
Rehabilitation Training after Spinal Cord Injury Affects Brain Structure and Function: From Mechanisms to Methods

Abstract
:1. Introduction
2. Effect of Rehabilitation Training on Brain Structure and Function
2.1. Physiological Mechanism of Exercise and Brain Structure and Function
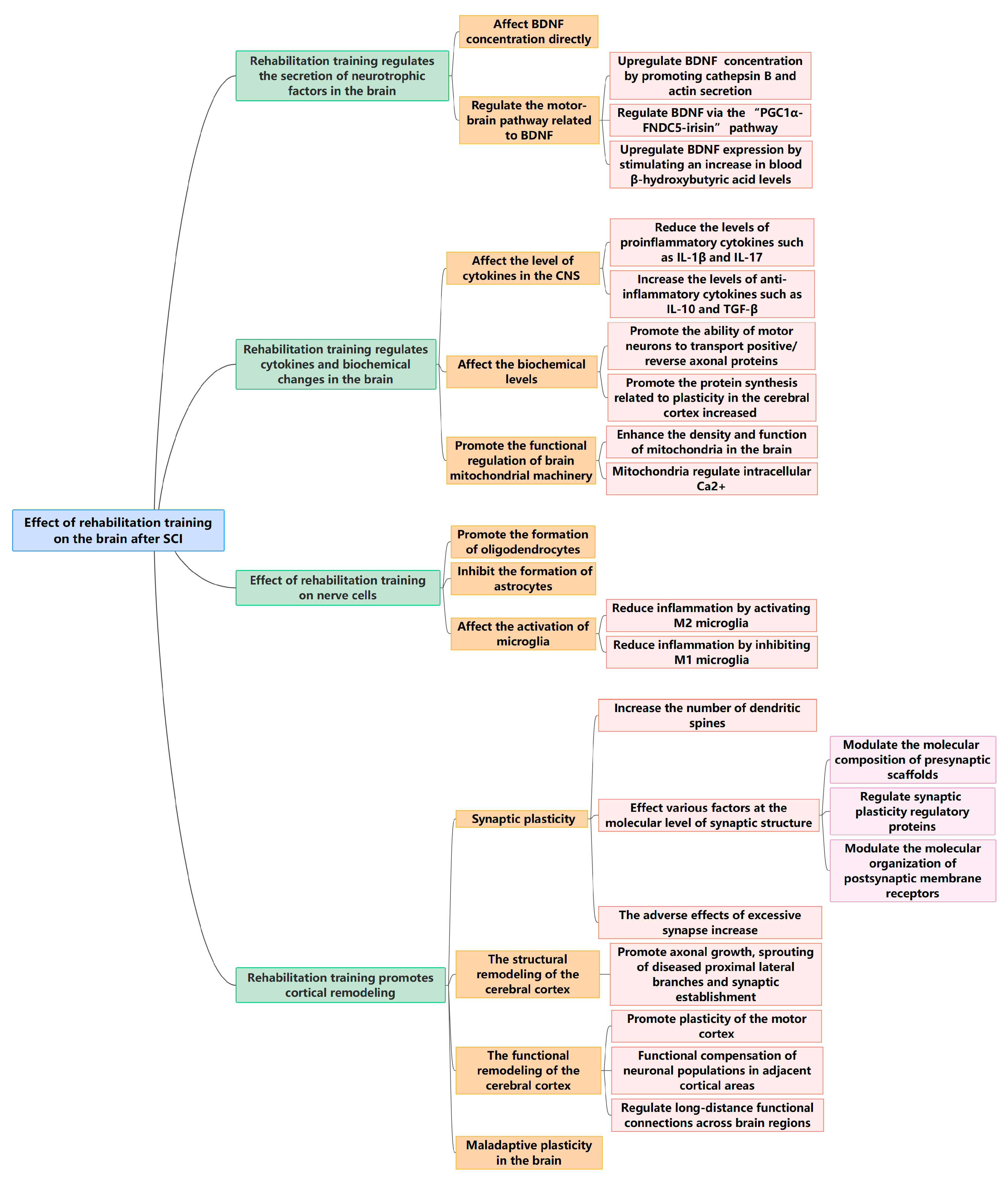
2.2. Effect of Rehabilitation Training on the Brain after SCI
2.2.1. Rehabilitation Training Regulates the Secretion of Neurotrophic Factors in the Brain
2.2.2. Rehabilitation Training Regulates Cytokines and Biochemical Changes in the Brain
2.2.3. Effect of Rehabilitation Training on Nerve Cells
Oligodendrocyte
Astrocytes
Microglia
2.2.4. Rehabilitation Training Promotes Cortical Remodeling
Synaptic Plasticity
Effect of Rehabilitation Training on the Structural Remodeling of the Cerebral Cortex
Effect of Rehabilitation Training on Functional Remodeling of the Cerebral Cortex
Maladaptive Plasticity in the Brain
2.2.5. Summary of This Section
3. Different Rehabilitation Techniques and Their Effects on Brain Reorganization after SCI
3.1. Rehabilitation Exercise Regulates the Brain
3.2. Epidural Electrical Stimulation Regulates the Brain
3.3. Exoskeleton Rehabilitation Robot Regulates Brain
3.4. Motor Imagination Rehabilitation Regulates the Brain
3.5. Comparison of the Advantages and Disadvantages of Different Rehabilitation Methods
4. Summary and Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Gross-Hemmi, M.H.; Post, M.W.; Ehrmann, C.; Fekete, C.; Hasnan, N.; Middleton, J.W.; Reinhardt, J.D.; Strom, V.; Stucki, G. Study Protocol of the International Spinal Cord Injury (InSCI) Community Survey. Am. J. Phys. Med. Rehabil. 2017, 96, S23–S34. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Bao, S.S.; Xu, M.; Rao, J.S. Importance of brain alterations in spinal cord injury. Sci. Prog. 2021, 104, 368504211031117. [Google Scholar] [CrossRef] [PubMed]
- Northcutt, R.G. Accommodations of the Nervous System: Body and Brain. A Trophic Theory of Neural Connections. Dale Purves. Harvard University Press, Cambridge, MA, 1988. viii, 231 pp., illus. $35. Science 1989, 244, 993. [Google Scholar] [CrossRef]
- Bilchak, J.N.; Caron, G.; Côté, M.P. Exercise-Induced Plasticity in Signaling Pathways Involved in Motor Recovery after Spinal Cord Injury. Int. J. Mol. Sci. 2021, 22, 4858. [Google Scholar] [CrossRef] [PubMed]
- Henderson, L.A.; Gustin, S.M.; Macey, P.M.; Wrigley, P.J.; Siddall, P.J. Functional reorganization of the brain in humans following spinal cord injury: Evidence for underlying changes in cortical anatomy. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 2630–2637. [Google Scholar] [CrossRef]
- Nandakumar, B.; Blumenthal, G.H.; Disse, G.D.; Desmond, P.C.; Ebinu, J.O.; Ricard, J.; Bethea, J.R.; Moxon, K.A. Exercise therapy guides cortical reorganization after midthoracic spinal contusion to enhance control of lower thoracic muscles, supporting functional recovery. Exp. Neurol. 2023, 364, 114394. [Google Scholar] [CrossRef]
- Lourenco, M.V.; Frozza, R.L.; de Freitas, G.B.; Zhang, H.; Kincheski, G.C.; Ribeiro, F.C.; Goncalves, R.A.; Clarke, J.R.; Beckman, D.; Staniszewski, A.; et al. Exercise-linked FNDC5/irisin rescues synaptic plasticity and memory defects in Alzheimer’s models. Nat. Med. 2019, 25, 165–175. [Google Scholar] [CrossRef]
- Noakes, T.; Spedding, M. Olympics: Run for your life. Nature 2012, 487, 295–296. [Google Scholar] [CrossRef]
- Kobilo, T.; Liu, Q.R.; Gandhi, K.; Mughal, M.; Shaham, Y.; van Praag, H. Running is the neurogenic and neurotrophic stimulus in environmental enrichment. Learn. Mem. 2011, 18, 605–609. [Google Scholar] [CrossRef]
- Mu, L.; Cai, J.; Gu, B.; Yu, L.; Li, C.; Liu, Q.S.; Zhao, L. Treadmill Exercise Prevents Decline in Spatial Learning and Memory in 3xTg-AD Mice through Enhancement of Structural Synaptic Plasticity of the Hippocampus and Prefrontal Cortex. Cells 2022, 11, 244. [Google Scholar] [CrossRef]
- Shen, W.; Jin, L.; Zhu, A.; Lin, Y.; Pan, G.; Zhou, S.; Cheng, J.; Zhang, J.; Tu, F.; Liu, C.; et al. Treadmill exercise enhances synaptic plasticity in the ischemic penumbra of MCAO mice by inducing the expression of Camk2a via CYFIP1 upregulation. Life Sci. 2021, 270, 119033. [Google Scholar] [CrossRef]
- Batouli, S.A.H.; Saba, V. At least eighty percent of brain grey matter is modifiable by physical activity: A review study. Behav. Brain Res. 2017, 332, 204–217. [Google Scholar] [CrossRef]
- Jurkiewicz, M.T.; Mikulis, D.J.; McIlroy, W.E.; Fehlings, M.G.; Verrier, M.C. Sensorimotor cortical plasticity during recovery following spinal cord injury: A longitudinal fMRI study. Neurorehabilit. Neural Repair. 2007, 21, 527–538. [Google Scholar] [CrossRef]
- Pedersen, B.K. Physical activity and muscle-brain crosstalk. Nat. Rev. Endocrinol. 2019, 15, 383–392. [Google Scholar] [CrossRef]
- Giudice, J.; Taylor, J.M. Muscle as a paracrine and endocrine organ. Curr. Opin. Pharmacol. 2017, 34, 49–55. [Google Scholar] [CrossRef]
- Hoffmann, C.; Weigert, C. Skeletal Muscle as an Endocrine Organ: The Role of Myokines in Exercise Adaptations. Cold Spring Harb. Perspect. Med. 2017, 7, a029793. [Google Scholar] [CrossRef] [PubMed]
- Pesce, M.; La Fratta, I.; Paolucci, T.; Grilli, A.; Patruno, A.; Agostini, F.; Bernetti, A.; Mangone, M.; Paoloni, M.; Invernizzi, M.; et al. From Exercise to Cognitive Performance: Role of Irisin. Appl. Sci. 2021, 11, 7120. [Google Scholar] [CrossRef]
- Severinsen, M.C.K.; Pedersen, B.K. Muscle-Organ Crosstalk: The Emerging Roles of Myokines. Endocr. Rev. 2020, 41, 594–609. [Google Scholar] [CrossRef]
- Kowianski, P.; Lietzau, G.; Czuba, E.; Waskow, M.; Steliga, A.; Morys, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- El Hayek, L.; Khalifeh, M.; Zibara, V.; Assaad, R.A.; Emmanuel, N.; Karnib, N.; El-Ghandour, R.; Nasrallah, P.; Bilen, M.; Ibrahim, P.; et al. Lactate Mediates the Effects of Exercise on Learning and Memory through SIRT1-Dependent Activation of Hippocampal Brain-Derived Neurotrophic Factor (BDNF). J. Neurosci. 2019, 39, 2369–2382. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, D.; Oliveira-Lima, O.C.; da Silva, T.V.; Faraco, C.C.F.; Leite, H.R.; Juliano, M.A.; dos Santos, D.M.; Bethea, J.R.; Brambilla, R.; Orian, J.M.; et al. Differential brain and spinal cord cytokine and BDNF levels in experimental autoimmune encephalomyelitis are modulated by prior and regular exercise. J. Neuroimmunol. 2013, 264, 24–34. [Google Scholar] [CrossRef]
