Neutralizing Effects of Small Molecule Inhibitors and Metal Chelators on Coagulopathic Viperinae Snake Venom Toxins

 , and
, and 
Abstract
:1. Introduction
2. Experimental
2.1. Chemicals
2.2. Venom Nanofractionation
2.3. Plasma Coagulation Activity Assay
2.4. Correlation of Biological Data with MS Data
3. Results
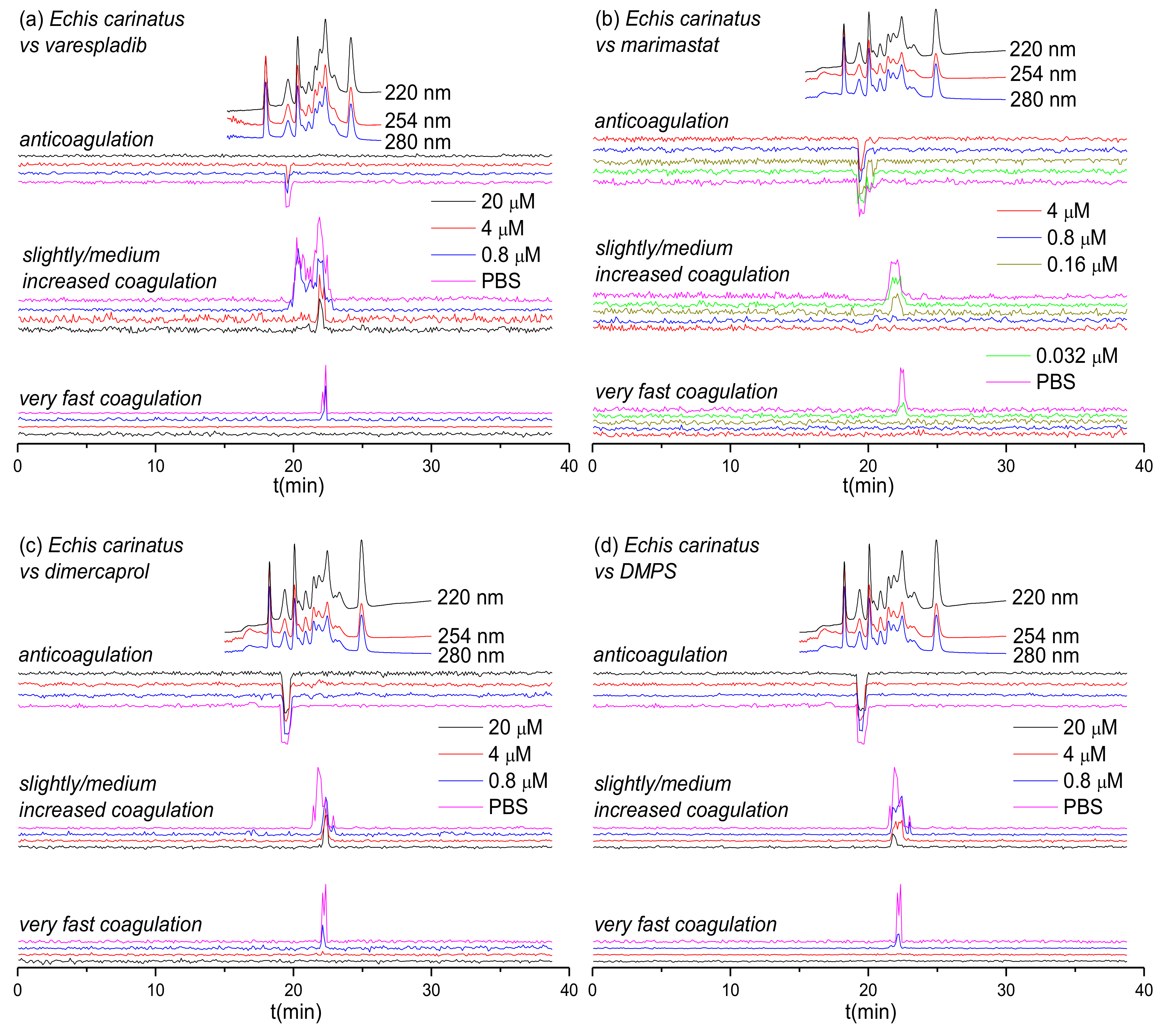
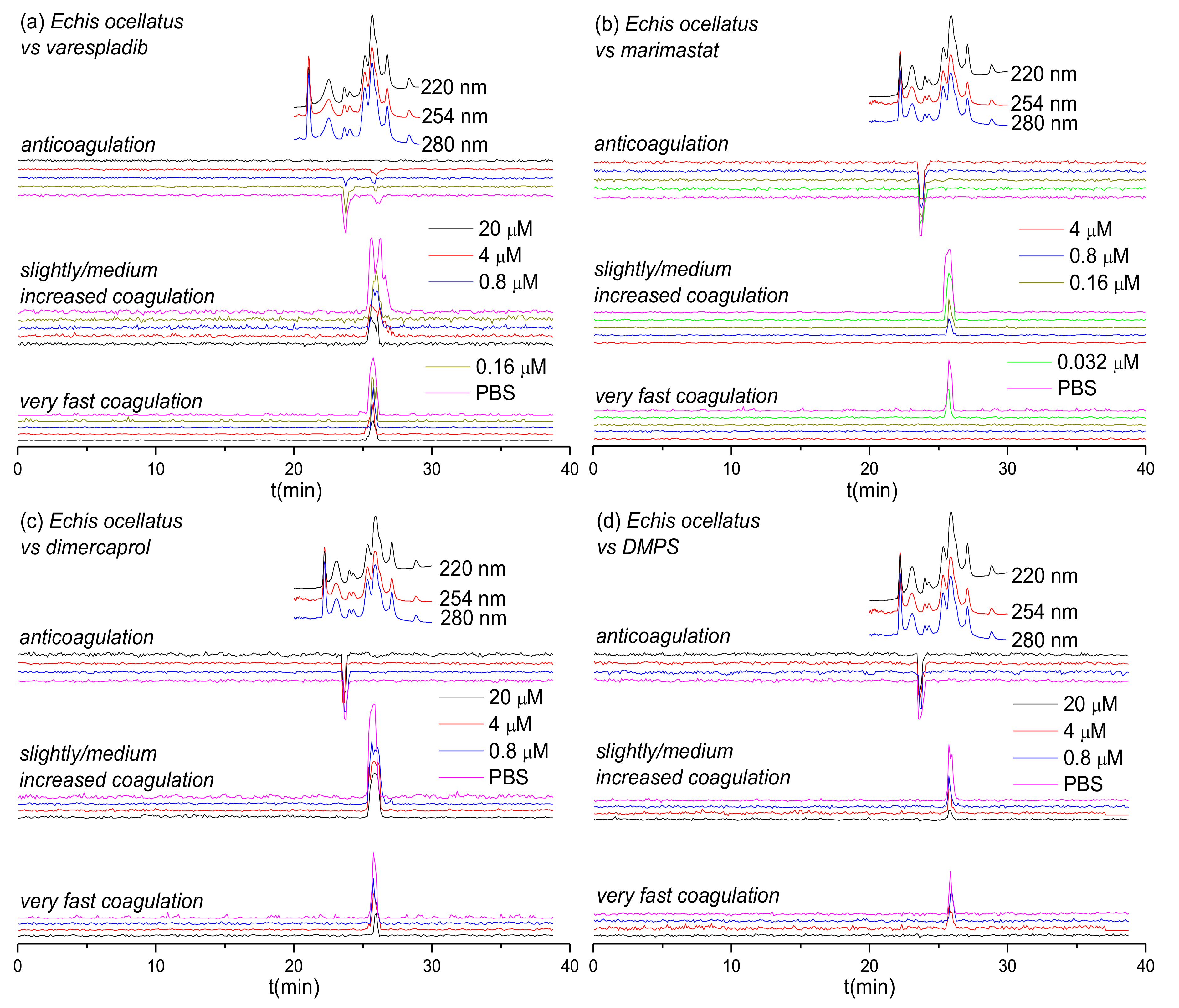
3.1. Inhibitory Effects of Varespladib, Marimastat, Dimercaprol and DMPS on Echis Venoms
3.2. Inhibitory Effect of Varespladib, Marimastat, Dimercaprol and DMPS on Daboia russelii Venom
3.3. Inhibitory Effects of Varespladib and Marimastat on Bitis arietans Venom
3.4. Identification of Coagulopathic Venom Toxins Neutralized by Small Molecule Inhibitors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Rogalski, A.; Soerensen, C.; Op den Brouw, B.; Lister, C.; Dashevsky, D.; Arbuckle, K.; Gloria, A.; Zdenek, C.N.; Casewell, N.R.; Gutiérrez, J.M. Differential procoagulant effects of saw-scaled viper (Serpentes: Viperidae: Echis) snake venoms on human plasma and the narrow taxonomic ranges of antivenom efficacies. Toxicol. Lett. 2017, 280, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Theakston, R.D.G.; Warrell, D.A. Confronting the neglected problem of snake bite envenoming: The need for a global partnership. PLoS Med. 2006, 3, e150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvete, J.J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M. Venoms, venomics, antivenomics. FEBS Lett. 2009, 583, 1736–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Q.; Clemetson, J.; Clemetson, K.J. Snake venoms and hemostasis. J. Thromb. Haemost. 2005, 3, 1791–1799. [Google Scholar] [CrossRef]
- Moura-da-Silva, A.; Butera, D.; Tanjoni, I. Importance of snake venom metalloproteinases in cell biology: Effects on platelets, inflammatory and endothelial cells. Curr. Pharm. Des. 2007, 13, 2893–2905. [Google Scholar] [CrossRef]
- Sant’Ana Malaque, C.M.; Gutiérrez, J.M. Snakebite Envenomation in Central and South America. Crit. Care Toxicol. 2016, 1–22. [Google Scholar] [CrossRef]
