Addition to Its quinone−methide system. J. Nat. Prod. 1998, 61, 1502. [Google Scholar] [CrossRef] [PubMed]
- Kashman, Y.; Hirsch, S.; Koehn, F.; Cross, S.S. Reiswigins A and B, novel antiviral diterpenes from a deep-water sponge. Tetrahedron Lett. 1987, 28, 5461–5464. [Google Scholar] [CrossRef]
- Cooper, G.J.; Davies−Coleman, M.T. Sesquiterpene hydroquinones from the South Africa soft coral Alcyonium fauri. Tetrahedron Lett. 1995, 36, 3265–3268. [Google Scholar]
- Tseng, Y.J.; Wang, S.K.; Duh, C.Y. Secosteroids and norcembranoids from the soft coral Sinularia nanolobata. Mar. Drugs 2013, 11, 3288–3296. [Google Scholar] [CrossRef] [PubMed]
- Groweiss, A.; Look, S.A.; Fenical, W. Solenolides, new antiinflammatory and antiviral diterpenoids from a marine octocoral of the genus Solenopodium. J. Org. Chem. 1988, 53, 2401–2406. [Google Scholar] [CrossRef]
- Qin, C.; Lin, X.; Lu, X.; Wan, J.; Zhou, X.; Liao, S.; Tu, Z.; Xu, S.; Liu, Y. Sesquiterpenoids and xanthones derivatives produced by sponge-derived fungus Stachybotry sp. HH1 ZSDS1F1−2. J. Antibiot. 2015, 68, 121–125. [Google Scholar] [CrossRef]
- Minagawa, K.; Kouzuki, S.; Yoshimoto, J.; Kawamura, Y.; Tani, H.; Iwata, T.; Terui, Y.; Yagi, S.; Hattori, N.; Fujiwara, T.; et al. Stachyflin and acetylstachyflin, novel anti−influenza A virus substances, produced by Stachybotrys sp. RF-7260. I. Isolation, structure elucidation and biological activities. J. Antibiot. 2002, 55, 155–164. [Google Scholar] [CrossRef]
- Wright, A.E.; Rueth, S.A.; Cross, S.S. An antiviral sesquiterpene hydroquinone from the marine sponge Strongylophora hartmani. Nat. Prod. 1991, 54, 1108–1111. [Google Scholar] [CrossRef]
- Sakemi, S.; Higa, T.; Jefford, C.W.; Bernardinelli, G. Venustatriol. A new, antiviral, triterpene tetracyclic ether from Laurencia venusta. Tetrahedron Lett. 1986, 27, 4287–4290. [Google Scholar] [CrossRef]
- Urones, J.G.; Araujo, M.E.M.; Brito, P.F.M.S.; Basabe, P.; Marcos, I.S.; Moro, R.F.; Lithgo, A.M.; Pineda, J. Meroterpenes from cystoseirausneoides II. Phytochemistry 1992, 31, 2105–2109. [Google Scholar] [CrossRef]
- Ding, L.; Munich, J.; Goerls, H.; Maier, A.; Fiebig, H.H.; Lin, W.H.; Hertweck, C. Xiamycin, a pentacyclic indolosesquiterpene with selective anti-HIV activity from a bacterial mangrove endophyte. Bioorg. Med. Chem. Lett. 2010, 20, 6685–6687. [Google Scholar] [CrossRef]
- Abrantes, J.L.; Barbosa, J.; Cavalcanti, D.; Pereira, R.C.; Frederico Fontes, C.L.; Teixeira, V.L.; Moreno Souza, T.L.; Paixão, I.C. The effects of the diterpenes isolated from the Brazilian brown algae Dictyota pfaffii and Dictyota menstrualis against the herpes simplex type-1 replicative cycle. Planta Med. 2010, 76, 339–344. [Google Scholar] [CrossRef] [Green Version]
- El−Hachem, N.; Haibe−Kains, B.; Khalil, A.; Kobeissy, F.H.; Nemer, G. AutoDock and AutoDock tools for protein−ligand docking: Beta-site amyloid precursor protein cleaving enzyme 1(BACE1) as a case study. Methods Mol. Biol. 2017, 1598, 391–403. [Google Scholar]