- Ozkul, C.; Guclu-Gunduz, A.; Irkec, C.; Fidan, I.; Aydin, Y.; Ozkan, T.; Yazici, G. Effect of combined exercise training on serum brain-derived neurotrophic factor, suppressors of cytokine signaling 1 and 3 in patients with multiple sclerosis. J. Neuroimmunol. 2018, 316, 121–129. [Google Scholar] [CrossRef]
- Pajonk, F.G.; Wobrock, T.; Gruber, O.; Scherk, H.; Berner, D.; Kaizl, I.; Kierer, A.; Muller, S.; Oest, M.; Meyer, T.; et al. Hippocampal plasticity in response to exercise in schizophrenia. Arch. Gen. Psychiatry 2010, 67, 133–143. [Google Scholar] [CrossRef]
- Wens, I.; Keytsman, C.; Deckx, N.; Cools, N.; Dalgas, U.; Eijnde, B.O. Brain derived neurotrophic factor in multiple sclerosis: Effect of 24 weeks endurance and resistance training. Eur. J. Neurol. 2016, 23, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Q.; Xie, C.; Wang, C.; Wang, Q.; Dong, C.; Fang, L.; Ding, J.; Wang, T. Blocking of BDNF-TrkB signaling inhibits the promotion effect of neurological function recovery after treadmill training in rats with spinal cord injury. Spinal Cord 2019, 57, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.Y.; Becke, A.; Berron, D.; Becker, B.; Sah, N.; Benoni, G.; Janke, E.; Lubejko, S.T.; Greig, N.H.; Mattison, J.A.; et al. Running-Induced Systemic Cathepsin B Secretion Is Associated with Memory Function. Cell Metab. 2016, 24, 332–340. [Google Scholar] [CrossRef]
- Chazeau, A.; Giannone, G. Organization and dynamics of the actin cytoskeleton during dendritic spine morphological remodeling. Cell. Mol. Life Sci. 2016, 73, 3053–3073. [Google Scholar] [CrossRef] [PubMed]
- Neasta, J.; Fiorenza, A.; He, D.Y.; Phamluong, K.; Kiely, P.A.; Ron, D. Activation of the cAMP Pathway Induces RACK1-Dependent Binding of β-Actin to BDNF Promoter. PLoS ONE 2016, 11, e0160948. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Farshbaf, M.J.; Ghaedi, K.; Megraw, T.L.; Curtiss, J.; Faradonbeh, M.S.; Vaziri, P.; Nasr-Esfahani, M.H. Does PGC1α/FNDC5/BDNF Elicit the Beneficial Effects of Exercise on Neurodegenerative Disorders? Neuromolecular Med. 2016, 18, 1–15. [Google Scholar] [CrossRef]
- Sleiman, S.F.; Henry, J.; Al-Haddad, R.; El Hayek, L.; Abou Haidar, E.; Stringer, T.; Ulja, D.; Karuppagounder, S.S.; Holson, E.B.; Ratan, R.R.; et al. Exercise promotes the expression of brain derived neurotrophic factor (BDNF) through the action of the ketone body beta-hydroxybutyrate. elife 2016, 5, e15092. [Google Scholar] [CrossRef]
- Komine, S.; Akiyama, K.; Warabi, E.; Oh, S.; Kuga, K.; Ishige, K.; Togashi, S.; Yanagawa, T.; Shoda, J. Exercise training enhances in vivo clearance of endotoxin and attenuates inflammatory responses by potentiating Kupffer cell phagocytosis. Sci. Rep. 2017, 7, 11977. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, D.; Brambilla, R.; Bracchi-Ricard, V.; Karmally, S.; Dellarole, A.; Carvalho-Tavares, J.; Bethea, J.R. Prior regular exercise improves clinical outcome and reduces demyelination and axonal injury in experimental autoimmune encephalomyelitis. J. Neurochem. 2016, 136 (Suppl. 1), 63–73. [Google Scholar] [CrossRef]
- Einstein, O.; Fainstein, N.; Touloumi, O.; Lagoudaki, R.; Hanya, E.; Grigoriadis, N.; Katz, A.; Ben-Hur, T. Exercise training attenuates experimental autoimmune encephalomyelitis by peripheral immunomodulation rather than direct neuroprotection. Exp. Neurol. 2018, 299, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Pryor, W.M.; Freeman, K.G.; Larson, R.D.; Edwards, G.L.; White, L.J. Chronic exercise confers neuroprotection in experimental autoimmune encephalomyelitis. J. Neurosci. Res. 2015, 93, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Nichol, K.E.; Poon, W.W.; Parachikova, A.I.; Cribbs, D.H.; Glabe, C.G.; Cotman, C.W. Exercise alters the immune profile in Tg2576 Alzheimer mice toward a response coincident with improved cognitive performance and decreased amyloid. J. Neuroinflammation 2008, 5, 13. [Google Scholar] [CrossRef]
- Lee, J.H.; Han, J.H.; Woo, J.H.; Jou, I. 25-Hydroxycholesterol suppress IFN-γ-induced inflammation in microglia by disrupting lipid raft formation and caveolin-mediated signaling endosomes. Free Radic. Biol. Med. 2022, 179, 252–265. [Google Scholar] [CrossRef]
- Cheng, T.; Huang, X.D.; Hu, X.F.; Wang, S.Q.; Chen, K.; Wei, J.A.; Yan, L.; So, W.A.; Yuan, T.F.; Zhang, L. Physical exercise rescues cocaine-evoked synaptic deficits in motor cortex. Mol. Psychiatry 2021, 26, 6187–6197. [Google Scholar] [CrossRef]
- Thomas, B.P.; Tarumi, T.; Sheng, M.; Tseng, B.; Womack, K.B.; Cullum, C.M.; Rypma, B.; Zhang, R.; Lu, H. Brain Perfusion Change in Patients with Mild Cognitive Impairment After 12 Months of Aerobic Exercise Training. J. Alzheimers Dis. 2020, 75, 617–631. [Google Scholar] [CrossRef]
- Dietrich, M.O.; Andrews, Z.B.; Horvath, T.L. Exercise-induced synaptogenesis in the hippocampus is dependent on UCP2-regulated mitochondrial adaptation. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 10766–10771. [Google Scholar] [CrossRef]
- Bayod, S.; del Valle, J.; Canudas, A.M.; Lalanza, J.F.; Sanchez-Roige, S.; Camins, A.; Escorihuela, R.M.; Pallàs, M. Long-term treadmill exercise induces neuroprotective molecular changes in rat brain. J. Appl. Physiol. 2011, 111, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. Dietary restriction, glycolysis, hormesis and ageing. Biogerontology 2007, 8, 221–224. [Google Scholar] [CrossRef]
- Cantó, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef]
- Marton, O.; Koltai, E.; Nyakas, C.; Bakonyi, T.; Zenteno-Savin, T.; Kumagai, S.; Goto, S.; Radak, Z. Aging and exercise affect the level of protein acetylation and SIRT1 activity in cerebellum of male rats. Biogerontology 2010, 11, 679–686. [Google Scholar] [CrossRef]
- Ravera, S.; Morelli, A.M.; Panfoli, I. Myelination increases chemical energy support to the axon without modifying the basic physicochemical mechanism of nerve conduction. Neurochem. Int. 2020, 141, 104883. [Google Scholar] [CrossRef] [PubMed]
- Markovinovic, A.; Greig, J.; Martín-Guerrero, S.M.; Salam, S.; Paillusson, S. Endoplasmic reticulum-mitochondria signaling in neurons and neurodegenerative diseases. J. Cell Sci. 2022, 135, jcs248534. [Google Scholar] [CrossRef]
- Orr, M.B.; Gensel, J.C. Spinal Cord Injury Scarring and Inflammation: Therapies Targeting Glial and Inflammatory Responses. Neurotherapeutics 2018, 15, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Liu, X.K.; Li, R.; Zhang, P.; Chu, Z.; Wang, C.L.; Liu, H.R.; Qi, J.; Lv, G.Y.; Wang, G.Y.; et al. Effect of glial cells on remyelination after spinal cord injury. Neural Regen. Res. 2017, 12, 1724–1732. [Google Scholar] [CrossRef]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the central nervous system: Structure, function, and pathology. Physiol. Rev. 2019, 99, 1381–1431. [Google Scholar] [CrossRef]
