Characterisation of a Novel A-Superfamily Conotoxin

 and
and
Abstract
:1. Introduction
2. Experimental Section
2.1. Peptide Synthesis, Purification and Characterisation
2.2. NMR Spectroscopy
2.3. Structure Calculations
2.4. Electrophysiological Measurements
2.5. FLIPRTetra Ion Channel Assays
3. Results
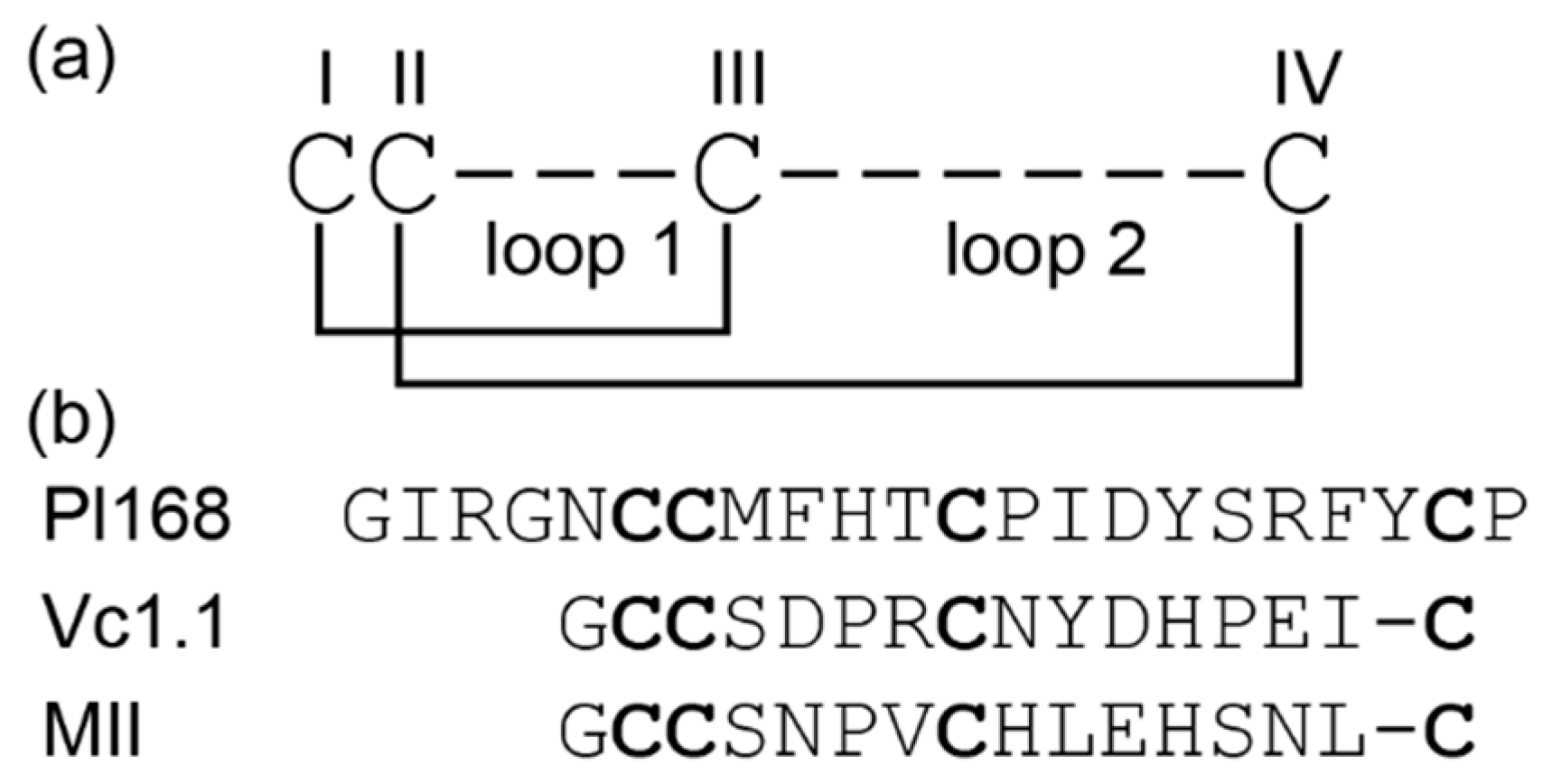
3.1. Peptide Synthesis and Characterisation
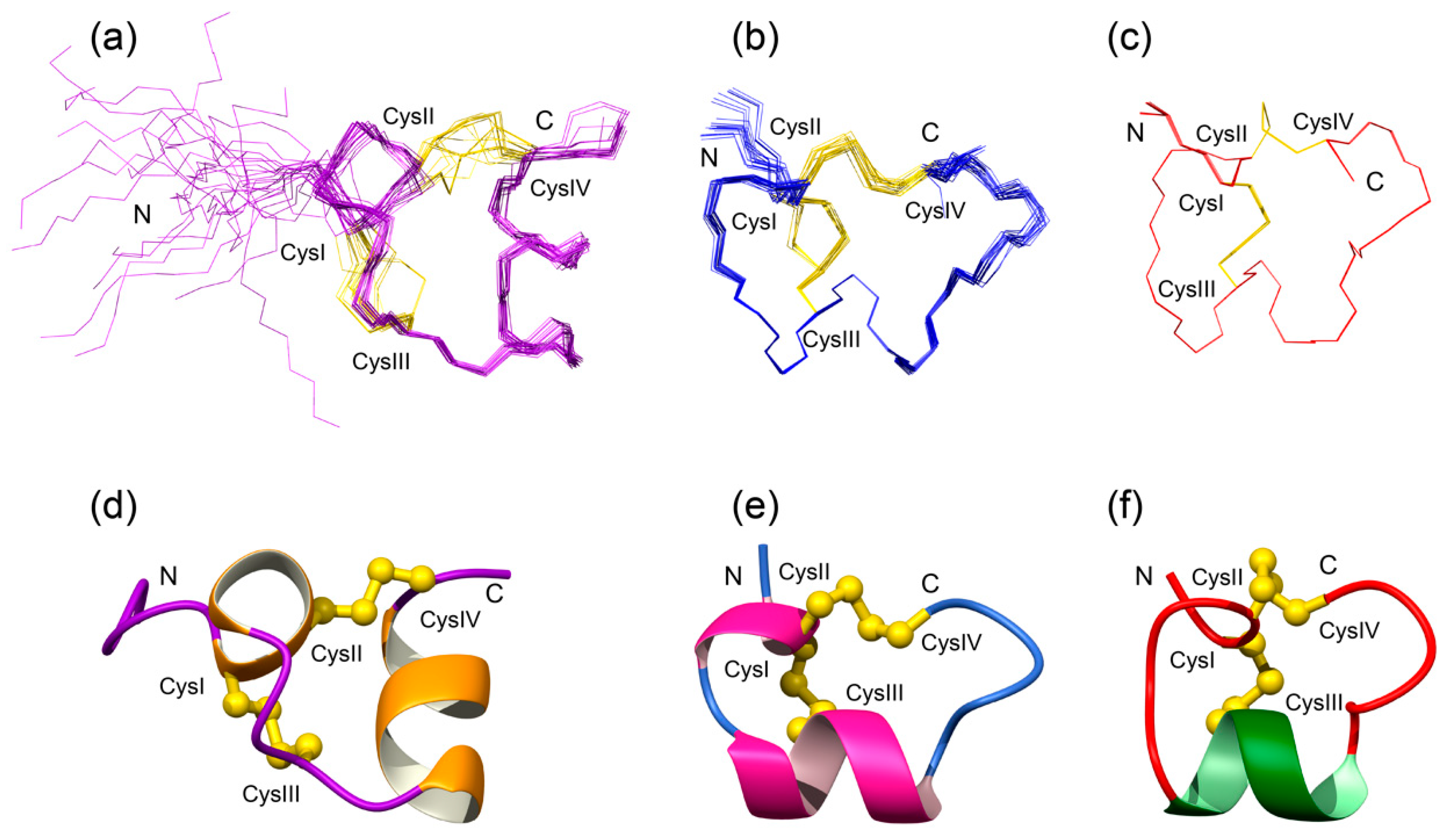
3.2. Structural Characterisation
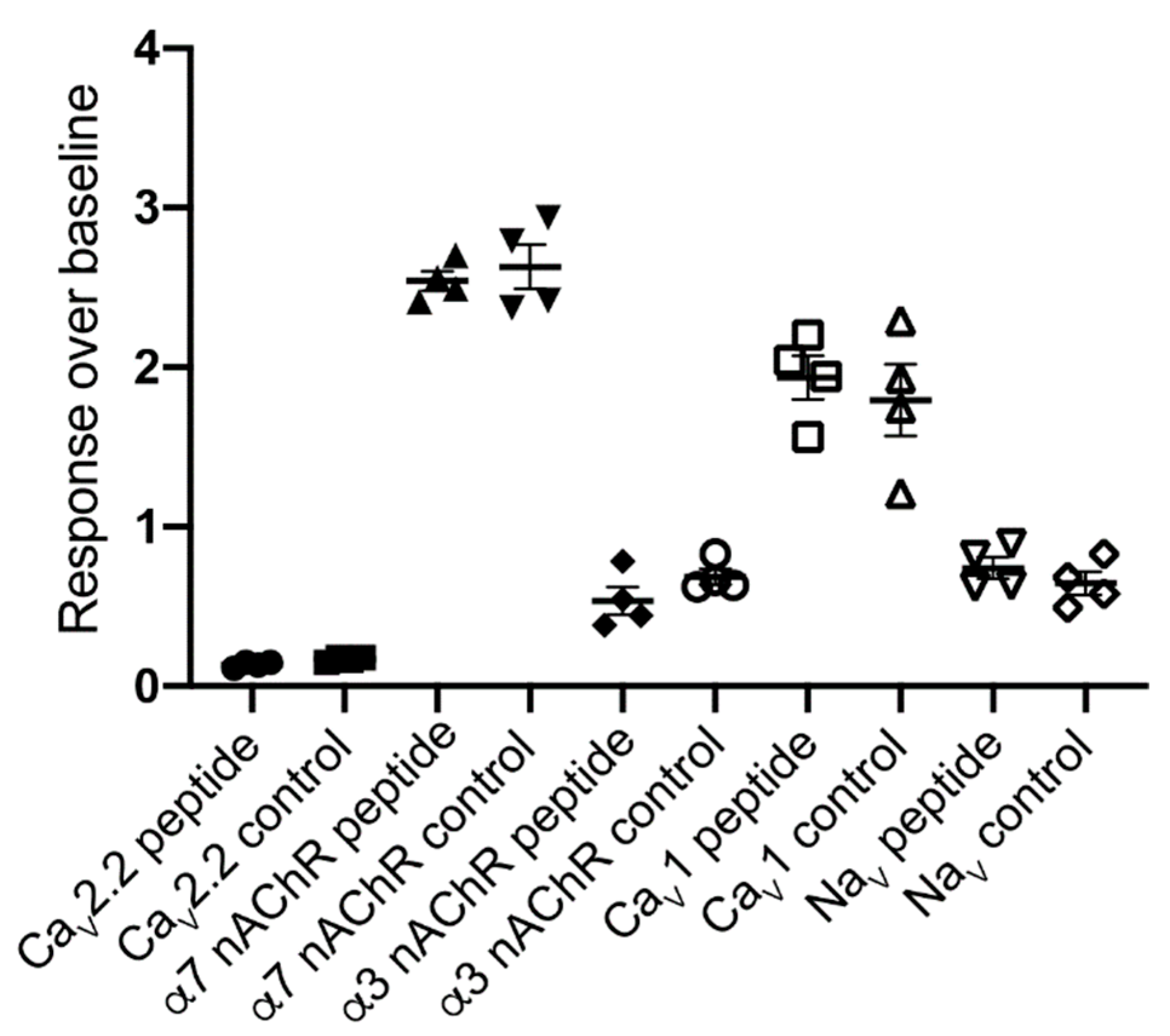
3.3. Electrophysiology
3.4. FLIPRTetra Ion Channel Assays
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mir, R.; Karim, S.; Kamal, M.A.; Wilson, C.; Mirza, Z. Conotoxins: Structure, Therapeutic Potential and Pharmacological Applications. Curr. Pharm. Des. 2016, 22, 582–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidtko, A.; Lotsch, J.; Freynhagen, R.; Geisslinger, G. Ziconotide for treatment of severe chronic pain. Lancet 2010, 375, 1569–1577. [Google Scholar] [CrossRef]
- Durek, T.; Craik, D.J. Therapeutic conotoxins: A US patent literature survey. Expert Opin. Ther. Pat. 2015, 25, 1159–1173. [Google Scholar] [CrossRef] [PubMed]
- Safavi-Hemami, H.; Brogan, S.E.; Olivera, B.M. Pain therapeutics from cone snail venoms: From Ziconotide to novel non-opioid pathways. J. Proteomics 2019, 190, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J. Discovery and development of the chi-conopeptide class of analgesic peptides. Toxicon 2012, 59, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.E.; White, H.S.; Wilcox, K.S. The effect of CGX-1007 and CI-1041, novel NMDA receptor antagonists, on NMDA receptor-mediated EPSCs. Epilepsy Res. 2004, 59, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Bernaldez-Sarabia, J.; Figueroa-Montiel, A.; Duenas, S.; Cervantes-Luevano, K.; Beltran, J.A.; Ortiz, E.; Jimenez, S.; Possani, L.D.; Paniagua-Solis, J.F.; Gonzalez-Canudas, J.; et al. The Diversified O-Superfamily in Californiconus californicus Presents a Conotoxin with Antimycobacterial Activity. Toxins 2019, 11, 128. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Liu, Y.Q.; Sun, Z.H.; Zhangsun, D.T.; Luo, S.L. Identification of nicotinic acetylcholine receptor subunits in different lung cancer cell lines and the inhibitory effect of alpha-conotoxin TxID on lung cancer cell growth. Eur. J. Pharm. 2019, 865, 172674. [Google Scholar] [CrossRef]
- Himaya, S.W.A.; Lewis, R.J. Venomics-accelerated cone snail venom peptide discovery. Int. J. Mol. Sci. 2018, 19, 788. [Google Scholar] [CrossRef] [Green Version]
- Jin, A.H.; Muttenthaler, M.; Dutertre, S.; Himaya, S.W.A.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Conotoxins: Chemistry and biology. Chem. Rev. 2019, 119, 11510–11549. [Google Scholar] [CrossRef]
- Robinson, S.D.; Norton, R.S. Conotoxin gene superfamilies. Mar. Drugs 2014, 12, 6058–6101. [Google Scholar] [CrossRef] [Green Version]
- Kaas, Q.; Westermann, J.C.; Halai, R.; Wang, C.K.; Craik, D.J. ConoServer, a database for conopeptide sequences and structures. Bioinformatics 2008, 24, 445–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaas, Q.; Yu, R.; Jin, A.H.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucleic Acids Res. 2012, 40, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Giribaldi, J.; Dutertre, S. Alpha-conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal nicotinic acetylcholine receptors. Neurosci. Lett. 2018, 679, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Azam, L.; McIntosh, J.M. Alpha-conotoxins as pharmacological probes of nicotinic acetylcholine receptors. Acta Pharm. Sin. 2009, 30, 771–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimohama, S.; Kawamata, J. Roles