- Ma, C.; Sacco, M.D.; Hurst, B.; Townsend, J.A.; Hu, Y.; Szeto, T.; Zhang, X.; Tarbet, B.; Marty, M.T.; Chen, Y.; et al. Boceprevir, GC-376, and calpain inhibitors II, XII inhibit SARS-CoV-2 viral replication by targeting the viral main protease. Cell Res. 2020, 30, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Peng, R.; Yuan, B.; Zhao, J.; Wang, M.; Wang, X.; Wang, Q.; Sun, Y.; Fan, Z.; Qi, J.; et al. Structural and biochemical characterization of the nsp12-nsp7-nsp8 core polymerase complex from SARS-CoV-2. Cell Rep. 2020, 31, 107774. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A. Drug-like properties and the causes of poor solubility and poor permeability. J. Pharmacol. Toxicol. Methods 2000, 44, 235–249. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara−Bell, J.; Lu, Y. Marine compounds and their antiviral activities. Antiviral. Res. 2010, 86, 231–240. [Google Scholar] [CrossRef]
- Putta, S.; Yarla, N.S.; Kilari, E.K.; Surekha, C.; Aliev, G.; Divakara, M.B.; Santosh, M.S.; Ramu, R.; Zameer, F.; Mn, N.P.; et al. Therapeutic potentials of triterpenes in diabetes and its associated complications. Curr. Top. Med. Chem. 2016, 16, 2532–2542. [Google Scholar] [CrossRef]
- Cox−Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and medicinal uses of terpenes. Med. Plants 2019, 333–359. [Google Scholar]
- Saeed, A.F.U.H.; Su, J.; Ouyang, S. Marine−derived drugs: Recent advances in cancer therapy and immune signaling. Biomed. Pharmacother. 2021, 134, 111091. [Google Scholar] [CrossRef]
- Nogueira, J.O.E.; Campolina, G.A.; Batista, L.R.; Alves, E.; Caetano, A.R.S.; Brandão, R.M.; Nelson, D.L.; Cardoso, M.D.G. Mechanism of action of various terpenes and phenylpropanoids against Escherichia coli and Staphylococcus aureus. FEMS Microbiol. Lett. 2021, 368, fnab052. [Google Scholar] [CrossRef]
- Al−Salihi, S.A.A.; Alberti, F. Naturally occurring terpenes: A promising class of organic molecules to address influenza pandemics. Nat. Prod. Bioprospect. 2021, 11, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Astani, A.; Schnitzler, P. Antiviral activity of monoterpenes beta−pinene and limonene against herpes simplex virus in vitro. Iran. J. Microbiol. 2014, 6, 149–155. [Google Scholar]
- De Clercq, E.; Li, G. Approved antiviral drugs over the past 50 years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kausar, S.; Said Khan, F.; Ishaq Mujeeb Ur Rehman, M.; Akram, M.; Riaz, M.; Rasool, G.; Hamid Khan, A.; Saleem, I.; Shamim, S.; Malik, A. A review: Mechanism of action of antiviral drugs. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211002621. [Google Scholar] [CrossRef] [PubMed]
- Valle, C.; Martin, B.; Touret, F.; Shannon, A.; Canard, B.; Guillemot, J.C.; Coutard, B.; Decroly, E. Drugs against SARS-CoV-2: What do we know about their mode of action? Rev. Med. Virol. 2020, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chatow, L.; Nudel, A.; Nesher, I.; Hayo Hemo, D.; Rozenberg, P.; Voropaev, H.; Winkler, I.; Levy, R.; Kerem, Z.; Yaniv, Z.; et al. In vitro evaluation of the activity of terpenes and cannabidiol against human coronavirus E229. Life 2021, 11, 290. [Google Scholar] [CrossRef]
