Scorpion Venom: Detriments and Benefits

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Scorpion Venom Compounds
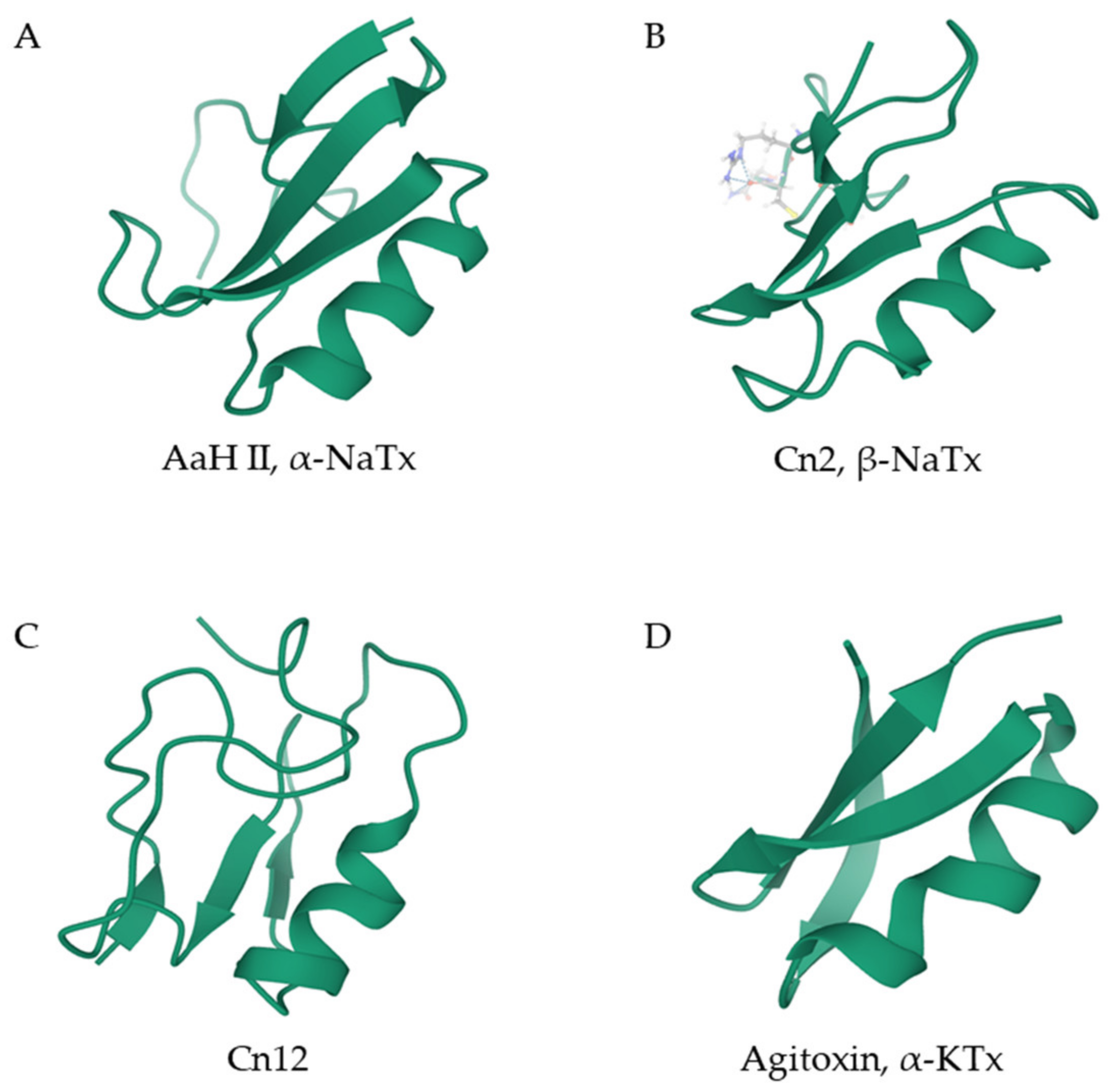
2.1. Peptides Containing CS α/β Motifs
2.1.1. Long Scorpion Toxins
2.1.2. Short Scorpion Toxins
2.2. Calcins
2.3. Non-Disulfide Bridged Peptides (NDBPs)
2.4. Enzymes
2.5. Other Venom Compounds
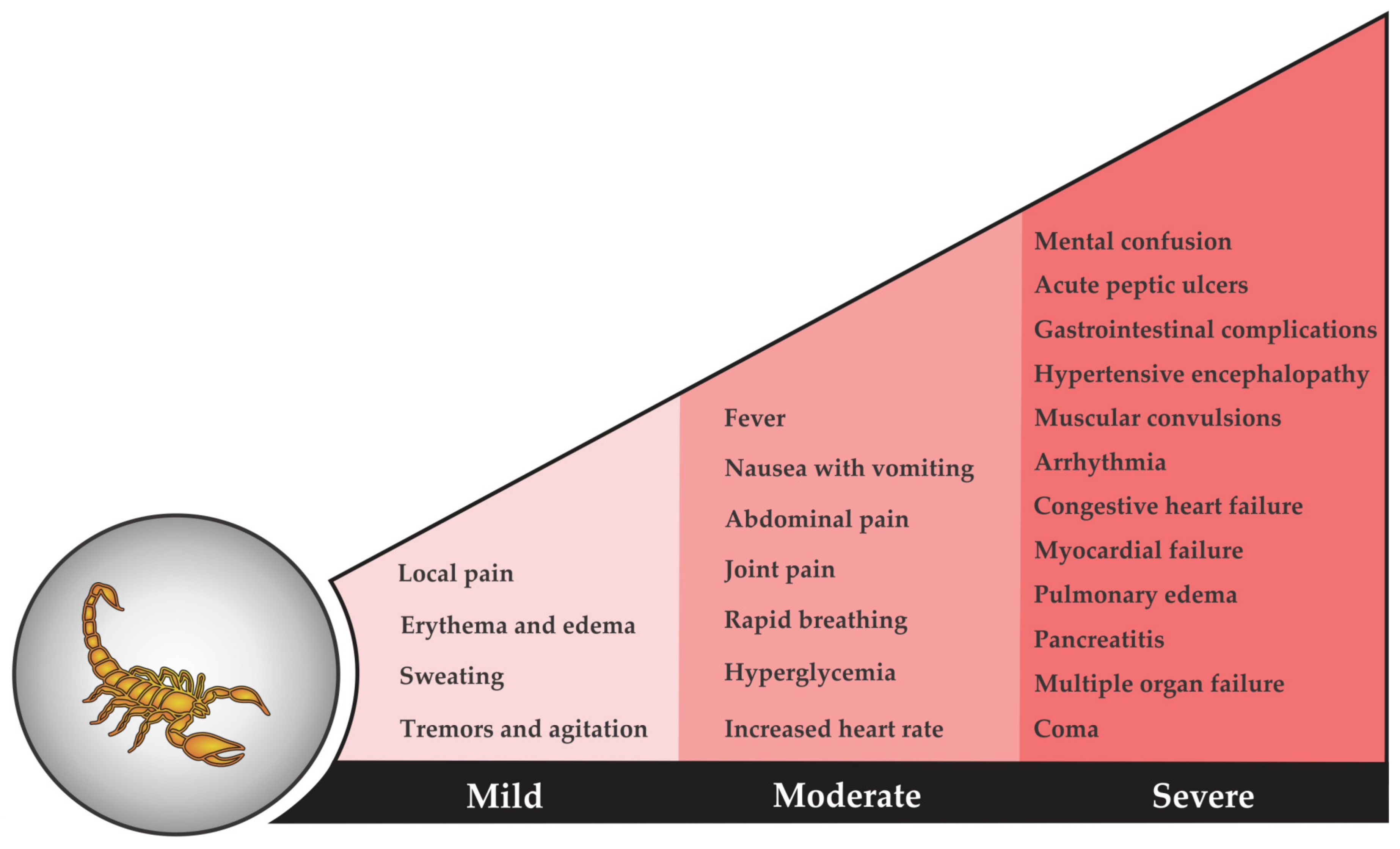
3. Detriments of Scorpion Venom: Scorpion Envenoming
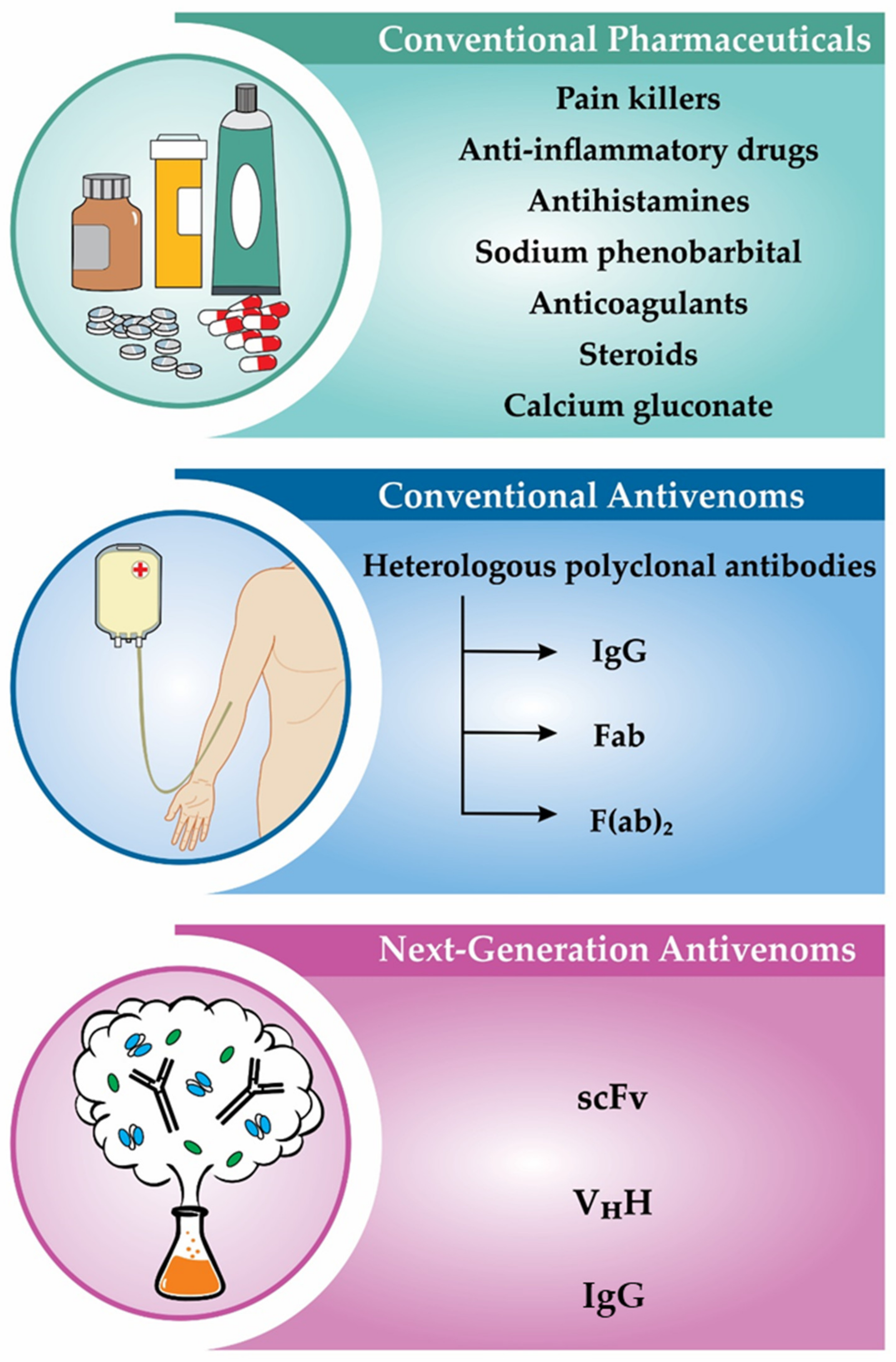
3.1. Scorpion Envenoming Treatment
3.1.1. Conventional Plasma-Derived Antivenoms
3.1.2. Recombinant Antivenoms
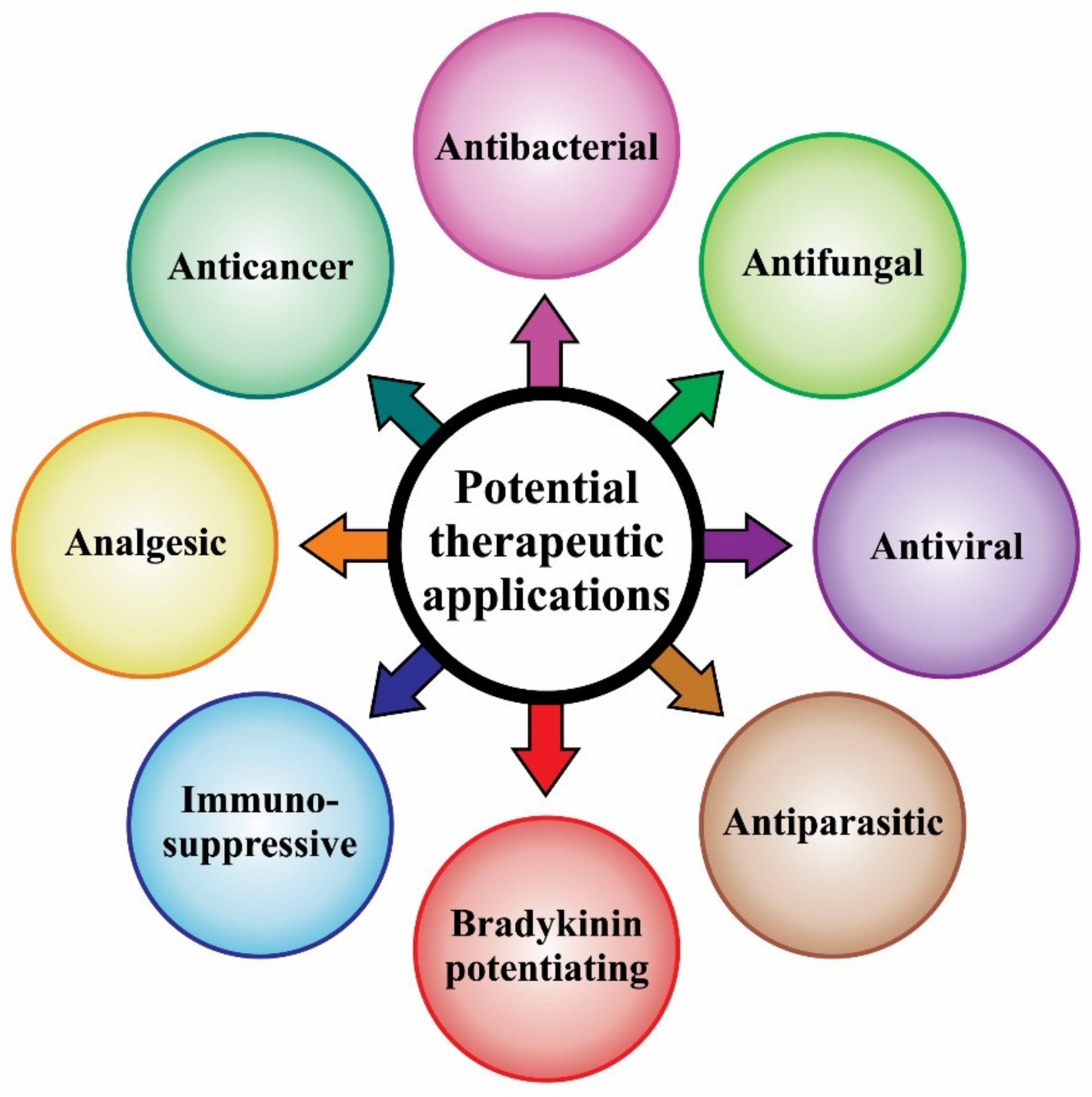
4. Benefits of Scorpion Venom: Ongoing Research on Scorpion Toxins with Potential Therapeutic Applications
4.1. Antibacterial Effects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Scorpion Species | Antibacterial Agent | MW (S–S Bridge) | Target | Reference |
|---|---|---|---|---|---|
| 2015 | A. aeneas | AaeAP1 | 2016.18 Da (0) | S. aureus | [96] |
| AaeAP2 | 1986.15 Da (0) | ||||
| 2015 | C. margaritatus | Cm38 | 2149 Da (2) | Klebsiella pneumonia | [97] |
| 2015 | T. stigmurus | Stigmurin | 1795.22 Da (0) | Gram-positive bacteria including S. aureus and Methicillin-resistant S. aureus (MRSA) | [98] |
| 2016 | M. gibbosus | Low molecular mass chitosan * | 3220 Da (0) | Bacterial species in general, including Listeria monocytogenes, Bacillus subtilis, Salmonella enteritidis, and the yeast Candida albicans | [99] |
| 2016 | Scorpio maurus palmatus (synthetic) | Smp-24 | 2578 Da (0) | Highest activity against Gram-positive bacteria, limited activity against C. albicans | [100] |
| Smp-43 | 4654.3 Da (0) | ||||
| 2017 | Isometrus maculatus | Im-4 | 1714 Da (0) | Gram-positive bacteria | [101] |
| Im-5 | 2803.7 Da (0) | Gram-positive and Gram-negative bacteria | |||
| Im-6 | 1707 Da (0) | Gram-positive bacteria | |||
| 2018 | T. obscurus | ToAP2 | 9486 Da (0) | Mycobacterium massiliense | [102] |
| 2018 | M. eupeus | Meucin-49 | 5574.93 Da (?) | Gram-positive bacteria | [103] |
| Meucin-18 | 2107.13 Da (0) | Gram-negative bacteria | |||
| 2018 | M. martensii | Marcin-18 | 2135.63 Da (0?) | Gram-positive bacteria, including some clinical antibiotic-resistant strains | [104] |
| M. gibbosus | Megicin-18 | 2068.04 Da (?) | |||
| M. eupeus | Meucin-18 | 2107.13 Da (0) | |||
| 2018 | Liocheles australasiae | LaIT2 | 6628.2 Da (3) | Gram-negative bacteria | [105] |
| N-LaIT2 | 3326 Da (?) | ||||
| 2018 | T. stigmurus | StigA6 | 1908 Da (0?) | Gram-positive and Gram-negative bacteria | [106] |
| StigA16 | 1949 Da (0?) | ||||
| 2019 | D. melici | Red 1,4-benzoquinone: 3,5-dimethoxy-2-(methylthio) | 168.15 Da | S. aureus | [53] |
| cyclohexa-2,5-diene-1,4-dione * | M. tuberculosis, including an MDR strain | ||||
| Blue 1,4-benzoquinone: 5-methoxy-2,3-bis(methylthio)cyclohexa-2,5-diene-1,4-dione * | |||||
| 2017 | U. yaschenkoi | UyCT1 | 1603.9 Da (0) | Gram-positive and Gram-negative bacteria | [107] |
| UyCT3 | 1433.7 Da (0) | ||||
| UyCT5 | 1442.7 Da (0) | ||||
| Uy17 | 1369.43 Da (0) | ||||
| Uy192 | 1459.98 Da (0) | ||||
| Uy234 | 1986.19 Da (0) | ||||
| 2019 | U. yaschenkoi | Uy234 | 1986.19 Da (0) | MDR bacteria, including β-hemolytic Streptococcus strains | [108] |
| Uy17 | 1369.43 Da (0) | ||||
| Uy192 | 1459.98 Da (0) | ||||
| 2017 | U. manicatus | Um2 | 2034.56 Da (?) | Gram-positive and Gram-negative bacteria | [107] |
| Um3 | 1577.23 Da (?) | ||||
| Um4 | 1428.58 Da (?) | ||||
| Um5 | 1508.82 Da (?) | ||||
| 2019 | T. serrulatus (hemolymph) | Serrulin | 3564 Da (?) | Gram-positive and Gram-negative bacteria | [109] |
| 2019 | A. australis hector | G-TI | 7390 Da (4, predicted) | B. cereus | [110] |
4.2. Antifungal Effects
4.3. Antiviral Effects
4.4. Antiparasitic Effects
