
BHLHE40 Maintains the Stemness of PαS Cells In Vitro by Targeting Zbp1 through the Wnt/β-Catenin Signaling Pathway
 , , , and
, , , and
Abstract
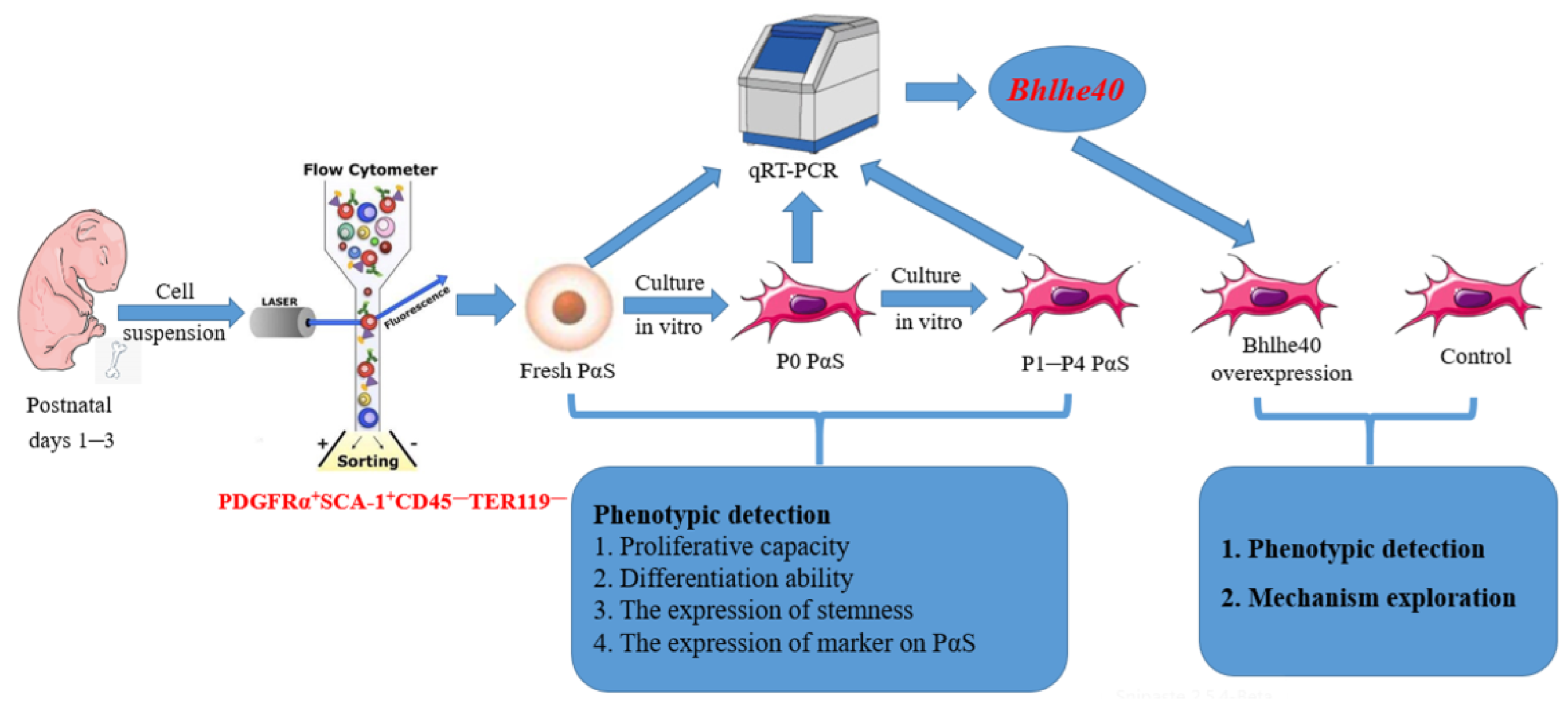
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Lentivirus Infection, RNA Transfection and Use of Inhibitor of Wnt/β-Catenin Signaling Pathway
2.3. Cell Proliferation Capacity Test