- Maduwage, K.; Isbister, G.K. Current treatment for venom-induced consumption coagulopathy resulting from snakebite. PLoS Negl. Trop. Dis. 2014, 8, e3220. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, S.; Slagboom, J.; Alomran, N.; Pla, D.; Alhamdi, Y.; King, S.I.; Bolton, F.M.; Gutiérrez, J.M.; Vonk, F.J.; Toh, C.-H. The paraspecific neutralisation of snake venom induced coagulopathy by antivenoms. Commun. Biol. 2018, 1, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Slagboom, J.; Kool, J.; Harrison, R.A.; Casewell, N.R. Haemotoxic snake venoms: Their functional activity, impact on snakebite victims and pharmaceutical promise. Br. J. Haematol. 2017, 177, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Kang, T.S.; Georgieva, D.; Genov, N.; Murakami, M.T.; Sinha, M.; Kumar, R.P.; Kaur, P.; Kumar, S.; Dey, S.; Sharma, S. Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. FEBS J. 2011, 278, 4544–4576. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Fujimura, Y.; Titani, K. Snake venom proteases affecting hemostasis and thrombosis. Biochim. Biophys. Acta 2000, 1477, 146–156. [Google Scholar] [CrossRef]
- Serrano, S.M.; Maroun, R.C. Snake venom serine proteinases: Sequence homology vs. substrate specificity, a paradox to be solved. Toxicon 2005, 45, 1115–1132. [Google Scholar] [CrossRef]
- Alvarez-Flores, M.P.; Faria, F.; de Andrade, S.A.; Chudzinski-Tavassi, A.M. Snake venom components affecting the coagulation system. In Snake Venoms; Gopalakrishnakone, P., Inagaki, H., Vogel, C.-W., Mukherjhee, A.K., Rahmy, T.R., Eds.; Springer: Dordrecht, Germany, 2017; pp. 417–436. [Google Scholar]
- Ramos, O.; Selistre-de-Araujo, H. Snake venom metalloproteases—structure and function of catalytic and disintegrin domains. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 142, 328–346. [Google Scholar] [CrossRef]
- Williams, H.F.; Layfield, H.J.; Vallance, T.; Patel, K.; Bicknell, A.B.; Trim, S.A.; Vaiyapuri, S. The urgent need to develop novel strategies for the diagnosis and treatment of snakebites. Toxins 2019, 11, 363. [Google Scholar] [CrossRef] [Green Version]
- Bulfone, T.C.; Samuel, S.P.; Bickler, P.E.; Lewin, M.R. Developing small molecule therapeutics for the Initial and adjunctive treatment of snakebite. J. Trop. Med. 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Resiere, D.; Gutiérrez, J.M.; Névière, R.; Cabié, A.; Hossein, M.; Kallel, H. Antibiotic therapy for snakebite envenoming. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, 1–2. [Google Scholar] [CrossRef]
