Mesenchymal Stem Cells: A Trump Card for the Treatment of Diabetes?


Abstract
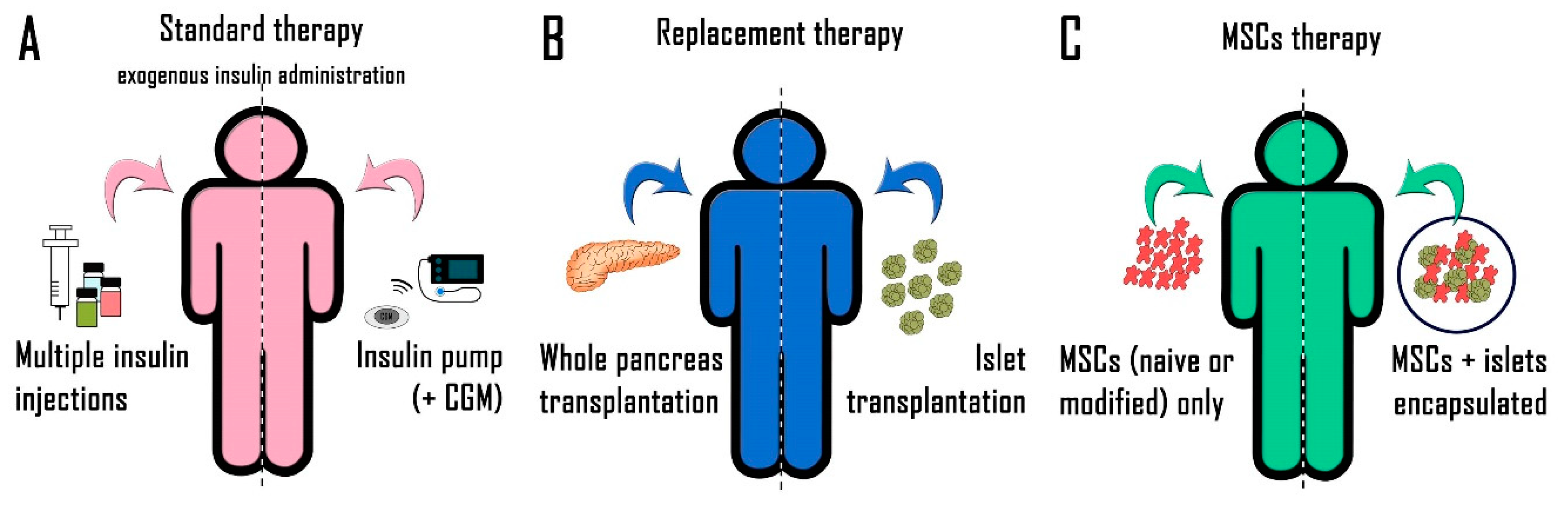
:1. Introduction
2. Diabetes Treatment
2.1. Exogenous Insulin Administration
2.2. New Therapeutic Approaches
2.2.1. Whole Pancreas Transplantation
2.2.2. Pancreatic Islet Transplantation
2.2.3. Beta-Cell Replacement
3. Mesenchymal Stem Cells
3.1. Administration of MSCs Alone
3.1.1. In Vitro Studies
3.1.2. In Vivo Studies
3.1.3. MSC Manipulation
3.2. Co-Transplantation of MSCs and Pancreatic Islets
3.2.1. Role of Soluble Factors
3.2.2. Role of Direct Contact
3.3. Clinical Use of MSCs
4. Conclusions
Funding
Conflicts of Interest
References
- Association, A.D. Standards of medical care in diabetes--2014. Diabetes Care 2014, 37, S14–S80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Mahato, R.I. Mesenchymal stem cell-based therapy for type 1 diabetes. Discov. Med. 2014, 17, 139–143. [Google Scholar] [PubMed]
- Ramírez-Domínguez, M. Historical Background of Pancreatic Islet Isolation. Adv. Exp. Med. Biol. 2016, 938, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pathak, V.; Pathak, N.M.; O’Neill, C.L.; Guduric-Fuchs, J.; Medina, R.J. Therapies for Type 1 Diabetes: Current Scenario and Future Perspectives. Clin. Med. Insights Endocrinol. Diabetes 2019, 12, 1179551419844521. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Pabbidi, R.M. Diabetic peripheral neuropathy: Role of reactive oxygen and nitrogen species. Cell Biochem. Biophys. 2013, 67, 373–383. [Google Scholar] [CrossRef]
- Maffi, P.; Secchi, A. Islet Transplantation Alone Versus Solitary Pancreas Transplantation: An Outcome-Driven Choice? Curr. Diab. Rep. 2019, 19, 26. [Google Scholar] [CrossRef]
- Tjernberg, J.; Ekdahl, K.N.; Lambris, J.D.; Korsgren, O.; Nilsson, B. Acute antibody-mediated complement activation mediates lysis of pancreatic islets cells and may cause tissue loss in clinical islet transplantation. Transplantation 2008, 85, 1193–1199. [Google Scholar] [CrossRef]
- Vaithilingam, V.; Bal, S.; Tuch, B.E. Encapsulated Islet Transplantation: Where Do We Stand? Rev. Diabet. Stud. 2017, 14, 51–78. [Google Scholar] [CrossRef] [Green Version]
- Millman, J.R.; Pagliuca, F.W. Autologous Pluripotent Stem Cell-Derived β-Like Cells for Diabetes Cellular Therapy. Diabetes 2017, 66, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Rezania, A.; Bruin, J.E.; Arora, P.; Rubin, A.; Batushansky, I.; Asadi, A.; O’Dwyer, S.; Quiskamp, N.; Mojibian, M.; Albrecht, T.; et al. Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol. 2014, 32, 1121–1133. [Google Scholar] [CrossRef]
- Pagliuca, F.W.; Millman, J.R.; Gürtler, M.; Segel, M.; Van Dervort, A.; Ryu, J.H.; Peterson, Q.P.; Greiner, D.; Melton, D.A. Generation of functional human pancreatic β cells in vitro. Cell 2014, 159, 428–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sackett, S.D.; Rodriguez, A.; Odorico, J.S. The Nexus of Stem Cell-Derived Beta-Cells and Genome Engineering. Rev. Diabet. Stud. 2017, 14, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, A.M.J. Gearing Up for Stem Cell-derived Beta Cells-Are We Ready? Transplantation 2018, 102, 1207–1208. [Google Scholar] [CrossRef] [PubMed]