4.5. Bradykinin-Potentiating Effects
4.6. Immunosuppressive Effects
4.7. Analgesic Effects
| Toxin Name | Scorpion Species | MW (S–S Bridge) | Target | Reference |
|---|---|---|---|---|
| BmK AS | M. martensii | 7701 Da (4) | TTX-R (Nav1.8, 1.9), TTX-S (Nav1.3); reduction of neural excitability; skeletal muscle RyR | [192,193,194,195] |
| BmK IT2 | M. martensii | 6650 Da (4) | TTX-R and TTX-S Nav | [192,196] |
| BmK IT-AP | M. martensii | 8157 Da (4) | N/A | [197] |
| BmK dITAP3 | M. martensii | 6740 Da (4) | N/A | [187] |
| BmK AEP/BmK ANEP | M. martensii | 6738 Da (4) | Nav1.1, Nav1.3, Nav1.6, Nav1.7 | [198,199,200] |
| BmK AS1 | M. martensii | 7712 Da (4) | TTX-R and TTX-S Nav, skeletal-muscle RyR-1 | [201] |
| BmK AGAP | M. martensii | 7281 Da (4) | Prevention of peripheral and spinal MAPKs expression; Decrease of spinal c-Fos expression; Nav1.7, Nav1.8, Nav1.4, Nav1.5; Cav | [185,202,203,204,205] |
| BmK Ang P1 | M. martensii | 8141 Da (4) | N/A | [206] |
| BmK Ang M1 | M. martensii | 7040 Da (4) | Nav, Kv | [114,207] |
| BmK(M)9 | M. martensii | 7106 Da (4) | Nav1.4, Nav1.5, Nav1.7 | [208,209] |
| BmK AGP-SYPU1 | M. martensii | 7227 Da (4) | N/A | [210] |
| BmK AGP-SYPU2 | M. martensii | 7457 Da (4) | Nav | [211,212] |
| BmK AGAP-SYPU2 | M. martensii | 7253 Da (4) | Nav (suspected) | [213] |
| BmK-YA | M. martensii | 871 Da (0) | µ, κ, δ-opioid receptor | [188] |
| BmKBTx | M. martensii | 6800 Da (3) | Nav1.7 | [214] |
| BmNaL-3SS2 | M. martensii | 7338.26 Da (3) | Nav1.7 | [214] |
| AmmVIII | A. mauretanicus mauretanicus | 7383 Da (4) | Nav1.2, endogenous opioid system, no data on other Navs yet | [186] |
| LqqIT2 | L. quinquestriatus quinquestriatus | 6845 Da (4) | Endogenous opioid system, no data on other Navs yet | [186] |
| TsNTxP | T. serrulatus | 6702 (4) | N/A (possibly glutamate release) | [191] |
| BotAF | B. occitanus tunetanus | 7446 Da (4) | Peripheral or spinal mechanisms | [198] |
| Hetlaxin | H. laoticus | 3665 Da (4) | Kv1.3, Kv1.1 | [184,215] |
4.8. Anticancer Effects
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chippaux, J.-P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef] [PubMed]
- WHO. Report of the Eleventh Meeting of the WHO Strategic and Technical Advisory Group for Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2018; pp. 1–28. [Google Scholar] [CrossRef]
- Chippaux, J.-P. Emerging options for the management of scorpion stings. Drug Des. Dev. Ther. 2012, 6, 165–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reckziegel, G.C.; Pinto, V.L.; Reckziegel, G.C.; Pinto, V.L. Scorpionism in Brazil in the years 2000 to 2012. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laustsen, A.H.; Solà, M.; Jappe, E.C.; Oscoz, S.; Lauridsen, L.P.; Engmark, M. Biotechnological Trends in Spider and Scorpion Antivenom Development. Toxins 2016, 8, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, W.R. The evolution and distribution of noxious species of scorpions (Arachnida: Scorpiones). J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24. [Google Scholar] [CrossRef] [Green Version]
- Hauke, T.J.; Herzig, V. Dangerous arachnids—Fake news or reality? Toxicon 2017, 138, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Mullen, G.; Durden, L. Medical and Veterinary Entomology. Ann. Trop. Med. Parasitol. 2019, 3. [Google Scholar] [CrossRef] [Green Version]
- Ward, M.J.; Ellsworth, S.A.; Nystrom, G.S. A global accounting of medically significant scorpions: Epidemiology, major toxins, and comparative resources in harmless counterparts. Toxicon 2018, 151, 137–155. [Google Scholar] [CrossRef]
- Petricevich, V.L. Scorpion Venom and the Inflammatory Response. Available online: https://www.hindawi.com/journals/mi/2010/903295/ (accessed on 12 January 2020).
