
Simultaneous Measurement of Changes in Neutrophil Granulocyte Membrane Potential, Intracellular pH, and Cell Size by Multiparametric Flow Cytometry

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval and Neutrophil Isolation
2.2. Fluorescent Staining
2.3. Stimulation and Flow Cytometric Measurement
2.4. Membrane Potential Analysis
2.5. Intracellular pH Analysis
2.6. Analysis of Cellular Size by Flow Cytometry and Electronic Current Exclusion
2.7. Human Ex Vivo Whole Blood Model
2.8. Data Analysis and Statistics
3. Results
3.1. Methodological Aspects
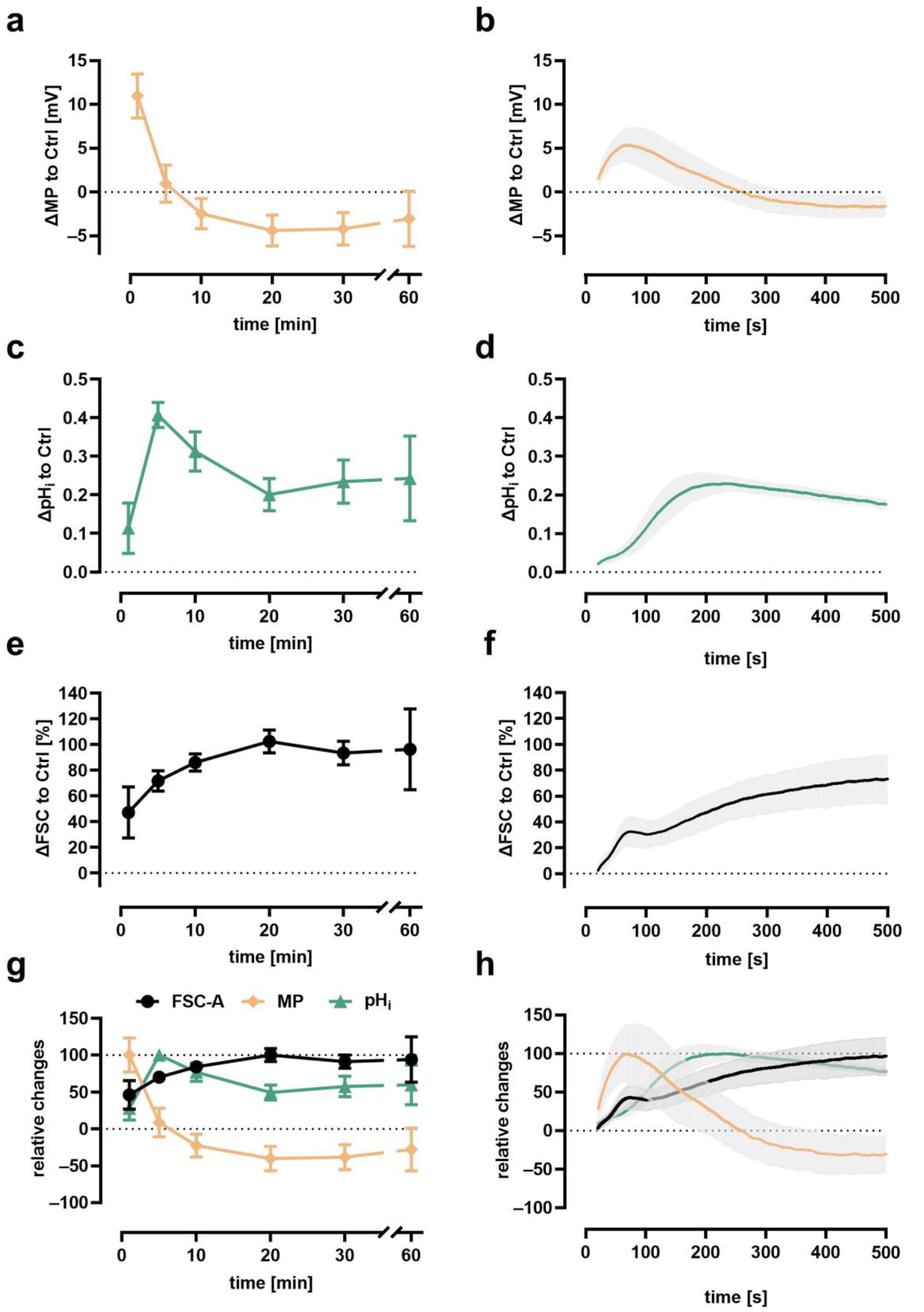
3.2. Time Course of Depolarization, Alkalization, and Cellular Swelling
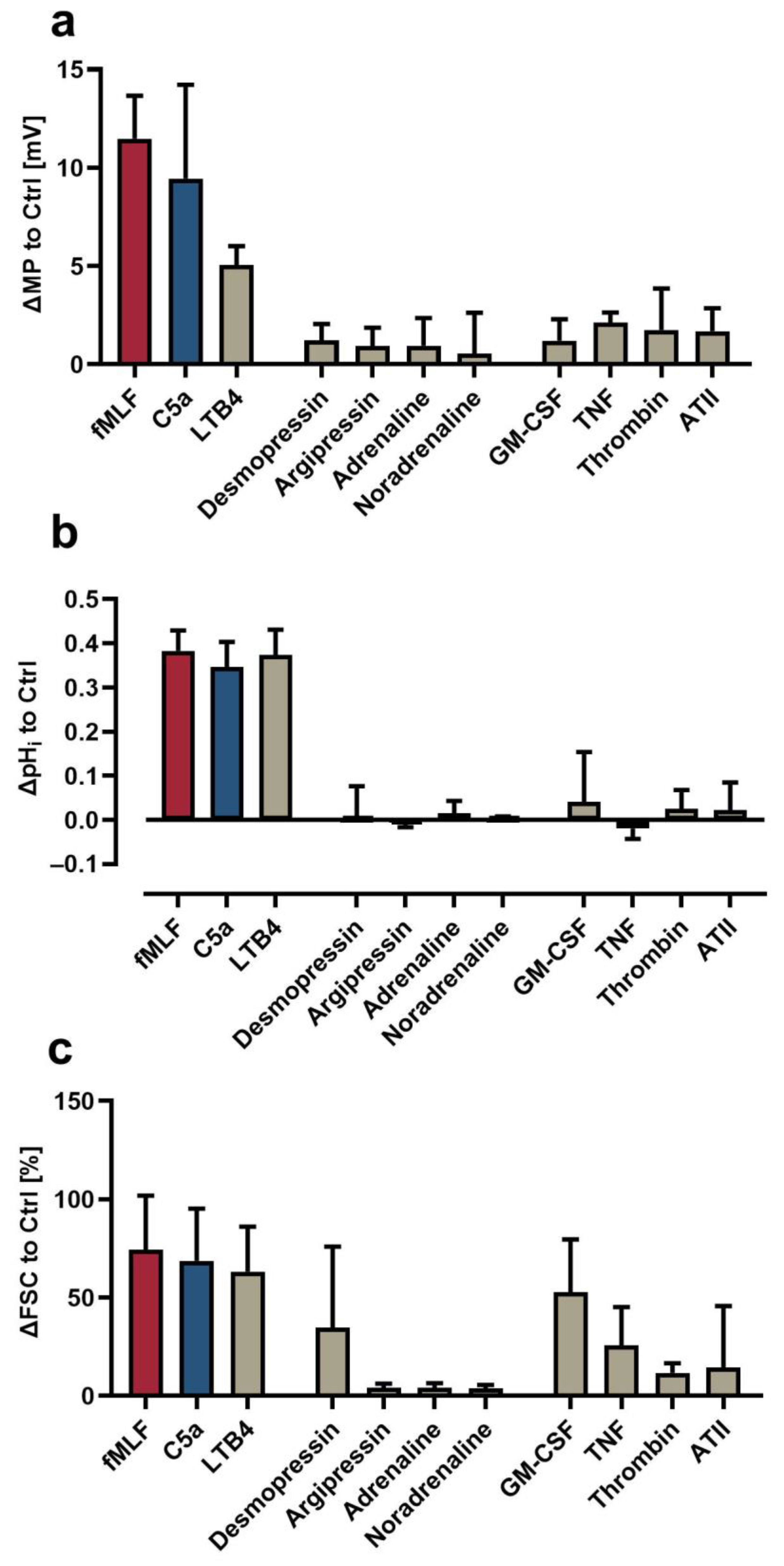
3.3. Comparison of the Neutrophil Response Elicited by Different Stimuli
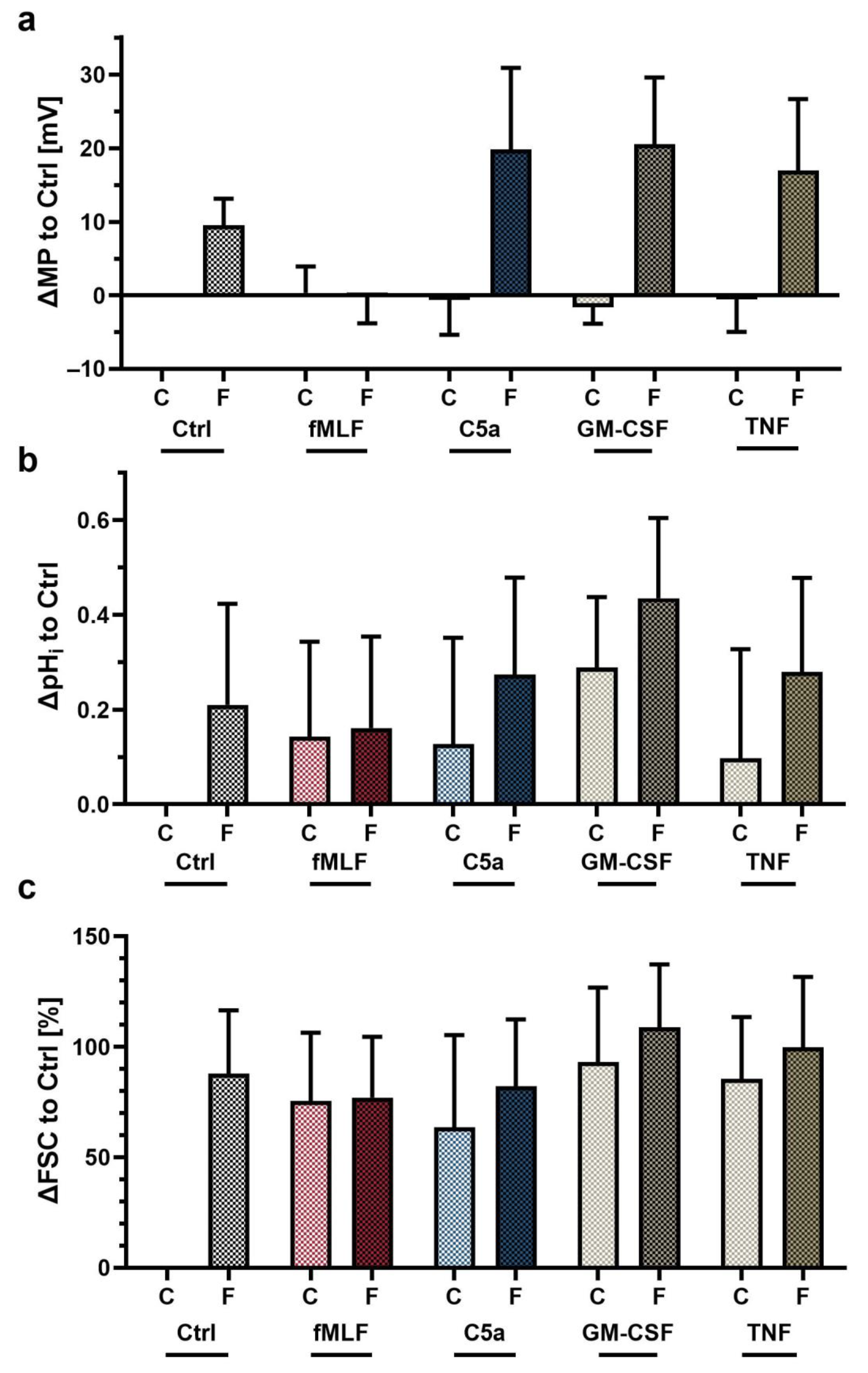
3.4. Exposure to Proinflammatory Stimuli In Vitro Changed the Neutrophil Response to fMLF
3.5. Endotoxemia in Whole Blood Markedly Alters the Neutrophil Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M.; Mumby, S.; Koenderman, L. Update on Neutrophil Function in Severe Inflammation. Front. Immunol. 2018, 9, 2171. [Google Scholar] [CrossRef] [Green Version]
- Kovtun, A.; Messerer, D.A.C.; Scharffetter-Kochanek, K.; Huber-Lang, M.; Ignatius, A. Neutrophils in Tissue Trauma of the Skin, Bone, and Lung: Two Sides of the Same Coin. J. Immunol. Res. 2018, 2018, 8173983. [Google Scholar] [CrossRef]
- Messerer, D.; Schmidt, H.; Frick, M.; Huber-Lang, M. Ion and Water Transport in Neutrophil Granulocytes and Its Impairment during Sepsis. Int. J. Mol. Sci. 2021, 22, 1699. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, S.; Rosowski, E.; Huttenlocher, A. Neutrophil migration in infection and wound repair: Going forward in reverse. Nat. Rev. Immunol. 2016, 16, 378–391. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Leopold, S.J.; Cranendonk, D.R.; Van Der Poll, T. Host innate immune responses to sepsis. Virulence 2013, 5, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messerer, D.A.C.; Halbgebauer, R.; Nilsson, B.; Pavenstädt, H.; Radermacher, P.; Huber-Lang, M. Immunopathophysiology of trauma-related acute kidney injury. Nat. Rev. Nephrol. 2020, 17, 91–111. [Google Scholar] [CrossRef]