of nicotinic acetylcholine receptors in the pathology and treatment of Alzheimer’s and Parkinson’s diseases. In Nicotinic Acetylcholine Receptor Signaling in Neuroprotection; Akaike, A., Shimohama, S., Misu, Y., Eds.; Springer: Singapore, 2018; pp. 137–158. [Google Scholar] [CrossRef] [Green Version]
- Hurst, R.; Rollema, H.; Bertrand, D. Nicotinic acetylcholine receptors: From basic science to therapeutics. Pharm. Ther. 2013, 137, 22–54. [Google Scholar] [CrossRef]
- Clark, R.J.; Jensen, J.; Nevin, S.T.; Callaghan, B.P.; Adams, D.J.; Craik, D.J. The engineering of an orally active conotoxin for the treatment of neuropathic pain. Angew. Chem. Int. Ed. Engl. 2010, 49, 6545–6548. [Google Scholar] [CrossRef]
- Castro, J.; Grundy, L.; Deiteren, A.; Harrington, A.M.; O’Donnell, T.; Maddern, J.; Moore, J.; Garcia-Caraballo, S.; Rychkov, G.Y.; Yu, R.; et al. Cyclic analogues of alpha-conotoxin Vc1.1 inhibit colonic nociceptors and provide analgesia in a mouse model of chronic abdominal pain. Br. J. Pharm. 2018, 175, 2384–2398. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Seymour, V.A.; Berecki, G.; Jia, X.; Akcan, M.; Adams, D.J.; Kaas, Q.; Craik, D.J. Less is More: Design of a Highly Stable Disulfide-Deleted Mutant of Analgesic Cyclic alpha-Conotoxin Vc1.1. Sci. Rep. 2015, 5, 13264. [Google Scholar] [CrossRef] [Green Version]
- Sadeghi, M.; Carstens, B.B.; Callaghan, B.P.; Daniel, J.T.; Tae, H.S.; O’Donnell, T.; Castro, J.; Brierley, S.M.; Adams, D.J.; Craik, D.J.; et al. Structure-activity studies reveal the molecular basis for GABAB-Receptor mediated inhibition of high voltage-activated calcium channels by alpha-conotoxin Vc1.1. ACS Chem. Biol. 2018, 13, 1577–1587. [Google Scholar] [CrossRef] [Green Version]