- De la Vega, R.C.R.; Vidal, N.; Possani, L.D. Chapter 59—Scorpion Peptides. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 423–429. ISBN 978-0-12-385095-9. [Google Scholar]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef]
- Valdivia, H.H.; Martin, B.M.; Ramírez, A.N.; Fletcher, P.L.; Possani, L.D. Isolation and pharmacological characterization of four novel Na+ channel-blocking toxins from the scorpion Centruroides noxius Hoffmann. J. Biochem. 1994, 116, 1383–1391. [Google Scholar] [CrossRef]
- Goyffon, M.; Tournier, J.-N. Scorpions: A Presentation. Toxins 2014, 6, 2137–2148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caliskan, F. Scorpion Venom Research Around the World: Turkish Scorpions. In Toxinology: Scorpion Venoms; Gopalakrishnakone, P., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 1–19. ISBN 978-94-007-6647-1. [Google Scholar]
- Del Río-Portilla, F.; Hernández-Marín, E.; Pimienta, G.; Coronas, F.V.; Zamudio, F.Z.; Rodríguez de la Vega, R.C.; Wanke, E.; Possani, L.D. NMR solution structure of Cn12, a novel peptide from the Mexican scorpion Centruroides noxius with a typical beta-toxin sequence but with alpha-like physiological activity. Eur. J. Biochem. 2004, 271, 2504–2516. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez de la Vega, R.C.; Possani, L.D. Current views on scorpion toxins specific for K+-channels. Toxicon 2004, 43, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Tytgat, J.; Chandy, K.G.; Garcia, M.L.; Gutman, G.A.; Martin-Eauclaire, M.F.; van der Walt, J.J.; Possani, L.D. A unified nomenclature for short-chain peptides isolated from scorpion venoms: Alpha-KTx molecular subfamilies. Trends Pharm. Sci. 1999, 20, 444–447. [Google Scholar] [CrossRef]
- Cremonez, C.M.; Maiti, M.; Peigneur, S.; Cassoli, J.S.; Dutra, A.A.A.; Waelkens, E.; Lescrinier, E.; Herdewijn, P.; de Lima, M.E.; Pimenta, A.M.C.; et al. Structural and Functional Elucidation of Peptide Ts11 Shows Evidence of a Novel Subfamily of Scorpion Venom Toxins. Toxins 2016, 8, 288. [Google Scholar] [CrossRef] [Green Version]
- Pucca, M.B.; Peigneur, S.; Cologna, C.T.; Cerni, F.A.; Zoccal, K.F.; Bordon, K.d.C.F.; Faccioli, L.H.; Tytgat, J.; Arantes, E.C. Electrophysiological characterization of the first Tityus serrulatus alpha-like toxin, Ts5: Evidence of a pro-inflammatory toxin on macrophages. Biochimie 2015, 115, 8–16. [Google Scholar] [CrossRef]
- Cerni, F.A.; Pucca, M.B.; Amorim, F.G.; de Castro Figueiredo Bordon, K.; Echterbille, J.; Quinton, L.; De Pauw, E.; Peigneur, S.; Tytgat, J.; Arantes, E.C. Isolation and characterization of Ts19 Fragment II, a new long-chain potassium channel toxin from Tityus serrulatus venom. Peptides 2016, 80, 9–17. [Google Scholar] [CrossRef]
- Jiménez-Vargas, J.M.; Restano-Cassulini, R.; Possani, L.D. Toxin modulators and blockers of hERG K(+) channels. Toxicon 2012, 60, 492–501. [Google Scholar] [CrossRef]
- Srinivasan, K.N.; Sivaraja, V.; Huys, I.; Sasaki, T.; Cheng, B.; Kumar, T.K.S.; Sato, K.; Tytgat, J.; Yu, C.; San, B.C.C.; et al. kappa-Hefutoxin1, a novel toxin from the scorpion Heterometrus fulvipes with unique structure and function. Importance of the functional diad in potassium channel selectivity. J. Biol. Chem. 2002, 277, 30040–30047. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Luo, F.; Feng, J.; Yang, W.; Zeng, D.; Zhao, R.; Cao, Z.; Liu, M.; Li, W.; Jiang, L.; et al. Genomic and Structural Characterization of Kunitz-Type Peptide LmKTT-1a Highlights Diversity and Evolution of Scorpion Potassium Channel Toxins. PLoS ONE 2013, 8, e60201. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.J.; Hill, J.M.; Little, M.J.; Nicholson, G.M.; King, G.F.; Alewood, P.F. Unique scorpion toxin with a putative ancestral fold provides insight into evolution of the inhibitor cystine knot motif. Proc. Natl. Acad. Sci. USA 2011, 108, 10478–10483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.; Harvey, P.J.; Craik, D.J.; Ronjat, M.; De Waard, M.; Zhu, S. Functional evolution of scorpion venom peptides with an inhibitor cystine knot fold. Biosci. Rep. 2013, 33, e00047. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Gurrola, G.B.; Zhang, J.; Valdivia, C.R.; SanMartin, M.; Zamudio, F.Z.; Zhang, L.; Possani, L.D.; Valdivia, H.H. Structure–function relationships of peptides forming the calcin family of ryanodine receptor ligands. J. Gen. Physiol. 2016, 147, 375–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Animal Toxins and Ion Channels—Abstract—Europe PMC. Available online: https://europepmc.org/article/med/10783702 (accessed on 12 January 2020).
- Lanner, J.T.; Georgiou, D.K.; Joshi, A.D.; Hamilton, S.L. Ryanodine Receptors: Structure, Expression, Molecular Details, and Function in Calcium Release. Cold Spring Harb. Perspect. Biol. 2010, 2, a003996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bers, D. Excitation-Contraction Coupling and Cardiac Contractile Force, 2nd ed.; Developments in Cardiovascular Medicine; Springer: Amsterdam, The Netherlands, 2001; ISBN 978-0-7923-7157-1. [Google Scholar]
- Smith, J.J.; Vetter, I.; Lewis, R.J.; Peigneur, S.; Tytgat, J.; Lam, A.; Gallant, E.M.; Beard, N.A.; Alewood, P.F.; Dulhunty, A.F. Multiple actions of φ-LITX-Lw1a on ryanodine receptors reveal a functional link between scorpion DDH and ICK toxins. Proc. Natl. Acad. Sci. USA 2013, 110, 8906–8911. [Google Scholar] [CrossRef] [Green Version]
- Boisseau, S.; Mabrouk, K.; Ram, N.; Garmy, N.; Collin, V.; Tadmouri, A.; Mikati, M.; Sabatier, J.-M.; Ronjat, M.; Fantini, J.; et al. Cell penetration properties of maurocalcine, a natural venom peptide active on the intracellular ryanodine receptor. Biochim. Biophys. Acta (BBA) Biomembr. 2006, 1758, 308–319. [Google Scholar] [CrossRef]
- Ram, N.; Weiss, N.; Texier-Nogues, I.; Aroui, S.; Andreotti, N.; Pirollet, F.; Ronjat, M.; Sabatier, J.-M.; Darbon, H.; Jacquemond, V.; et al. Design of a disulfide-less, pharmacologically inert, and chemically competent analog of maurocalcine for the efficient transport of impermeant compounds into cells. J. Biol. Chem. 2008, 283, 27048–27056. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira-Mendes, B.B.R.; Horta, C.C.R.; do Carmo, A.O.; Biscoto, G.L.; Sales-Medina, D.F.; Leal, H.G.; Brandão-Dias, P.F.P.; Miranda, S.E.M.; Aguiar, C.J.; Cardoso, V.N.; et al. CPP-Ts: A new intracellular calcium channel modulator and a promising tool for drug delivery in cancer cells. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- King, J.V.L.; Emrick, J.J.; Kelly, M.J.S.; Herzig, V.; King, G.F.; Medzihradszky, K.F.; Julius, D. A Cell-Penetrating Scorpion Toxin Enables Mode-Specific Modulation of TRPA1 and Pain. Cell 2019, 178, 1362–1374.e16. [Google Scholar] [CrossRef] [PubMed]
- Almaaytah, A.; Albalas, Q. Scorpion venom peptides with no disulfide bridges: A review. Peptides 2014, 51, 35–45. [Google Scholar] [CrossRef]
- Quintero-Hernández, V.; Ramírez-Carreto, S.; Romero-Gutiérrez, M.T.; Valdez-Velázquez, L.L.; Becerril, B.; Possani, L.D.; Ortiz, E. Transcriptome Analysis of Scorpion Species Belonging to the Vaejovis Genus. PLoS ONE 2015, 10, e0117188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.-C.; Corzo, G.; Hahin, R. Scorpion Venom Peptides without Disulfide Bridges. IUBMB Life 2005, 57, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Pinheiro-Junior, E.L.; Zoccal, K.F.; Bordon, K.d.C.F.; Amorim, F.G.; Peigneur, S.; Vriens, K.; Thevissen, K.; Cammue, B.P.A.; et al. Non-disulfide-bridged peptides from Tityus serrulatus venom: Evidence for proline-free ACE-inhibitors. Peptides 2016, 82, 44–51. [Google Scholar] [CrossRef]
- Bordon, K.C.F.; Wiezel, G.A.; Amorim, F.G.; Arantes, E.C. Arthropod venom Hyaluronidases: Biochemical properties and potential applications in medicine and biotechnology. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girish, K.S.; Kemparaju, K. The magic glue hyaluronan and its eraser hyaluronidase: A biological overview. Life Sci. 2007, 80, 1921–1943. [Google Scholar] [CrossRef]
- De Oliveira-Mendes, B.B.R.; Miranda, S.E.M.; Sales-Medina, D.F.; de Magalhães, B.F.; Kalapothakis, Y.; de Souza, R.P.; Cardoso, V.N.; de Barros, A.L.B.; Guerra-Duarte, C.; Kalapothakis, E.; et al. Inhibition of Tityus serrulatus venom hyaluronidase affects venom biodistribution. PLoS Negl. Trop. Dis. 2019, 13, e0007048. [Google Scholar] [CrossRef]
- Schwartz, E.F.; Camargos, T.S.; Zamudio, F.Z.; Silva, L.P.; Bloch, C.; Caixeta, F.; Schwartz, C.A.; Possani, L.D. Mass spectrometry analysis, amino acid sequence and biological activity of venom components from the Brazilian scorpion Opisthacanthus cayaporum. Toxicon 2008, 51, 1499–1508. [Google Scholar] [CrossRef]
- Incamnoi, P.; Patramanon, R.; Thammasirirak, S.; Chaveerach, A.; Uawonggul, N.; Sukprasert, S.; Rungsa, P.; Daduang, J.; Daduang, S. Heteromtoxin (HmTx), a novel heterodimeric phospholipase A2 from Heterometrus laoticus scorpion venom. Toxicon 2013, 61, 62–71. [Google Scholar] [CrossRef]
- Fletcher, P.L.; Fletcher, M.D.; Weninger, K.; Anderson, T.E.; Martin, B.M. Vesicle-associated Membrane Protein (VAMP) Cleavage by a New Metalloprotease from the Brazilian Scorpion Tityus serrulatus. J. Biol. Chem. 2010, 285, 7405–7416. [Google Scholar] [CrossRef] [Green Version]
- Brazón, J.; Guerrero, B.; D’Suze, G.; Sevcik, C.; Arocha-Piñango, C.L. Fibrin(ogen)olytic enzymes in scorpion (Tityus discrepans) venom. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 168, 62–69. [Google Scholar] [CrossRef]
- Kazemi, S.M.; Sabatier, J.-M. Venoms of Iranian Scorpions (Arachnida, Scorpiones) and Their Potential for Drug Discovery. Molecules 2019, 24, 2670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, F.M.; Pimenta, A.M.C.; De Figueiredo, S.G.; Santoro, M.M.; Martin-Eauclaire, M.F.; Diniz, C.R.; De Lima, M.E. Enzymes with gelatinolytic activity can be found in Tityus bahiensis and Tityus serrulatus venoms. Toxicon 2002, 40, 1041–1045. [Google Scholar] [CrossRef]
- Cordeiro, F.A.; Coutinho, B.M.; Wiezel, G.A.; de Bordon, K.C.F.; Bregge-Silva, C.; Rosa-Garzon, N.G.; Cabral, H.; Ueberheide, B.; Arantes, E.C.; Cordeiro, F.A.; et al. Purification and enzymatic characterization of a novel metalloprotease from Lachesis muta rhombeata snake venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 32. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A. Snake venom metalloproteinases: Their role in the pathogenesis of local tissue damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Mandal, M.B.; Deshpande, S.B. Role of serotonergic mechanism in gastric contractions induced by Indian Red Scorpion (Mesobuthus tamulus) venom. Indian J. Pharm. 2009, 41, 255–257. [Google Scholar] [CrossRef] [Green Version]