- Sônego, F.; Castanheira, F.; Ferreira, R.G.; Kanashiro, A.; Leite, C.A.V.G.; Nascimento, D.C.; Colon, D.; Borges, V.D.F.; Alves-Filho, J.C.; Cunha, F.Q. Paradoxical Roles of the Neutrophil in Sepsis: Protective and Deleterious. Front. Immunol. 2016, 7, 155. [Google Scholar] [CrossRef] [Green Version]
- Singer, M.; Deutschman, C.S.; Seymour, C.C.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.C.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Alves-Filho, J.C.; Spiller, F.; Cunha, F.Q. Neutrophil paralysis in sepsis. Shock 2010, 34, 15–21. [Google Scholar] [CrossRef]
- Morris, A.C.; Brittan, M.; Wilkinson, T.S.; McAuley, D.F.; Antonelli, J.; McCulloch, C.; Barr, L.C.; McDonald, N.A.; Dhaliwal, K.; Jones, R.O.; et al. C5a-mediated neutrophil dysfunction is RhoA-dependent and predicts infection in critically ill patients. Blood 2011, 117, 5178–5188. [Google Scholar] [CrossRef] [PubMed]
- Huber-Lang, M.S.; Younkin, E.M.; Sarma, J.V.; McGuire, S.R.; Lu, K.T.; Guo, R.F.; Padgaonkar, V.A.; Curnutte, J.T.; Erickson, R.; Ward, P.A. Complement-Induced Impairment of Innate Immunity During Sepsis. J. Immunol. 2002, 169, 3223–3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messerer, D.A.C.; Denk, S.; Föhr, K.J.; Halbgebauer, R.; Braun, C.K.; Hönes, F.; Harant, J.; Fauler, M.; Frick, M.; Nußbaum, B.L.; et al. Complement C5a Alters the Membrane Potential of Neutrophils during Hemorrhagic Shock. Mediat. Inflamm. 2018, 2018, 2052356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denk, S.; Neher, M.D.; Messerer, D.A.C.; Wiegner, R.; Nilsson, B.; Rittirsch, D.; Nilsson-Ekdahl, K.; Weckbach, S.; Ignatius, A.; Kalbitz, M.; et al. Complement C5a Functions as a Master Switch for the pH Balance in Neutrophils Exerting Fundamental Immunometabolic Effects. J. Immunol. 2017, 198, 4846–4854. [Google Scholar] [CrossRef] [Green Version]
