Increased ACh-Associated Immunoreactivity in Autonomic Centers in PTZ Kindling Model of Epilepsy

 ,
, 
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Method
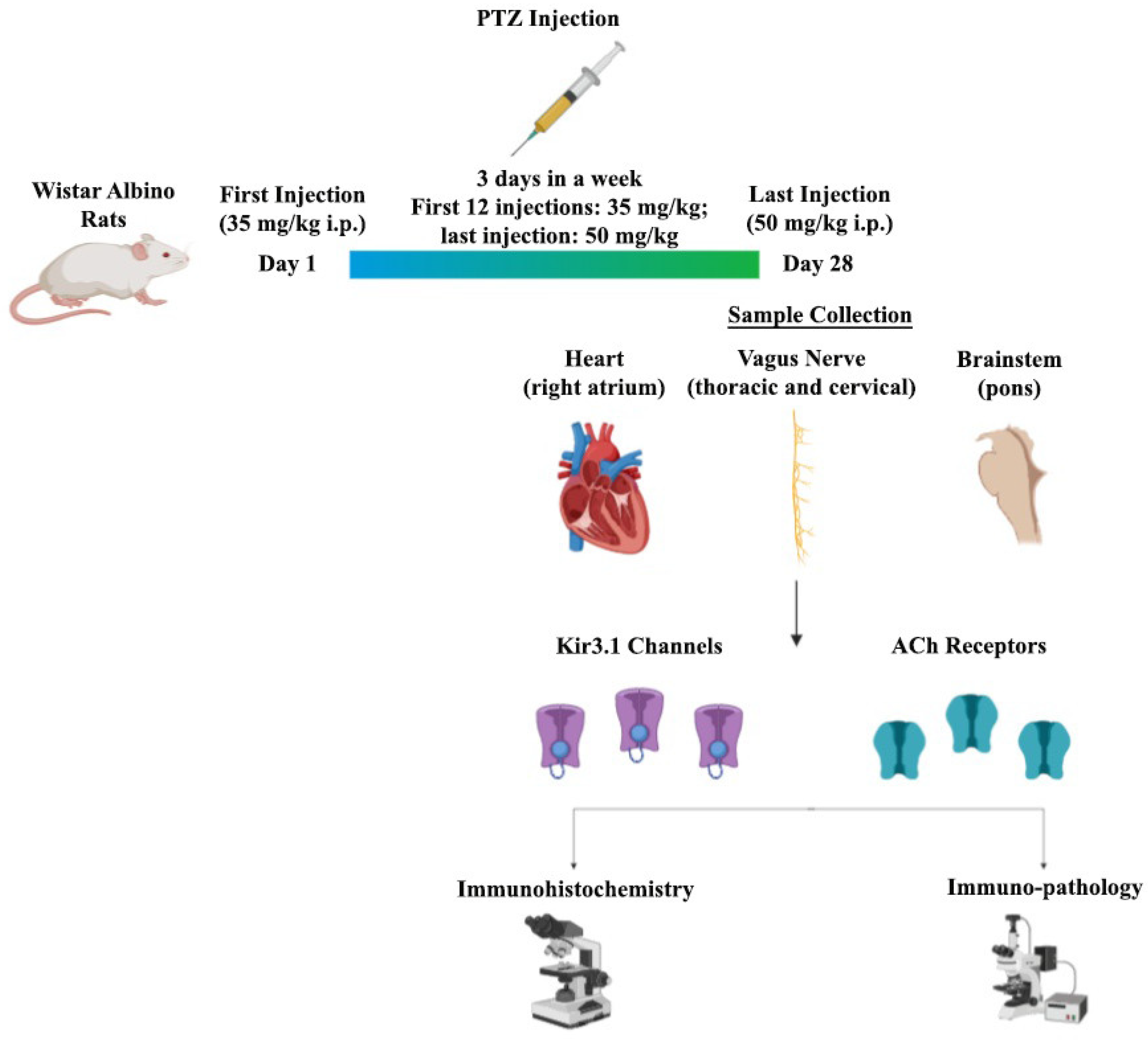
2.1. Experimental Epilepsy Model
2.1.1. Control Groups
2.1.2. PTZ Kindling Groups (n = 10 per each)
- Phase 0:
- No response to PTZ;
- Phase 1:
- Continuous ear and facial twitching;
- Phase 2:
- Myoclonic body jerks;
- Phase 3:
- Clonic forelimb convulsions;
- Phase 4:
- Tonic–clonic seizures;
- Phase 5:
- Generalized tonic–clonic seizures;
- Phase 6:
- Death.