- McKenzie, I.A.; Ohayon, D.; Li, H.; de Faria, J.P.; Emery, B.; Tohyama, K.; Richardson, W.D. Motor skill learning requires active central myelination. Science 2014, 346, 318–322. [Google Scholar] [CrossRef]
- Zheng, J.; Sun, X.; Ma, C.; Li, B.-m.; Luo, F. Voluntary wheel running promotes myelination in the motor cortex through Wnt signaling in mice. Mol. Brain 2019, 12, 85. [Google Scholar] [CrossRef]
- Kim, T.W.; Sung, Y.H. Regular exercise promotes memory function and enhances hippocampal neuroplasticity in experimental autoimmune encephalomyelitis mice. Neuroscience 2017, 346, 173–181. [Google Scholar] [CrossRef]
- Krityakiarana, W.; Espinosa-Jeffrey, A.; Ghiani, C.A.; Zhao, P.M.; Topaldjikian, N.; Gomez-Pinilla, F.; Yamaguchi, M.; Kotchabhakdi, N.; de Vellis, J. Voluntary exercise increases oligodendrogenesis in spinal cord. Int. J. Neurosci. 2010, 120, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Agata, V.; Magrì, B.; Roggio, F.; Castorina, A.; Ravalli, S.; Di Rosa, M.; Musumeci, G. Neuroprotective Effects of Physical Activity via the Adaptation of Astrocytes. Cells 2021, 10, 1542. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zuo, Y.X.; Jiang, R.T. Astrocyte morphology: Diversity, plasticity, and role in neurological diseases. CNS Neurosci. Ther. 2019, 25, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J.; Eroglu, C. Cell Biology of Astrocyte-Synapse Interactions. Neuron 2017, 96, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, Y.; Wang, Y.; Song, L.; Zhang, R.; Du, Y. Long-term treadmill exercise attenuates Aβ burdens and astrocyte activation in APP/PS1 mouse model of Alzheimer’s disease. Neurosci. Lett. 2018, 666, 70–77. [Google Scholar] [CrossRef]
- Lee, S.U.; Kim, D.Y.; Park, S.H.; Choi, D.H.; Park, H.W.; Han, T.R. Mild to moderate early exercise promotes recovery from cerebral ischemia in rats. Can. J. Neurol. Sci. 2009, 36, 443–449. [Google Scholar] [CrossRef]
- Fiuza-Luces, C.; Santos-Lozano, A.; Joyner, M.; Carrera-Bastos, P.; Picazo, O.; Zugaza, J.L.; Izquierdo, M.; Ruilope, L.M.; Lucia, A. Exercise benefits in cardiovascular disease: Beyond attenuation of traditional risk factors. Nat. Rev. Cardiol. 2018, 15, 731–743. [Google Scholar] [CrossRef]
- Chu, F.; Shi, M.; Zheng, C.; Shen, D.; Zhu, J.; Zheng, X.; Cui, L. The roles of macrophages and microglia in multiple sclerosis and experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2018, 318, 1–7. [Google Scholar] [CrossRef]
- Kohman, R.A.; Bhattacharya, T.K.; Wojcik, E.; Rhodes, J.S. Exercise reduces activation of microglia isolated from hippocampus and brain of aged mice. J. Neuroinflammation 2013, 10, 114. [Google Scholar] [CrossRef]
- Benson, C.; Paylor, J.W.; Tenorio, G.; Winship, I.; Baker, G.; Kerr, B.J. Voluntary wheel running delays disease onset and reduces pain hypersensitivity in early experimental autoimmune encephalomyelitis (EAE). Exp. Neurol. 2015, 271, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.K.; Michaels, N.J.; Ilyntskyy, S.; Keough, M.B.; Kovalchuk, O.; Yong, V.W. Multimodal Enhancement of Remyelination by Exercise with a Pivotal Role for Oligodendroglial PGC1α. Cell Rep. 2018, 24, 3167–3179. [Google Scholar] [CrossRef] [PubMed]
- Clark, T.A.; Sullender, C.; Jacob, D.; Zuo, Y.; Dunn, A.K.; Jones, T.A. Rehabilitative Training Interacts with Ischemia-Instigated Spine Dynamics to Promote a Lasting Population of New Synapses in Peri-Infarct Motor Cortex. J. Neurosci. Off. J. Soc. Neurosci. 2019, 39, 8471–8483. [Google Scholar] [CrossRef]
- Dietrich, A.; McDaniel, W.F. Endocannabinoids and exercise. Br. J. Sports Med. 2004, 38, 536–541. [Google Scholar] [CrossRef]
- Opendak, M.; Gould, E. Adult neurogenesis: A substrate for experience-dependent change. Trends Cogn. Sci. 2015, 19, 151–161. [Google Scholar] [CrossRef]
- Houle, J.D.; Côté, M.P. Axon regeneration and exercise-dependent plasticity after spinal cord injury. Ann. N. Y. Acad. Sci. 2013, 1279, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.A.-O.; Lozinski, B.; Yong, V.A.-O. Exercise in multiple sclerosis and its models: Focus on the central nervous system outcomes. J. Neurosci. Res. 2020, 98, 509–523. [Google Scholar] [CrossRef]
- Cassilhas, R.; Lee, K.; Fernandes, J.; Oliveira, M.; Tufik, S.; Meeusen, R.; de Mello, M. Spatial memory is improved by aerobic and resistance exercise through divergent molecular mechanisms. Neuroscience 2012, 202, 309–317. [Google Scholar] [CrossRef]
- Lista, I.; Sorrentino, G. Biological mechanisms of physical activity in preventing cognitive decline. Cell Mol. Neurobiol. 2010, 30, 493–503. [Google Scholar] [CrossRef]
- Oestreicher, A.B.; De Graan, P.N.; Gispen, W.H.; Verhaagen, J.; Schrama, L.H. B-50, the growth associated protein-43: Modulation of cell morphology and communication in the nervous system. Prog. Neurobiol. 1997, 53, 627–686. [Google Scholar] [CrossRef]
- Mizutani, K.; Sonoda, S.; Yamada, K.; Beppu, H.; Shimpo, K. Alteration of protein expression profile following voluntary exercise in the perilesional cortex of rats with focal cerebral infarction. Brain Res. 2011, 1416, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Thiele, C.; Hannah, M.J.; Fahrenholz, F.; Huttner, W.B. Cholesterol binds to synaptophysin and is required for biogenesis of synaptic vesicles. Nat. Cell Biol. 2000, 2, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Rapp, S.; Baader, M.; Hu, M.; Jennen-Steinmetz, C.; Henn, F.A.; Thome, J. Differential regulation of synaptic vesicle proteins by antidepressant drugs. Pharmacogenomics J. 2004, 4, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ke, C.K.; Su, Y.; Wan, C.X. Exercise Intervention Promotes the Growth of Synapses and Regulates Neuroplasticity in Rats with Ischemic Stroke Through Exosomes. Front. Neurol. 2021, 12, 752595. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.M.; Gomez-Puertas, P.; Tümer, Z. Neurodevelopmental Disorders Associated with PSD-95 and Its Interaction Partners. Int. J. Mol. Sci. 2022, 23, 4390. [Google Scholar] [CrossRef] [PubMed]
- Pagnussat, A.S.; Simao, F.; Anastacio, J.R.; Mestriner, R.G.; Michaelsen, S.M.; Castro, C.C.; Salbego, C.; Netto, C.A. Effects of skilled and unskilled training on functional recovery and brain plasticity after focal ischemia in adult rats. Brain Res. 2012, 1486, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.-C.; Yang, Y.-R.; Wang, R.-Y. Effects of exercise intensity on spatial memory performance and hippocampal synaptic plasticity in transient brain ischemic rats. PLoS ONE 2013, 8, e78163. [Google Scholar] [CrossRef]