- Boopathi, S.; Poma, A.B.; Kolandaivel, P. Novel 2019 coronavirus structure, mechanism of action, antiviral drug promises and rule out against its treatment. J. Biomol. Struct. Dyn. 2021, 39, 3409–3418. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial Activity of terpenes and terpenoids present in essential oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nweze, J.A.; Mbaoji, F.N.; Li, Y.M.; Yang, L.Y.; Huang, S.S.; Chigor, V.N.; Eze, E.A.; Pan, L.X.; Zhang, T.; Yang, D.F. Potentials of marine natural products against malaria, leishmaniasis, and trypanosomiasis parasites: A review of recent articles. Infect. Dis. Poverty. 2021, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Sobral, M.V.; Xavier, A.L.; Lima, T.C.; de Sousa, D.P. Antitumor activity of monoterpenes found in essential oils. Sci. World J. 2014, 2014, 953451. [Google Scholar] [CrossRef] [PubMed]
- Bahi, A.; Al Mansouri, S.; Al Memari, E.; Al Ameri, M.; Nurulain, S.M.; Ojha, S. β-Caryophyllene, a CB2 receptor agonist produces multiple behavioral changes relevant to anxiety and depression in mice. Physiol. Behav. 2014, 135, 119–124. [Google Scholar] [CrossRef]
- Guzmán−Gutiérrez, S.L.; Bonilla−Jaime, H.; Gómez−Cansino, R.; Reyes-Chilpa, R. Linalool and β-pinene exert their antidepressant-like activity through the monoaminergic pathway. Life Sci. 2015, 128, 24–29. [Google Scholar] [CrossRef]
- Brahmachari, G. Discovery and Development of Antidiabetic Agents from Natural Products Natural Product Drug Discovery; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Hosseini, M.; Chen, W.; Xiao, D.; Wang, C. Computational molecular docking and virtual screening revealed promising SARS-CoV-2 drugs. Precis. Clin. Med. 2021, 4, 1–16. [Google Scholar] [CrossRef]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Development of antibacterial conjugates using sulfamethoxazole with monocyclic terpenes: A systematic medicinal chemistry based computational approach. Comput. Method Program. Biomed. 2017, 140, 185–194. [Google Scholar] [CrossRef]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. ProTox-II: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug−likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Zhang, X.; Zhu, J.; Turpoff, A.; Chen, G.; Morrill, C.; Huang, S.; Lennox, W.; Kakarla, R.; Liu, R.; et al. Structure-activity relationship (SAR) optimization of 6-(indol-2-yl)pyridine-3-sulfonamides: Identification of potent, selective, and orally bioavailable small molecules targeting hepatitis C (HCV) NS4B. J. Med. Chem. 2014, 57, 2121–2135. [Google Scholar] [CrossRef]
- Machado, D.; Girardini, M.; Viveiros, M.; Pieroni, M. Challenging the drug-likeness dogma for new drug discovery in tuberculosis. Front Microbiol. 2018, 9, 1367. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N.; Hussain, T. Isoniazid-phytochemical conjugation: A new approach for potent and less toxic anti-TB drug development. Chem. Biol. Drug Des. 2020, 96, 714–730. [Google Scholar] [CrossRef]