- Al-Asmari, A.K.; Kunnathodi, F.; Al Saadon, K.; Idris, M.M. Elemental analysis of scorpion venoms. J. Venom Res. 2016, 7, 16–20. [Google Scholar] [PubMed]
- Carcamo-Noriega, E.N.; Sathyamoorthi, S.; Banerjee, S.; Gnanamani, E.; Mendoza-Trujillo, M.; Mata-Espinosa, D.; Hernández-Pando, R.; Veytia-Bucheli, J.I.; Possani, L.D.; Zare, R.N. 1,4-Benzoquinone antimicrobial agents against Staphylococcus aureus and Mycobacterium tuberculosis derived from scorpion venom. Proc. Natl. Acad. Sci. USA 2019, 116, 12642–12647. [Google Scholar] [CrossRef] [Green Version]
- Thien, T.V.; Anh, H.N.; Trang, N.T.T.; Trung, P.V.; Khoa, N.C.; Osipov, A.V.; Dubovskii, P.V.; Ivanov, I.A.; Arseniev, A.S.; Tsetlin, V.I.; et al. Low-molecular-weight compounds with anticoagulant activity from the scorpion Heterometrus laoticus venom. Dokl. Biochem. Biophys. 2017, 476, 316–319. [Google Scholar] [CrossRef]
- Tran, T.V.; Hoang, A.N.; Nguyen, T.T.T.; Phung, T.V.; Nguyen, K.C.; Osipov, A.V.; Ivanov, I.A.; Tsetlin, V.I.; Utkin, Y.N. Anticoagulant Activity of Low-Molecular Weight Compounds from Heterometrus laoticus Scorpion Venom. Toxins 2017, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Cupo, P.; Cupo, P. Clinical update on scorpion envenoming. Rev. Soc. Bras. Med. Trop. 2015, 48, 642–649. [Google Scholar] [CrossRef] [Green Version]
- Pucca, M.B.; Cerni, F.A.; Pinheiro Junior, E.L.; Bordon, K.d.C.F.; Amorim, F.G.; Cordeiro, F.A.; Longhim, H.T.; Cremonez, C.M.; Oliveira, G.H.; Arantes, E.C. Tityus serrulatus venom—A lethal cocktail. Toxicon 2015, 108, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Zoccal, K.F.; Sorgi, C.A.; Hori, J.I.; Paula-Silva, F.W.G.; Arantes, E.C.; Serezani, C.H.; Zamboni, D.S.; Faccioli, L.H. Opposing roles of LTB4 and PGE2 in regulating the inflammasome-dependent scorpion venom-induced mortality. Nat. Commun. 2016, 7, 10760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksel, G.; Güler, S.; Doğan, N.Ö.; Çorbacioğlu, Ş.K. A randomized trial comparing intravenous paracetamol, topical lidocaine, and ice application for treatment of pain associated with scorpion stings. Hum. Exp. Toxicol. 2015, 34, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V. Prazosin: A Pharmacological Antidote for Scorpion Envenomation. J. Trop. Pediatr. 2006, 52, 150–151. [Google Scholar] [CrossRef]
- Al Abri, S.; Al Rumhi, M.; Al Mahruqi, G.; Shakir, A.S. Scorpion Sting Management at Tertiary and Secondary Care Emergency Departments. Oman Med. J. 2019, 34, 9–13. [Google Scholar] [CrossRef]
- Mesquita, M.B.S.; Moraes-Santos, T.; Moraes, M.F.D. Phenobarbital blocks the lung edema induced by centrally injected tityustoxin in adult Wistar rats. Neurosci. Lett. 2002, 332, 119–122. [Google Scholar] [CrossRef]
- Ahmed, H.O.; Ranj, A.H. Clinical-demographic aspects of scorpion sting in al sulaimaneyah province: How frequent is hypocalcaemia in the victims? Eur. Sci. J. 2013, 9, 276–288. [Google Scholar]
- Rodrigo, C.; Gnanathasan, A. Management of scorpion envenoming: A systematic review and meta-analysis of controlled clinical trials. Syst. Rev. 2017, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- León, G.; Segura, A.; Herrera, M.; Otero, R.; França, F.O.d.S.; Barbaro, K.C.; Cardoso, J.L.C.; Wen, F.H.; de Medeiros, C.R.; Prado, J.C.L.; et al. Human heterophilic antibodies against equine immunoglobulins: Assessment of their role in the early adverse reactions to antivenom administration. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 1115–1119. [Google Scholar] [CrossRef] [Green Version]
- Laustsen, A. Toxin-centric development approach for next-generation antivenoms. Toxicon 2018, 150, 195–197. [Google Scholar] [CrossRef] [Green Version]
- Laustsen, A.H.; María Gutiérrez, J.; Knudsen, C.; Johansen, K.H.; Bermúdez-Méndez, E.; Cerni, F.A.; Jürgensen, J.A.; Ledsgaard, L.; Martos-Esteban, A.; Øhlenschlæger, M.; et al. Pros and cons of different therapeutic antibody formats for recombinant antivenom development. Toxicon 2018, 146, 151–175. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Engmark, M.; Clouser, C.; Timberlake, S.; Vigneault, F.; Gutiérrez, J.M.; Lomonte, B. Exploration of immunoglobulin transcriptomes from mice immunized with three-finger toxins and phospholipases A2 from the Central American coral snake, Micrurus nigrocinctus. PeerJ 2017, 5, e2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pucca, M.B.; Cerni, F.A.; Janke, R.; Bermúdez-Méndez, E.; Ledsgaard, L.; Barbosa, J.E.; Laustsen, A.H. History of Envenoming Therapy and Current Perspectives. Front. Immunol. 2019, 10, 1598. [Google Scholar] [CrossRef] [PubMed]
- Sifi, A.; Adi-Bessalem, S.; Laraba-Djebari, F. Development of a new approach of immunotherapy against scorpion envenoming: Avian IgYs an alternative to equine IgGs. Int. Immunopharmacol. 2018, 61, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Navarro, D.; Vargas, M.; Herrera, M.; Segura, Á.; Gómez, A.; Villalta, M.; Ramírez, N.; Williams, D.; Gutiérrez, J.M.; León, G. Development of a chicken-derived antivenom against the taipan snake (Oxyuranus scutellatus) venom and comparison with an equine antivenom. Toxicon 2016, 120, 1–8. [Google Scholar] [CrossRef]
- Leiva, C.L.; Cangelosi, A.; Mariconda, V.; Farace, M.; Geoghegan, P.; Brero, L.; Fernández-Miyakawa, M.; Chacana, P. IgY-based antivenom against Bothrops alternatus: Production and neutralization efficacy. Toxicon 2019, 163, 84–92. [Google Scholar] [CrossRef]
- Liu, J.; He, Q.; Wang, W.; Zhou, B.; Li, B.; Zhang, Y.; Luo, C.; Chen, D.; Tang, J.; Yu, X. Preparation and neutralization efficacy of IgY antibodies raised against Deinagkistrodon acutus venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 22. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, A.; Montero, Y.; Jimenez, E.; Zerpa, N.; Parrilla, P.; Malavé, C. IgY antibodies anti-Tityus caripitensis venom: Purification and neutralization efficacy. Toxicon 2013, 74, 208–214. [Google Scholar] [CrossRef]
- Aguilar, I.; Sánchez, E.E.; Girón, M.E.; Estrella, A.; Guerrero, B.; Rodriguez-Acosta, F.A. Coral snake antivenom produced in chickens (Gallus domesticus). Rev. Inst. Med. Trop. São Paulo 2014, 56, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Sevcik, C.; Díaz, P.; D’Suze, G. On the presence of antibodies against bovine, equine and poultry immunoglobulins in human IgG preparations, and its implications on antivenom production. Toxicon 2008, 51, 10–16. [Google Scholar] [CrossRef]
- Laustsen, A.H. Recombinant Antivenoms. Ph.D. Thesis, Faculty of Health and Medical Science, University of Copenhagen, Copenhagen, Denmark, 2016. [Google Scholar]
- Pucca, M.B.; Zoccal, K.F.; Roncolato, E.C.; Bertolini, T.B.; Campos, L.B.; Cologna, C.T.; Faccioli, L.H.; Arantes, E.C.; Barbosa, J.E. Serrumab: A human monoclonal antibody that counters the biochemical and immunological effects of Tityus serrulatus venom. J. Immunotoxicol. 2012, 9, 173–183. [Google Scholar] [CrossRef]
- Riaño-Umbarila, L.; Gómez-Ramírez, I.V.; Ledezma-Candanoza, L.M.; Olamendi-Portugal, T.; Rodríguez-Rodríguez, E.R.; Fernández-Taboada, G.; Possani, L.D.; Becerril, B. Generation of a Broadly Cross-Neutralizing Antibody Fragment against Several Mexican Scorpion Venoms. Toxins 2019, 11, 32. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, C.; Ledsgaard, L.; Dehli, R.I.; Ahmadi, S.; Sørensen, C.V.; Laustsen, A.H. Engineering and design considerations for next-generation snakebite antivenoms. Toxicon 2019, 167, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Dorrestijn, N. Integrating Engineering, Manufacturing, and Regulatory Considerations in the Development of Novel Antivenoms. Toxins 2018, 10, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, E.R.; Liu, J.L.; Zabetakis, D.; Anderson, G.P. Enhancing Stability of Camelid and Shark Single Domain Antibodies: An Overview. Front. Immunol. 2017, 8, 865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yardehnavi, N.; Behdani, M.; Pooshang Bagheri, K.; Mahmoodzadeh, A.; Khanahmad, H.; Shahbazzadeh, D.; Habibi-Anbouhi, M.; Ghassabeh, G.H.; Muyldermans, S. A camelid antibody candidate for development of a therapeutic agent against Hemiscorpius lepturus envenomation. FASEB J. 2014, 28, 4004–4014. [Google Scholar] [CrossRef] [Green Version]
- Debie, P.; Lafont, C.; Defrise, M.; Hansen, I.; van Willigen, D.M.; van Leeuwen, F.W.B.; Gijsbers, R.; D’Huyvetter, M.; Devoogdt, N.; Lahoutte, T.; et al. Size and affinity kinetics of nanobodies influence targeting and penetration of solid tumours. J. Control. Release 2020, 317, 34–42. [Google Scholar] [CrossRef]
- Alirahimi, E.; Kazemi-Lomedasht, F.; Shahbazzadeh, D.; Habibi-Anbouhi, M.; Hosseininejad Chafi, M.; Sotoudeh, N.; Ghaderi, H.; Muyldermans, S.; Behdani, M. Nanobodies as novel therapeutic agents in envenomation. Biochim. Biophys. Acta (BBA) Gen. Subj. 2018, 1862, 2955–2965. [Google Scholar] [CrossRef]
- Hmila, I.; Saerens, D.; Ben Abderrazek, R.; Vincke, C.; Abidi, N.; Benlasfar, Z.; Govaert, J.; El Ayeb, M.; Bouhaouala-Zahar, B.; Muyldermans, S. A bispecific nanobody to provide full protection against lethal scorpion envenoming. FASEB J. 2010, 24, 3479–3489. [Google Scholar] [CrossRef]
- Hmila, I.; Cosyns, B.; Tounsi, H.; Roosens, B.; Caveliers, V.; Abderrazek, R.B.; Boubaker, S.; Muyldermans, S.; El Ayeb, M.; Bouhaouala-Zahar, B.; et al. Pre-clinical studies of toxin-specific nanobodies: Evidence of in vivo efficacy to prevent fatal disturbances provoked by scorpion envenoming. Toxicol. Appl. Pharm. 2012, 264, 222–231. [Google Scholar] [CrossRef]
- Darvish, M.; Behdani, M.; Shokrgozar, M.A.; Pooshang-Bagheri, K.; Shahbazzadeh, D. Development of protective agent against Hottentotta saulcyi venom using camelid single-domain antibody. Mol. Immunol. 2015, 68, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Kerkis, I.; de Brandão, P.; da Silva, A.R.; Pompeia, C.; Tytgat, J.; de Sá Junior, P.L. Toxin bioportides: Exploring toxin biological activity and multifunctionality. Cell. Mol. Life Sci. 2017, 74, 647–661. [Google Scholar] [CrossRef] [PubMed]
- 131-I-TM-601 Study in Adults with Recurrent High-Grade Glioma—Phase 2. Available online: https://clinicaltrials.gov/ct2/show/NCT00114309 (accessed on 23 June 2019).