- Denk, S.; Taylor, R.P.; Wiegner, R.; Cook, E.M.; Lindorfer, M.A.; Pfeiffer, K.; Paschke, S.; Eiseler, T.; Weiss, M.; Barth, E.; et al. Complement C5a-Induced Changes in Neutrophil Morphology During Inflammation. Scand. J. Immunol. 2017, 86, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Hug, S.; Bernhard, S.; Stratmann, A.E.P.; Erber, M.; Wohlgemuth, L.; Knapp, C.L.; Bauer, J.M.; Vidoni, L.; Fauler, M.; Föhr, K.J.; et al. Activation of Neutrophil Granulocytes by Platelet-Activating Factor Is Impaired During Experimental Sepsis. Front. Immunol. 2021, 12, 580. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, S.; Hug, S.; Stratmann, A.E.P.; Erber, M.; Vidoni, L.; Knapp, C.L.; Thomaß, B.D.; Fauler, M.; Nilsson, B.; Ekdahl, K.N.; et al. Interleukin 8 Elicits Rapid Physiological Changes in Neutrophils That Are Altered by Inflammatory Conditions. J. Innate Immun. 2021, 13, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Immler, R.; Simon, S.I.; Sperandio, M. Calcium signalling and related ion channels in neutrophil recruitment and function. Eur. J. Clin. Investig. 2018, 48, e12964. [Google Scholar] [CrossRef] [PubMed]
- Forehand, J.R.; Pabst, M.J.; Phillips, A.W.; Johnston, R.B. Lipopolysaccharide priming of human neutrophils for an enhanced respiratory burst. Role of intracellular free calcium. J. Clin. Investig. 1989, 83, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, H.; Aharonovitz, O.; Alexander, R.T.; Touret, N.; Furuya, W.; Orlowski, J.; Grinstein, S. Na+/H+exchange and pH regulation in the control of neutrophil chemokinesis and chemotaxis. Am. J. Physiol. Physiol. 2008, 294, C526–C534. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, M.A.; Carretta, M.D.; Teuber, S.E.; Zárate, C.; Carcamo, L.; Concha, I.I.; Burgos, R.A. fMLP-Induced IL-8 Release Is Dependent on NADPH Oxidase in Human Neutrophils. J. Immunol. Res. 2015, 2015, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Hesselink, L.; Heeres, M.; Paraschiakos, F.; Berg, M.T.; Huisman, A.; Hoefer, I.E.; de Groot, M.C.; Van Solinge, W.; Dijkgraaf, M.; Hellebrekers, P.; et al. A Rise in Neutrophil Cell Size Precedes Organ Dysfunction After Trauma. Shock 2019, 51, 439–446. [Google Scholar] [CrossRef]