2.4. Alkaline Phosphatase (ALP) Staining and Activity
2.5. Alizarin Red S (ARS) Staining and Quantification
2.6. PαS Cells in Well Plates Stained with Alcian Blue
2.7. Chondrosphere Sections Stained with Alcian Blue and Sirius Red
2.8. RNA Extraction and Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sense Strand/Sense Primer (5′–3′) | Antisense Strand/Antisense Primer (5′–3′) |
|---|---|---|
| si-RNA | ||
| si-Zbp1 | GCCUGCAACAUGGAGCAUATT | UAUGCUCCAUGUUG-CAGGCTT |
| Si-NC | UUCUCCGAACGUGUCACGUTT | ACGUGACACGUUCGGAGAATT |
| Primers | ||
| mGapdh | TGGCCTTCCGTGTTCCTAC | GAGTTGCTGTTGAAGTCGCA |
| mAlp | CCAACTCTTTTGTGCCAGAGA | GGCTACATTGGTGTTGAGCTTTT |
| mRunx2 | AGAGTCAGATTACAGATCCCAGG | TGGCTCTTCTTACTGA-GAGAGG |
| mNanog | AGGACAGGTTTCAGAAGCAGA | CCATTGCTAG-TCTTCAACCACTG |
| mSox2 | GCGGAGTGGAAACTTTTGTCC | GGGAAGCGTGTACTTATCCTTCT |
| mOct4 | AGAGGATCACCTTGGGGTACA | CGAA-GCGACAGATGGTGGTC |
| mPdgfrα | TATCCTCCCAAACGAGAATGAGA | GTGGTTGTAGTAG-CAAGTGTACC |
| mSca-1 | CTGACTGGAAAGCCGAAACTC | CGACCCGTCCTTT-GAATTTCT |