- Sala, C.; Piech, V.; Wilson, N.R.; Passafaro, M.; Liu, G.; Sheng, M. Regulation of dendritic spine morphology and synaptic function by Shank and Homer. Neuron 2001, 31, 115–130. [Google Scholar] [CrossRef]
- Xu, B.; Zang, K.; Ruff, N.L.; Zhang, Y.A.; McConnell, S.K.; Stryker, M.P.; Reichardt, L.F. Cortical degeneration in the absence of neurotrophin signaling: Dendritic retraction and neuronal loss after removal of the receptor TrkB. Neuron 2000, 26, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Bramham, C.R.; Wells, D.G. Dendritic mRNA: Transport, translation and function. Nat. Rev. Neurosci. 2007, 8, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Andreska, T.; Lüningschrör, P.; Sendtner, M. Regulation of TrkB cell surface expression-a mechanism for modulation of neuronal responsiveness to brain-derived neurotrophic factor. Cell Tissue Res. 2020, 382, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Palasz, E.; Wysocka, A.; Gasiorowska, A.; Chalimoniuk, M.; Niewiadomski, W.; Niewiadomska, G. BDNF as a Promising Therapeutic Agent in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 1170. [Google Scholar] [CrossRef]
- Yasuda, R.; Hayashi, Y.; Hell, J.W. CaMKII: A central molecular organizer of synaptic plasticity, learning and memory. Nat. Rev. Neurosci. 2022, 23, 666–682. [Google Scholar] [CrossRef] [PubMed]
- Langnaese, K.; Seidenbecher, C.; Wex, H.; Seidel, B.; Hartung, K.; Appeltauer, U.; Garner, A.; Voss, B.; Mueller, B.; Garner, C.C.; et al. Protein components of a rat brain synaptic junctional protein preparation. Brain Res. Mol. Brain Res. 1996, 42, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Fiorenza, M.; Gunnarsson, T.P.; Hostrup, M.; Iaia, F.M.; Schena, F.; Pilegaard, H.; Bangsbo, J. Metabolic stress-dependent regulation of the mitochondrial biogenic molecular response to high-intensity exercise in human skeletal muscle. J. Physiol. Lond. 2018, 596, 2823–2840. [Google Scholar] [CrossRef]
- Skovgaard, C.; Almquist, N.W.; Bangsbo, J. The effect of repeated periods of speed endurance training on performance, running economy, and muscle adaptations. Scand. J. Med. Sci. Sports 2018, 28, 381–390. [Google Scholar] [CrossRef]
- Liu, W.; Xia, Y.; Kuang, H.; Wang, Z.; Liu, S.; Tang, C.; Yin, D. Proteomic Profile of Carbonylated Proteins Screen the Regulation of Calmodulin-Dependent Protein Kinases-AMPK-Beclin1 in Aerobic Exercise-Induced Autophagy in Middle-Aged Rat Hippocampus. Gerontology 2019, 65, 620–633. [Google Scholar] [CrossRef]
- Xing, Y.; Bai, Y.A.-O. A Review of Exercise-Induced Neuroplasticity in Ischemic Stroke: Pathology and Mechanisms. Mol. Neurobiol. 2020, 57, 4218–4231. [Google Scholar] [CrossRef]
- Schwenk, J.; Boudkkazi, S.; Kocylowski, M.K.; Brechet, A.; Zolles, G.; Bus, T.; Costa, K.; Kollewe, A.; Jordan, J.; Bank, J.; et al. An ER Assembly Line of AMPA-Receptors Controls Excitatory Neurotransmission and Its Plasticity. Neuron 2019, 104, 680–692.e689. [Google Scholar] [CrossRef]
- Real, C.C.; Ferreira, A.F.; Hernandes, M.S.; Britto, L.R.; Pires, R.S. Exercise-induced plasticity of AMPA-type glutamate receptor subunits in the rat brain. Brain Res. 2010, 1363, 63–71. [Google Scholar] [CrossRef]
- Kintz, N.; Petzinger, G.M.; Akopian, G.; Ptasnik, S.; Williams, C.; Jakowec, M.W.; Walsh, J.P. Exercise modifies alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor expression in striatopallidal neurons in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse. J. Neurosci. Res. 2013, 91, 1492–1507. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Wang, D.; Duan, Y.C.; Zhai, L.; Shi, L.; Guo, F. Aerobic Exercise Prevents Depression via Alleviating Hippocampus Injury in Chronic Stressed Depression Rats. Brain Sci. 2021, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.W.; Seo, J.H.; Baek, S.B.; Kim, C.J.; Kim, T.W. Treadmill exercise inhibits hippocampal apoptosis through enhancing N-methyl-D-aspartate receptor expression in the MK-801-induced schizophrenic mice. J. Exerc. Rehabil. 2014, 10, 218–224. [Google Scholar] [CrossRef]
- Tang, G.; Gudsnuk, K.; Kuo, S.H.; Cotrina, M.L.; Rosoklija, G.; Sosunov, A.; Sonders, M.S.; Kanter, E.; Castagna, C.; Yamamoto, A.; et al. Loss of mTOR-dependent macroautophagy causes autistic-like synaptic pruning deficits. Neuron 2014, 83, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Obi-Nagata, K.; Suzuki, N.; Miyake, R.; MacDonald, M.L.; Fish, K.N.; Ozawa, K.; Nagahama, K.; Okimura, T.; Tanaka, S.; Kano, M.; et al. Distorted neurocomputation by a small number of extra-large spines in psychiatric disorders. Sci. Adv. 2023, 9, eade5973. [Google Scholar] [CrossRef] [PubMed]
- Freund, P.; Weiskopf, N.; Ward, N.S.; Hutton, C.; Gall, A.; Ciccarelli, O.; Craggs, M.; Friston, K.; Thompson, A.J.; Freund, P.; et al. Disability, atrophy and cortical reorganization following spinal cord injury. Brain J. Neurol. 2011, 134, 1610–1622. [Google Scholar] [CrossRef] [PubMed]
- Jurkiewicz, M.T.; Crawley, A.P.; Verrier, M.C.; Fehlings, M.G.; Mikulis, D.J. Somatosensory cortical atrophy after spinal cord injury: A voxel-based morphometry study. Neurology 2006, 66, 762–764. [Google Scholar] [CrossRef]
- Wrigley, P.; Gustin, S.; Macey, P.; Nash, P.; Gandevia, S.; Macefield, V.; Siddall, P.; Henderson, L. Anatomical changes in human motor cortex and motor pathways following complete thoracic spinal cord injury. Cereb. Cortex 2009, 19, 224–232. [Google Scholar] [CrossRef]
- Freund, P.; Weiskopf, N.; Ashburner, J.; Wolf, K.; Sutter, R.; Altmann, D.R.; Friston, K.; Thompson, A.; Curt, A. MRI investigation of the sensorimotor cortex and the corticospinal tract after acute spinal cord injury: A prospective longitudinal study. Lancet Neurol. 2013, 12, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.S.; Zhao, C.; Chen, H.W.; Feng, T.; Guo, X.J.; Xu, M.; Rao, J.S. NT3 treatment alters spinal cord injury-induced changes in the gray matter volume of rhesus monkey cortex. Sci. Rep. 2022, 12, 5919. [Google Scholar] [CrossRef] [PubMed]