- Albulescu, L.-O.; Hale, M.S.; Ainsworth, S.; Alsolaiss, J.; Crittenden, E.; Calvete, J.J.; Evans, C.; Wilkinson, M.C.; Harrison, R.A.; Kool, J. Preclinical validation of a repurposed metal chelator as an early-intervention therapeutic for hemotoxic snakebite. Sci. Transl. Med. 2020, 12, eaay8314. [Google Scholar] [CrossRef]
- Abraham, E.; Naum, C.; Bandi, V.; Gervich, D.; Lowry, S.F.; Wunderink, R.; Schein, R.M.; Macias, W.; Skerjanec, S.; Dmitrienko, A. Efficacy and safety of LY315920Na/S-5920, a selective inhibitor of 14-kDa group IIA secretory phospholipase A2, in patients with suspected sepsis and organ failure. Crit. Care Med. 2003, 31, 718–728. [Google Scholar] [CrossRef]
- Nicholls, S.J.; Kastelein, J.J.; Schwartz, G.G.; Bash, D.; Rosenson, R.S.; Cavender, M.A.; Brennan, D.M.; Koenig, W.; Jukema, J.W.; Nambi, V. Varespladib and cardiovascular events in patients with an acute coronary syndrome: The VISTA-16 randomized clinical trial. JAMA 2014, 311, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Lewin, M.; Samuel, S.; Merkel, J.; Bickler, P. Varespladib (LY315920) appears to be a potent, broad-spectrum, inhibitor of snake venom phospholipase A2 and a possible pre-referral treatment for envenomation. Toxins 2016, 8, 248. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, J.M.; Lewin, M.R.; Williams, D.; Lomonte, B. Varespladib (LY315920) and methyl varespladib (LY333013) abrogate or delay lethality induced by presynaptically acting neurotoxic snake venoms. Toxins 2020, 12, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan-Quirós, W.; Fernández, J.; Gutiérrez, J.M.; Lewin, M.R.; Lomonte, B. Neutralizing properties of LY315920 toward snake venom group I and II myotoxic phospholipases A2. Toxicon 2019, 157, 1–7. [Google Scholar] [CrossRef]
- Bittenbinder, M.A.; Zdenek, C.N.; Op den Brouw, B.; Youngman, N.J.; Dobson, J.S.; Naude, A.; Vonk, F.J.; Fry, B.G. Coagulotoxic cobras: Clinical implications of strong anticoagulant actions of African spitting Naja venoms that are not neutralised by antivenom but are by LY315920 (Varespladib). Toxins 2018, 10, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zdenek, C.N.; Youngman, N.J.; Hay, C.; Dobson, J.; Dunstan, N.; Allen, L.; Milanovic, L.; Fry, B.G. Anticoagulant activity of black snake (Elapidae: Pseudechis) venoms: Mechanisms, potency, and antivenom efficacy. Toxicol. Lett. 2020, 330, 176–184. [Google Scholar] [CrossRef]