- Saleemi, M.A.; Ahmad, B.; Benchoula, K.; Vohra, M.S.; Mea, H.J.; Chong, P.P.; Palanisamy, N.K.; Wong, E.H. Emergence and molecular mechanisms of SARS-CoV-2 and HIV to target host cells and potential therapeutics. Infect. Genet. Evol. 2020, 85, 104583. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, D.; Giovanetti, M.; Salemi, M.; Prosperi, M.; De Flora, C.; Junior Alcantara, L.C.; Angeletti, S.; Ciccozzi, M. The global spread of 2019-nCoV: A molecular evolutionary analysis. Pathog. Glob. Health 2020, 114, 64–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogara, P.A.; Omage, F.B.; Bolzan, G.R.; Delgado, C.P.; Aschner, M.; Orian, L.; Teixeira Rocha, J.B. In silico studies on the interaction between Mpro and PLpro from SARS-CoV-2 and Ebselen, its metabolites and derivatives. Mol. Inform. 2021, 40, e2100028. [Google Scholar] [CrossRef]
- Ojha, P.K.; Kar, S.; Krishna, J.G.; Roy, K.; Leszczynski, J. Therapeutics for COVID-19: From computation to practices-where we are, where we are heading to. Mol. Divers 2020, 22, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.S.; Panda, S.K.; Luyten, W. Phytochemicals against SARS-CoV as potential drug leads. Biomed. J. 2021, 44, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Kangabam, R.; Sahoo, S.; Ghosh, A.; Roy, R.; Silla, Y.; Misra, N.; Suar, M. Next−generation computational tools and resources for coronavirus research: From detection to vaccine discovery. Comput. Biol. Med. 2021, 128, 104158. [Google Scholar] [CrossRef]
- Sayed, A.M.; Alhadrami, H.A.; El-Gendy, A.O.; Shamikh, Y.I.; Belbahri, L.; Hassan, H.M.; Abdelmohsen, U.R.; Rateb, M.E. Microbial natural products as potential inhibitors of SARS-CoV-2 main protease (Mpro). Microorganisms 2020, 8, 970. [Google Scholar] [CrossRef]
- McKee, D.L.; Sternberg, A.; Stange, U.; Laufer, S.; Naujokat, C. Candidate drugs against SARS-CoV-2 and COVID-19. Pharmacol. Res. 2020, 157, 104859. [Google Scholar] [CrossRef]
- Keller, T.H.; Pichota, A.; Yin, Z. A practical view of ‘druggability’. Curr. Opin. Chem. Biol. 2006, 10, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Phytochemical conjugation as a potential semisynthetic approach toward reactive and reuse of obsolete sulfonamides against pathogenic bacteria. Drug Dev. Res. 2020, 82, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Weiss, C.; Carriere, M.; Fusco, L.; Capua, I.; Regla−Nava, J.A.; Pasquali, M.; Scoott, J.A.; Vitale, F.; Unal, M.A.; Mattevi, C.; et al. Toward nanotechnology-enabled approaches against the COVID-19 pandemic. ACS Nano 2020, 14, 6383–6406. [Google Scholar] [CrossRef]





{kind=link}
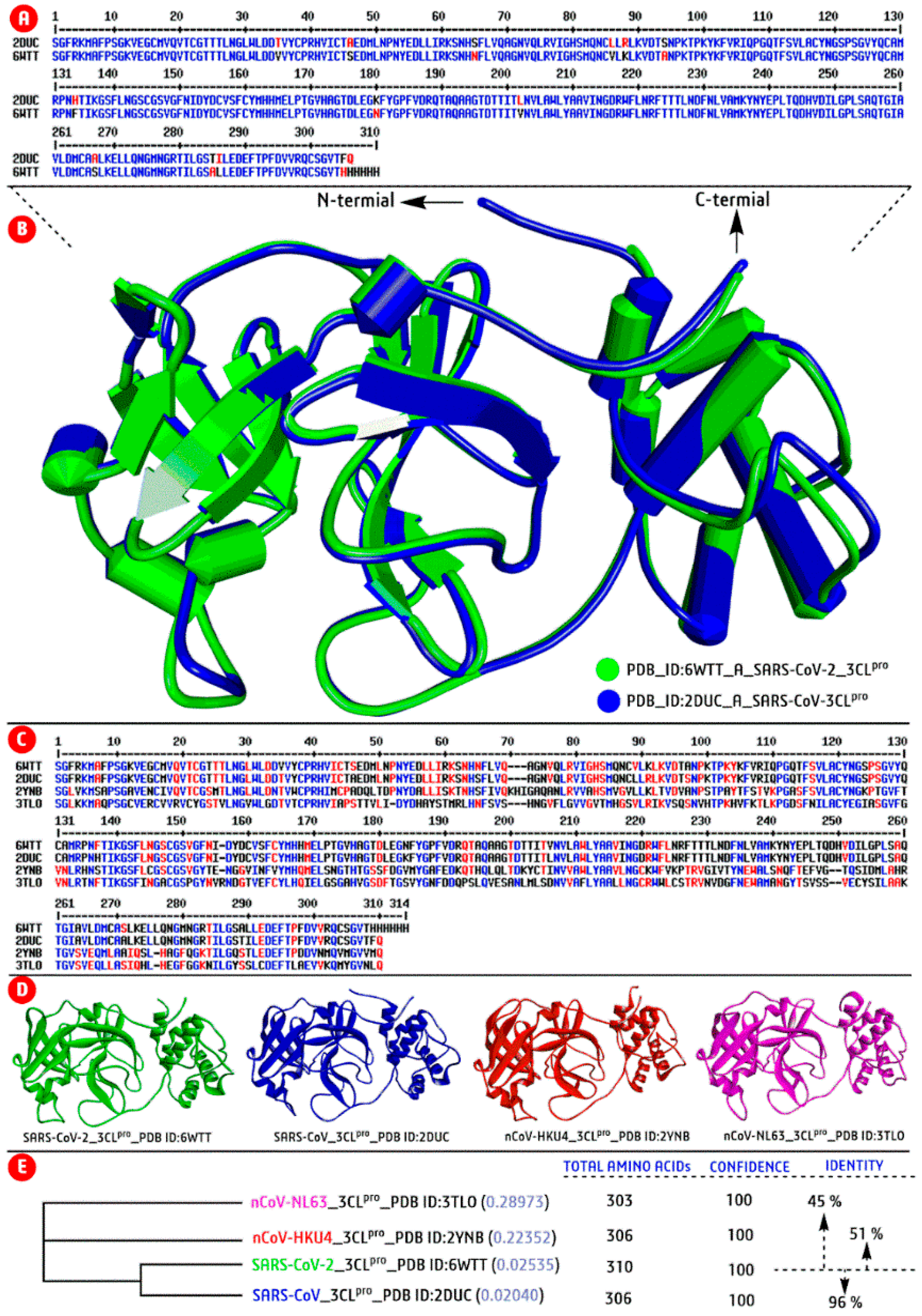{kind=link}
{kind=link}
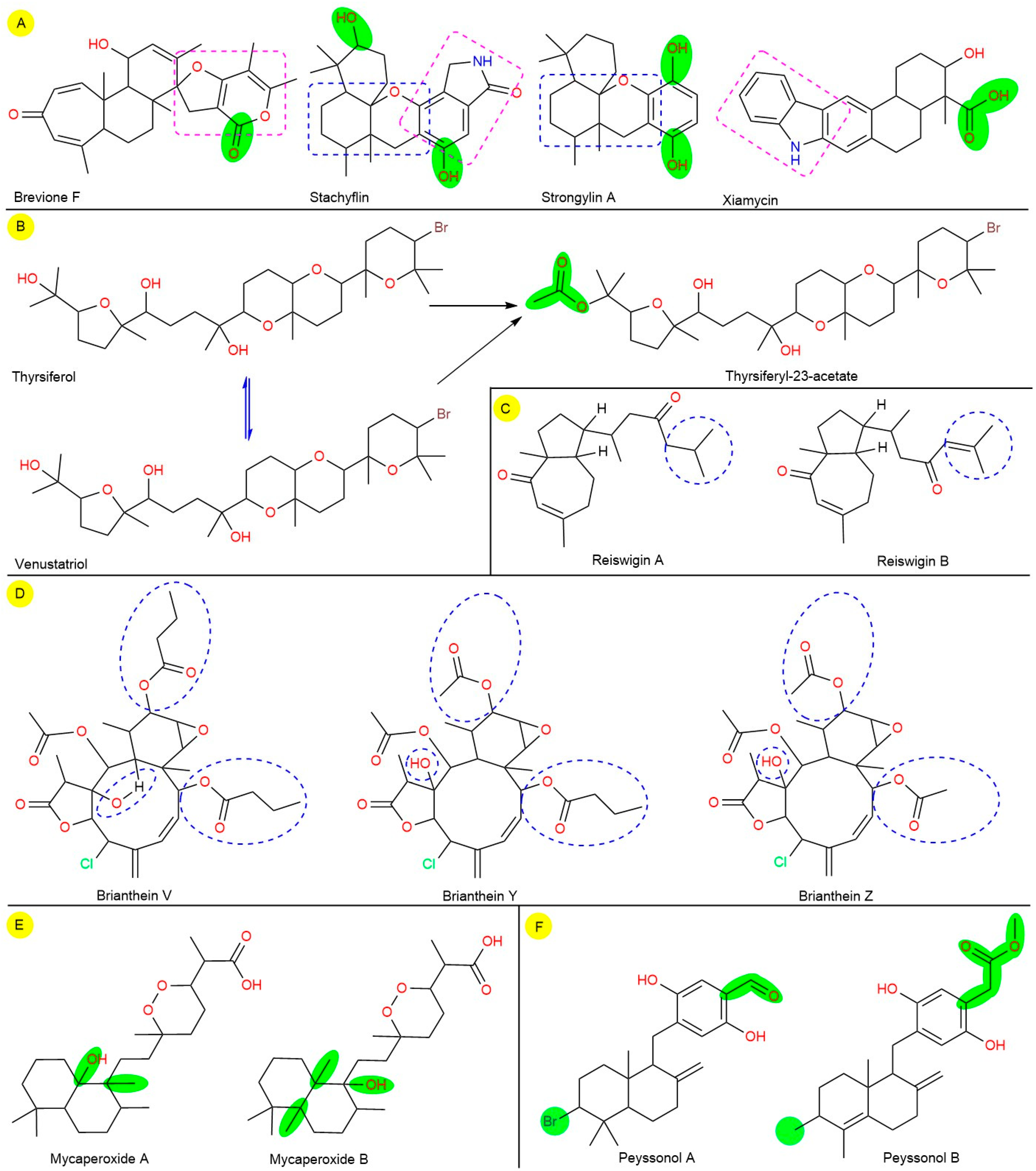{kind=link}
| Sl. No. | Name of the Terpenoid (Terpenoid Type) | Marine Source (Organism Type) | Antiviral Activity | Recorded IC50/EC50 (µM) | References |
|---|---|---|---|---|---|
| 1. | AcDa-1 (Diterpene) | Dictyota menstrualis (Algae) | HIV-1 | IC50: 35–70 | [27] |
| 2. | Acetyl ehrenberoxide B (Diterpenoid) | Sarcophyton ehrenbergi (Soft coral) | HCMV | EC50:22.0 | [28] |
| 3. | Alismol (Sesquiterpenoid) | Litophyton arboretum (Soft coral) | HIV-1 | IC50:7.20 | [29] |
| 4. | Aphidicolin (Diterpene) | Cephalosporium aphidicola (Sponge) | HSV, HCMV | IC50: 0.59 | [30] |