- El Chemaly, A.; Okochi, Y.; Sasaki, M.; Arnaudeau, S.; Okamura, Y.; Demaurex, N. VSOP/Hv1 proton channels sustain calcium entry, neutrophil migration, and superoxide production by limiting cell depolarization and acidification. J. Exp. Med. 2009, 207, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, L.M.; Chappell, J.B.; Jones, O.T.G. The superoxide-generating NADPH oxidase of human neutrophils is electrogenic and associated with an H+ channel. Biochem. J. 1987, 246, 325–329. [Google Scholar] [CrossRef] [PubMed]
- DeCoursey, T.E.; Morgan, D.; Cherny, V.V. The voltage dependence of NADPH oxidase reveals why phagocytes need proton channels. Nat. Cell Biol. 2003, 422, 531–534. [Google Scholar] [CrossRef] [PubMed]
- DeCoursey, T.E. The intimate and controversial relationship between voltage-gated proton channels and the phagocyte NADPH oxidase. Immunol. Rev. 2016, 273, 194–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiszt, M.; Kapus, A.; Német, K.; Farkas, L.; Ligeti, E. Regulation of Capacitative Ca2+ Influx in Human Neutrophil Granulocytes. J. Biol. Chem. 1997, 272, 26471–26478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowski, A.; Grinstein, S. A Noninvasive Fluorimetric Procedure for Measurement of Membrane Potential. Quantification of the NADPH Oxidase-Induced Depolarization in Activated Neutrophils. J. Biol. Chem. 1999, 274, 26098–26104. [Google Scholar] [CrossRef] [Green Version]
- Schrenzel, J.; Serrander, L.; Bánfi, B.; Nüße, O.; Fouyouzi, R.; Lew, D.P.; Demaurex, N.; Krause, K.-H. Electron currents generated by the human phagocyte NADPH oxidase. Nature 1998, 392, 734–737. [Google Scholar] [CrossRef]
- Sandoval, A.J.; Riquelme, J.P.; Carretta, M.D.; Hancke, J.L.; Hidalgo, M.A.; Burgos, R.A. Store-operated calcium entry mediates intracellular alkalinization, ERK1/2, and Akt/PKB phosphorylation in bovine neutrophils. J. Leukoc. Biol. 2007, 82, 1266–1277. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.; Capasso, M.; Musset, B.; Cherny, V.V.; Ríos, E.; Dyer, M.J.S.; DeCoursey, T.E. Voltage-gated proton channels maintain pH in human neutrophils during phagocytosis. Proc. Natl. Acad. Sci. USA 2009, 106, 18022–18027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coakley, R.J.; Taggart, C.; McElvaney, N.G.; O’Neill, S.J. Cytosolic pH and the inflammatory microenvironment modulate cell death in human neutrophils after phagocytosis. Blood 2002, 100, 3383–3391. [Google Scholar] [CrossRef] [PubMed]