2.2. Dissection of VN
2.3. Histological and Pathological Staining
2.4. Statistical Analysis
3. Results
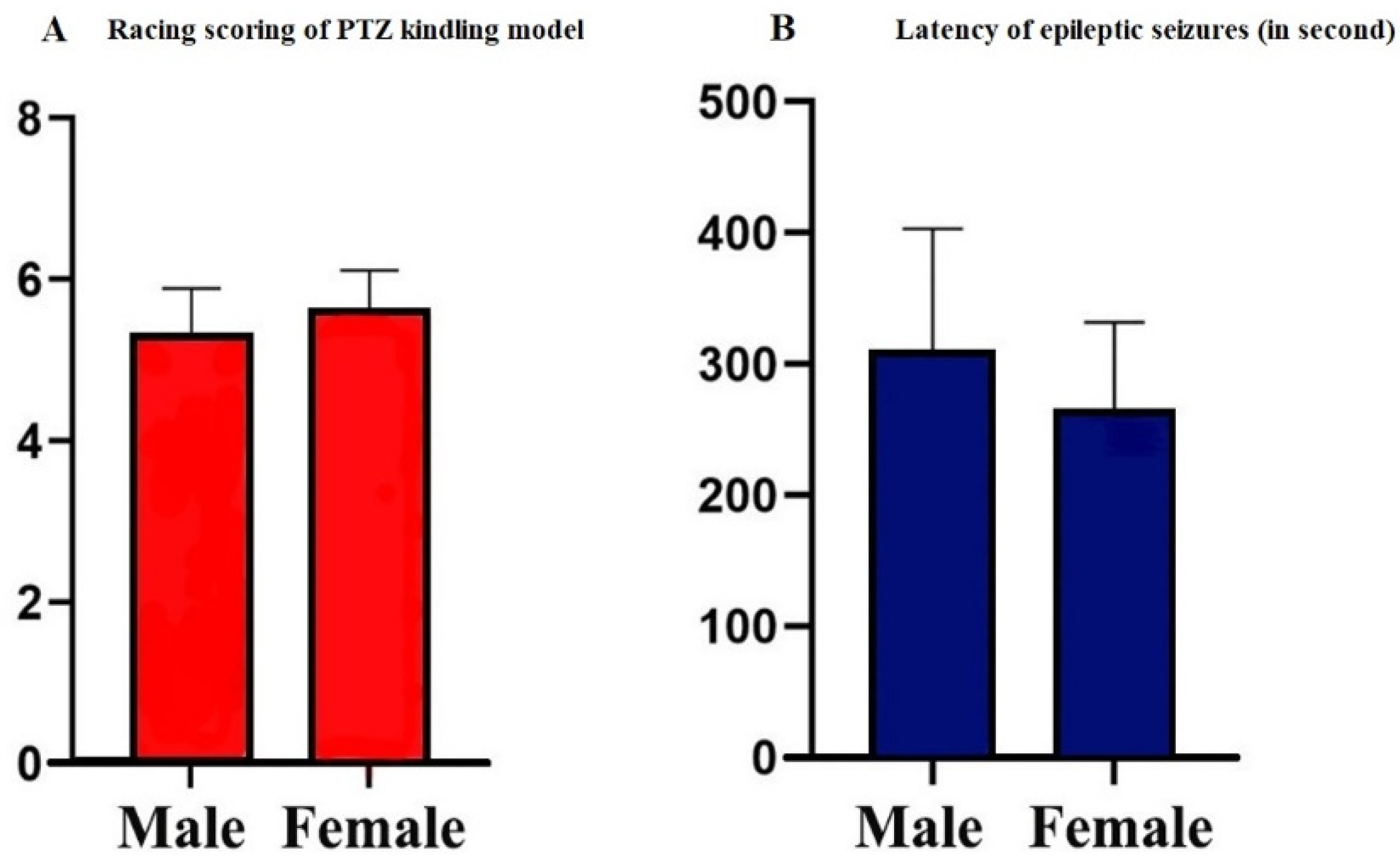
3.1. Scores of PTZ Kindling Epilepsy Model
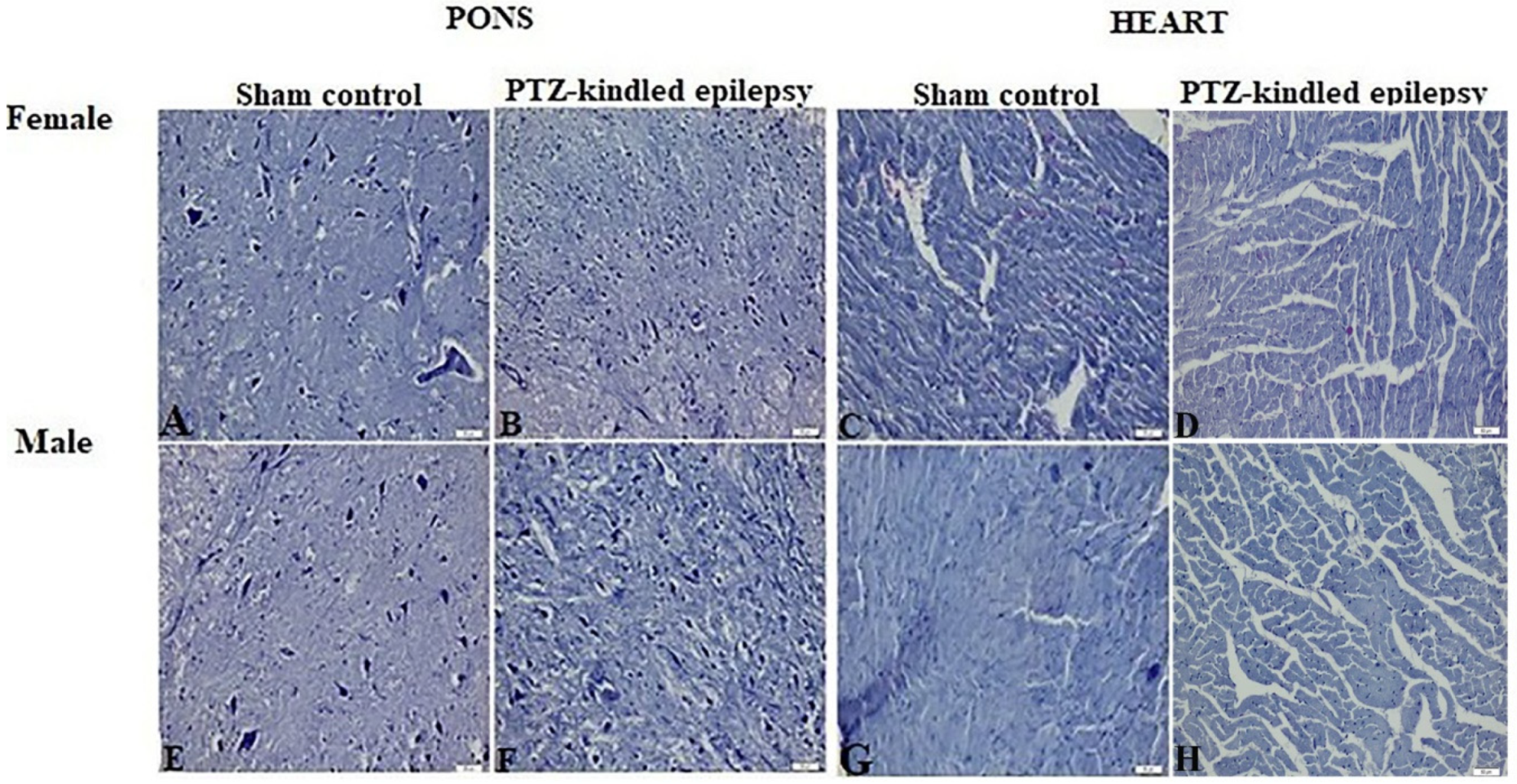
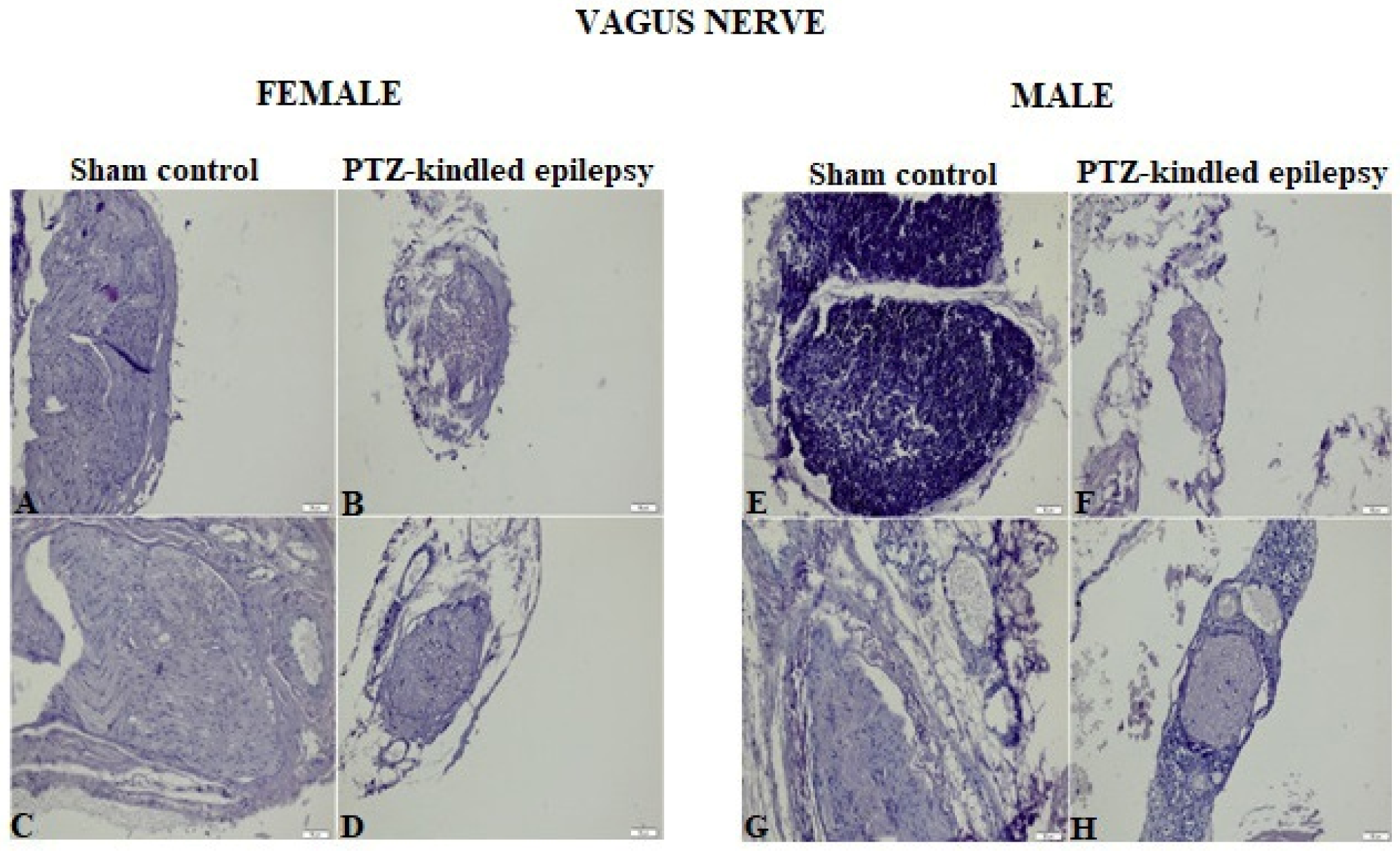
3.2. Histopathological Evaluation
3.3. Immunohistochemical Findings
3.3.1. Brainstem
3.3.2. VN
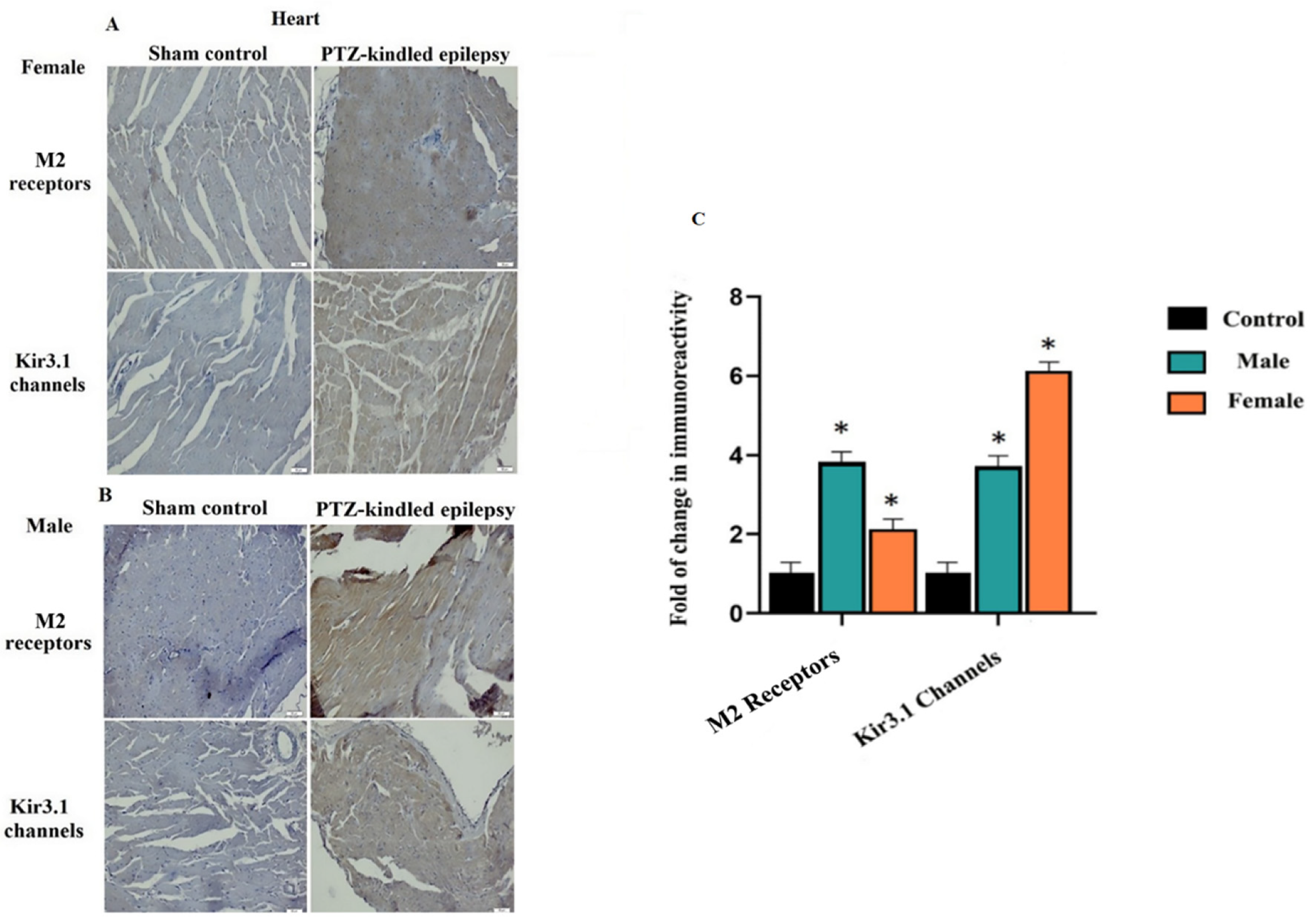
3.3.3. Cardiac Tissue
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ravindran, K.; Powell, K.L.; Todaro, M.; O’Brien, T.J. The pathophysiology of cardiac dysfunction in epilepsy. Epilepsy Res. 2016, 127, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Zeiler, F.; Zeiler, K.; Teitelbaum, J.; Gillman, L.; West, M. VNS for refractory status epilepticus. Epilepsy Res. 2015, 112, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Grimonprez, A.; Raedt, R.; Dauwe, I.; Mollet, L.; Larsen, L.E.; Meurs, A.; De Herdt, V.; Wadman, W.; Delbeke, J.; Vonck, K. Vagus nerve stimulation has antidepressant effects in the kainic acid model for temporal lobe epilepsy. Brain Stimul. 2015, 8, 13–20. [Google Scholar] [CrossRef]
- Barot, N.; Nei, M. Autonomic aspects of sudden unexpected death in epilepsy (SUDEP). Clin. Auton. Res. 2019, 29, 151–160. [Google Scholar] [CrossRef]
- Sevcencu, C.; Struijk, J.J. Autonomic alterations and cardiac changes in epilepsy. Epilepsia 2010, 51, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Ansakorpi, H.; Korpelainen, J.T.; Suominen, K.; Tolonen, U.; Myllylä, V.V.; Isojärvi, J.I. Interictal cardiovascular autonomic responses in patients with temporal lobe epilepsy. Epilepsia 2000, 41, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Ferri, R.; Curzi-Dascalova, L.; Arzimanoglou, A.; Bourgeois, M.; Beaud, C.; Nunes, M.L.; Elia, M.; Musumeci, S.A.; Tripodi, M. Heart rate variability during sleep in children with partial epilepsy. J. Sleep Res. 2002, 11, 153–160. [Google Scholar] [CrossRef]