- Underwood, C.; Min, D.; Lyons, J.; Hambley, T. The interaction of metal ions and Marimastat with matrix metalloproteinase 9. J. Inorg. Biochem. 2003, 95, 165–170. [Google Scholar] [CrossRef]
- Peterson, M.; Porter, K.; Loftus, I.; Thompson, M.; London, N. Marimastat inhibits neointimal thickening in a model of human arterial intimal hyperplasia. Eur. J. Vasc. Endovasc. Surg. 2000, 19, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Curran, S.; Murray, G.I. Matrix metalloproteinases in tumour invasion and metastasis. J. Pathol. 1999, 189, 300–308. [Google Scholar] [CrossRef]
- Rasmussen, H.S.; McCann, P.P. Matrix metalloproteinase inhibition as a novel anticancer strategy: A review with special focus on batimastat and marimastat. Pharmacol. Ther. 1997, 75, 69–75. [Google Scholar] [CrossRef]
- Evans, J.; Stark, A.; Johnson, C.; Daniel, F.; Carmichael, J.; Buckels, J.; Imrie, C.; Brown, P.; Neoptolemos, J. A phase II trial of marimastat in advanced pancreatic cancer. Br. J. Cancer 2001, 85, 1865. [Google Scholar] [CrossRef] [PubMed]
- Winer, A.; Adams, S.; Mignatti, P. Matrix metalloproteinase inhibitors in cancer therapy: Turning past failures into future successes. Mol. Cancer Ther. 2018, 17, 1147–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbaum, E.; Zahurak, M.; Sinibaldi, V.; Carducci, M.A.; Pili, R.; Laufer, M.; DeWeese, T.L.; Eisenberger, M.A. Marimastat in the treatment of patients with biochemically relapsed prostate cancer: A prospective randomized, double-blind, phase I/II trial. Clin. Cancer Res. 2005, 11, 4437–4443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, J.; Zhao, J.; Clingan, P.; Morris, D. Randomised double blind placebo control study of adjuvant treatment with the metalloproteinase inhibitor, Marimastat in patients with inoperable colorectal hepatic metastases: Significant survival advantage in patients with musculoskeletal side-effects. Anticancer Res. 2003, 23, 639–645. [Google Scholar] [PubMed]