| mBhlhe40 | CTGTCAGGGATGGATTTTGCC | GCTGTCTTCGCTCCGTTTTATTC |
| mBglap | CTGACCTCACAGATCCCAAGC | TGGTCTGATAGCTCGTCACAAG |
| mCol1a1 | TAAGGGTCCCCAATGGTGAGA | GGGTCCCTCGACTCC-TACAT |
| mAcan | CCTGCTACTTCATCGACCCC | AGATGCTGTTGACTCGAACCT |
| mSox9 | CGGAACAGACTCACATCTCTCC | GCTTGCACGTCGGTTTTGG |
| mComp | ACTGCCTGCGTTCTAGTGC | CGCCGCATTAGTCTCCTGAA |
| mZbp-1 | AAGAGTCCCCTGCGATTATTTG | TCTGGATGGCGTTT-GAATTGG |
| mWnt3a | CTCCTCTCGGATACCTCTTAGTG | GCATGATCTCCACGTAG-TTCCTG |
| mCtnnb1 | ATGGAGCCGGACAGAAAAGC | CTTGCCACTCAGGGAAGGA |
| mCcnd1 | GCGTACCCTGACACCAATCTC | CTCCTCTTCGCACTTCTGCTC |
| mSpp1 | AGCAAGAAACTCTTCCAAGCAA | GTGAGATTCGTCAGAT-TCATCCG |
2.9. Enzyme Linked Immunosorbent Assay (ELISA)
2.10. Chromatin Immunoprecipitation (ChIP) Assay