- Henry, T.R. Therapeutic mechanisms of vagus nerve stimulation. Neurology 2002, 59, S3–S14. [Google Scholar] [CrossRef]
- Tata, A.M.; Velluto, L.; D’Angelo, C.; Reale, M. Cholinergic system dysfunction and neurodegenerative diseases: Cause or effect? Cns Neurol. Disord. -Drug Targets (Former. Curr. Drug Targets-Cns Neurol. Disord.) 2014, 13, 1294–1303. [Google Scholar] [CrossRef]
- Holt, R.L.; Arehart, E.; Hunanyan, A.; Fainberg, N.A.; Mikati, M.A. Pediatric sudden unexpected death in epilepsy: What have we learned from animal and human studies, and can we prevent it? Semin. Pediatr. Neurol. 2016, 23, 127–133. [Google Scholar] [CrossRef]
- Massey, C.A.; Sowers, L.P.; Dlouhy, B.J.; Richerson, G.B. Mechanisms of sudden unexpected death in epilepsy: The pathway to prevention. Nat. Rev. Neurol. 2014, 10, 271. [Google Scholar] [CrossRef] [Green Version]
- Damasceno, D.D.; Savergnini, S.Q.; Gomes, E.R.; Guatimosim, S.; Ferreira, A.J.; Doretto, M.C.; Almeida, A.P. Cardiac dysfunction in rats prone to audiogenic epileptic seizures. Seizure 2013, 22, 259–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazan, R., Jr.; de Oliveira, M.; Oliveira, J.A.C.; Salgado, H.C.; Garcia-Cairasco, N. Changes in autonomic control of the cardiovascular system in the Wistar audiogenic rat (WAR) strain. Epilepsy Behav. 2011, 22, 666–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devinsky, O.; Hesdorffer, D.C.; Thurman, D.J.; Lhatoo, S.; Richerson, G. Sudden unexpected death in epilepsy: Epidemiology, mechanisms, and prevention. Lancet Neurol. 2016, 15, 1075–1088. [Google Scholar] [CrossRef]