- van der Torren, C.R.; Zaldumbide, A.; Duinkerken, G.; Brand-Schaaf, S.H.; Peakman, M.; Stangé, G.; Martinson, L.; Kroon, E.; Brandon, E.P.; Pipeleers, D.; et al. Immunogenicity of human embryonic stem cell-derived beta cells. Diabetologia 2017, 60, 126–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velazco-Cruz, L.; Song, J.; Maxwell, K.G.; Goedegebuure, M.M.; Augsornworawat, P.; Hogrebe, N.J.; Millman, J.R. Acquisition of Dynamic Function in Human Stem Cell-Derived β Cells. Stem. Cell Reports 2019, 12, 351–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremmel, D.M.; Mitchell, S.A.; Sackett, S.D.; Odorico, J.S. Mimicking nature-made beta cells: Recent advances towards stem cell-derived islets. Curr. Opin. Organ. Transplant. 2019, 24, 574–581. [Google Scholar] [CrossRef]
- Cho, J.; D’Antuono, M.; Glicksman, M.; Wang, J.; Jonklaas, J. A review of clinical trials: Mesenchymal stem cell transplant therapy in type 1 and type 2 diabetes mellitus. Am. J. Stem. Cells 2018, 7, 82–93. [Google Scholar]
- Scuteri, A.; Donzelli, E.; Rodriguez-Menendez, V.; Ravasi, M.; Monfrini, M.; Bonandrini, B.; Figliuzzi, M.; Remuzzi, A.; Tredici, G. A double mechanism for the mesenchymal stem cells’ positive effect on pancreatic islets. PLoS ONE 2014, 9, e84309. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.; Kahlenberg, S.; Hornsby, P. Therapeutic potential of mesenchymal stem cells for diabetes. J. Mol. Endocrinol. 2017, 59, R109–R120. [Google Scholar] [CrossRef]
- Stiner, R.; Alexander, M.; Liu, G.; Liao, W.; Liu, Y.; Yu, J.; Pone, E.J.; Zhao, W.; Lakey, J.R.T. Transplantation of stem cells from umbilical cord blood as therapy for type I diabetes. Cell Tissue Res. 2019, 378, 155–162. [Google Scholar] [CrossRef]
- Nojehdehi, S.; Soudi, S.; Hesampour, A.; Rasouli, S.; Soleimani, M.; Hashemi, S.M. Immunomodulatory effects of mesenchymal stem cell-derived exosomes on experimental type-1 autoimmune diabetes. J. Cell Biochem. 2018, 119, 9433–9443. [Google Scholar] [CrossRef] [PubMed]
- Kuljanin, M.; Bell, G.I.; Sherman, S.E.; Lajoie, G.A.; Hess, D.A. Proteomic characterisation reveals active Wnt-signalling by human multipotent stromal cells as a key regulator of beta cell survival and proliferation. Diabetologia 2017, 60, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Mesples, A.; Majeed, N.; Zhang, Y.; Hu, X. Early immunotherapy using autologous adult stem cells reversed the effect of anti-pancreatic islets in recently diagnosed type 1 diabetes mellitus: Preliminary results. Med. Sci. Monit. 2013, 19, 852–857. [Google Scholar] [CrossRef]