- Marx, U.C.; Daly, N.L.; Craik, D.J. NMR of conotoxins: Structural features and an analysis of chemical shifts of post-translationally modified amino acids. Magn. Reson. Chem. 2006, 44, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.H.; Vetter, I.; Himaya, S.W.; Alewood, P.F.; Lewis, R.J.; Dutertre, S. Transcriptome and proteome of Conus planorbis identify the nicotinic receptors as primary target for the defensive venom. Proteomics 2015, 15, 4030–4040. [Google Scholar] [CrossRef] [PubMed]
- Skinner, S.P.; Fogh, R.H.; Boucher, W.; Ragan, T.J.; Mureddu, L.G.; Vuister, G.W. CcpNmr AnalysisAssign: A flexible platform for integrated NMR analysis. J. Biomol. NMR 2016, 66, 111–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wüthrich, K. NMR of Proteins and Nucleic Acids; Wiley-Interscience: New York, NY, USA, 1986. [Google Scholar]
- Wishart, D.S.; Bigam, C.G.; Holm, A.; Hodges, R.S.; Sykes, B.D. 1H, 13C and 15N random coil NMR chemical shifts of the common amino acids. I. Investigations of nearest-neighbor effects. J. Biomol. NMR 1995, 5, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Wurz, J.M.; Kazemi, S.; Schmidt, E.; Bagaria, A.; Guntert, P. NMR-based automated protein structure determination. Arch. Biochem. Biophys. 2017, 628, 24–32. [Google Scholar] [CrossRef]
- Shen, Y.; Bax, A. Protein backbone and sidechain torsion angles predicted from NMR chemical shifts using artificial neural networks. J. Biomol. NMR 2013, 56, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Koradi, R.; Billeter, M.; Wuthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14. [Google Scholar] [CrossRef]
- Beissner, M.; Dutertre, S.; Schemm, R.; Danker, T.; Sporning, A.; Grubmuller, H.; Nicke, A. Efficient binding of 4/7 alpha-conotoxins to nicotinic alpha4beta2 receptors is prevented by Arg185 and Pro195 in the alpha4 subunit. Mol. Pharm. 