- Avery, M.C.; Krichmar, J.L. Neuromodulatory systems and their interactions: a review of models, theories, and experiments. Front. Neural Circuits 2017, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Hibino, H.; Inanobe, A.; Furutani, K.; Murakami, S.; Findlay, I.; Kurachi, Y. Inwardly rectifying potassium channels: Their structure, function, and physiological roles. Physiol. Rev. 2010, 90, 291–366. [Google Scholar] [CrossRef] [Green Version]
- Ten Eick, R.; Nawrath, H.; McDonald, T.F.; Trautwein, W. On the mechanism of the negative inotropic effect of acetylcholine. Pflügers Arch. 1976, 361, 207–213. [Google Scholar] [CrossRef]
- Myers, K.A.; Bello-Espinosa, L.E.; Symonds, J.D.; Zuberi, S.M.; Clegg, R.; Sadleir, L.G.; Buchhalter, J.; Scheffer, I.E. Heart rate variability in epilepsy: A potential biomarker of sudden unexpected death in epilepsy risk. Epilepsia 2018, 59, 1372–1380. [Google Scholar] [CrossRef] [Green Version]
- Lüscher, C.; Slesinger, P.A. Emerging roles for G protein-gated inwardly rectifying potassium (GIRK) channels in health and disease. Nat. Rev. Neurosci. 2010, 11, 301–315. [Google Scholar] [CrossRef]
- Touhara, K.K.; MacKinnon, R. Molecular basis of signaling specificity between GIRK channels and GPCRs. Elife 2018, 7, e42908. [Google Scholar] [CrossRef]
- Hilger, D.; Masureel, M.; Kobilka, B.K. Structure and dynamics of GPCR signaling complexes. Nat. Struct. Mol. Biol. 2018, 25, 4. [Google Scholar] [CrossRef] [PubMed]
- Downes, G.B.; Granato, M. Acetylcholinesterase function is dispensable for sensory neurite growth but is critical for neuromuscular synapse stability. Dev. Biol. 2004, 270, 232–245. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.; Behrens, C.J.; Heinemann, U. Cholinergic dysfunction in temporal lobe epilepsy. Epilepsia 2007, 48, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, G.; Njunting, M.; Ivens, S.; Tolner, E.; Behrens, C.J.; Gross, M.; Soreq, H.; Heinemann, U.; Friedman, A. Acetylcholine-induced seizure-like activity and modified cholinergic gene expression in chronically epileptic rats. Eur. J. Neurosci. 2008, 27, 965–975. [Google Scholar] [CrossRef]