| 5. | Atomaric acid (Meroditerpenoid) | Stypopodium zonale (Brown algae) | HMPV | IC50: 7.96 | [31] |
| 6. | Avarol (Sesquiterpene hydroquinone) | Dysidea avara (Sponge) | HIV-1 | IC50: 0.30 | [32] |
| 7. | Brevione F (Spiroditerpenoid) | Penicillium sp. (Fungus) | HIV-1 | EC50: 14.7 | [33] |
| 8. | Brianthein V (Diterpene) | Briarurn asbestinum (Soft coral) | CoV-A59 | EC50: 83.75 | [34] |
| 9. | Brianthein Y (Diterpene) | Briarurn asbestinum (Soft coral) | CoV-A59 | EC50: 702.98 | [34] |
| 10. | Brianthein Z (Diterpene) | Briarurn asbestinum (Soft coral) | HSV-1 CoV-A59 | EC50: 147.87 | [34] |
| 11. | Bromophycolide A (Diterpene benzoate) | Callophycus serratus (Red algae) | HIV-1 | IC50: 9.1–9.8 | [34] |
| 12. | Capillobenzofuranol (Furano−sesquiterpenoid) | Sinularia capillosa (Soft coral) | HCMV | EC50: 13.5 | [35] |
| 13. | Capilloquinol (Diterpene quinoid) | Sinularia capillosa (Soft coral) | HCMV | ED50 11.24 | [36] |
| 14. | Curcuphenol (Sequiterpene) | Didiscus axeata (Sponge) | HCV | EC50: 31.2 | [37] |
| 15. | Cyanthiwigin B (Diterpene) | Myrmekoderma styx (Sponge) | HIV-1 HBV | EC50: 42.1 EC50: >333.33 | [38] |
| 16. | Da-1 (Diterpene) | Dictyota menstrualis (Brown algae) | HIV-1 | IC50:10−40 | [27] |
| 17. | Dehydrofurodendin (Sesterterpene) | Lendenfeldia sp. (Sponge) | HIV-1 | IC50: 3.2−5.6 | [39] |
| 18. | Dolabelladienol A (Diterpene) | Dictyota pfaffii (Brown algae) | HIV-1 | EC50: 2.9 ± 0.2 | [40] |
| 19. | Dolabelladienol B (Diterpene) | Dictyota pfaffii (Brown algae) | HIV-1 | EC50: 4.10 | [40] |
| 20. | Ehrenbergol A (Diterpenoid) | Sarcophyton ehrenbergi (Soft coral) | HCMV | IC50: 132.1 | [41] |
| 21. | Ehrenbergol B (Diterpenoid) | Sarcophyton ehrenbergi (Soft coral) | HCMV | IC50: 13.73 | [41] |
| 22. | Ehrenbergol C (Diterpenoid) | Sarcophyton ehrenbergi (Soft coral) | HCMV | EC50: 52.91 | [28] |
| 23. | Epispongiadiol (Diterpene) | Spongia sp. (Sponge) | HSV-1 | IC50: 37.65 | [42] |
| 24. | Epitaondiol (Meroditerpenoid) | Stypopodium zonale (Brown algae) | HMPV | IC50: 2.45 | [31] |
| 25. | Erythro-N-dodecanoyl-doco sasphinga-(4E,8E)-dienine (Sesquiterpenoid) | Litophyton arboreum (Soft coral) | HIV-1 | IC50: 4.80 | [43] |
| 26. | Gyrosanol A (Diterpenoid) | Sinularia gyrosa (Soft coral) | HCMV | IC50: 2.60 | [35] |
| 27. | Gyrosanol C (Diterpenoid) | Sinularia gyrosa (Soft coral) | HCMV | IC50: 3.70 | [35] |
| 28. | Halitunal (Diterpene aldehyde) | Halimeda tuna (Green seaweed) | CoV-A59 | IC50: 56.49 | [44] |
| 29. | Helioporin A (Diterpene) | Heliopora coerulea (Blue coral) | HSV-1 | IC50: 12.19 | [45] |
| 30. | Helioporin B (Diterpene) | Heliopora coerulea (Blue coral) | HSV-1 | IC50: 4.84 | [45] |
| 31. | Hyrtiosal (Sesterterpenoid) | Hyrtios erectus (Sponge) | HIV-1 | IC50: 9.60 | [46] |
| 32. | Ilimaquinone (Sesquiterpenoid quinones) | Dysudea arenaria (Sponge) | HIV-1 | IC50: 16.4 | [47] |
| 33. | Integric acid (Sesquiterpenoid) | Xylaria sp. (Ascomycetous fungi) | HIV-1 | IC50: 5−20 | [48] |
| 34. | Isospongiadiol (Diterpene) | Spongia sp. (Sponge) | HSV-1 | IC50: 6.02 | [42] |