- Levin, V.A.; Phuphanich, S.; Yung, W.A.; Forsyth, P.A.; Del Maestro, R.; Perry, J.R.; Fuller, G.N.; Baillet, M. Randomized, double-blind, placebo-controlled trial of marimastat in glioblastoma multiforme patients following surgery and irradiation. J. Neuro Oncol. 2006, 78, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Bramhall, S.; Schulz, J.; Nemunaitis, J.; Brown, P.; Baillet, M.; Buckels, J. A double-blind placebo-controlled, randomised study comparing gemcitabine and marimastat with gemcitabine and placebo as first line therapy in patients with advanced pancreatic cancer. Br. J. Cancer 2002, 87, 161. [Google Scholar] [CrossRef]
- Howes, J.-M.; Theakston, R.D.G.; Laing, G. Neutralization of the haemorrhagic activities of viperine snake venoms and venom metalloproteinases using synthetic peptide inhibitors and chelators. Toxicon 2007, 49, 734–739. [Google Scholar] [CrossRef]
- Zhang, D.; Botos, I.; Gomis-Rüth, F.-X.; Doll, R.; Blood, C.; Njoroge, F.G.; Fox, J.W.; Bode, W.; Meyer, E.F. Structural interaction of natural and synthetic inhibitors with the venom metalloproteinase, atrolysin C (form d). Proc. Natl. Acad. Sci. USA 1994, 91, 8447–8451. [Google Scholar] [CrossRef] [Green Version]
- Nagase, H.; Woessner, J.F. Matrix metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar] [CrossRef] [Green Version]
- Rucavado, A.; Escalante, T.; Gutiérrez, J.M. Effect of the metalloproteinase inhibitor batimastat in the systemic toxicity induced by Bothrops asper snake venom: Understanding the role of metalloproteinases in envenomation. Toxicon 2004, 43, 417–424. [Google Scholar] [CrossRef]
- Arias, A.S.; Rucavado, A.; Gutiérrez, J.M. Peptidomimetic hydroxamate metalloproteinase inhibitors abrogate local and systemic toxicity induced by Echis ocellatus (saw-scaled) snake venom. Toxicon 2017, 132, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Layfield, H.J.; Williams, H.F.; Ravishankar, D.; Mehmi, A.; Sonavane, M.; Salim, A.; Vaiyapuri, R.; Lakshminarayanan, K.; Vallance, T.M.; Bicknell, A.B. Repurposing cancer drugs batimastat and marimastat to inhibit the activity of a group I metalloprotease from the venom of the western diamondback rattlesnake, Crotalus atrox. Toxins 2020, 12, 309. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Model List of Essential Medicines, 20th list (March 2017). 2017. Available online: http://www.who.int/medicines/publications/essentialmedicines/20th_EML2017_FINAL_amendedAug2017.pdf?ua=1 (accessed on 15 November 2017).