- Furman, M.; Zhan, Q.; McCafferty, C.; Lerner, B.A.; Motelow, J.E.; Meng, J.; Ma, C.; Buchanan, G.F.; Witten, I.B.; Deisseroth, K. Optogenetic stimulation of cholinergic brainstem neurons during focal limbic seizures: Effects on cortical physiology. Epilepsia 2015, 56, e198–e202. [Google Scholar] [CrossRef] [Green Version]
- Broncel, A.; Bocian, R.; Kłos-Wojtczak, P.; Konopacki, J. Medial septal cholinergic mediation of hippocampal theta rhythm induced by vagal nerve stimulation. PLoS ONE 2018, 13, e0206532. [Google Scholar] [CrossRef]
- Castle, M.; Comoli, E.; Loewy, A. Autonomic brainstem nuclei are linked to the hippocampus. Neuroscience 2005, 134, 657–669. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of seizure activity by electrical stimulation: II. Motor seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
- Löscher, W. Animal models of seizures and epilepsy: Past, present, and future role for the discovery of antiseizure drugs. Neurochem. Res. 2017, 42, 1873–1888. [Google Scholar] [CrossRef]
- Powley, T.L.; Prechtl, J.C.; Fox, E.A.; Berthoud, H.-R. Anatomical considerations for surgery of the rat abdominal vagus: Distribution, paraganglia and regeneration. J. Auton. Nerv. Syst. 1983, 9, 79–97. [Google Scholar] [CrossRef]
- Devinsky, O. Effects of seizures on autonomic and cardiovascular function. Epilepsy Curr. 2004, 4, 43–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCorry, L.K. Physiology of the autonomic nervous system. Am. J. Pharm. Educ. 2007, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brimijoin, S. Molecular forms of acetylcholinesterase in brain, nerve and muscle: Nature, localization and dynamics. Prog. Neurobiol. 1983, 21, 291–322. [Google Scholar] [CrossRef]