- Grinstein, S.; Swallow, C.J.; Rotstein, O.D. Regulation of cytoplasmic pH in phagocytic cell function and dysfunction. Clin. Biochem. 1991, 24, 241–247. [Google Scholar] [CrossRef]
- Simchowitz, L.; Cragoe, E.J. Regulation of human neutrophil chemotaxis by intracellular pH. J. Biol. Chem. 1986, 261, 6492–6500. [Google Scholar] [CrossRef]
- Lewis, A.; Di Ciano, C.; Rotstein, O.D.; Kapus, A. Osmotic stress activates Rac and Cdc42 in neutrophils: Role in hypertonicity-induced actin polymerization. Am. J. Physiol. Physiol. 2002, 282, C271–C279. [Google Scholar] [CrossRef]
- DeCoursey, T.E.; Cherny, V.V.; Zhou, W.; Thomas, L.L. Simultaneous activation of NADPH oxidase-related proton and electron currents in human neutrophils. Proc. Natl. Acad. Sci. USA 2000, 97, 6885–6889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawa, K. Electrophysiological properties of three types of granulocytes in circulating blood of the newt. J. Physiol. 1989, 415, 211–231. [Google Scholar] [CrossRef] [Green Version]
- Morgan, D.; DeCoursey, T.E. Analysis of Electrophysiological Properties and Responses of Neutrophils. In Neutrophil Methods and Protocols; Quinn, M.T., DeLeo, F.R., Eds.; Humana Press: Totowa, NJ, USA, 2014; Volume 1124, pp. 121–158. ISBN 978-1-62703-844-7. [Google Scholar]
- Nanda, A.; Gukovskaya, A.; Tseng, J.; Grinstein, S. Activation of vacuolar-type proton pumps by protein kinase C. Role in neutrophil pH regulation. J. Biol. Chem. 1992, 267, 22740–22746. [Google Scholar] [CrossRef]
- Demaurex, N.; Downey, G.; Waddell, T.K.; Grinstein, S. Intracellular pH regulation during spreading of human neutrophils. J. Cell Biol. 1996, 133, 1391–1402. [Google Scholar] [CrossRef] [Green Version]