2012, 82, 711–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, A.; Starobova, H.; Inserra, M.C.; Jin, A.H.; Deuis, J.R.; Dutertre, S.; Lewis, R.J.; Alewood, P.F.; Daly, N.L.; Vetter, I. alpha-Conotoxin MrIC is a biased agonist at alpha7 nicotinic acetylcholine receptors. Biochem. Pharm. 2015, 94, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.R.; McArthur, J.R.; Brust, A.; Bhola, R.F.; Rosengren, K.J.; Ragnarsson, L.; Dutertre, S.; Alewood, P.F.; Christie, M.J.; Adams, D.J.; et al. Novel analgesic omega-conotoxins from the vermivorous cone snail Conus moncuri provide new insights into the evolution of conopeptides. Sci. Rep. 2018, 8, 13397. [Google Scholar] [CrossRef]
- Vetter, I.; Mozar, C.A.; Durek, T.; Wingerd, J.S.; Alewood, P.F.; Christie, M.J.; Lewis, R.J. Characterisation of Na(v) types endogenously expressed in human SH-SY5Y neuroblastoma cells. Biochem. Pharm. 2012, 83, 1562–1571. [Google Scholar] [CrossRef]
- Vetter, I.; Lewis, R.J. Characterization of endogenous calcium responses in neuronal cell lines. Biochem. Pharm. 2010, 79, 908–920. [Google Scholar] [CrossRef] [PubMed]
- El Hamdaoui, Y.; Wu, X.; Clark, R.J.; Giribaldi, J.; Anangi, R.; Craik, D.J.; King, G.F.; Dutertre, S.; Kaas, Q.; Herzig, V.; et al. Periplasmic expression of 4/7 alpha-Conotoxin TxIA analogs in E. coli favors ribbon isomer formation—Suggestion of a binding bode at the alpha7 nAChR. Front. Pharm. 2019, 10, 577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, R.J.; Fischer, H.; Nevin, S.T.; Adams, D.J.; Craik, D.J. The synthesis, structural characterization, and receptor specificity of the alpha-conotoxin Vc1.1. J. Biol. Chem. 2006, 281, 23254–23263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.M.; Oomen, C.J.; Miranda, L.P.; Bingham, J.P.; Alewood, P.F.; Craik, D.J. Three-dimensional solution structure of alpha-conotoxin MII by NMR spectroscopy: Effects of solution environment on helicity. Biochemistry 1998, 37, 15621–15630. [Google Scholar] [CrossRef] [PubMed]