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide therapeutics from venom: Current status and potential. Bioorg. Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef]
- Jackson, N.; Czaplewski, L.; Piddock, L.J.V. Discovery and development of new antibacterial drugs: Learning from experience? J. Antimicrob. Chemother. 2018, 73, 1452–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bontems, F.; Roumestand, C.; Gilquin, B.; Ménez, A.; Toma, F. Refined structure of charybdotoxin: Common motifs in scorpion toxins and insect defensins. Science 1991, 254, 1521–1523. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, G. Insights into Antimicrobial Peptides from Spiders and Scorpions. Protein Pept. Lett. 2016, 23, 707–721. [Google Scholar] [CrossRef] [Green Version]
- Harrison, P.L.; Abdel-Rahman, M.A.; Miller, K.; Strong, P.N. Antimicrobial peptides from scorpion venoms. Toxicon 2014, 88, 115–137. [Google Scholar] [CrossRef]
- Du, Q.; Hou, X.; Wang, L.; Zhang, Y.; Xi, X.; Wang, H.; Zhou, M.; Duan, J.; Wei, M.; Chen, T.; et al. AaeAP1 and AaeAP2: Novel Antimicrobial Peptides from the Venom of the Scorpion, Androctonus aeneas: Structural Characterisation, Molecular Cloning of Biosynthetic Precursor-Encoding cDNAs and Engineering of Analogues with Enhanced Antimicrobial and Anticancer Activities. Toxins 2015, 7, 219–237. [Google Scholar] [CrossRef] [Green Version]
- Dueñas-Cuellar, R.A.; Kushmerick, C.; Naves, L.A.; Batista, I.F.C.; Guerrero-Vargas, J.A.; Pires, O.R.; Fontes, W.; Castro, M.S. Cm38: A new antimicrobial peptide active against Klebsiella pneumoniae is homologous to Cn11. Protein Pept. Lett. 2015, 22, 164–172. [Google Scholar] [CrossRef]
- De Melo, E.T.; Estrela, A.B.; Santos, E.C.G.; Machado, P.R.L.; Farias, K.J.S.; Torres, T.M.; Carvalho, E.; Lima, J.P.M.S.; Silva-Júnior, A.A.; Barbosa, E.G.; et al. Structural characterization of a novel peptide with antimicrobial activity from the venom gland of the scorpion Tityus stigmurus: Stigmurin. Peptides 2015, 68, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Kaya, M.; Baran, T.; Asan-Ozusaglam, M.; Cakmak, Y.S.; Tozak, K.O.; Mol, A.; Mentes, A.; Sezen, G. Extraction and characterization of chitin and chitosan with antimicrobial and antioxidant activities from cosmopolitan Orthoptera species (Insecta). Biotechnol. Bioproc. E 2015, 20, 168–179. [Google Scholar] [CrossRef]
- Harrison, P.L.; Abdel-Rahman, M.A.; Strong, P.N.; Tawfik, M.M.; Miller, K. Characterisation of three alpha-helical antimicrobial peptides from the venom of Scorpio maurus palmatus. Toxicon 2016, 117, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, M.; Kitanaka, A.; Yakio, M.; Yamazaki, Y.; Nakagawa, Y.; Miyagawa, H. Complete de novo sequencing of antimicrobial peptides in the venom of the scorpion Isometrus maculatus. Toxicon 2017, 139, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Marques-Neto, L.M.; Trentini, M.M.; das Neves, R.C.; Resende, D.P.; Procopio, V.O.; da Costa, A.C.; Kipnis, A.; Mortari, M.R.; Schwartz, E.F.; Junqueira-Kipnis, A.P. Antimicrobial and Chemotactic Activity of Scorpion-Derived Peptide, ToAP2, against Mycobacterium massiliensis. Toxins 2018, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Dalziel, J.; Tanzi, S.; Zhu, S. Meucin-49, a multifunctional scorpion venom peptide with bactericidal synergy with neurotoxins. Amino Acids 2018, 50, 1025–1043. [Google Scholar] [CrossRef]
- Liu, G.; Yang, F.; Li, F.; Li, Z.; Lang, Y.; Shen, B.; Wu, Y.; Li, W.; Harrison, P.L.; Strong, P.N.; et al. Therapeutic Potential of a Scorpion Venom-Derived Antimicrobial Peptide and Its Homologs Against Antibiotic-Resistant Gram-Positive Bacteria. Front. Microbiol. 2018, 9, 1159. [Google Scholar] [CrossRef]
- Juichi, H.; Ando, R.; Ishido, T.; Miyashita, M.; Nakagawa, Y.; Miyagawa, H. Chemical synthesis of a two-domain scorpion toxin LaIT2 and its single-domain analogs to elucidate structural factors important for insecticidal and antimicrobial activities. J. Pept. Sci. 2018, 24, e3133. [Google Scholar] [CrossRef]
- Parente, A.M.S.; Daniele-Silva, A.; Furtado, A.A.; Melo, M.A.; Lacerda, A.F.; Queiroz, M.; Moreno, C.; Santos, E.; Rocha, H.A.O.; Barbosa, E.G.; et al. Analogs of the Scorpion Venom Peptide Stigmurin: Structural Assessment, Toxicity, and Increased Antimicrobial Activity. Toxins 2018, 10, 161. [Google Scholar] [CrossRef] [Green Version]
- Luna-Ramirez, K.; Tonk, M.; Rahnamaeian, M.; Vilcinskas, A. Bioactivity of Natural and Engineered Antimicrobial Peptides from Venom of the Scorpions Urodacus yaschenkoi and U. manicatus. Toxins 2017, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Cesa-Luna, C.; Muñoz-Rojas, J.; Saab-Rincon, G.; Baez, A.; Morales-García, Y.E.; Juárez-González, V.R.; Quintero-Hernández, V. Structural characterization of scorpion peptides and their bactericidal activity against clinical isolates of multidrug-resistant bacteria. PLoS ONE 2019, 14, e0222438. [Google Scholar] [CrossRef] [Green Version]
- De Jesus Oliveira, T.; de Oliveira, U.C.; da Silva, P.I., Jr. Serrulin: A Glycine-Rich Bioactive Peptide from the Hemolymph of the Yellow Tityus serrulatus Scorpion. Toxins 2019, 11, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerouti, K.; Khemili, D.; Laraba-Djebari, F.; Hammoudi-Triki, D. Nontoxic fraction of scorpion venom reduces bacterial growth and inflammatory response in a mouse model of infection. Toxin Rev. 2019, 1–15. [Google Scholar] [CrossRef]
- Luna-Ramirez, K.; Skaljac, M.; Grotmann, J.; Kirfel, P.; Vilcinskas, A. Orally Delivered Scorpion Antimicrobial Peptides Exhibit Activity against Pea Aphid (Acyrthosiphon pisum) and Its Bacterial Symbionts. Toxins 2017, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Pedron, C.N.; Torres, M.D.T.; Lima, J.A.; da Silva, P.I.; Silva, F.D.; Oliveira, V.X. Novel designed VmCT1 analogs with increased antimicrobial activity. Eur. J. Med. Chem. 2017, 126, 456–463. [Google Scholar] [CrossRef]
- Amorim-Carmo, B.; Daniele-Silva, A.; Parente, A.M.S.; Furtado, A.A.; Carvalho, E.; Oliveira, J.W.F.; Santos, E.C.G.; Silva, M.S.; Silva, S.R.B.; Silva, A.A., Jr.; et al. Potent and Broad-Spectrum Antimicrobial Activity of Analogs from the Scorpion Peptide Stigmurin. Int. J. Mol. Sci. 2019, 20, 623. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.-Y.; Mi, Z.-M.; Cheng, G.-F.; Shen, W.-Q.; Xiao, X.; Liu, X.-M.; Liang, X.-T.; Yu, D.-Q. Purification and characterization of a new peptide with analgesic effect from the scorpion Buthus martensi Karch. J. Pept. Res. 2004, 64, 33–41. [Google Scholar] [CrossRef]
- Zeng, X.-C.; Wang, S.-X.; Zhu, Y.; Zhu, S.-Y.; Li, W.-X. Identification and functional characterization of novel scorpion venom peptides with no disulfide bridge from Buthus martensii Karsch. Peptides 2004, 25, 143–150. [Google Scholar] [CrossRef]
- Garcia, F.; Villegas, E.; Espino-Solis, G.P.; Rodriguez, A.; Paniagua-Solis, J.F.; Sandoval-Lopez, G.; Possani, L.D.; Corzo, G. Antimicrobial peptides from arachnid venoms and their microbicidal activity in the presence of commercial antibiotics. J. Antibiot. 2013, 66, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Pal, M. Morbidity and Mortality Due to Fungal Infections. J. Appl. Microbiol. Biochem. 2017, 1, 1–3. [Google Scholar] [CrossRef]
- Cabezas-Quintario, M.A.; Guerrero, C.; Gomez, P.; Perez-Fernandez, E. Prevalence of invasive fungal infections detected at necropsy in a medium-sized hospital: A 15-year review of autopsy findings. Rev. Esp. Patol. 2016, 49, 76–80. [Google Scholar] [CrossRef]
- Kullberg, B.J.; Arendrup, M.C. Invasive Candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Patterson, T.F. Multidrug-Resistant Candida: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2017, 216, S445–S451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, R.J.A.; Estrela, A.B.; Nascimento, A.K.L.; Melo, M.M.A.; Torres-Rêgo, M.; Lima, E.O.; Rocha, H.A.O.; Carvalho, E.; Silva-Junior, A.A.; Fernandes-Pedrosa, M.F. Characterization of TistH, a multifunctional peptide from the scorpion Tityus stigmurus: Structure, cytotoxicity and antimicrobial activity. Toxicon 2016, 119, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Guilhelmelli, F.; Vilela, N.; Smidt, K.S.; de Oliveira, M.A.; da Cunha Morales Álvares, A.; Rigonatto, M.C.L.; da Silva Costa, P.H.; Tavares, A.H.; de Freitas, S.M.; Nicola, A.M.; et al. Activity of Scorpion Venom-Derived Antifungal Peptides against Planktonic Cells of Candida spp. and Cryptococcus neoformans and Candida albicans Biofilms. Front. Microbiol. 2016, 7, 1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santussi, W.M.; Bordon, K.C.F.; Rodrigues Alves, A.P.N.; Cologna, C.T.; Said, S.; Arantes, E.C. Antifungal Activity against Filamentous Fungi of Ts1, a Multifunctional Toxin from Tityus serrulatus Scorpion Venom. Front. Microbiol. 2017, 8, 984. [Google Scholar] [CrossRef]
- Woolhouse, M.; Scott, F.; Hudson, Z.; Howey, R.; Chase-Topping, M. Human viruses: Discovery and emergence. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 2864–2871. [Google Scholar] [CrossRef] [Green Version]
- Da Mata, É.C.G.; Mourão, C.B.F.; Rangel, M.; Schwartz, E.F. Antiviral activity of animal venom peptides and related compounds. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 3. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhao, Z.; Zhou, D.; Chen, Y.; Hong, W.; Cao, L.; Yang, J.; Zhang, Y.; Shi, W.; Cao, Z.; et al. Virucidal activity of a scorpion venom peptide variant mucroporin-M1 against measles, SARS-CoV and influenza H5N1 viruses. Peptides 2011, 32, 1518–1525. [Google Scholar] [CrossRef]