- Ezquer, F.; Ezquer, M.; Contador, D.; Ricca, M.; Simon, V.; Conget, P. The antidiabetic effect of mesenchymal stem cells is unrelated to their transdifferentiation potential but to their capability to restore Th1/Th2 balance and to modify the pancreatic microenvironment. Stem. Cells 2012, 30, 1664–1674. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem. Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Hao, H.; Tong, C.; Cheng, Y.; Liu, J.; Pang, Y.; Si, Y.; Guo, Y.; Zang, L.; Mu, Y.; et al. Human umbilical cord-derived mesenchymal stem cells elicit macrophages into an anti-inflammatory phenotype to alleviate insulin resistance in type 2 diabetic rats. Stem Cells 2016, 34, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Hao, H.; Cheng, Y.; Gao, J.; Liu, J.; Xie, Z.; Zhang, Q.; Zang, L.; Han, W.; Mu, Y. The homing of human umbilical cord-derived mesenchymal stem cells and the subsequent modulation of macrophage polarization in type 2 diabetic mice. Int. Immunopharmacol. 2018, 60, 235–245. [Google Scholar] [CrossRef]
- Rackham, C.L.; Amisten, S.; Persaud, S.J.; King, A.J.F.; Jones, P.M. Mesenchymal stromal cell secretory factors induce sustained improvements in islet function pre- and post-transplantation. Cytotherapy 2018, 20, 1427–1436. [Google Scholar] [CrossRef] [Green Version]
- Monfrini, M.; Donzelli, E.; Rodriguez-Menendez, V.; Ballarini, E.; Carozzi, V.A.; Chiorazzi, A.; Meregalli, C.; Canta, A.; Oggioni, N.; Crippa, L.; et al. Therapeutic potential of Mesenchymal Stem Cells for the treatment of diabetic peripheral neuropathy. Exp. Neurol. 2017, 288, 75–84. [Google Scholar] [CrossRef]
- Banerjee, M.; Kumar, A.; Bhonde, R.R. Reversal of experimental diabetes by multiple bone marrow transplantation. Biochem. Biophys. Res. Commun. 2005, 328, 318–325. [Google Scholar] [CrossRef]
- Domouky, A.M.; Hegab, A.S.; Al-Shahat, A.; Raafat, N. Mesenchymal stem cells and differentiated insulin producing cells are new horizons for pancreatic regeneration in type I diabetes mellitus. Int. J. Biochem. Cell Biol. 2017, 87, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.Y.; Zhang, Z.; Qian, G.S. Mesenchymal stem cells to treat diabetic neuropathy: A long and strenuous way from bench to the clinic. Cell Death Discov. 2016, 2, 16055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.R.; Zhang, N.K.; Zhang, Y.; Chen, Y.; Wang, L.; Zhu, Y.; Tang, H.H. Overexpression of apelin in Wharton’ jelly mesenchymal stem cell reverses insulin resistance and promotes pancreatic β cell proliferation in type 2 diabetic rats. Stem. Cell Res. Ther. 2018, 9, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdipour, E.; Salmasi, Z.; Sabeti, N. Potential of stem cell-derived exosomes to regenerate β islets through Pdx-1 dependent mechanism in a rat model of type 1 diabetes. J. Cell Physiol. 2019, 234, 20310–20321. [Google Scholar] [CrossRef]
- Montanari, E.; Meier, R.P.H.; Mahou, R.; Seebach, J.D.; Wandrey, C.; Gerber-Lemaire, S.; Buhler, L.H.; Gonelle-Gispert, C. Multipotent mesenchymal stromal cells enhance insulin secretion from human islets via N-cadherin interaction and prolong function of transplanted encapsulated islets in mice. Stem. Cell Res. Ther. 2017, 8, 199. [Google Scholar] [CrossRef]
- Rackham, C.L.; Dhadda, P.K.; Chagastelles, P.C.; Simpson, S.J.; Dattani, A.A.; Bowe, J.E.; Jones, P.M.; King, A.J. Pre-culturing islets with mesenchymal stromal cells using a direct contact configuration is beneficial for transplantation outcome in diabetic mice. Cytotherapy 2013, 15, 449–459. [Google Scholar] [CrossRef]