- Messerer, D.A.C.; Vidoni, L.; Erber, M.; Stratmann, A.E.P.; Bauer, J.M.; Braun, C.K.; Hug, S.; Adler, A.; Ekdahl, K.N.; Nilsson, B.; et al. Animal-Free Human Whole Blood Sepsis Model to Study Changes in Innate Immunity. Front. Immunol. 2020, 11, 571992. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Chang, H.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhingra, N.; Diepart, M.; Dziekan, G.; Khamassi, S.; Otaiza, F.; Wilburn, S.; Al-Abdulrazzak, R.; Bertsche, P.K.; Damani, N.; Lin, C.; et al. WHO Guidelines on Drawing Blood: Best Practices in Phlebotomy; World Health Organization: Geneva, Switzerland, 2010; pp. 1–105. [Google Scholar]
- Goldman, D.E. Potential, impedance, and rectification in membranes. J. Gen. Physiol. 1943, 27, 37–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klapperstück, T.; Glanz, D.; Hanitsch, S.; Klapperstück, M.; Markwardt, F.; Wohlrab, J. Calibration procedures for the quantitative determination of membrane potential in human cells using anionic dyes. Cytom. Part. A 2013, 83A, 612–626. [Google Scholar] [CrossRef]
- Jones, G.S.; Van Dyke, K.; Castranova, V. Purification of human granulocytes by centrifugal elutriation and measurement of transmembrane potential. J. Cell. Physiol. 1980, 104, 425–431. [Google Scholar] [CrossRef]
- Simchowitz, L.; Spilberg, I.; De Weer, P. Sodium and potassium fluxes and membrane potential of human neutrophils: Evidence for an electrogenic sodium pump. J. Gen. Physiol. 1982, 79, 453–479. [Google Scholar] [CrossRef]
- Chow, S.; Hedley, D.; Tannock, I. Flow Cytometric Calibration of Intracellular PH Measurements in Viable Cells Using Mixtures of Weak Acids and Bases. Cytometry 1996, 24, 360–367. [Google Scholar] [CrossRef]
- Klapperstück, T.; Glanz, D.; Klapperstück, M.; Wohlrab, J. Methodological aspects of measuring absolute values of membrane potential in human cells by flow cytometry. Cytom. Part. A 2009, 75A, 593–608. [Google Scholar] [CrossRef]