| 35. | Liouvilloside A (Trisulfated triterpene glycoside) | Staurocucumis liouvillei (Sea cucumber) | HSV-1 | IC50: 6.96 | [49] |
| 36. | Liouvilloside B (Trisulfated triterpene glycoside) | Staurocucumis liouvillei (Sea cucumber) | HSV-1 | IC50: 6.93 | [49] |
| 37. | Lobohedleolide (Cembrane diterpenoid) | Lobophytum sp. (Soft coral) | HIV-1 | IC50: 10.90 | [50] |
| 38. | Mycaperoxide A (Norsesterterpene) | Mycale sp. (Sponge) | HSV-1, VSV | IC50: 0.6−4.0 | [51] |
| 39. | Mycaperoxide B (Norsesterterpene) | Mycale sp. (Sponge) | HSV-1, VSV | IC50: 0.58−2.35 | [51] |
| 40. | Peyssonol A (Sesquiterpene hydroquinone) | Peyssonnelia sp. (Red algae) | HIV-1 HIV-2 | IC50: 6.40 IC50: 21.30 | [52] |
| 41. | Peyssonol B (Sesquiterpene hydroquinone) | Peyssonnelia sp. (Red algae) | HIV-1 HIV-2 | IC50: 4.30 IC50: 14.70 | [52] |
| 42. | Pseudopterosin P (Diterpene glycoside) | Pseudopterogorgia elisabethae (Sponge) | HSV-1 & VZV, HCMV | EC50: 2.90 EC50: 2.60 | [53] |
| 43. | Puupehedione (Shikimatesequiterpene) | Verongida and Dictyoceratida sp. (Sponge) | HIV-1 | IC50: 3.01 | [54] |
| 44. | Reiswigin A (Diterpene) | Epipolasis reiswigi (Sponge) | HSV-1 & VSV CoV-A59 | EC50: 6.57 EC50: 65.7 | [55] |
| 45. | Reiswigin B (Diterpene) | Epipolasis reiswigi (Sponge) | HSV-1 & VSV, CoV-A59 | EC50: 6.62 EC50: 66.22 | [55] |
| 46. | Rietone (Triprenylhydro−quinone) | Alcyonium fauri (Soft coral) | HIV-1 | IC50:9.32 | [56] |
| 47. | Secopseudopterosin H (Diterpene glycoside) | Pseudopterogorgia elisabethae (Sponge) | HSV-1, VZV & HCMV | IC50: >10 | [53] |
| 48. | Seco−pseudopterosin I (Diterpene glycoside) | Pseudopterogorgia elisabethae (Sponge) | HSV-1, VZV & HCMV | IC50: >10 | [53] |
| 49. | Secocrassumol (Seco−cembranoid diterpene) | Lobophytum crissum (Soft coral) | HCMV | IC50: 12.69 | [43] |
| 50. | Sinuleptolide (Diterpene) | Sinulariana nolobata (Soft coral) | HCMV | ED50: 5.51 | [57] |
| 51. | Smenospongine (Sesquiterpenoid quinone) | Dysudea arenaria (Sponge) | HIV-1 | IC50: 176.1 | [47] |
| 52. | Smenotronic acid (Sesquiterpenoid quinone) | Dysudea arenaria (Sponge) | HIV-1 | IC50: 130.4 | [47] |
| 53. | Solenolide A (Diterpenoid lactone) | Solenopodium sp. (Marine octocoral) | Rhinovirus, HSV | IC50: 0.70 | [58] |
| 54. | Solenolide E (Diterpenoid lactone) | Solenopodium sp. (Marine octocoral) | Rhinovirus, HSV | IC50: 28.40 | [58] |
| 55. | Spongiadiol (Furanoditerpene) | Spongia sp. (Sponge) | HSV-1 | IC50: 0.75 | [42] |
| 56. | Stachybogrisephenone B (Sesquiterpenoid) | Stachybotry sp. HH1 ZSDS1F1-2 (Sponge) | EV71 | IC50: 30.10 | [59] |
| 57. | Stachyflin (Terpenoid) | Stachybotrys sp. RF−7260 (Fungus) | H1N1 | IC50: 0.003 | [60] |
| 58. | Strongylin A (Sesquiterpene hydroquinone) | Strongylophor ahartmani (Sponge) | H1N1 | IC50: 18.89 | [61] |
| 59. | Thyrsiferol (Triterpene) | Laurencia venusta (Red algae) | HSV-1, CoV−A59, VSV | ED50: 0.16–0.82 | [62] |
| 60. | Thyrsiferyl-23-acetate (Triterpene) | Laurencia venusta (Red algae) | HSV-1, CoV-A59, VSV | ED50: 0.15–0.77 | [62] |
| 61. | Usneoidol E (Meroterpene) | Cystoseira usneoides (Brown seaweed) | HSV, VSV | IC50: 7.62 IC50: 7.83 | [63] |