- Leuzinger, W. Structure and function of acetylcholinesterase. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 1969; Volume 31, pp. 241–245. [Google Scholar]
- Dobrzynski, H.; Marples, D.D.; Musa, H.; Yamanushi, T.T.; Hendersonxyl, Z.; Takagishi, Y.; Honjo, H.; Kodama, I.; Boyett, M.R. Distribution of the muscarinic K+ channel proteins Kir3. 1 and Kir3. 4 in the ventricle, atrium, and sinoatrial node of heart. J. Histochem. Cytochem. 2001, 49, 1221–1234. [Google Scholar] [CrossRef] [Green Version]
- Bettahi, I.; Marker, C.L.; Roman, M.I.; Wickman, K. Contribution of the Kir3. 1 subunit to the muscarinic-gated atrial potassium channel IKACh. J. Biol. Chem. 2002, 277, 48282–48288. [Google Scholar] [CrossRef] [Green Version]
- Celada, P.; Puig, M.; Artigas, F. Serotonin modulation of cortical neurons and networks. Front. Integr. Neurosci. 2013, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Safaai, H.; Neves, R.; Eschenko, O.; Logothetis, N.K.; Panzeri, S. Modeling the effect of locus coeruleus firing on cortical state dynamics and single-trial sensory processing. Proc. Natl. Acad. Sci. USA 2015, 112, 12834–12839. [Google Scholar] [CrossRef] [Green Version]
- Neves, R.M.; van Keulen, S.; Yang, M.; Logothetis, N.K.; Eschenko, O. Locus coeruleus phasic discharge is essential for stimulus-induced gamma oscillations in the prefrontal cortex. J. Neurophysiol. 2018, 119, 904–920. [Google Scholar] [CrossRef]
- Jefferys, J.G.; Arafat, M.A.; Irazoqui, P.P.; Lovick, T.A. Brainstem activity, apnea, and death during seizures induced by intrahippocampal kainic acid in anaesthetized rats. Epilepsia 2019, 60, 2346–2358. [Google Scholar] [CrossRef]