- Sun, Y.; Shi, H.; Yin, S.; Ji, C.; Zhang, X.; Zhang, B.; Wu, P.; Shi, Y.; Mao, F.; Yan, Y.; et al. Human Mesenchymal Stem Cell Derived Exosomes Alleviate Type 2 Diabetes Mellitus by Reversing Peripheral Insulin Resistance and Relieving β-Cell Destruction. ACS Nano 2018, 12, 7613–7628. [Google Scholar] [CrossRef]
- Xiang, C.; Xie, Q.P. Protection of mouse pancreatic islet function by co-culture with hypoxia pre-treated mesenchymal stromal cells. Mol. Med. Rep. 2018, 18, 2589–2598. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Xu, C.; Zhou, S.; Liu, X.; Wang, J.; Qian, S.; Xin, Y.; Gao, Y.; Zhu, Y.; Tang, X. PAX4 promotes PDX1-induced differentiation of mesenchymal stem cells into insulin-secreting cells. Am. J. Transl. Res. 2017, 9, 874–886. [Google Scholar]
- Bader, A.M.; Klose, K.; Bieback, K.; Korinth, D.; Schneider, M.; Seifert, M.; Choi, Y.H.; Kurtz, A.; Falk, V.; Stamm, C. Hypoxic Preconditioning Increases Survival and Pro-Angiogenic Capacity of Human Cord Blood Mesenchymal Stromal Cells In Vitro. PLoS ONE 2015, 10, e0138477. [Google Scholar] [CrossRef]
- Xu, T.; Lv, Z.; Chen, Q.; Guo, M.; Wang, X.; Huang, F. Vascular endothelial growth factor over-expressed mesenchymal stem cells-conditioned media ameliorate palmitate-induced diabetic endothelial dysfunction through PI-3K/AKT/m-TOR/eNOS and p38/MAPK signaling pathway. Biomed. Pharmacother 2018, 106, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, S.; Yoo, K.H.; Sym, S.J.; Khang, D. Mesenchymal stem cell therapy assisted by nanotechnology: A possible combinational treatment for brain tumor and central nerve regeneration. Int. J. Nanomedicine 2019, 14, 5925–5942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arzouni, A.A.; Vargas-Seymour, A.; Dhadda, P.K.; Rackham, C.L.; Huang, G.C.; Choudhary, P.; King, A.J.F.; Jones, P.M. Characterization of the Effects of Mesenchymal Stromal Cells on Mouse and Human Islet Function. Stem. Cells Transl. Med. 2019, 8, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borg, D.J.; Welzel, P.B.; Grimmer, M.; Friedrichs, J.; Weigelt, M.; Wilhelm, C.; Prewitz, M.; Stißel, A.; Hommel, A.; Kurth, T.; et al. Macroporous biohybrid cryogels for co-housing pancreatic islets with mesenchymal stromal cells. Acta Biomater 2016, 44, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, P.O.; Schwarcz, E.; Korsgren, O.; Le Blanc, K. Preserved β-cell function in type 1 diabetes by mesenchymal stromal cells. Diabetes 2015, 64, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Sbano, P.; Cuccia, A.; Mazzanti, B.; Urbani, S.; Giusti, B.; Lapini, I.; Rossi, L.; Abbate, R.; Marseglia, G.; Nannetti, G.; et al. Use of donor bone marrow mesenchymal stem cells for treatment of skin allograft rejection in a preclinical rat model. Arch. Dermatol. Res. 2008, 300, 115–124. [Google Scholar] [CrossRef] [Green Version]
- van de Vyver, M. Intrinsic Mesenchymal Stem Cell Dysfunction in Diabetes Mellitus: Implications for Autologous Cell Therapy. Stem. Cells Dev. 2017, 26, 1042–1053. [Google Scholar] [CrossRef]