| 62. | Usneoidol Z (Meroterpene) | Cystoseira usneoides (Brown seaweed) | HSV, VSV | IC50: 8.47 IC50: 13.13 | [63] |
| 63. | Venustatriol (Triterpene) | Laurencia venusta (Red algae) | HSV-1, CoV-A59, VSV | ED50: 0.16–0.82 | [62] |
| 64. | Xiamycin (Pentacyclic indolosesquiterpene) | Bruguiera gymnorrhiza (Marine mangrove bacteria) | HIV | IC50: 5–20 | [64] |
| 65. | 7β-acetoxy-24-methyl cholesta-5-24(28)-diene-3,19-diol (Sesquiterpenoid) | Litophyton arboreum (Soft coral) | HIV-1 | IC50: 4.85 | [43] |
| 66. | (7Z)-Lobohedleolide (Cembranoid diterpene) | Lobophytum sp. (Soft coral) | HIV-1 | EC50: 13.93 | [50] |
| 67. | 8,10,18-trihydroxy-2,6-dolabelladiene (Diterpene) | Dictyota pfaffii (Brown algae) | HSV-1 | EC50: 5.10 | [65] |
| 68. | 17-dimethyl amino-lobohedleolide (Cembranoid diterpene) | Lobophytum sp. (Soft coral) | HIV-1 | IC50: 8.80 | [50] |
| Sl. No. | Docking Score | Drug Likeness Score | LD50 Score (mg/kg) | BA Score | Toxicity Profiles | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mpro/3CLpro | RdRp | HT | CG | IT | MG | CT | TC | ||||
| 1. | −7.4 | −6.7 | 0.42 | 1400 | 0.55 |  |  | | | | IV |
| 2. | −7.0 | −7.1 | −1.02 | 2000 | 0.55 | | | |  | | IV |
| 3. | −6.5 | −7.0 | −1.03 | 1200 | 0.55 | | | | | | IV |
| 4. | −6.8 | −6.9 | −0.15 | 500 | 0.55 | | | | | | IV |
| 5. | −7.8 | −7.1 | 0.58 | 750 | 0.56 | | | | | | IV |
| 6. | −7.9 | −7.5 | 0.41 | 2300 | 0.55 | | |  | | | V |
| 7. | −8.4 | −8.6 | 0.06 | 5 | 0.55 | | | | | | I |
| 8. | −6.5 | −7.5 | −0.77 | 7 | 0.55 | | | | | | II |
| 9. | −6.6 | −7.0 | 0.29 | 7 | 0.55 | | | | | | II |
| 10. | −7.0 | −6.6 | −0.99 | 7 | 0.55 | | | | | | II |
| 11. | −7.4 | −8.0 | 0.28 | 1000 | 0.17 | | | | | | IV |
| 12. | −8.0 | −7.3 | 0.29 | 500 | 0.55 | | | | | | IV |
| 13. | −7.7 | −7.7 | −0.75 | 500 | 0.55 | | | | | | IV |
| 14. | −5.4 | −6.1 | 0.28 | 1500 | 0.55 | | | | | | IV |
| 15. | −7.2 | −7.1 | 0.03 | 2200 | 0.55 | | | | | | V |
| 16. | −6.4 | −6.2 | 0.20 | 750 | 0.55 | | | | | | IV |
| 17. | −7.5 | −5.7 | −1.14 | 600 | 0.55 | | | | | | IV |
| 18. | −6.8 | −7.0 | −0.17 | 5000 | 0.55 | | | | | | V |
| 19. | −6.7 | −6.8 | −0.17 | 5000 | 0.55 | | | | | | V |
| 20. | −3.4 | −3.3 | −1.28 | 500 | 0.55 | | | | | | III |
| 21. | −2.9 | −3.5 | −0.41 | 500 | 0.55 | | | | | | III |
| 22. | −3.0 | −3.2 | −0.41 | 500 | 0.55 | | | | | | III |
| 23. | −6.7 | −7.0 | −0.34 | 79 | 0.55 | | | | | | III |
| 24. | −8.0 | −8.0 | −0.41 | 1000 | 0.55 | | | | | | IV |
| 25. | −6.8 | −7.7 | −1.08 | 2000 | 0.55 | | | | | | IV |
| 26. | −7.1 | −7.9 | −0.10 | 1016 | 0.55 | | | | | | IV |
| 27. | −6.1 | −7.0 | −0.81 | 2000 | 0.55 | | | | | | IV |
| 28. | −6.5 | −6.8 | −0.54 | 950 | 0.56 | | | | | | IV |
| 29. | −7.8 | −7.5 | −0.78 | 500 | 0.55 | | | | | | IV |
| 30. | −6.5 | −7.7 | −0.63 | 4000 | 0.55 | | | | | | V |
| 31. | −7.8 | −7.8 | −0.29 | 79 | 0.55 | | | | | | III |
| 32. | −7.3 | −7.2 | 0.10 | 2800 | 0.56 | | | | | | V |
| 33. | −6.7 | −6.5 | 0.25 | 5000 | 0.56 | | | | | | IV |
| 34. | −7.1 | −7.1 | −0.25 | 79 | 0.55 | | | | | | III |
| 35. | −7.3 | −8.8 | 0.50 | 4000 | 0.17 | | | | | | V |
| 36. | −7.5 | −8.9 | 0.45 | 4000 | 0.17 | | | | | | V |