- Zhao, Z.; Hong, W.; Zeng, Z.; Wu, Y.; Hu, K.; Tian, X.; Li, W.; Cao, Z. Mucroporin-M1 Inhibits Hepatitis B Virus Replication by Activating the Mitogen-activated Protein Kinase (MAPK) Pathway and Down-regulating HNF4α in Vitro and in Vivo. J. Biol. Chem. 2012, 287, 30181–30190. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Lang, Y.; Ji, Z.; Xia, Z.; Han, Y.; Cheng, Y.; Liu, G.; Sun, F.; Zhao, Y.; Gao, M.; et al. A scorpion venom peptide Ev37 restricts viral late entry by alkalizing acidic organelles. J. Biol. Chem. 2018, 294, 182–194. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Zhang, R.; Hong, W.; Cheng, Y.; Wang, H.; Lang, Y.; Ji, Z.; Wu, Y.; Li, W.; Xie, Y.; et al. Histidine-rich Modification of a Scorpion-derived Peptide Improves Bioavailability and Inhibitory Activity against HSV-1. Theranostics 2018, 8, 199–211. [Google Scholar] [CrossRef]
- Yu, Y.; Deng, Y.-Q.; Zou, P.; Wang, Q.; Dai, Y.; Yu, F.; Du, L.; Zhang, N.-N.; Tian, M.; Hao, J.-N.; et al. A peptide-based viral inactivator inhibits Zika virus infection in pregnant mice and fetuses. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Li, F.; Xia, Z.; Guo, X.; Gao, M.; Sun, F.; Cheng, Y.; Wu, Y.; Li, W.; Ali, S.A.; et al. The Scorpion Venom Peptide Smp76 Inhibits Viral Infection by Regulating Type-I Interferon Response. Virol. Sin. 2018, 33, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-S.; Tseng, C.-K.; Lin, C.-K.; Hsu, Y.-C.; Wu, Y.-H.; Hsieh, C.-L.; Lee, J.-C. Celastrol inhibits dengue virus replication via up-regulating type I interferon and downstream interferon-stimulated responses. Antivir. Res. 2017, 137, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Simner, P.J. Medical Parasitology Taxonomy Update: January 2012 to December 2015. J. Clin. Microbiol. 2017, 55, 43–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gockel-Blessing, E.A. Clinical Parasitology: A Practical Approach, 2nd ed.; Elsevier Saunders: St. Louis, MO, USA, 2013; ISBN 978-1-4160-6044-4. [Google Scholar]
- Kappagoda, S.; Singh, U.; Blackburn, B.G. Antiparasitic Therapy. Mayo Clin. Proc. 2011, 86, 561–583. [Google Scholar] [CrossRef] [Green Version]
- Frearson, J.A.; Wyatt, P.G.; Gilbert, I.H.; Fairlamb, A.H. Target assessment for antiparasitic drug discovery. Trends Parasitol. 2007, 23, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Conde, R.; Zamudio, F.Z.; Rodríguez, M.H.; Possani, L.D. Scorpine, an anti-malaria and anti-bacterial agent purified from scorpion venom. FEBS Lett. 2000, 471, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Carballar-Lejarazú, R.; Rodríguez, M.H.; de la Cruz Hernández-Hernández, F.; Ramos-Castañeda, J.; Possani, L.D.; Zurita-Ortega, M.; Reynaud-Garza, E.; Hernández-Rivas, R.; Loukeris, T.; Lycett, G.; et al. Recombinant scorpine: A multifunctional antimicrobial peptide with activity against different pathogens. Cell. Mol. Life Sci. 2008, 65, 3081–3092. [Google Scholar] [CrossRef]
- Gao, B.; Xu, J.; Rodriguez, M.d.C.; Lanz-Mendoza, H.; Hernández-Rivas, R.; Du, W.; Zhu, S. Characterization of two linear cationic antimalarial peptides in the scorpion Mesobuthus eupeus. Biochimie 2010, 92, 350–359. [Google Scholar] [CrossRef]
- Symeonidou, I.; Arsenopoulos, K.; Tzilves, D.; Soba, B.; Gabriël, S.; Papadopoulos, E. Human taeniasis/cysticercosis: A potentially emerging parasitic disease in Europe. Ann. Gastroenterol. 2018, 31, 406–412. [Google Scholar] [CrossRef]
- Taeniasis/Cysticercosis. Available online: https://www.who.int/news-room/fact-sheets/detail/taeniasis-cysticercosis (accessed on 22 January 2020).
- Toledo, A.; Larralde, C.; Fragoso, G.; Gevorkian, G.; Manoutcharian, K.; Hernández, M.; Acero, G.; Rosas, G.; López-Casillas, F.; Garfias, C.K.; et al. Towards a Taenia solium Cysticercosis Vaccine: An Epitope Shared by Taenia crassiceps and Taenia solium Protects Mice against Experimental Cysticercosis. Infect. Immun. 1999, 67, 2522–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Solis, D.; Toledano, Y.; Rodríguez-Lima, O.; Cano-Sánchez, P.; Ramírez-Cordero, B.E.; Landa, A.; de la Vega, R.C.R.; del Rio-Portilla, F. Solution structure and antiparasitic activity of scorpine-like peptides from Hoffmannihadrurus gertschi. FEBS Lett. 2016, 590, 2286–2296. [Google Scholar] [CrossRef] [PubMed]
- Chagas Disease. Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 22 January 2020).
- Borges, A.; Silva, S.; Op den Camp, H.J.M.; Velasco, E.; Alvarez, M.; Alfonzo, M.J.M.; Jorquera, A.; De Sousa, L.; Delgado, O. In vitro leishmanicidal activity of Tityus discrepans scorpion venom. Parasitol. Res. 2006, 99, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Golias, C.; Charalabopoulos, A.; Stagikas, D.; Charalabopoulos, K.; Batistatou, A. The kinin system—Bradykinin: Biological effects and clinical implications. Multiple role of the kinin system—Bradykinin. Hippokratia 2007, 11, 124–128. [Google Scholar] [PubMed]
- Riordan, J.F. Angiotensin-I-converting enzyme and its relatives. Genome Biol. 2003, 4, 225. [Google Scholar] [CrossRef] [Green Version]
- Camargo, A.C.M.; Ianzer, D.; Guerreiro, J.R.; Serrano, S.M.T. Bradykinin-potentiating peptides: Beyond captopril. Toxicon 2012, 59, 516–523. [Google Scholar] [CrossRef]
- Zeng, X.-C.; Wang, S.; Nie, Y.; Zhang, L.; Luo, X. Characterization of BmKbpp, a multifunctional peptide from the Chinese scorpion Mesobuthus martensii Karsch: Gaining insight into a new mechanism for the functional diversification of scorpion venom peptides. Peptides 2012, 33, 44–51. [Google Scholar] [CrossRef]
- Rocha-Resende, C.; Leão, N.M.; de Lima, M.E.; Santos, R.A.; Pimenta, A.M.d.C.; Verano-Braga, T. Moving pieces in a cryptomic puzzle: Cryptide from Tityus serrulatus Ts3 Nav toxin as potential agonist of muscarinic receptors. Peptides 2017, 98, 70–77. [Google Scholar] [CrossRef]
- Ferreira, L.A.F.; Alves, E.W.; Henriques, O.B. Peptide T, a novel bradykinin potentiator isolated from Tityus serrulatus scorpion venom. Toxicon 1993, 31, 941–947. [Google Scholar] [CrossRef]
- Verano-Braga, T.; Figueiredo-Rezende, F.; Melo, M.N.; Lautner, R.Q.; Gomes, E.R.M.; Mata-Machado, L.T.; Murari, A.; Rocha-Resende, C.; Elena de Lima, M.; Guatimosim, S.; et al. Structure-function studies of Tityus serrulatus Hypotensin-I (TsHpt-I): A new agonist of B(2) kinin receptor. Toxicon 2010, 56, 1162–1171. [Google Scholar] [CrossRef]
- Machado, R.J.A.; Junior, L.G.M.; Monteiro, N.K.V.; Silva-Júnior, A.A.; Portaro, F.C.V.; Barbosa, E.G.; Braga, V.A.; Fernandes-Pedrosa, M.F. Homology modeling, vasorelaxant and bradykinin-potentiating activities of a novel hypotensin found in the scorpion venom from Tityus stigmurus. Toxicon 2015, 101, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begg, A.C.; Stewart, F.A.; Vens, C. Strategies to improve radiotherapy with targeted drugs. Nat. Rev. Cancer 2011, 11, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Elshater, A.-E.; Salman, M.; Abd-Elhady, A. Physiological studies on the effect of a bradykinin potentiating factor (BPF) isolated from scorpion venom on the burnt skin of alloxan-induced diabetic Guinea pigs. Egypt. Acad. J. Biol. Sci. C Physiol. Mol. Biol. 2011, 3, 5–15. [Google Scholar] [CrossRef]
- Mohan, G.; Hamna, T.P.A.; Jijo, A.J.; Devi, S.K.M.; Narayanasamy, A.; Vellingiri, B. Recent advances in radiotherapy and its associated side effects in cancer—A review. J. Basic Appl. Zool. 2019, 80, 14. [Google Scholar] [CrossRef]
- Hasan, H.F.; Radwan, R.R.; Galal, S.M. Bradykinin-potentiating factor isolated from Leiurus quinquestriatus scorpion venom alleviates cardiomyopathy in irradiated rats via remodelling of the RAAS pathway. Clin. Exp. Pharm. Physiol. 2020, 47, 263–273. [Google Scholar] [CrossRef]
- Adi-Bessalem, S.; Hammoudi-Triki, D.; Laraba-Djebari, F. Scorpion Venom Interactions with the Immune System. In Scorpion Venoms; Gopalakrishnakone, P., Possani, L.D.F., Schwartz, E., Rodríguez de la Vega, R.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 87–107. ISBN 978-94-007-6403-3. [Google Scholar]
- Bertazzi, D.T.; de Assis-Pandochi, A.I.; Caleiro Seixas Azzolini, A.E.; Talhaferro, V.L.; Lazzarini, M.; Arantes, E.C. Effect of Tityus serrulatus scorpion venom and its major toxin, TsTX-I, on the complement system in vivo. Toxicon 2003, 41, 501–508. [Google Scholar] [CrossRef]
- Adi-Bessalem, S.; Hammoudi-Triki, D.; Laraba-Djebari, F. Pathophysiological effects of Androctonus australis hector scorpion venom: Tissue damages and inflammatory response. Exp. Toxicol. Pathol. 2008, 60, 373–380. [Google Scholar] [CrossRef]
- Magalhães, M.M.; Pereira, M.E.; Amaral, C.F.; Rezende, N.A.; Campolina, D.; Bucaretchi, F.; Gazzinelli, R.T.; Cunha-Melo, J.R. Serum levels of cytokines in patients envenomed by Tityus serrulatus scorpion sting. Toxicon 1999, 37, 1155–1164. [Google Scholar] [CrossRef]
- Fukuhara, Y.D.M.; Reis, M.L.; Dellalibera-Joviliano, R.; Cunha, F.Q.C.; Donadi, E.A. Increased plasma levels of IL-1beta, IL-6, IL-8, IL-10 and TNF-alpha in patients moderately or severely envenomed by Tityus serrulatus scorpion sting. Toxicon 2003, 41, 49–55. [Google Scholar] [CrossRef]
- Petricevich, V.L. Balance between pro- and anti-inflammatory cytokines in mice treated with Centruroides noxius scorpion venom. Mediat. Inflamm. 2006, 2006, 54273. [Google Scholar] [CrossRef] [Green Version]
- Abdoon, N.A.; Fatani, A.J. Correlation between blood pressure, cytokines and nitric oxide in conscious rabbits injected with Leiurus quinquestriatus quinquestriatus scorpion venom. Toxicon 2009, 54, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Petricevich, V.L. Cytokine and nitric oxide production following severe envenomation. Curr. Drug. Targets Inflamm. Allergy 2004, 3, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Huang, J.; Yuan, X.; Peng, B.; Liu, W.; Han, S.; He, X. Toxins Targeting the KV1.3 Channel: Potential Immunomodulators for Autoimmune Diseases. Toxins 2015, 7, 1749–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, I.S.; Ferreira, I.G.; Alexandre-Silva, G.M.; Cerni, F.A.; Cremonez, C.M.; Arantes, E.C.; Zottich, U.; Pucca, M.B.; Oliveira, I.S.; Ferreira, I.G.; et al. Scorpion toxins targeting Kv1.3 channels: Insights into immunosuppression. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25. [Google Scholar] [CrossRef] [Green Version]