- Sydnor, V.J.; Larsen, B.; Bassett, D.S.; Alexander-Bloch, A.; Fair, D.A.; Liston, C.; Mackey, A.P.; Milham, M.P.; Pines, A.; Roalf, D.R.; et al. Neurodevelopment of the association cortices: Patterns, mechanisms, and implications for psychopathology. Neuron 2021, 109, 2820–2846. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.D.; Stojic, R.S.; Killackey, H.P.; Rhoades, R.W. Source of inappropriate receptive fields in cortical somatotopic maps from rats that sustained neonatal forelimb removal. J. Neurophysiol. 1999, 81, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, P.; Dai, Y.; Heckman, C.J. Effects of exercise training on alpha-motoneurons. J. Appl. Physiol. 2006, 101, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Giszter, S.; Davies, M.R.; Ramakrishnan, A.; Udoekwere, U.I.; Kargo, W.J. Trunk sensorimotor cortex is essential for autonomous weight-supported locomotion in adult rats spinalized as P1/P2 neonates. J. Neurophysiol. 2008, 100, 839–851. [Google Scholar] [CrossRef]
- Kao, T.; Shumsky, J.S.; Knudsen, E.B.; Murray, M.; Moxon, K.A. Functional role of exercise-induced cortical organization of sensorimotor cortex after spinal transection. J. Neurophysiol. 2011, 106, 2662–2674. [Google Scholar] [CrossRef]
- Gomez-Pinilla, F.; Ying, Z.; Zhuang, Y. Brain and spinal cord interaction: Protective effects of exercise prior to spinal cord injury. PLoS ONE 2012, 7, e32298. [Google Scholar] [CrossRef]
- Matsubayashi, K.; Nagoshi, N.; Komaki, Y.; Kojima, K.; Shinozaki, M.; Tsuji, O.; Iwanami, A.; Ishihara, R.; Takata, N.; Matsumoto, M.; et al. Assessing cortical plasticity after spinal cord injury by using resting-state functional magnetic resonance imaging in awake adult mice. Sci. Rep. 2018, 8, 14406. [Google Scholar] [CrossRef]
- Kaushal, M.; Oni-Orisan, A.; Chen, G.; Li, W.; Leschke, J.; Ward, B.D.; Kalinosky, B.; Budde, M.D.; Schmit, B.D.; Li, S.J.; et al. Evaluation of Whole-Brain Resting-State Functional Connectivity in Spinal Cord Injury: A Large-Scale Network Analysis Using Network-Based Statistic. J. Neurotrauma 2017, 34, 1278–1282. [Google Scholar] [CrossRef]
- Hou, J.M.; Sun, T.S.; Xiang, Z.M.; Zhang, J.Z.; Zhang, Z.C.; Zhao, M.; Zhong, J.F.; Liu, J.; Zhang, H.; Liu, H.L.; et al. Alterations of resting-state regional and network-level neural function after acute spinal cord injury. Neuroscience 2014, 277, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Moxon, K.A.; Oliviero, A.; Aguilar, J.; Foffani, G. Cortical reorganization after spinal cord injury: Always for good? Neuroscience 2014, 283, 78–94. [Google Scholar] [CrossRef]
- Sato, G.; Osumi, M.; Morioka, S. Effects of wheelchair propulsion on neuropathic pain and resting electroencephalography after spinal cord injury. J. Rehabil. Med. 2017, 49, 136–143. [Google Scholar] [CrossRef]
- Curt, A.; Alkadhi, H.; Crelier, G.R.; Boendermaker, S.H.; Hepp-Reymond, M.C.; Kollias, S.S. Changes of non-affected upper limb cortical representation in paraplegic patients as assessed by fMRI. Brain J. Neurol. 2002, 125, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Bruehlmeier, M.; Dietz, V.; Leenders, K.L.; Roelcke, U.; Missimer, J.; Curt, A. How does the human brain deal with a spinal cord injury? Eur. J. Neurosci. 1998, 10, 3918–3922. [Google Scholar] [CrossRef]
- Kerr, A.L.; Cheng, S.Y.; Jones, T.A. Experience-dependent neural plasticity in the adult damaged brain. J. Commun. Disord. 2011, 44, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Zhao, C.; Rao, J.S.; Guo, X.J.; Bao, S.S.; He, L.W.; Zhao, W.; Liu, Z.; Yang, Z.Y.; Li, X.G. Different macaque brain network remodeling after spinal cord injury and NT3 treatment. iScience 2023, 26, 106784. [Google Scholar] [CrossRef]
- Sawada, M.; Kato, K.; Kunieda, T.; Mikuni, N.; Miyamoto, S.; Onoe, H.; Isa, T.; Nishimura, Y. Function of the nucleus accumbens in motor control during recovery after spinal cord injury. Science 2015, 350, 98–101. [Google Scholar] [CrossRef]
- Chisholm, A.E.; Peters, S.; Borich, M.R.; Boyd, L.A.; Lam, T. Short-term cortical plasticity associated with feedback-error learning after locomotor training in a patient with incomplete spinal cord injury. Phys. Ther. 2015, 95, 257–266. [Google Scholar] [CrossRef]
- Peng, Y.; Liu, J.; Hua, M.; Liang, M.; Yu, C. Enhanced Effective Connectivity from Ipsilesional to Contralesional M1 in Well-Recovered Subcortical Stroke Patients. Front. Neurol. 2019, 10, 909. [Google Scholar] [CrossRef]
- Hubbard, I.J.; Carey, L.M.; Budd, T.W.; Levi, C.; McElduff, P.; Hudson, S.; Bateman, G.; Parsons, M.W. A Randomized Controlled Trial of the Effect of Early Upper-Limb Training on Stroke Recovery and Brain Activation. Neurorehabilit. Neural Repair 2015, 29, 703–713. [Google Scholar] [CrossRef]
- Hoffman, L.R.; Field-Fote, E.C. Functional and corticomotor changes in individuals with tetraplegia following unimanual or bimanual massed practice training with somatosensory stimulation: A pilot study. J. Neurol. Phys. Ther. JNPT 2010, 34, 193–201. [Google Scholar] [CrossRef]
- Lotze, M.; Ladda, A.M.; Stephan, K.M. Cerebral plasticity as the basis for upper limb recovery following brain damage. Neurosci. Biobehav. Rev. 2019, 99, 49–58. [Google Scholar] [CrossRef]
- Gustin, S.M.; Peck, C.C.; Cheney, L.B.; Macey, P.M.; Murray, G.M.; Henderson, L.A. Pain and plasticity: Is chronic pain always associated with somatosensory cortex activity and reorganization? J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 14874–14884. [Google Scholar] [CrossRef] [PubMed]
- Makin, T.R.; Scholz, J.; Filippini, N.; Henderson Slater, D.; Tracey, I.; Johansen-Berg, H. Phantom pain is associated with preserved structure and function in the former hand area. Nat. Commun. 2013, 4, 1570. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Voulalas, P.; Ji, Y.; Masri, R. Post-translational modification of cortical GluA receptors in rodents following spinal cord lesion. Neuroscience 2016, 316, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Beeler, J.A.; Petzinger, G.; Jakowec, M.W. The Enemy within: Propagation of Aberrant Corticostriatal Learning to Cortical Function in Parkinson’s Disease. Front. Neurol. 2013, 4, 134. [Google Scholar] [CrossRef] [PubMed]
- Nudo, R.J.; Milliken, G.W.; Jenkins, W.M.; Merzenich, M.M. Use-dependent alterations of movement representations in primary motor cortex of adult squirrel monkeys. J. Neurosci. Off. J. Soc. Neurosci. 1996, 16, 785–807. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zheng, Y.; Wei, J.A.; Ouyang, H.; Huang, X.; Zhang, F.; Lai, C.S.W.; Ren, C.; So, K.F.; Zhang, L. Exercise training improves motor skill learning via selective activation of mTOR. Sci. Adv. 2019, 5, eaaw1888. [Google Scholar] [CrossRef]
- Charalambous, C.C.; Helm, E.E.; Lau, K.A.; Morton, S.M.; Reisman, D.S. The feasibility of an acute high-intensity exercise bout to promote locomotor learning after stroke. Top. Stroke Rehabil. 2018, 25, 83–89. [Google Scholar] [CrossRef]