- Tian, R.; Shi, R. Dimercaprol is an acrolein scavenger that mitigates acrolein-mediated PC-12 cells toxicity and reduces acrolein in rat following spinal cord injury. J. Neurochem. 2017, 141, 708–720. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Kumar, R.; Khadwal, A.; Singhi, S. Accidental inorganic mercury chloride poisoning in a 2-year old child. Indian J. Pediatrics 2010, 77, 1153–1155. [Google Scholar] [CrossRef]
- Kathirgamanathan, K.; Angaran, P.; Lazo-Langner, A.; Gula, L.J. Cardiac conduction block at multiple levels caused by arsenic trioxide therapy. Can. J. Cardiol. 2013, 29, 130.e5–130.e6. [Google Scholar] [CrossRef]
- Yajima, Y.; Kawaguchi, M.; Yoshikawa, M.; Okubo, M.; Tsukagoshi, E.; Sato, K.; Katakura, A. The effects of 2,3-dimercapto-1-propanesulfonic acid (DMPS) and meso-2,3-dimercaptosuccinic acid (DMSA) on the nephrotoxicity in the mouse during repeated cisplatin (CDDP) treatments. J. Pharmacol. Sci. 2017, 134, 108–115. [Google Scholar] [CrossRef]
- Aldhaheri, S.R.; Jeelani, R.; Kohan-Ghadr, H.R.; Khan, S.N.; Mikhael, S.; Washington, C.; Morris, R.T.; Abu-Soud, H.M. Dimercapto-1-propanesulfonic acid (DMPS) induces metaphase II mouse oocyte deterioration. Free Radic. Biol. Med. 2017, 112, 445–451. [Google Scholar] [CrossRef]
- Zietek, B.M.; Mayar, M.; Slagboom, J.; Bruyneel, B.; Vonk, F.J.; Somsen, G.W.; Casewell, N.R.; Kool, J. Liquid chromatographic nanofractionation with parallel mass spectrometric detection for the screening of plasmin inhibitors and (metallo) proteinases in snake venoms. Anal. Bioanal. Chem. 2018, 410, 5751–5763. [Google Scholar] [CrossRef] [Green Version]
- Mladic, M.; Zietek, B.M.; Iyer, J.K.; Hermarij, P.; Niessen, W.M.; Somsen, G.W.; Kini, R.M.; Kool, J. At-line nanofractionation with parallel mass spectrometry and bioactivity assessment for the rapid screening of thrombin and factor Xa inhibitors in snake venoms. Toxicon 2016, 110, 79–89. [Google Scholar] [CrossRef]
- Xie, C.; Slagboom, J.; Albulescu, L.-O.; Bruyneel, B.; Still, K.; Vonk, F.J.; Somsen, G.W.; Casewell, N.R.; Kool, J. Antivenom neutralization of coagulopathic snake venom toxins assessed by bioactivity profiling using nanofractionation analytics. Toxins 2020, 12, 53. [Google Scholar] [CrossRef] [Green Version]
- Slagboom, J.; Mladić, M.; Xie, C.; Kazandjian, T.D.; Vonk, F.; Somsen, G.W.; Casewell, N.R.; Kool, J. High throughput screening and identification of coagulopathic snake venom proteins and peptides using nanofractionation and proteomics approaches. PLoS Negl. Trop. Dis. 2020, 14, e0007802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.; Albulescu, L.-O.; Still, K.; Slagboom, J.; Zhao, Y.; Jiang, Z.; Somsen, G.W.; Vonk, F.J.; Casewell, N.R.; Kool, J. Varespladib inhibits the phospholipase A2 and coagulopathic activities of venom components from hemotoxic snakes. Biomedicines 2020, 8, 165. [Google Scholar] [CrossRef] [PubMed]