- Peigneur, S.; Devi, P.; Seldeslachts, A.; Ravichandran, S.; Quinton, L.; Tytgat, J. Structure-function elucidation of a new alpha-conotoxin, MilIA, from Conus milneedwardsi. Mar. Drugs 2019, 17, 535. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Connectivity | Fold | Target Function 1 |
|---|---|---|
| Cys6-Cys12, Cys7-Cys21 | Globular | 0.046 ± 0.037 |
| Cys6-Cys21, Cys7-Cys12 | Ribbon | 1.23 ± 0.099 |
| Cys6-Cys7, Cys12-Cys21 | Beads | 3.3 ± 0.090 |
| Experimental Restraints | |
|---|---|
| Interproton distance restraints | |
| Intraresidue | 57 |
| Sequential | 50 |
| Medium range (i–j < 5) | 17 |
| Long range (i–j ≥ 5) | 8 |
| Total | 132 |
| Dihedral-angle restraints | 30 |
| Hydrogen bonds (2 per bond) | 12 |
| R.m.s. deviations from mean coordinate structure (Å) (in residues 6–21) | |
| Backbone atoms | 0.44 +/− 0.14 |
| All heavy atoms | 1.51 +/− 0.25 |
| Ramachandran Statistics * | |
| % in most favoured region | 77.8 |
| % in additionally allowed region | 22.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, D.T.; Bansal, P.S.; Carter, D.A.; Vetter, I.; Nicke, A.; Dutertre, S.; Daly, N.L. Characterisation of a Novel A-Superfamily Conotoxin. Biomedicines 2020, 8, 128. https://doi.org/10.3390/biomedicines8050128
Wilson DT, Bansal PS, Carter DA, Vetter I, Nicke A, Dutertre S, Daly NL. Characterisation of a Novel A-Superfamily Conotoxin. Biomedicines. 2020; 8(5):128. https://doi.org/10.3390/biomedicines8050128
Chicago/Turabian StyleWilson, David T., Paramjit S. Bansal, David A. Carter, Irina Vetter, Annette Nicke, Sébastien Dutertre, and Norelle L. Daly. 2020. "Characterisation of a Novel A-Superfamily Conotoxin" Biomedicines 8, no. 5: 128. https://doi.org/10.3390/biomedicines8050128