- Krakauer, J.W. Motor learning: Its relevance to stroke recovery and neurorehabilitation. Curr. Opin. Neurol. 2006, 19, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Olafson, E.R.; Jamison, K.W.; Sweeney, E.M.; Liu, H.; Wang, D.; Bruss, J.E.; Boes, A.D.; Kuceyeski, A. Functional connectome reorganization relates to post-stroke motor recovery and structural and functional disconnection. Neuroimage 2021, 245, 118642. [Google Scholar] [CrossRef]
- Belviranli, M.; Okudan, N. Differential effects of voluntary and forced exercise trainings on spatial learning ability and hippocampal biomarkers in aged female rats. Neurosci. Lett. 2022, 773, 136499. [Google Scholar] [CrossRef]
- Gumus, H.; Ilgin, R.; Koc, B.; Yuksel, O.; Kizildag, S.; Guvendi, G.; Karakilic, A.; Kandis, S.; Hosgorler, F.; Ates, M.; et al. A combination of ketogenic diet and voluntary exercise ameliorates anxiety and depression-like behaviors in Balb/c mice. Neurosci. Lett. 2022, 770, 136443. [Google Scholar] [CrossRef]
- Hayashi, N.; Himi, N.; Nakamura-Maruyama, E.; Okabe, N.; Sakamoto, I.; Hasegawa, T.; Miyamoto, O. Improvement of motor function induced by skeletal muscle contraction in spinal cord-injured rats. Spine J. 2019, 19, 1094–1105. [Google Scholar] [CrossRef]
- Sánchez-Ventura, J.; Giménez-Llort, L.; Penas, C.; Udina, E. Voluntary wheel running preserves lumbar perineuronal nets, enhances motor functions and prevents hyperreflexia after spinal cord injury. Exp. Neurol. 2021, 336, 113533. [Google Scholar] [CrossRef]
- Evans, N.H.; Suri, C.; Field-Fote, E.C. Walking and Balance Outcomes Are Improved Following Brief Intensive Locomotor Skill Training but Are Not Augmented by Transcranial Direct Current Stimulation in Persons with Chronic Spinal Cord Injury. Front. Hum. Neurosci. 2022, 16, 849297. [Google Scholar] [CrossRef]
- Ozturk, E.D.; Lapointe, M.S.; Kim, D.I.; Hamner, J.W.; Tan, C.O. Effect of 6-Month Exercise Training on Neurovascular Function in Spinal Cord Injury. Med. Sci. Sports Exerc. 2021, 53, 38–46. [Google Scholar] [CrossRef]
- Quaney, B.M.; Boyd, L.A.; McDowd, J.M.; Zahner, L.H.; He, J.; Mayo, M.S.; Macko, R.F. Aerobic exercise improves cognition and motor function poststroke. Neurorehabilit. Neural Repair. 2009, 23, 879–885. [Google Scholar] [CrossRef]
- Rademeyer, H.J.; Gauthier, C.; Zariffa, J.; Walden, K.; Jeji, T.; McCullum, S.; Musselman, K.E. Using activity-based therapy for individuals with spinal cord injury or disease: Interviews with physical and occupational therapists in rehabilitation hospitals. J. Spinal Cord Med. 2023, 46, 298–308. [Google Scholar] [CrossRef]
- Angeli, C.A.; Boakye, M.; Morton, R.A.; Vogt, J.; Benton, K.; Chen, Y.S.; Ferreira, C.K.; Harkema, S.J. Recovery of Over-Ground Walking after Chronic Motor Complete Spinal Cord Injury. N. Engl. J. Med. 2018, 379, 1244–1250. [Google Scholar] [CrossRef]
- Chapman, L.; Cooper-Knock, J.; Shaw, P.J. Physical activity as an exogenous risk factor for amyotrophic lateral sclerosis: A review of the evidence. Brain J. Neurol. 2023, 146, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Julian, T.H.; Glascow, N.; Barry, A.D.F.; Moll, T.; Harvey, C.; Klimentidis, Y.C.; Newell, M.; Zhang, S.; Snyder, M.P.; Cooper-Knock, J.; et al. Physical exercise is a risk factor for amyotrophic lateral sclerosis: Convergent evidence from Mendelian randomisation, transcriptomics and risk genotypes. EBioMedicine 2021, 68, 103397. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.K.; Labban, J.D.; Gapin, J.I.; Etnier, J.L. The effects of acute exercise on cognitive performance: A meta-analysis. Brain Res. 2012, 1453, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Lambourne, K.; Tomporowski, P. The effect of exercise-induced arousal on cognitive task performance: A meta-regression analysis. Brain Res. 2010, 1341, 12–24. [Google Scholar] [CrossRef]
- McMorris, T. The acute exercise-cognition interaction: From the catecholamines hypothesis to an interoception model. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2021, 170, 75–88. [Google Scholar] [CrossRef]
- Marquez-Chin, C.; Popovic, M.R. Functional electrical stimulation therapy for restoration of motor function after spinal cord injury and stroke: A review. Biomed. Eng. Online 2020, 19, 34. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.H.; Gattas, S.; Brown, N.J.; Hong, J.D.; Limbo, J.N.; Chan, A.Y.; Oh, M.Y. Epidural electrical stimulation for spinal cord injury. Neural Regen. Res. 2021, 16, 2367–2375. [Google Scholar] [CrossRef]
- Eisdorfer, J.T.; Smit, R.D.; Keefe, K.M.; Lemay, M.A.; Smith, G.M.; Spence, A.J. Epidural Electrical Stimulation: A Review of Plasticity Mechanisms That Are Hypothesized to Underlie Enhanced Recovery from Spinal Cord Injury with Stimulation. Front. Mol. Neurosci. 2020, 13, 163. [Google Scholar] [CrossRef]
- Carhart, M.R.; He, J.P.; Herman, R.; D’Luzansky, S.; Willis, W.T. Epidural spinal-cord stimulation facilitates recovery of functional walking following incomplete spinal-cord injury. IEEE Trans. Neural Syst. Rehabilitation Eng. 2004, 12, 32–42. [Google Scholar] [CrossRef]
- Capogrosso, M.; Wenger, N.; Raspopovic, S.; Musienko, P.; Beauparlant, J.; Luciani, L.B.; Courtine, G.; Micera, S. A computational model for epidural electrical stimulation of spinal sensorimotor circuits. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 19326–19340. [Google Scholar] [CrossRef]
- Nishimura, Y.; Perlmutter, S.I.; Fetz, E.E. Restoration of upper limb movement via artificial corticospinal and musculospinal connections in a monkey with spinal cord injury. Front. Neural Circuits 2013, 7, 57. [Google Scholar] [CrossRef]
- Zimmermann, J.B.; Jackson, A. Closed-loop control of spinal cord stimulation to restore hand function after paralysis. Front. Neurosci. 2014, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Holinski, B.J.; A Mazurek, K.; Everaert, D.G.; Toossi, A.; Lucas-Osma, A.M.; Troyk, P.; Etienne-Cummings, R.; Stein, R.B.; Mushahwar, V.K. Intraspinal microstimulation produces over-ground walking in anesthetized cats. J. Neural Eng. 2016, 13, 056016. [Google Scholar] [CrossRef] [PubMed]
- Wenger, N.; Moraud, E.M.; Raspopovic, S.; Bonizzato, M.; DiGiovanna, J.; Musienko, P.; Morari, M.; Micera, S.; Courtine, G. Closed-loop neuromodulation of spinal sensorimotor circuits controls refined locomotion after complete spinal cord injury. Sci. Transl. Med. 2014, 6, 255ra133. [Google Scholar] [CrossRef] [PubMed]
- Formento, E.; Minassian, K.; Wagner, F.; Mignardot, J.B.; Le Goff-Mignardot, C.G.; Rowald, A.; Bloch, J.; Micera, S.; Capogrosso, M.; Courtine, G. Electrical spinal cord stimulation must preserve proprioception to enable locomotion in humans with spinal cord injury. Nat. Neurosci. 2018, 21, 1728–1741. [Google Scholar] [CrossRef] [PubMed]