- Dorr, A.E.; Debonnel, G. Effect of vagus nerve stimulation on serotonergic and noradrenergic transmission. J. Pharmacol. Exp. Ther. 2006, 318, 890–898. [Google Scholar] [CrossRef]
- Patodia, S.; Somani, A.; O’Hare, M.; Venkateswaran, R.; Liu, J.; Michalak, Z.; Ellis, M.; Scheffer, I.E.; Diehl, B.; Sisodiya, S.M. The ventrolateral medulla and medullary raphe in sudden unexpected death in epilepsy. Brain 2018, 141, 1719–1733. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.G.; Bateman, L.M.; Nei, M.; Goldman, A.M.; Laxer, K.D. Brainstem atrophy in focal epilepsy destabilizes brainstem-brain interactions: Preliminary findings. Neuroimage Clin. 2019, 23, 101888. [Google Scholar] [CrossRef] [PubMed]
- Seki, A.; Green, H.R.; Lee, T.D.; Hong, L.; Tan, J.; Vinters, H.V.; Chen, P.-S.; Fishbein, M.C. Sympathetic nerve fibers in human cervical and thoracic vagus nerves. Heart Rhythm 2014, 11, 1411–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randall, W.C.; Rohse, W.G. The augmentor action of the sympathetic cardiac nerves. Circ. Res. 1956, 4, 470–475. [Google Scholar] [CrossRef] [Green Version]
- Frei, M.G.; Osorio, I. Left vagus nerve stimulation with the neurocybernetic prosthesis has complex effects on heart rate and on its variability in humans. Epilepsia 2001, 42, 1007–1016. [Google Scholar] [CrossRef]
- Akyüz, E.; Tiber, P.M.; Beker, M.; Akbaş, F. Expression of cardiac inwardly rectifying potassium channels in pentylenetetrazole kindling model of epilepsy in rats. Cell. Mol. Biol. (Noisy-Le-Grand) 2018, 64, 47–54. [Google Scholar]
- Mueller, S.G.; Bateman, L.M.; Laxer, K.D. Evidence for brainstem network disruption in temporal lobe epilepsy and sudden unexplained death in epilepsy. Neuroimage Clin. 2014, 5, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Moore, B.M.; Jerry Jou, C.; Tatalovic, M.; Kaufman, E.S.; Kline, D.D.; Kunze, D.L. The Kv1. 1 null mouse, a model of sudden unexpected death in epilepsy (SUDEP). Epilepsia 2014, 55, 1808–1816. [Google Scholar] [CrossRef] [Green Version]