| 37. | −6.7 | −7.3 | −1.40 | 1000 | 0.56 | | | | | | IV |
| 38. | −6.9 | −7.5 | −0.25 | 10000 | 0.56 | | | | | | VI |
| 39. | −6.4 | −6.8 | −0.69 | 10000 | 0.55 | | | | | | VI |
| 40. | −6.3 | −7.3 | −0.48 | 2300 | 0.55 | | | | | | V |
| 41. | −7.5 | −7.1 | 0.09 | 860 | 0.55 | | | | | | IV |
| 42. | −7.7 | −7.6 | 0.56 | 3000 | 0.55 | | | | | | V |
| 43. | −7.8 | −7.9 | −0.47 | 105 | 0.56 | | | | | | III |
| 44. | −6.1 | −6.5 | −0.28 | 5000 | 0.55 | | | | | | V |
| 45. | −6.1 | −6.0 | −0.11 | 400 | 0.55 | | | | | | IV |
| 46. | −6.1 | −7.4 | −0.32 | 400 | 0.55 | | | | | | IV |
| 47. | −7.4 | −7.3 | 0.29 | 3000 | 0.55 | | | | | | V |
| 48. | −6.9 | −7.5 | 1.08 | 3000 | 0.55 | | | | | | V |
| 49. | −7.1 | −6.4 | 0.41 | 3000 | 0.55 | | | | | | IV |
| 50. | −8.0 | −7.5 | −1.12 | 1000 | 0.55 | | | | | | IV |
| 51. | −7.2 | −7.5 | 0.29 | 100 | 0.56 | | | | | | III |
| 52. | −7.2 | −7.0 | −0.07 | 8 | 0.56 | | | | | | II |
| 53. | −6.5 | −7.0 | −0.40 | 7 | 0.55 | | | | | | II |
| 54. | −7.5 | −7.3 | −0.27 | 750 | 0.55 | | | | | | III |
| 55. | −7.5 | −7.3 | −0.34 | 79 | 0.55 | | | | | | III |
| 56. | −6.0 | −6.4 | −0.27 | 3200 | 0.55 | | | | | | V |
| 57. | −8.4 | −8.7 | 0.34 | 4000 | 0.55 | | | | | | V |
| 58. | −8.3 | −7.4 | 0.39 | 1000 | 0.55 | | | | | | IV |
| 59. | −8.2 | −9.2 | −0.33 | 2600 | 0.55 | | | | | | V |
| 60. | −8.0 | −8.6 | −0.40 | 5000 | 0.55 | | | | | | V |
| 61. | −6.7 | −6.2 | 0.47 | 2000 | 0.55 | | | | | | IV |
| 62. | −6.3 | −6.7 | 0.47 | 2000 | 0.55 | | | | | | IV |
| 63. | −7.6 | −8.7 | −0.33 | 2600 | 0.56 | | | | | | V |
| 64. | −8.4 | −9.3 | −0.02 | 1000 | 0.56 | | | | | | IV |
| 65. | −6.8 | −7.8 | −0.08 | 2000 | 0.55 | | | | | | IV |
| 66. | −7.3 | −7.6 | −0.84 | 1200 | 0.56 | | | | | | IV |
| 67. | −7.6 | −6.9 | −0.04 | 5000 | 0.55 | | | | | | IV |
| 68. | −6.6 | −7.0 | −0.25 | 1000 | 0.55 | | | | | | IV |
| 69. | −7.5 | ND | 0.60 | 245 | 0.55 | | | | | | III |
| 70. | −8.4 | ND | 1.10 | 5000 | 0.55 | | | | | | V |
| 71. | ND | −5.7 | −0.87 | 2000 | 0.55 | | | | | | IV |
| 72. | ND | −7.4 | 0.13 | 1200 | 0.17 | | | | | | IV |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahoo, A.; Fuloria, S.; Swain, S.S.; Panda, S.K.; Sekar, M.; Subramaniyan, V.; Panda, M.; Jena, A.K.; Sathasivam, K.V.; Fuloria, N.K. Potential of Marine Terpenoids against SARS-CoV-2: An In Silico Drug Development Approach. Biomedicines 2021, 9, 1505. https://doi.org/10.3390/biomedicines9111505
Sahoo A, Fuloria S, Swain SS, Panda SK, Sekar M, Subramaniyan V, Panda M, Jena AK, Sathasivam KV, Fuloria NK. Potential of Marine Terpenoids against SARS-CoV-2: An In Silico Drug Development Approach. Biomedicines. 2021; 9(11):1505. https://doi.org/10.3390/biomedicines9111505
Chicago/Turabian StyleSahoo, Alaka, Shivkanya Fuloria, Shasank S. Swain, Sujogya K. Panda, Mahendran Sekar, Vetriselvan Subramaniyan, Maitreyee Panda, Ajaya K. Jena, Kathiresan V. Sathasivam, and Neeraj Kumar Fuloria. 2021. "Potential of Marine Terpenoids against SARS-CoV-2: An In Silico Drug Development Approach" Biomedicines 9, no. 11: 1505. https://doi.org/10.3390/biomedicines9111505