- Wagner, F.B.; Mignardot, J.B.; Le Goff-Mignardot, C.G.; Demesmaeker, R.; Komi, S.; Capogrosso, M.; Rowald, A.; Seáñez, I.; Caban, M.; Pirondini, E.; et al. Targeted neurotechnology restores walking in humans with spinal cord injury. Nature 2018, 563, 65–71. [Google Scholar] [CrossRef]
- Darrow, D.; Balser, D.; Netoff, T.I.; Krassioukov, A.; Phillips, A.; Parr, A.; Samadani, U. Epidural Spinal Cord Stimulation Facilitates Immediate Restoration of Dormant Motor and Autonomic Supraspinal Pathways after Chronic Neurologically Complete Spinal Cord Injury. J. Neurotrauma 2019, 36, 2325–2336. [Google Scholar] [CrossRef]
- Pena Pino, I.; Hoover, C.; Venkatesh, S.; Ahmadi, A.; Sturtevant, D.; Patrick, N.; Freeman, D.; Parr, A.; Samadani, U.; Balser, D.; et al. Long-Term Spinal Cord Stimulation After Chronic Complete Spinal Cord Injury Enables Volitional Movement in the Absence of Stimulation. Front. Syst. Neurosci. 2020, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Siu, R.; Brown, E.H.; Mesbah, S.; Gonnelli, F.; Pisolkar, T.; Edgerton, V.R.; Ovechkin, A.V.; Gerasimenko, Y.P. Novel Noninvasive Spinal Neuromodulation Strategy Facilitates Recovery of Stepping after Motor Complete Paraplegia. J. Clin. Med. 2022, 11, 3670. [Google Scholar] [CrossRef]
- Asboth, L.; Friedli, L.; Beauparlant, J.; Martinez-Gonzalez, C.; Anil, S.; Rey, E.; Baud, L.; Pidpruzhnykova, G.; Anderson, M.A.; Shkorbatova, P.; et al. Cortico-reticulo-spinal circuit reorganization enables functional recovery after severe spinal cord contusion. Nat. Neurosci. 2018, 21, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Shahabi, P.; Karimi, P.; Soltani-Zangbar, H.; Morshedi, M.; Bani, S.; Jafarzadehgharehziaaddin, M.; Sadeghzadeh-Oskouei, B.; Ahmadalipour, A. Impacts of epidural electrical stimulation on Wnt signaling, FAAH, and BDNF following thoracic spinal cord injury in rat. J. Cell. Physiol. 2020, 235, 9795–9805. [Google Scholar] [CrossRef] [PubMed]
- Sivanesan, E.; Stephens, K.E.; Huang, Q.; Chen, Z.; Ford, N.C.; Duan, W.; He, S.Q.; Gao, X.; Linderoth, B.; Raja, S.N.; et al. Spinal cord stimulation prevents paclitaxel-induced mechanical and cold hypersensitivity and modulates spinal gene expression in rats. Pain Rep. 2019, 4, e785. [Google Scholar] [CrossRef] [PubMed]
- Stampacchia, G.; Gazzotti, V.; Olivieri, M.; Andrenelli, E.; Bonaiuti, D.; Calabro, R.S.; Carmignano, S.M.; Cassio, A.; Fundaro, C.; Companini, I.; et al. Gait robot-assisted rehabilitation in persons with spinal cord injury: A scoping review. NeuroRehabilitation 2022, 51, 609–647. [Google Scholar] [CrossRef] [PubMed]
- Morone, G.; Cocchi, I.; Paolucci, S.; Iosa, M. Robot-assisted therapy for arm recovery for stroke patients: State of the art and clinical implication. Expert. Rev. Med. Devices 2020, 17, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Banala, S.K.; Kim, S.H.; Agrawal, S.K.; Scholz, J.P. Robot assisted gait training with active leg exoskeleton (ALEX). IEEE Trans. Neural Syst. Rehabil. Eng. 2009, 17, 2–8. [Google Scholar] [CrossRef]
- Meng, Q.; Zeng, Q.; Xie, Q.; Fei, C.; Kong, B.; Lu, X.; Wang, H.; Yu, H. Flexible lower limb exoskeleton systems: A review. NeuroRehabilitation 2022, 50, 367–390. [Google Scholar] [CrossRef]
- Fang, C.Y.; Tsai, J.L.; Li, G.S.; Lien, A.S.Y.; Chang, Y.J. Effects of Robot-Assisted Gait Training in Individuals with Spinal Cord Injury: A Meta-analysis. BioMed Res. Int. 2020, 2020, 2102785. [Google Scholar] [CrossRef]
- Benito-Penalva, J.; Edwards, D.J.; Opisso, E.; Cortes, M.; Lopez-Blazquez, R.; Murillo, N.; Costa, U.; Tormos, J.M.; Vidal-Samsó, J.; Valls-Solé, J.; et al. Gait training in human spinal cord injury using electromechanical systems: Effect of device type and patient characteristics. Arch. Phys. Med. Rehabil. 2012, 93, 404–412. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Mutsuzaki, H.; Koseki, K.; Endo, Y.; Hashizume, Y.; Nakazawa, R.; Aoyama, T.; Yozu, A.; Kohno, Y. Gait Training Using a Wearable Robotic Device for Non-Traumatic Spinal Cord Injury: A Case Report. Geriatr. Orthop. Surg. Rehabil. 2020, 11, 2151459320956960. [Google Scholar] [CrossRef]
- Bersch, I.; Alberty, M.; Fridén, J. Robot-assisted training with functional electrical stimulation enhances lower extremity function after spinal cord injury. Artif. Organs 2022, 46, 2009–2014. [Google Scholar] [CrossRef]
- Alwardat, M.; Etoom, M.; Al Dajah, S.; Schirinzi, T.; Di Lazzaro, G.; Salimei, P.S.; Mercuri, N.B.; Pisani, A. Effectiveness of robot-assisted gait training on motor impairments in people with Parkinson’s disease: A systematic review and meta-analysis. Int. J. Rehabil. Res. 2018, 41, 287–296. [Google Scholar] [CrossRef]
- Seo, J.S.; Yang, H.S.; Jung, S.; Kang, C.S.; Jang, S.; Kim, D.H. Effect of reducing assistance during robot-assisted gait training on step length asymmetry in patients with hemiplegic stroke: A randomized controlled pilot trial. Medicine 2018, 97, e11792. [Google Scholar] [CrossRef]
- Calabrò, R.S.; Cacciola, A.; Bertè, F.; Manuli, A.; Leo, A.; Bramanti, A.; Naro, A.; Milardi, D.; Bramanti, P. Robotic gait rehabilitation and substitution devices in neurological disorders: Where are we now? Neurol. Sci. 2016, 37, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.L.; Ramos-Murguialday, A.; Birbaumer, N.; Hoffmann, U.; Luft, A. Neurophysiology of robot-mediated training and therapy: A perspective for future use in clinical populations. Front. Neurol. 2013, 4, 184. [Google Scholar] [CrossRef] [PubMed]
- Oza, C.S.; Giszter, S.F. Trunk robot rehabilitation training with active stepping reorganizes and enriches trunk motor cortex representations in spinal transected rats. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 7174–7189. [Google Scholar] [CrossRef] [PubMed]
- Calabrò, R.S.; Naro, A.; Russo, M.; Bramanti, P.; Carioti, L.; Balletta, T.; Buda, A.; Manuli, A.; Filoni, S.; Bramanti, A. Shaping neuroplasticity by using powered exoskeletons in patients with stroke: A randomized clinical trial. J. Neuroeng. Rehabil. 2018, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.; Solis-Escalante, T.; Grieshofer, P.; Neuper, C.; Müller-Putz, G.; Scherer, R. Level of participation in robotic-assisted treadmill walking modulates midline sensorimotor EEG rhythms in able-bodied subjects. Neuroimage 2012, 63, 1203–1211. [Google Scholar] [CrossRef]
- Savaki, H.E.; Raos, V. Action perception and motor imagery: Mental practice of action. Prog. Neurobiol. 2019, 175, 107–125. [Google Scholar] [CrossRef]