- Sakmann, B.; Noma, A.; Trautwein, W. Acetylcholine activation of single muscarinic K+ channels in isolated pacemaker cells of the mammalian heart. Nature 1983, 303, 250–253. [Google Scholar] [CrossRef]
- Dergacheva, O.; Weigand, L.A.; Dyavanapalli, J.; Mares, J.; Wang, X.; Mendelowitz, D. Function and modulation of premotor brainstem parasympathetic cardiac neurons that control heart rate by hypoxia-, sleep-, and sleep-related diseases including obstructive sleep apnea. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 212, pp. 39–58. [Google Scholar]
- Ruffoli, R.; Giorgi, F.S.; Pizzanelli, C.; Murri, L.; Paparelli, A.; Fornai, F. The chemical neuroanatomy of vagus nerve stimulation. J. Chem. Neuroanat. 2011, 42, 288–296. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akyüz, E.; Doğanyiğit, Z.; Paudel, Y.N.; Kaymak, E.; Yilmaz, S.; Uner, A.; Shaikh, M.F. Increased ACh-Associated Immunoreactivity in Autonomic Centers in PTZ Kindling Model of Epilepsy. Biomedicines 2020, 8, 113. https://doi.org/10.3390/biomedicines8050113
Akyüz E, Doğanyiğit Z, Paudel YN, Kaymak E, Yilmaz S, Uner A, Shaikh MF. Increased ACh-Associated Immunoreactivity in Autonomic Centers in PTZ Kindling Model of Epilepsy. Biomedicines. 2020; 8(5):113. https://doi.org/10.3390/biomedicines8050113
Chicago/Turabian StyleAkyüz, Enes, Züleyha Doğanyiğit, Yam Nath Paudel, Emin Kaymak, Seher Yilmaz, Arda Uner, and Mohd. Farooq Shaikh. 2020. "Increased ACh-Associated Immunoreactivity in Autonomic Centers in PTZ Kindling Model of Epilepsy" Biomedicines 8, no. 5: 113. https://doi.org/10.3390/biomedicines8050113