- Bräuner, T.; Hülser, D.F.; Strasser, R.J. Comparative measurements of membrane potentials with microelectrodes and voltage-sensitive dyes. Biochim. Biophys. Acta Biomembr. 1984, 771, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Epps, D.E.; Wolfe, M.L.; Groppi, V. Characterization of the steady-state and dynamic fluorescence properties of the potential-sensitive dye bis-(1,3-dibutylbarbituric acid)trimethine oxonol (Dibac4(3)) in model systems and cells. Chem. Phys. Lipids 1994, 69, 137–150. [Google Scholar] [CrossRef]
- Bashford, C.; Pasternak, C. Plasma membrane potential of neutrophils generated by the Na+ pump. Biochim. Biophys. Acta Biomembr. 1985, 817, 174–180. [Google Scholar] [CrossRef]
- Martin, A.M.; Nauseef, W.; Clark, A.R. Depolarization blunts the oxidative burst of human neutrophils. Parallel effects of monoclonal antibodies, depolarizing buffers, and glycolytic inhibitors. J. Immunol. 1988, 140, 3928–3935. [Google Scholar]
- Seligmann, B.E.; Gallin, J.I. Comparison of indirect probes of membrane potential utilized in studies of human neutrophils. J. Cell. Physiol. 1983, 115, 105–115. [Google Scholar] [CrossRef]
- Han, J.; Burgess, K. Fluorescent Indicators for Intracellular pH. Chem. Rev. 2009, 110, 2709–2728. [Google Scholar] [CrossRef] [PubMed]
- Loiselle, F.B.; Casey, J.R. Measurement of Intracellular pH. In Membrane Transporters; Humana Press: Totowa, NJ, USA, 2003; Volume 227, pp. 259–280. ISBN 978-1-59259-387-3. [Google Scholar]
- Alwazeer, D.; Cachon, R. Ion-selective electrode integrated in small-scale bioreactor for continuous intracellular pH determination in Lactobacillus plantarum. Folia Microbiol. 2019, 65, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yao, S.; Madou, M. A long-term stable iridium oxide pH electrode. Sens. Actuators B Chem. 2002, 81, 313–315. [Google Scholar] [CrossRef]
- Wille, C.; Eiseler, T.; Langenberger, S.-T.; Richter, J.; Mizuno, K.; Radermacher, P.; Knippschild, U.; Huber-Lang, M.; Seufferlein, T.; Paschke, S. PKD regulates actin polymerization, neutrophil deformability, and transendothelial migration in response to fMLP and trauma. J. Leukoc. Biol. 2018, 104, 615–630. [Google Scholar] [CrossRef]
- Hesselink, L.; Spijkerman, R.; De Fraiture, E.; Bongers, S.; Van Wessem, K.J.P.; Vrisekoop, N.; Koenderman, L.; Leenen, L.P.H.; Hietbrink, F. New automated analysis to monitor neutrophil function point-of-care in the intensive care unit after trauma. Intensiv. Care Med. Exp. 2020, 8, 1–12. [Google Scholar] [CrossRef]