- Tong, Y.; Pendy, J.T., Jr.; Li, W.A.; Du, H.; Zhang, T.; Geng, X.; Ding, Y. Motor Imagery-Based Rehabilitation: Potential Neural Correlates and Clinical Application for Functional Recovery of Motor Deficits after Stroke. Aging Dis. 2017, 8, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Gowda, A.S.; Memon, A.N.; Bidika, E.; Salib, M.; Rallabhandi, B.; Fayyaz, H. Investigating the Viability of Motor Imagery as a Physical Rehabilitation Treatment for Patients with Stroke-Induced Motor Cortical Damage. Cureus J. Med. Sci. 2021, 13, e14001. [Google Scholar] [CrossRef]
- Carvalho, R.; Azevedo, E.; Marques, P.; Dias, N.; Cerqueira, J.J. Physiotherapy based on problem-solving in upper limb function and neuroplasticity in chronic stroke patients: A case series. J. Eval. Clin. Pract. 2018, 24, 552–560. [Google Scholar] [CrossRef]
- Grangeon, M.; Charvier, K.; Guillot, A.; Rode, G.; Collet, C. Using sympathetic skin responses in individuals with spinal cord injury as a quantitative evaluation of motor imagery abilities. Phys. Ther. 2012, 92, 831–840. [Google Scholar] [CrossRef]
- Sabbah, P.; de Schonen, S.; Leveque, C.; Gay, S.; Pfefer, F.; Nioche, C.; Sarrazin, J.-L.; Barouti, H.; Tadie, M.; Cordoliani, Y.-S.; et al. Sensorimotor cortical activity in patients with complete spinal cord injury: A functional magnetic resonance imaging study. J. Neurotrauma 2002, 19, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wan, L.; Qin, W.; Zheng, W.; Qi, Z.; Chen, N.; Li, K. Functional Preservation and Reorganization of Brain during Motor Imagery in Patients with Incomplete Spinal Cord Injury: A Pilot fMRI Study. Front. Hum. Neurosci. 2016, 10, 46. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, F.; Guillot, A.; Mateo, S.; Daligault, S.; Delpuech, C.; Rode, G.; Collet, C. Neuroplasticity of prehensile neural networks after quadriplegia. Neuroscience 2014, 274, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zheng, W.; Liang, T.; Yang, Y.; Yang, B.; Chen, X.; Chen, Q.; Li, X.; Lu, J.; Li, B.; et al. Brain Activation Evoked by Motor Imagery in Pediatric Patients with Complete Spinal Cord Injury. AJNR Am. J. Neuroradiol. 2023, 44, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Mokienko, O.A.; Chernikova, L.A.; Frolov, A.A.; Bobrov, P.D. Motor Imagery and its Practical Application. Zhurnal Vysshei Nervnoi Deyatelnosti Imeni IP Pavlova 2013, 63, 195–204. [Google Scholar] [CrossRef]
- Sims, C.; Waldron, R.; Marcellin-Little, D.J. Rehabilitation and Physical Therapy for the Neurologic Veterinary Patient. Vet. Clin. N. Am. Small Anim. Pract. 2015, 45, 123–143. [Google Scholar] [CrossRef]


{kind=link}
| Types | Characteristics | Advantage | Shortcoming | Effect | Applicability |
|---|---|---|---|---|---|
| Rehabilitation exercise | Easy to operate, convenient, and low-cost; the most commonly used clinical method for the rehabilitation of spinal cord injury (SCI). | Practicability, operability, and low cost; the most effective rehabilitation means to promote brain reorganization after SCI at present. | Exercise acts on the distal limbs, indirectly stimulates the CNS of the brain; rehabilitation training takes a long time. | One of the most important quantifiable methods of functional recovery after SCI, which activates the cerebral cortex by acting on the distal limb. | It can be applied to people with various degrees of SCI and at various stages of treatment after SCI, but it is mainly used for incomplete SCI. |
| Epidural electrical stimulation (EES) | It can induce movement through a sequence of pulsed currents and makes effective use of the residual function of spinal nerves and muscles. | It can enhance the neural plasticity of the motor cortex and activate the excitability of the corticospinal tract. | The use of implantable electrodes may cause postoperative infection. Prolonged application of electrical stimulation can cause pain in patients. | Combined with rehabilitation exercise, patients can improve their exercise ability in a short period of time. | EES is suitable for severe and complete SCI. It can be used for the early rehabilitation of SCI and patients with severe motor dysfunction. |
| Exoskeleton rehabilitation robot | It can simulate the structure characteristics of the human body and has the characteristics of high strength, repeatability, and interaction. | It can provide repeatable and specific movement therapy and promote the rehabilitation of fine movements after SCI. It can objectively quantify rehabilitation parameters and training output. | The structure is complex and expensive, and the weight and body shape should be considered. | It can be used to provide standard movement training for injured limbs. It has a better effect on brain reorganization than routine exercise training. | For patients with severe and complete SCI, it can provide more effective support. It is used for rehabilitation training of patients with severe motor dysfunction after SCI. |
| Motor imagination (MI) | Makes full use of the patient’s subjective intentions and acts directly on the cerebral cortex. | It has a wide range of applications, motivates patients, and guides patients to achieve appropriate brain activation patterns related to tasks. | The standard of treatment and the best intensity of intervention are not clear. The effect depends on the feedback content and the status of the patient. | It can help patients with SCI activate the activity of the cerebral cortex and realize the self-regulation of the functional brain networks. Combining with other rehabilitation training methods is beneficial to the better effect of MI. | It can be applied to individuals with complete and incomplete SCI and at various stages of treatment after SCI. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.-W.; Guo, X.-J.; Zhao, C.; Rao, J.-S. Rehabilitation Training after Spinal Cord Injury Affects Brain Structure and Function: From Mechanisms to Methods. Biomedicines 2024, 12, 41. https://doi.org/10.3390/biomedicines12010041
He L-W, Guo X-J, Zhao C, Rao J-S. Rehabilitation Training after Spinal Cord Injury Affects Brain Structure and Function: From Mechanisms to Methods. Biomedicines. 2024; 12(1):41. https://doi.org/10.3390/biomedicines12010041
Chicago/Turabian StyleHe, Le-Wei, Xiao-Jun Guo, Can Zhao, and Jia-Sheng Rao. 2024. "Rehabilitation Training after Spinal Cord Injury Affects Brain Structure and Function: From Mechanisms to Methods" Biomedicines 12, no. 1: 41. https://doi.org/10.3390/biomedicines12010041