- Lam, J.; Wulff, H. The Lymphocyte Potassium Channels Kv1.3 and KCa3.1 as Targets for Immunosuppression. Drug Dev. Res. 2011, 72, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Pucca, M.B.; Bertolini, T.B.; Cerni, F.A.; Bordon, K.C.F.; Peigneur, S.; Tytgat, J.; Bonato, V.L.; Arantes, E.C. Immunosuppressive evidence of Tityus serrulatus toxins Ts6 and Ts15: Insights of a novel K(+) channel pattern in T cells. Immunology 2016, 147, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.; Ding, L.; Yang, W.; Chai, L.; Sun, Y.; Yang, X.; Li, D.; Zhang, H.; Li, W.; Cao, Z.; et al. St20, a new venomous animal derived natural peptide with immunosuppressive and anti-inflammatory activities. Toxicon 2017, 127, 37–43. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Lewis, R.J. Sodium channels and pain: From toxins to therapies. Br. J. Pharmacol. 2018, 175, 2138–2157. [Google Scholar] [CrossRef] [Green Version]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International Union of Pharmacology. XLVII. Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharm. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef]
- Deuis, J.R.; Zimmermann, K.; Romanovsky, A.A.; Possani, L.D.; Cabot, P.J.; Lewis, R.J.; Vetter, I. An animal model of oxaliplatin-induced cold allodynia reveals a crucial role for Nav1.6 in peripheral pain pathways. Pain 2013, 154, 1749–1757. [Google Scholar] [CrossRef] [Green Version]
- Osteen, J.D.; Herzig, V.; Gilchrist, J.; Emrick, J.J.; Zhang, C.; Wang, X.; Castro, J.; Garcia-Caraballo, S.; Grundy, L.; Rychkov, G.Y.; et al. Selective spider toxins reveal a role for Nav1.1 channel in mechanical pain. Nature 2016, 534, 494–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.J.; Reimann, F.; Nicholas, A.K.; Thornton, G.; Roberts, E.; Springell, K.; Karbani, G.; Jafri, H.; Mannan, J.; Raashid, Y.; et al. An SCN9A channelopathy causes congenital inability to experience pain. Nature 2006, 444, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Phatarakijnirund, V.; Mumm, S.; McAlister, W.H.; Novack, D.V.; Wenkert, D.; Clements, K.L.; Whyte, M.P. Congenital insensitivity to pain: Fracturing without apparent skeletal pathobiology caused by an autosomal dominant, second mutation in SCN11A encoding voltage-gated sodium channel 1.9. Bone 2016, 84, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Peigneur, S.; Cologna, C.T.; Cremonez, C.M.; Mille, B.G.; Pucca, M.B.; Cuypers, E.; Arantes, E.C.; Tytgat, J. A gamut of undiscovered electrophysiological effects produced by Tityus serrulatus toxin 1 on NaV-type isoforms. Neuropharmacology 2015, 95, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Cologna, C.T.; Peigneur, S.; Rustiguel, J.K.; Nonato, M.C.; Tytgat, J.; Arantes, E.C. Investigation of the relationship between the structure and function of Ts2, a neurotoxin from Tityus serrulatus venom. FEBS J. 2012, 279, 1495–1504. [Google Scholar] [CrossRef]
- Motin, L.; Durek, T.; Adams, D.J. Modulation of human Nav1.7 channel gating by synthetic α-scorpion toxin OD1 and its analogs. Channels 2016, 10, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Da Mata, D.O.; Tibery, D.V.; Campos, L.A.; Camargos, T.S.; Peigneur, S.; Tytgat, J.; Schwartz, E.F. Subtype Specificity of β-Toxin Tf1a from Tityus fasciolatus in Voltage Gated Sodium Channels. Toxins 2018, 10, 339. [Google Scholar] [CrossRef] [Green Version]
- Chow, C.Y.; Chin, Y.K.-Y.; Walker, A.A.; Guo, S.; Blomster, L.V.; Ward, M.J.; Herzig, V.; Rokyta, D.R.; King, G.F. Venom Peptides with Dual Modulatory Activity on the Voltage-Gated Sodium Channel Nav1.1 Provide Novel Leads for Development of Antiepileptic Drugs. ACS Pharm. Transl. Sci. 2020, 3, 119–134. [Google Scholar] [CrossRef]
- Pucca, M.B.; Cerni, F.A.; Cordeiro, F.A.; Peigneur, S.; Cunha, T.M.; Tytgat, J.; Arantes, E.C. Ts8 scorpion toxin inhibits the Kv4.2 channel and produces nociception in vivo. Toxicon 2016, 119, 244–252. [Google Scholar] [CrossRef]
- Hakim, M.A.; Jiang, W.; Luo, L.; Li, B.; Yang, S.; Song, Y.; Lai, R. Scorpion Toxin, BmP01, Induces Pain by Targeting TRPV1 Channel. Toxins 2015, 7, 3671–3687. [Google Scholar] [CrossRef] [Green Version]
- Hoang, A.N.; Vo, H.D.M.; Vo, N.P.; Kudryashova, K.S.; Nekrasova, O.V.; Feofanov, A.V.; Kirpichnikov, M.P.; Andreeva, T.V.; Serebryakova, M.V.; Tsetlin, V.I.; et al. Vietnamese Heterometrus laoticus scorpion venom: Evidence for analgesic and anti-inflammatory activity and isolation of new polypeptide toxin acting on Kv1.3 potassium channel. Toxicon 2014, 77, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, C.; Chen, J.; Du, J.; Zhang, J.; Li, G.; Jin, X.; Wu, C. AGAP, a new recombinant neurotoxic polypeptide, targets the voltage-gated calcium channels in rat small diameter DRG neurons. Biochem. Biophys. Res. Commun. 2014, 452, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Martin-Eauclaire, M.-F.; Abbas, N.; Sauze, N.; Mercier, L.; Berge-Lefranc, J.-L.; Condo, J.; Bougis, P.E.; Guieu, R. Involvement of endogenous opioid system in scorpion toxin-induced antinociception in mice. Neurosci. Lett. 2010, 482, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Guan, R.-J.; Wang, C.-G.; Wang, M.; Wang, D.-C. A depressant insect toxin with a novel analgesic effect from scorpion Buthus martensii Karsch. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 2001, 1549, 9–18. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Wang, Z.; Zhang, X.; Liang, X.; Civelli, O. BmK-YA, an Enkephalin-Like Peptide in Scorpion Venom. PLoS ONE 2012, 7, e40417. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Hu, P.; Wu, W.; Wang, Y. Peptides with therapeutic potential in the venom of the scorpion Buthus martensii Karsch. Peptides 2019, 115, 43–50. [Google Scholar] [CrossRef]
- Renata, M.; Siqueira Cunh, A.O. New Perspectives in Drug Discovery Using Neuroactive Molecules from the Venom of Arthropods. In An Integrated View of the Molecular Recognition and Toxinology—From Analytical Procedures to Biomedical Applications; InTech: London, UK, 2013; Volume I, p. 13. [Google Scholar]
- Rigo, F.K.; Bochi, G.V.; Pereira, A.L.; Adamante, G.; Ferro, P.R.; Dal-Toé De Prá, S.; Milioli, A.M.; Damiani, A.P.; da Silveira Prestes, G.; Dalenogare, D.P.; et al. TsNTxP, a non-toxic protein from Tityus serrulatus scorpion venom, induces antinociceptive effects by suppressing glutamate release in mice. Eur. J. Pharmacol. 2019, 855, 65–74. [Google Scholar] [CrossRef]
- Chen, B.; Ji, Y. Antihyperalgesia effect of BmK AS, a scorpion toxin, in rat by intraplantar injection. Brain Res. 2002, 952, 322–326. [Google Scholar] [CrossRef]
- Chen, J.; Feng, X.-H.; Shi, J.; Tan, Z.-Y.; Bai, Z.-T.; Liu, T.; Ji, Y.-H. The anti-nociceptive effect of BmK AS, a scorpion active polypeptide, and the possible mechanism on specifically modulating voltage-gated Na+ currents in primary afferent neurons. Peptides 2006, 27, 2182–2192. [Google Scholar] [CrossRef]
- Cui, Y.; Song, Y.-B.; Ma, L.; Liu, Y.-F.; Li, G.-D.; Wu, C.-F.; Zhang, J.-H. Site-directed Mutagenesis of the Toxin from the Chinese Scorpion Buthus martensii Karsch (BmKAS): Insight into Sites Related to Analgesic Activity. Arch. Pharm. Res. 2010, 33, 1633–1639. [Google Scholar] [CrossRef]
- Liu, Z.R.; Tao, J.; Dong, B.Q.; Ding, G.; Cheng, Z.J.; He, H.Q.; Ji, Y.H. Pharmacological kinetics of BmK AS, a sodium channel site 4-specific modulator on Nav1.3. Neurosci. Bull. 2012, 28, 209–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Z.-Y.; Xiao, H.; Mao, X.; Wang, C.-Y.; Zhao, Z.-Q.; Ji, Y.-H. The inhibitory effects of BmK IT2, a scorpion neurotoxin on rat nociceptive flexion reflex and a possible mechanism for modulating voltage-gated Na+ channels. Neuropharmacology 2001, 40, 352–357. [Google Scholar] [CrossRef]
- Xiong, Y.M.; Lan, Z.D.; Wang, M.; Liu, B.; Liu, X.Q.; Fei, H.; Xu, L.G.; Xia, Q.C.; Wang, C.G.; Da-Cheng, W.; et al. Molecular characterization of a new excitatory insect neurotoxin with an analgesic effect on mice from the scorpion Buthus martensi Karsch. Toxicon 1999, 37, 1165–1180. [Google Scholar] [CrossRef]
- Maatoug, R.; Jebali, J.; Guieu, R.; De Waard, M.; Kharrat, R. BotAF, a new Buthus occitanus tunetanus scorpion toxin, produces potent analgesia in rodents. Toxicon 2018, 149, 72–85. [Google Scholar] [CrossRef]
- Song, Y.; Liu, Z.; Zhang, Q.; Li, C.; Jin, W.; Liu, L.; Zhang, J.; Zhang, J. Investigation of Binding Modes and Functional Surface of Scorpion Toxins ANEP to Sodium Channels 1.7. Toxins 2017, 9, 387. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wu, Y.; Zou, X.; Tang, Q.; Zhao, F.; Cao, Z. BmK AEP, an Anti-Epileptic Peptide Distinctly Affects the Gating of Brain Subtypes of Voltage-Gated Sodium Channels. Int. J. Mol. Sci. 2019, 20, 729. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.-Y.; Mao, X.; Xiao, H.; Zhao, Z.-Q.; Ji, Y.-H. Buthus martensi Karsch agonist of skeletal-muscle RyR-1, a scorpion active polypeptide: Antinociceptive effect on rat peripheral nervous system and spinal cord, and inhibition of voltage-gated Na+ currents in dorsal root ganglion neurons. Neurosci. Lett. 2001, 297, 65–68. [Google Scholar] [CrossRef]
- Mao, Q.; Ruan, J.; Cai, X.; Lu, W.; Ye, J.; Yang, J.; Yang, Y.; Sun, X.; Cao, J.; Cao, P. Antinociceptive Effects of Analgesic-Antitumor Peptide (AGAP), a Neurotoxin from the Scorpion Buthus martensii Karsch, on Formalin-Induced Inflammatory Pain through a Mitogen-Activated Protein Kinases–Dependent Mechanism in Mice. PLoS ONE 2013, 8, e78239. [Google Scholar] [CrossRef]