- Kornicka, K.; Houston, J.; Marycz, K. Dysfunction of Mesenchymal Stem Cells Isolated from Metabolic Syndrome and Type 2 Diabetic Patients as Result of Oxidative Stress and Autophagy may Limit Their Potential Therapeutic Use. Stem Cell Rev. Rep. 2018, 14, 337–345. [Google Scholar] [CrossRef] [Green Version]
- El-Badawy, A.; El-Badri, N. Clinical Efficacy of Stem Cell Therapy for Diabetes Mellitus: A Meta-Analysis. PLoS ONE 2016, 11, e0151938. [Google Scholar] [CrossRef] [Green Version]
- Foudah, D.; Redaelli, S.; Donzelli, E.; Bentivegna, A.; Miloso, M.; Dalpra, L.; Tredici, G. Monitoring the genomic stability of in vitro cultured rat bone-marrow-derived mesenchymal stem cells. Chromosome Res. 2009, 17, 1025–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tappenbeck, N.; Schröder, H.M.; Niebergall-Roth, E.; Hassinger, F.; Dehio, U.; Dieter, K.; Kraft, K.; Kerstan, A.; Esterlechner, J.; Frank, N.Y.; et al. In vivo safety profile and biodistribution of GMP-manufactured human skin-derived ABCB5-positive mesenchymal stromal cells for use in clinical trials. Cytotherapy 2019, 21, 546–560. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Authors | Source of MSCs | Application | Suggested Mechanism |
|---|---|---|---|
| Banerjee et al., 2005 [30] | Mouse BM-MSCs | T1D | Islet regeneration |
| Domouky et al., 2017 [31] | Rat BM-MSCs | T1D | Islet regeneration |
| Ezquer et al., 2012 [24] | Mouse BM-MSCs | T1D | |
| Gao et al., 2018 [33] | Human Wharton’s jelly MSCs | T2D | Immunomodulatory effect |
| Kuljanin et al., 2017 [22] | Human BM-MSCs | T1D | Immunomodulatory effect |
| Mahdipour et al., 2019 [34] | Human menstrual blood-derived MSCs | T1D | Islet regeneration |
| Monfrini et al., 2017 [29] | Rat BM-MSCs | T1D | Islet regeneration |
| Montanari et al., 2017 [35] | Human BM-MSCs | T1D | Increased Islet function |
| Nojehdehi et al., 2018 [21] | Mouse Adipose MSCs | T1D | Islet regeneration |
| Rackham et al., 2013 [36] | Mouse Adipose MSCs | T1D | Increased Islet function |
| Rackham et al., 2018 [28] | Mouse Adipose MSCs | T1D | Islet regeneration |
| Sun et al., 2018 [37] | Human umbilical cord-derived MSCs | T2D | Increased Islet function |
| Xiang et al., 2018 [38] | Mouse BM-MSCs | T1D | Islet regeneration |
| Xie et al., 2016 [26] | Human umbilical cord-derived MSCs | T2D | Increased Islet function |
| Yin et al., 2018 [27] | Human umbilical cord-derived MSCs | T2D | Immunomodulatory effect |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donzelli, E.; Scuteri, A. Mesenchymal Stem Cells: A Trump Card for the Treatment of Diabetes? Biomedicines 2020, 8, 112. https://doi.org/10.3390/biomedicines8050112
Donzelli E, Scuteri A. Mesenchymal Stem Cells: A Trump Card for the Treatment of Diabetes? Biomedicines. 2020; 8(5):112. https://doi.org/10.3390/biomedicines8050112
Chicago/Turabian StyleDonzelli, Elisabetta, and Arianna Scuteri. 2020. "Mesenchymal Stem Cells: A Trump Card for the Treatment of Diabetes?" Biomedicines 8, no. 5: 112. https://doi.org/10.3390/biomedicines8050112