- Spijkerman, R.; Hesselink, L.; Hellebrekers, P.; Vrisekoop, N.; Hietbrink, F.; Leenen, L.P.; Koenderman, L. Automated flow cytometry enables high performance point-of-care analysis of leukocyte phenotypes. J. Immunol. Methods 2019, 474, 112646. [Google Scholar] [CrossRef]
- Coakley, R.J.; Taggart, C.; Greene, C.; McElvaney, N.G.; O’Neill, S.J. Ambient pCO2 modulates intracellular pH, intracellular oxidant generation, and interleukin-8 secretion in human neutrophils. J. Leukoc. Biol. 2002, 71, 603–610. [Google Scholar]
- Coakley, R.J.; Taggart, C.; Canny, G.; Greally, P.; O’Neill, S.J.; McElvaney, N.G. Altered intracellular pH regulation in neutrophils from patients with cystic fibrosis. Am. J. Physiol. Cell. Mol. Physiol. 2000, 279, L66–L74. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stratmann, A.E.P.; Wohlgemuth, L.; Erber, M.E.; Bernhard, S.; Hug, S.; Fauler, M.; Vidoni, L.; Mohamed, A.O.K.; Thomaß, B.D.; Münnich, F.; et al. Simultaneous Measurement of Changes in Neutrophil Granulocyte Membrane Potential, Intracellular pH, and Cell Size by Multiparametric Flow Cytometry. Biomedicines 2021, 9, 1504. https://doi.org/10.3390/biomedicines9111504
Stratmann AEP, Wohlgemuth L, Erber ME, Bernhard S, Hug S, Fauler M, Vidoni L, Mohamed AOK, Thomaß BD, Münnich F, et al. Simultaneous Measurement of Changes in Neutrophil Granulocyte Membrane Potential, Intracellular pH, and Cell Size by Multiparametric Flow Cytometry. Biomedicines. 2021; 9(11):1504. https://doi.org/10.3390/biomedicines9111504
Chicago/Turabian StyleStratmann, Alexander Elias Paul, Lisa Wohlgemuth, Maike Elisabeth Erber, Stefan Bernhard, Stefan Hug, Michael Fauler, Laura Vidoni, Adam Omar Khalaf Mohamed, Bertram Dietrich Thomaß, Frederik Münnich, and et al. 2021. "Simultaneous Measurement of Changes in Neutrophil Granulocyte Membrane Potential, Intracellular pH, and Cell Size by Multiparametric Flow Cytometry" Biomedicines 9, no. 11: 1504. https://doi.org/10.3390/biomedicines9111504