- Still, K.; Nandlal, R.S.; Slagboom, J.; Somsen, G.W.; Casewell, N.R.; Kool, J. Multipurpose HTS coagulation analysis: Assay development and assessment of coagulopathic snake venoms. Toxins 2017, 9, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Das, D.; Iyer, J.K.; Kini, R.M.; Doley, R. Unveiling the complexities of Daboia russelii venom, a medically important snake of India, by tandem mass spectrometry. Toxicon 2015, 107, 266–281. [Google Scholar] [CrossRef]
- Hiremath, V.; Urs, A.N.; Joshi, V.; Suvilesh, K.; Savitha, M.; Amog, P.U.; Rudresha, G.; Yariswamy, M.; Vishwanath, B. Differential action of medically important Indian BIG FOUR snake venoms on rodent blood coagulation. Toxicon 2016, 110, 19–26. [Google Scholar] [CrossRef]
- Hiremath, V.; Yariswamy, M.; Nanjaraj Urs, A.; Joshi, V.; Suvilesh, K.; Ramakrishnan, C.; Nataraju, A.; Vishwanath, B. Differential action of Indian BIG FOUR snake venom toxins on blood coagulation. Toxin Rev. 2014, 33, 23–32. [Google Scholar] [CrossRef]
- Albulescu, L.-O.; Xie, C.; Ainsworth, S.; Alsolaiss, J.; Crittenden, E.; Dawson, C.A.; Softley, R.; Bartlett, K.E.; Harrison, R.A.; Kool, J.; et al. A combination of two small molecule toxin inhibitors provides pancontinental preclinical efficacy against viper snakebite. bioRxiv 2020. (preprint). [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Zhang, D.; Xiao, H.; Xiong, S.; Huang, C. Exploration of the inhibitory potential of varespladib for snakebite envenomation. Molecules 2018, 23, 391. [Google Scholar] [CrossRef] [Green Version]
- Kini, R.M.; Sidhu, S.S.; Laustsen, A.H. Biosynthetic oligoclonal antivenom (BOA) for snakebite and next-generation treatments for snakebite victims. Toxins 2018, 10, 534. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, C.; Ledsgaard, L.; Dehli, R.I.; Ahmadi, S.; Sørensen, C.V.; Laustsen, A.H. Engineering and design considerations for next-generation snakebite antivenoms. Toxicon 2019, 167, 67–75. [Google Scholar] [CrossRef]
- Lewin, M.R.; Gutiérrez, J.M.; Samuel, S.P.; Herrera, M.; Bryan-Quirós, W.; Lomonte, B.; Bickler, P.E.; Bulfone, T.C.; Williams, D.J. Delayed oral LY333013 rescues mice from highly neurotoxic, lethal doses of Papuan Taipan (Oxyuranus scutellatus) venom. Toxins 2018, 10, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewin, M.R.; Gilliam, L.L.; Gilliam, J.; Samuel, S.P.; Bulfone, T.C.; Bickler, P.E.; Gutiérrez, J.M. Delayed LY333013 (oral) and LY315920 (intravenous) reverse severe neurotoxicity and rescue juvenile pigs from lethal doses of Micrurus fulvius (Eastern Coral snake) venom. Toxins 2018, 10, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiorino, R.M.; Dart, R.C.; Carter, D.E.; Aposhian, H.V. Determination and metabolism of dithiol chelating agents. XII. Metabolism and pharmacokinetics of sodium 2, 3-dimercaptopropane-1-sulfonate in humans. J. Pharmacol. Exp. Ther. 1991, 259, 808–814. [Google Scholar] [PubMed]
- Planas-Bohne, F.; Gabard, B.; Schäffer, E. Toxicological studies on sodium 2, 3-dimercaptopropane-1-sulfonate in the rat. In Arzneimittel-Forschung; Thieme Medical Publishers: Leipzig, Germany, 1980; Volume 30, pp. 1291–1294. [Google Scholar]