2.11. Ectopic Osteogenesis and Chondrogenesis In Vivo
2.12. Statistical Analyses
3. Results
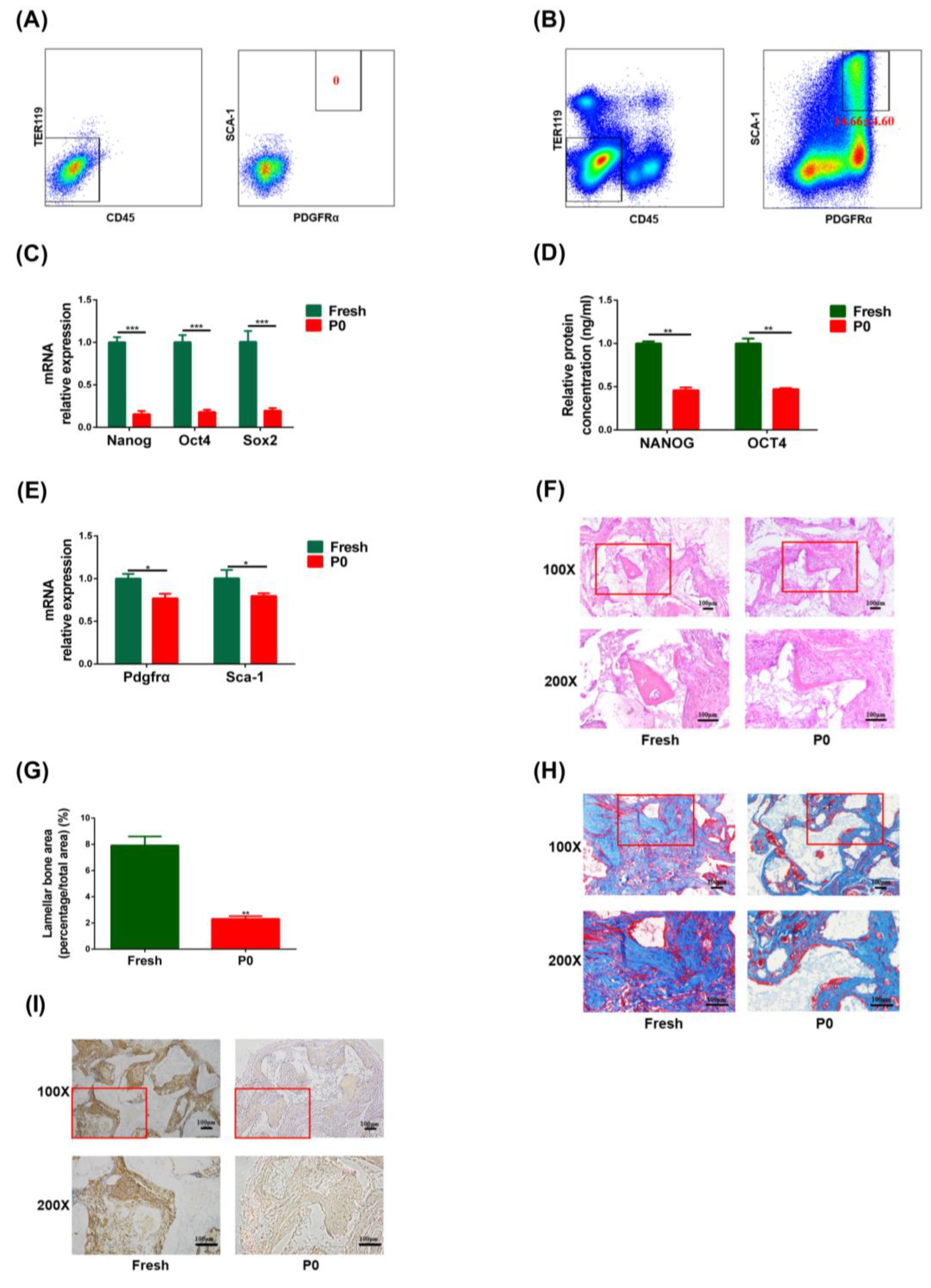
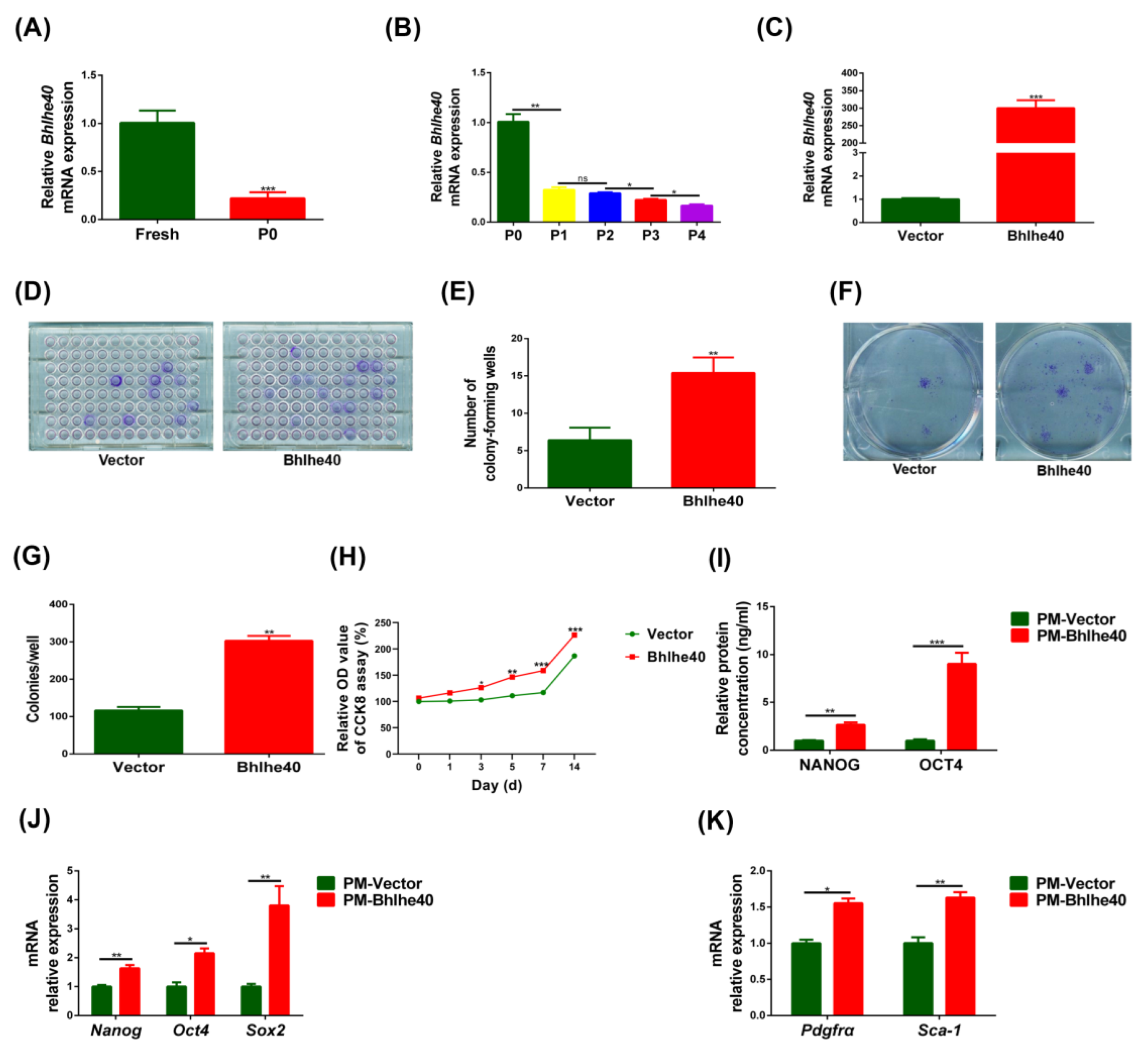
3.1. The Stemness of Fresh PαS Cells Was Superior to That of In Vitro P0 Cells
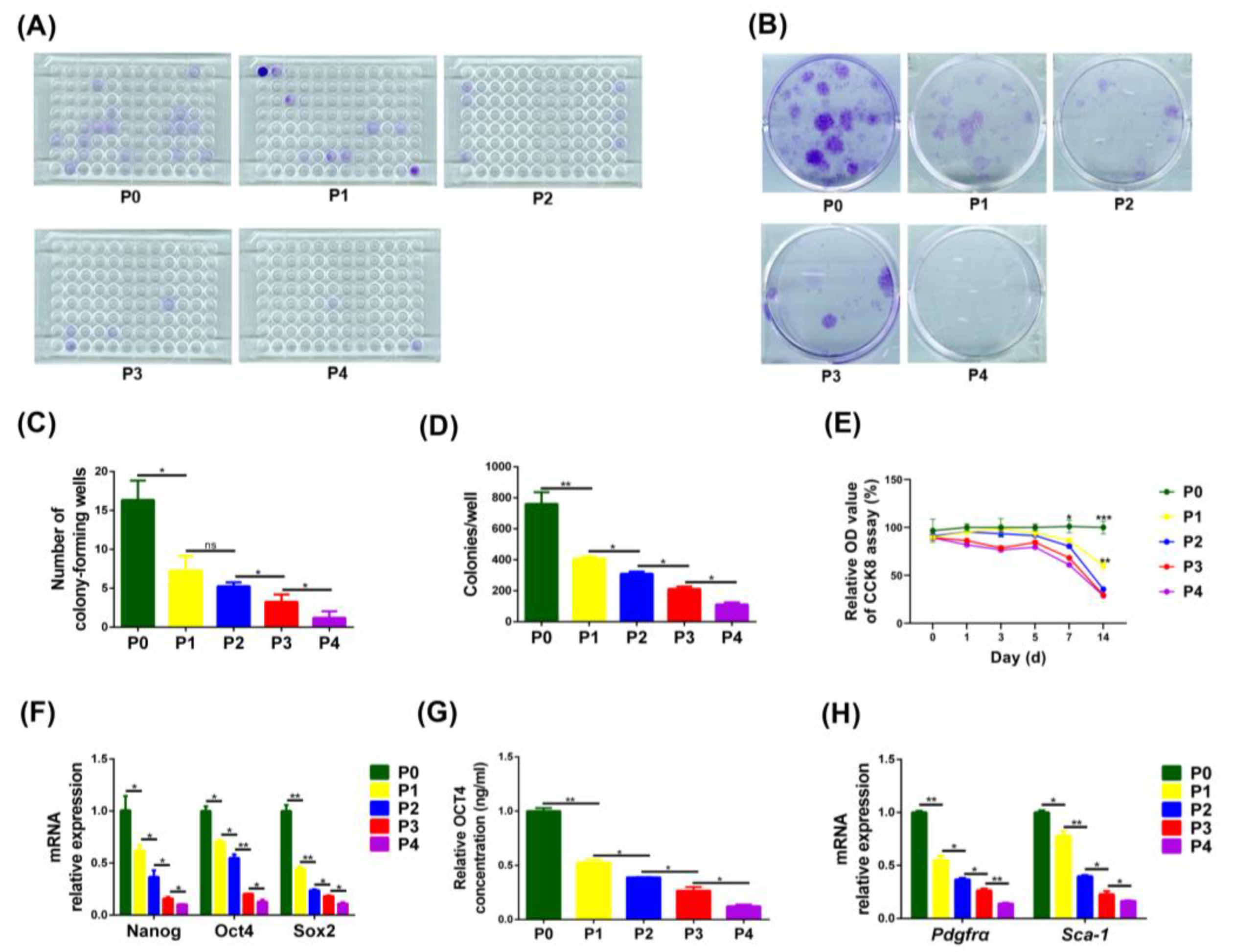