- Ruan, J.-P.; Mao, Q.-H.; Lu, W.-G.; Cai, X.-T.; Chen, J.; Li, Q.; Fu, Q.; Yan, H.-J.; Cao, J.-L.; Cao, P. Inhibition of spinal MAPKs by scorpion venom peptide BmK AGAP produces a sensory-specific analgesic effect. Mol. Pain 2018, 14, 1–11. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, J.-L.; Ming, H.-Y.; Zhang, Y.-N.; Dun, Y.-Q.; Zhang, J.-H.; Song, Y.-B. Insights into the binding mode and functional components of the analgesic-antitumour peptide from Buthus martensii Karsch to human voltage-gated sodium channel 1.7 based on dynamic simulation analysis. J. Biomol. Struct. Dyn. 2019, 38, 1–12. [Google Scholar] [CrossRef]
- Xu, Y.; Meng, X.; Hou, X.; Sun, J.; Kong, X.; Sun, Y.; Liu, Z.; Ma, Y.; Niu, Y.; Song, Y.; et al. A mutant of the Buthus martensii Karsch antitumor-analgesic peptide exhibits reduced inhibition to hNav1.4 and hNav1.5 channels while retaining analgesic activity. J. Biol. Chem. 2017, 292, 18270–18280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, R.-J.; Wang, M.; Wang, D.-C.; Wang, D.-C. A new insect neurotoxin AngP1 with analgesic effect from the scorpion Buthus martensii Karsch: Purification and characterization. J. Pept. Res. 2001, 58, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-M.; Liu, W.; Zhu, R.-H.; Wang, D.-C.; Goudet, C.; Tytgat, J. Roles of disulfide bridges in scorpion toxin BmK M1 analyzed by mutagenesis. J. Pept. Res. 2002, 60, 247–256. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, Z.; Shao, J.; Song, Y.; Li, C.; Li, C.; Zhao, Y.; Liu, Y.; Wei, T.; Wu, C.; et al. The role of Ser54 in the antinociceptive activity of BmK9, a neurotoxin from the scorpion Buthus martensii Karsch. Toxicon 2011, 58, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Liu, S.; Zhang, Y.; Qin, C.; Xu, L.; Li, W.; Cao, Z.; Li, W.; Wu, Y. Expression of recombinant α-toxin BmKM9 from scorpion Buthus martensii Karsch and its functional characterization on sodium channels. Peptides 2018, 99, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Cui, Y.; Song, Y.-B.; Liu, Y.-F.; Zhang, R.; Wu, C.-F.; Zhang, J.-H. Purification, characterization and functional expression of a new peptide with an analgesic effect from Chinese scorpion Buthus martensii Karsch (BmK AGP-SYPU1). Biomed. Chromatogr. 2011, 25, 801–807. [Google Scholar] [CrossRef]
- Zhang, R.; Cui, Y.; Zhang, X.; Yang, Z.; Zhao, Y.; Song, Y.; Wu, C.; Zhang, J. Soluble expression, purification and the role of C-terminal glycine residues in scorpion toxin BmK AGP-SYPU2. BMB Rep. 2010, 43, 801–806. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.-S.; Zhang, R.; Xu, Y.; Cui, Y.; Liu, Y.-F.; Song, Y.-B.; Zhang, H.-X.; Zhang, J.-H. The role of glycine residues at the C-terminal peptide segment in antinociceptive activity: A molecular dynamics simulation. J. Mol. Model. 2013, 19, 1295–1299. [Google Scholar] [CrossRef]
- Shao, J.-H.; Cui, Y.; Zhao, M.-Y.; Wu, C.-F.; Liu, Y.-F.; Zhang, J.-H. Purification, characterization, and bioactivity of a new analgesic-antitumor peptide from Chinese scorpion Buthus martensii Karsch. Peptides 2014, 53, 89–96. [Google Scholar] [CrossRef]
- Lin, S.; Wang, X.; Hu, X.; Zhao, Y.; Zhao, M.; Zhang, J.; Cui, Y. Recombinant Expression, Functional Characterization of Two Scorpion Venom Toxins with Three Disulfide Bridges from the Chinese Scorpion Buthus martensii Karsch. Protein Pept. Lett. 2017, 24, 235–240. [Google Scholar] [CrossRef]
- Anh, H.N.; Hoang, V.D.M.; Kudryashova, K.S.; Nekrasova, O.V.; Feofanov, A.V.; Andreeva, T.V.; Tsetlin, V.I.; Utkin, Y.N. Hetlaxin, a new toxin from the Heterometrus laoticus scorpion venom, interacts with voltage-gated potassium channel Kv1.3. Dokl. Biochem. Biophys. 2013, 449, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Srairi-Abid, N.; Othman, H.; Aissaoui, D.; BenAissa, R. Anti-tumoral effect of scorpion peptides: Emerging new cellular targets and signaling pathways. Cell Calcium 2019, 80, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Gómez Rave, L.J.; Muñoz Bravo, A.X.; Sierra Castrillo, J.; Román Marín, L.M.; Corredor Pereira, C. Scorpion Venom: New Promise in the Treatment of Cancer. Acta Biológica Colomb. 2019, 24, 213–223. [Google Scholar] [CrossRef]
- Akef, H.M. Anticancer and antimicrobial activities of scorpion venoms and their peptides. Toxin Rev. 2019, 38, 41–53. [Google Scholar] [CrossRef]
- Uzair, B.; Bint-e-Irshad, S.; Khan, B.A.; Azad, B.; Mahmood, T.; Rehman, M.U.; Braga, V.A. Scorpion Venom Peptides as a Potential Source for Human Drug Candidates. Protein Pept. Lett. 2018, 25, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Bernardes-Oliveira, E.; Farias, K.J.S.; Gomes, D.L.; de Araújo, J.M.G.; da Silva, W.D.; Rocha, H.A.O.; Donadi, E.A.; de Fernandes-Pedrosa, M.F.; de Crispim, J.C.O. Tityus serrulatus Scorpion Venom Induces Apoptosis in Cervical Cancer Cell Lines. Evid. Based Complementary Altern. Med. 2019, 2019, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Peng, Y.; Wang, Z.; Chai, X.; Lv, Z.; Song, Q. Venom from the scorpion Heterometrus liangi inhibits HeLa cell proliferation by inducing p21 expression. Biologia 2018, 73, 1099–1108. [Google Scholar] [CrossRef]
- Al-Asmari, A.K.; Riyasdeen, A.; Islam, M. Scorpion Venom Causes Apoptosis by Increasing Reactive Oxygen Species and Cell Cycle Arrest in MDA-MB-231 and HCT-8 Cancer Cell Lines. J. Evid. Based Integr. Med. 2018, 23, 215658721775179. [Google Scholar] [CrossRef] [Green Version]
- Giovannini, C.; Baglioni, M.; Baron Toaldo, M.; Cescon, M.; Bolondi, L.; Gramantieri, L. Vidatox 30 CH has tumor activating effect in hepatocellular carcinoma. Sci. Rep. 2017, 7, 44685. [Google Scholar] [CrossRef]
- Díaz-García, A.; Ruiz-Fuentes, J.L.; Rodríguez-Sánchez, H.; Fraga Castro, J.A. Rhopalurus junceus scorpion venom induces apoptosis in the triple negative human breast cancer cell line MDA-MB-231. J. Venom Res. 2017, 8, 9–13. [Google Scholar] [PubMed]
- Li, B.; Lyu, P.; Xi, X.; Ge, L.; Mahadevappa, R.; Shaw, C.; Kwok, H.F. Triggering of cancer cell cycle arrest by a novel scorpion venom-derived peptide—Gonearrestide. J. Cell. Mol. Med. 2018, 22, 4460–4473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BenAissa, R.; Othman, H.; Villard, C.; Peigneur, S.; Mlayah-Bellalouna, S.; Abdelkafi-Koubaa, Z.; Marrakchi, N.; Essafi-Benkhadir, K.; Tytgat, J.; Luis, J.; et al. AaHIV a sodium channel scorpion toxin inhibits the proliferation of DU145 prostate cancer cells. Biochem. Biophys. Res. Commun. 2019, 521, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-F.; Ma, R.-L.; Wang, S.-L.; Duan, Z.-Y.; Zhang, J.-H.; Wu, L.-J.; Wu, C.-F. Expression of an antitumor–analgesic peptide from the venom of Chinese scorpion Buthus martensii karsch in Escherichia coli. Protein Expr. Purif. 2003, 27, 253–258. [Google Scholar] [CrossRef]
- Kampo, S.; Ahmmed, B.; Zhou, T.; Owusu, L.; Anabah, T.W.; Doudou, N.R.; Kuugbee, E.D.; Cui, Y.; Lu, Z.; Yan, Q.; et al. Scorpion Venom Analgesic Peptide, BmK AGAP Inhibits Stemness, and Epithelial-Mesenchymal Transition by Down-Regulating PTX3 in Breast Cancer. Front. Oncol. 2019, 9, 21. [Google Scholar] [CrossRef]
- Tang, D.; Yang, Y.; Xiao, Z.; Xu, J.; Yang, Q.; Dai, H.; Liang, S.; Tang, C.; Dong, H.; Liu, Z. Scorpion toxin inhibits the voltage-gated proton channel using a Zn2+-like long-range conformational coupling mechanism. Br. J. Pharmacol. 2020, 177, 2351–2364. [Google Scholar] [CrossRef]
- Fernández, A.; Pupo, A.; Mena-Ulecia, K.; Gonzalez, C. Pharmacological modulation of proton channel hv1 in cancer therapy: Future perspectives. Mol. Pharmacol. 2016, 90, 385–402. [Google Scholar] [CrossRef] [Green Version]
- Tong-ngam, P.; Roytrakul, R.; Hathaitip, S. BmKn-2 scorpion venom peptide for killing oral cancer cells by apoptosis. Asian Pac. J. Cancer Prev. 2015, 16, 2807–2811. [Google Scholar] [CrossRef] [Green Version]
- Satitmanwiwat, S.; Changsangfa, C.; Khanuengthong, A.; Promthep, K.; Roytrakul, S.; Arpornsuwan, T.; Saikhun, K.; Sritanaudomchai, H. The scorpion venom peptide BmKn2 induces apoptosis in cancerous but not in normal human oral cells. Biomed. Pharm. 2016, 84, 1042–1050. [Google Scholar] [CrossRef]
- Khamessi, O.; Ben Mabrouk, H.; ElFessi-Magouri, R.; Kharrat, R. RK1, the first very short peptide from Buthus occitanus tunetanus inhibits tumor cell migration, proliferation and angiogenesis. Biochem. Biophys. Res. Commun. 2018, 499, 1–7. [Google Scholar] [CrossRef]




| Year | Scorpion Species | Antifungal Agent | MW (S–S Bridge) | Target | Reference |
|---|---|---|---|---|---|
| 2015 | T. stigmurus | Stigmurin | 1795.22 Da (0) | C. albicans, C. krusei, and C. glabrata | [98] |
| 2015 | A. aeneas | AaeAP1 | 2016.18 Da (0) | C. albicans | [96] |
| AaeAP2 | 1986.15 Da (0) | ||||
| 2016 | T. stigmurus | Hypotensin TistH | 2700 Da (0) | C. albicans, C. tropicalis and Aspergillus flavus | [121] |
| 2016 | T. obscurus | ToAcP, ToAP1, ToAP2, ToAP3, ToAP4 | ? (0) | Cryptococcus neoforman and Candida species | [122] |
| 2017 | T. serrulatus | Ts1 | 8300 Da (3) | A. nidulans | [123] |
| 2018 | T. stigmurus | StigA6 | 1908 Da (0?) | C. albicans, C. krusei, and C. glabrata | [106] |
| StigA16 | 1949 Da (0?) | ||||
| 2019 | T. serrulatus (hemolymph) | Serrulin | 3564 Da (0) | A. niger and C. albicans | [109] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmadi, S.; Knerr, J.M.; Argemi, L.; Bordon, K.C.F.; Pucca, M.B.; Cerni, F.A.; Arantes, E.C.; Çalışkan, F.; Laustsen, A.H. Scorpion Venom: Detriments and Benefits. Biomedicines 2020, 8, 118. https://doi.org/10.3390/biomedicines8050118
Ahmadi S, Knerr JM, Argemi L, Bordon KCF, Pucca MB, Cerni FA, Arantes EC, Çalışkan F, Laustsen AH. Scorpion Venom: Detriments and Benefits. Biomedicines. 2020; 8(5):118. https://doi.org/10.3390/biomedicines8050118
Chicago/Turabian StyleAhmadi, Shirin, Julius M. Knerr, Lídia Argemi, Karla C. F. Bordon, Manuela B. Pucca, Felipe A. Cerni, Eliane C. Arantes, Figen Çalışkan, and Andreas H. Laustsen. 2020. "Scorpion Venom: Detriments and Benefits" Biomedicines 8, no. 5: 118. https://doi.org/10.3390/biomedicines8050118