- Domingo, J.L.; Ortega, A.; Bosque, M.; Corbella, J. Evaluation of the developmental effects on mice after prenatal, or pre-and postnatal exposure to 2, 3-dimercaptopropane-1-sulfonic acid (DMPS). Life Sci. 1990, 46, 1287–1292. [Google Scholar] [CrossRef]
- Sharma, S.K.; Chappuis, F.; Jha, N.; Bovier, P.A.; Loutan, L.; Koirala, S. Impact of snake bites and determinants of fatal outcomes in southeastern Nepal. Am. J. Trop. Med. Hyg. 2004, 71, 234–238. [Google Scholar] [CrossRef]
- Abubakar, S.; Habib, A.; Mathew, J. Amputation and disability following snakebite in Nigeria. Tropical Doctor 2010, 40, 114–116. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Peak Retention Time (min) | Mascot Results Matching the Exact Mass | Exact Mass from MS Data | Exact Mass from Mascot Data | Toxin Class | Activity | Dose Required for Full Inhibition |
|---|---|---|---|---|---|---|---|
| E. carinatus | 19.1–19.9 | PA2A1_ECHCA | – | 16310 | PLA2 | Anticoagulant | 20 μM varespladib |
| 19.9–23.1 | – | – | – | – | Procoagulant | 0.8 μM marimastat | |
| E. ocellatus | 23.4–24.4 | PA2A5_ECHOC | 13856.138 | 13856 | PLA2 | Anticoagulant | 4 μM varespladib |
| 25.1–27.1 | VM3E2_ECHOC | – | 69426 | SVMP | Procoagulant | 0.16 μM marimastat/20 μM DMPS | |
| 25.1–27.1 | VM3E6_ECHOC | – | 57658 | SVMP | Procoagulant | 0.16 μM marimastat/20 μM DMPS | |
| 25.1–27.1 | SL1_ECHOC | – | 16601 | CTL | Procoagulant | 0.16 μM marimastat/20 μM DMPS | |
| 25.1–27.1 | SL124_ECHOC | – | 16882 | CTL | Procoagulant | 0.16 μM marimastat/20 μM DMPS | |
| D. russelii | 18.6–21.5 | PA2B8_DABRR | 13587.225 | 13587 | PLA2 | Anticoagulant | 20 μM varespladib |
| 18.6–21.5 | PA2B5_DABRR | – | 13587 | PLA2 | Anticoagulant | 20 μM varespladib | |
| 18.6–21.5 | PA2B3_DABRR | – | 13687 | PLA2 | Anticoagulant | 20 μM varespladib | |
| 21.5–22.8 | – | – | – | – | Procoagulant | 4 μM marimastat | |
| B. arietans | 16.7–17.1 | SLA_BITAR | – | 14935 | CTL | Anticoagulant | 0.8 μM varespladib |
| 16.7–17.1 | SLB_BITAR | – | 14798 | CTL | Anticoagulant | 0.8 μM varespladib |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, C.; Albulescu, L.-O.; Bittenbinder, M.A.; Somsen, G.W.; Vonk, F.J.; Casewell, N.R.; Kool, J. Neutralizing Effects of Small Molecule Inhibitors and Metal Chelators on Coagulopathic Viperinae Snake Venom Toxins. Biomedicines 2020, 8, 297. https://doi.org/10.3390/biomedicines8090297
Xie C, Albulescu L-O, Bittenbinder MA, Somsen GW, Vonk FJ, Casewell NR, Kool J. Neutralizing Effects of Small Molecule Inhibitors and Metal Chelators on Coagulopathic Viperinae Snake Venom Toxins. Biomedicines. 2020; 8(9):297. https://doi.org/10.3390/biomedicines8090297
Chicago/Turabian StyleXie, Chunfang, Laura-Oana Albulescu, Mátyás A. Bittenbinder, Govert W. Somsen, Freek J. Vonk, Nicholas R. Casewell, and Jeroen Kool. 2020. "Neutralizing Effects of Small Molecule Inhibitors and Metal Chelators on Coagulopathic Viperinae Snake Venom Toxins" Biomedicines 8, no. 9: 297. https://doi.org/10.3390/biomedicines8090297