3.2. The Proliferative Capacity, Expression of Stemness Genes and Cell Surface Markers in PαS Cells Decreased Gradually from P0 to P4
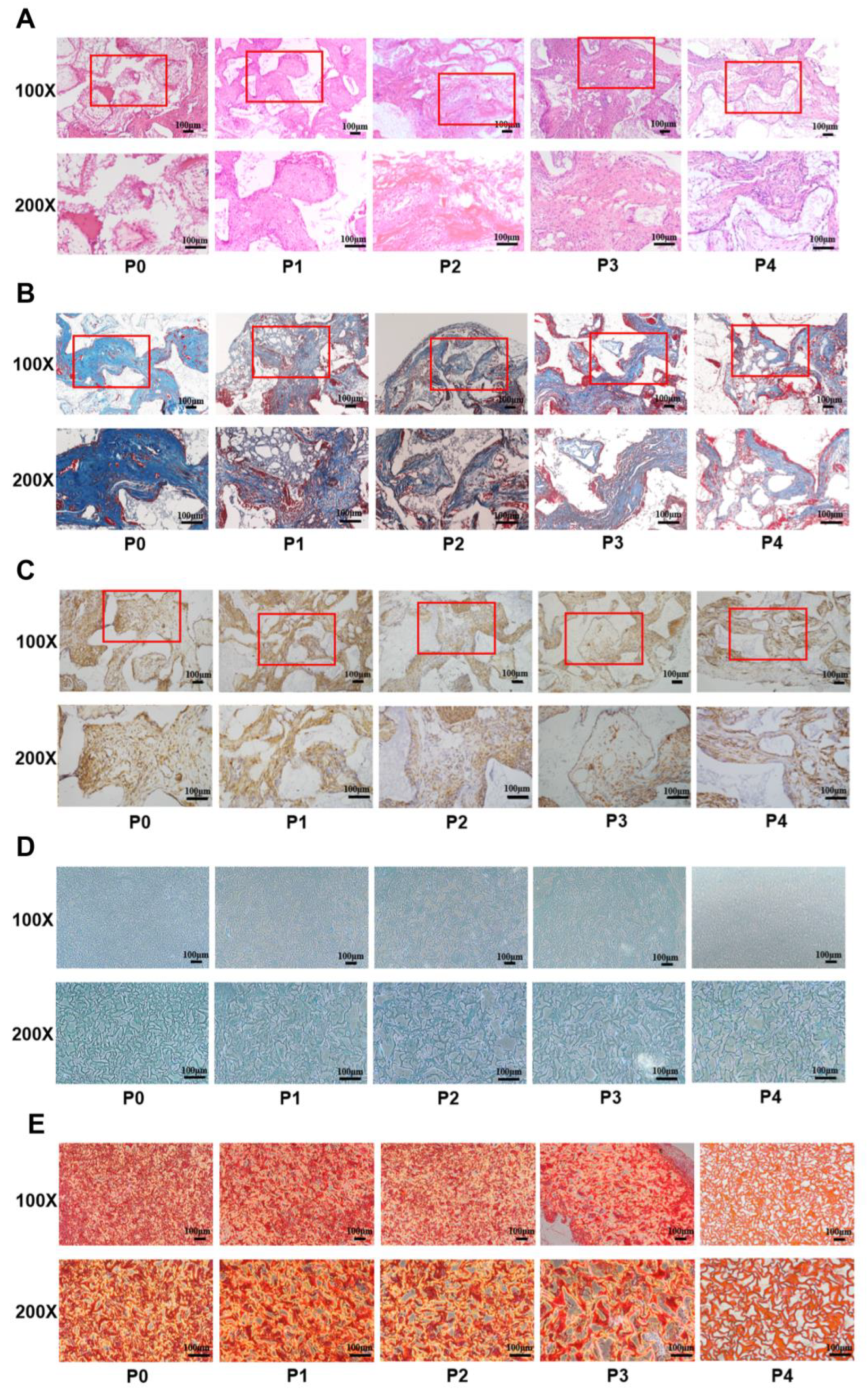
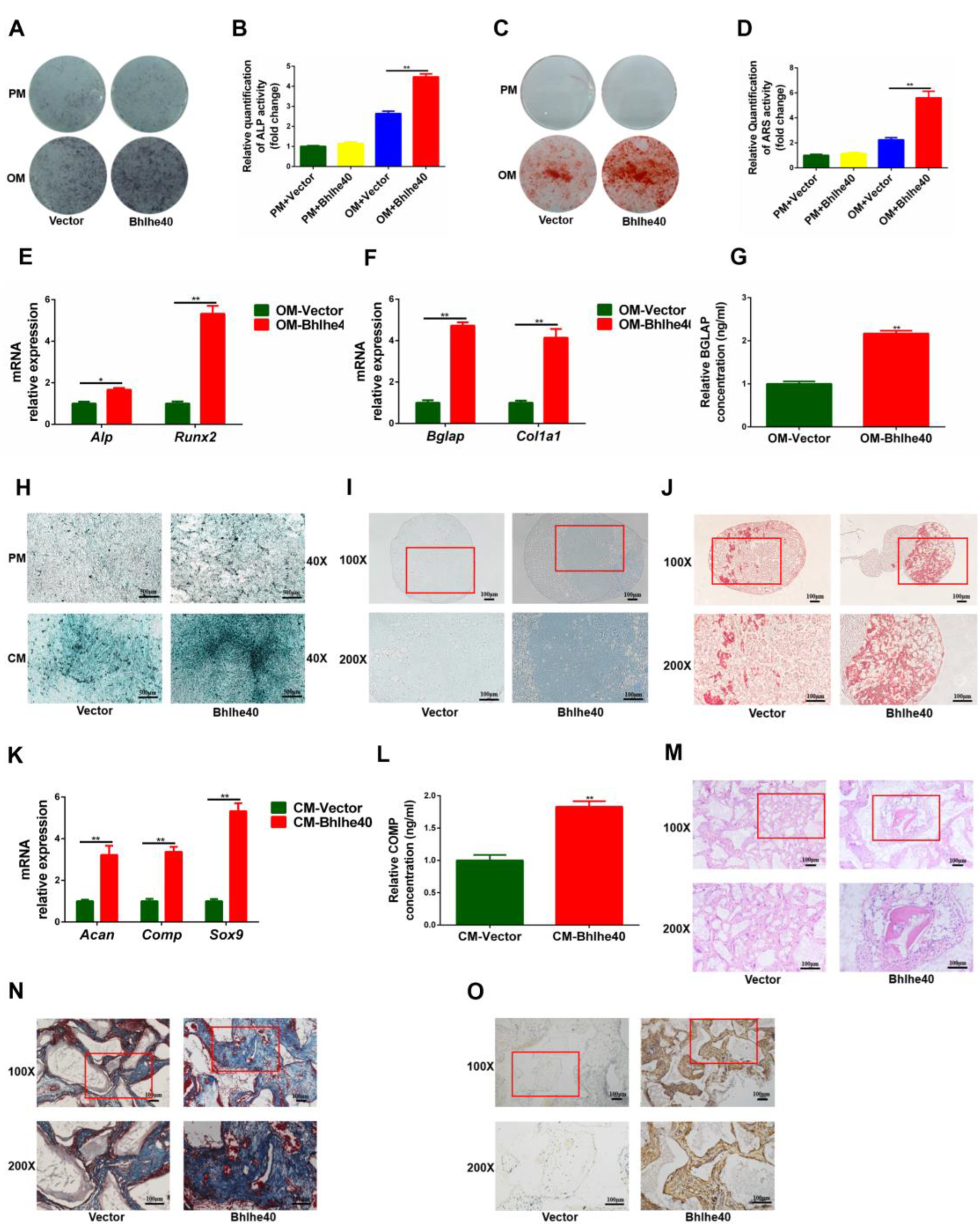
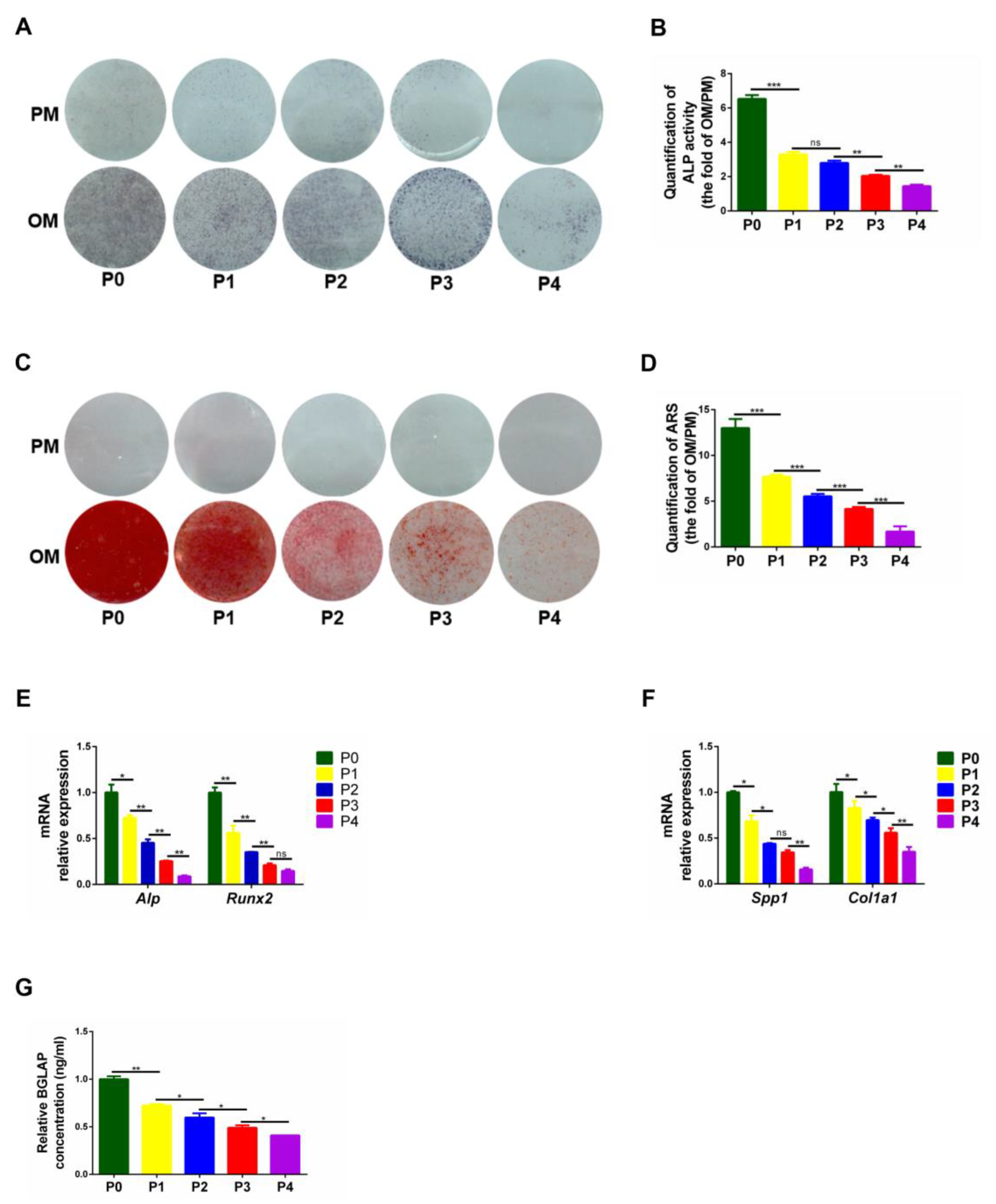
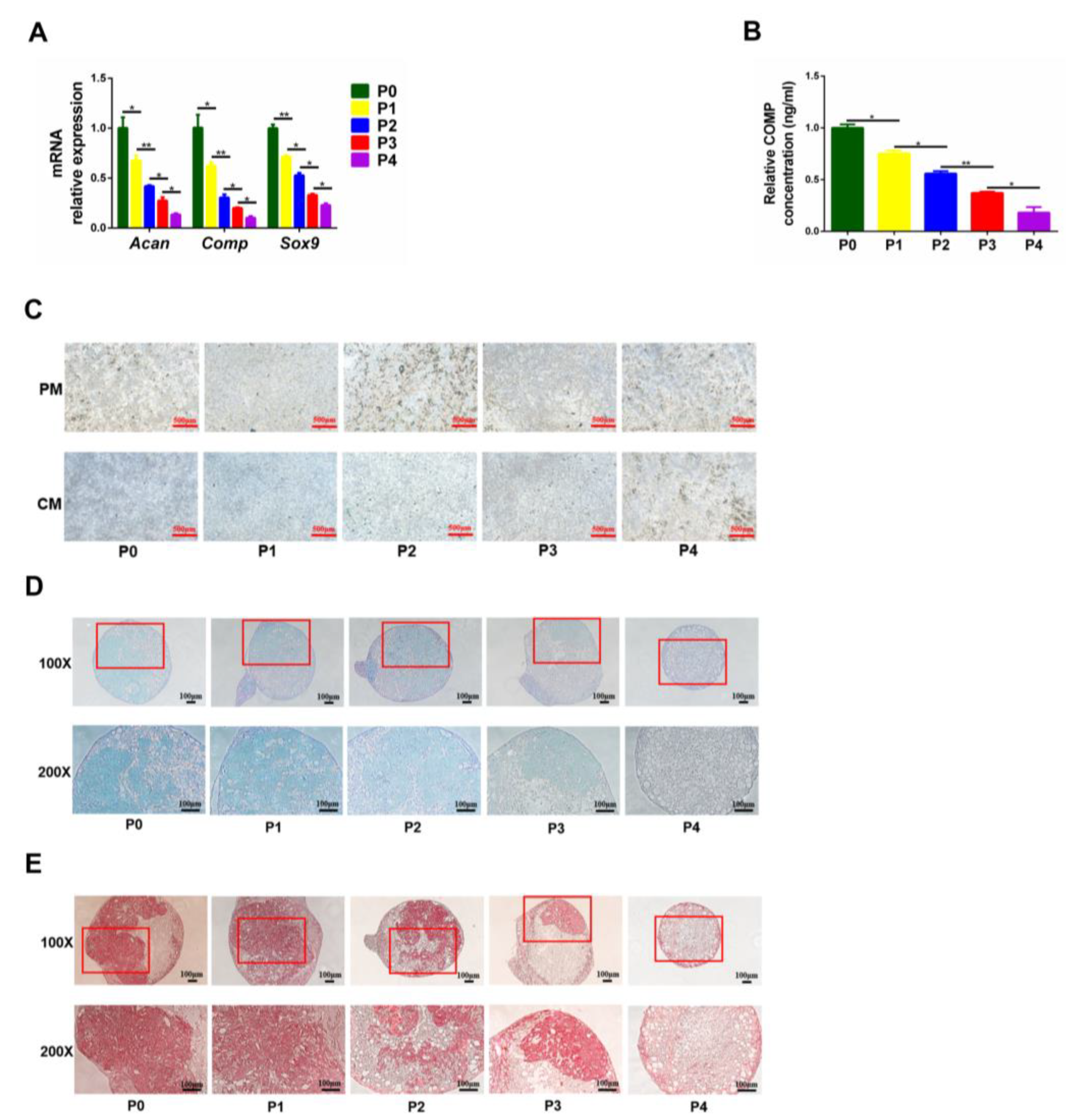
3.3. The Osteogenic and Chondrogenic Differentiation Capacity of PαS Cells Decreased Gradually from P0 to P4
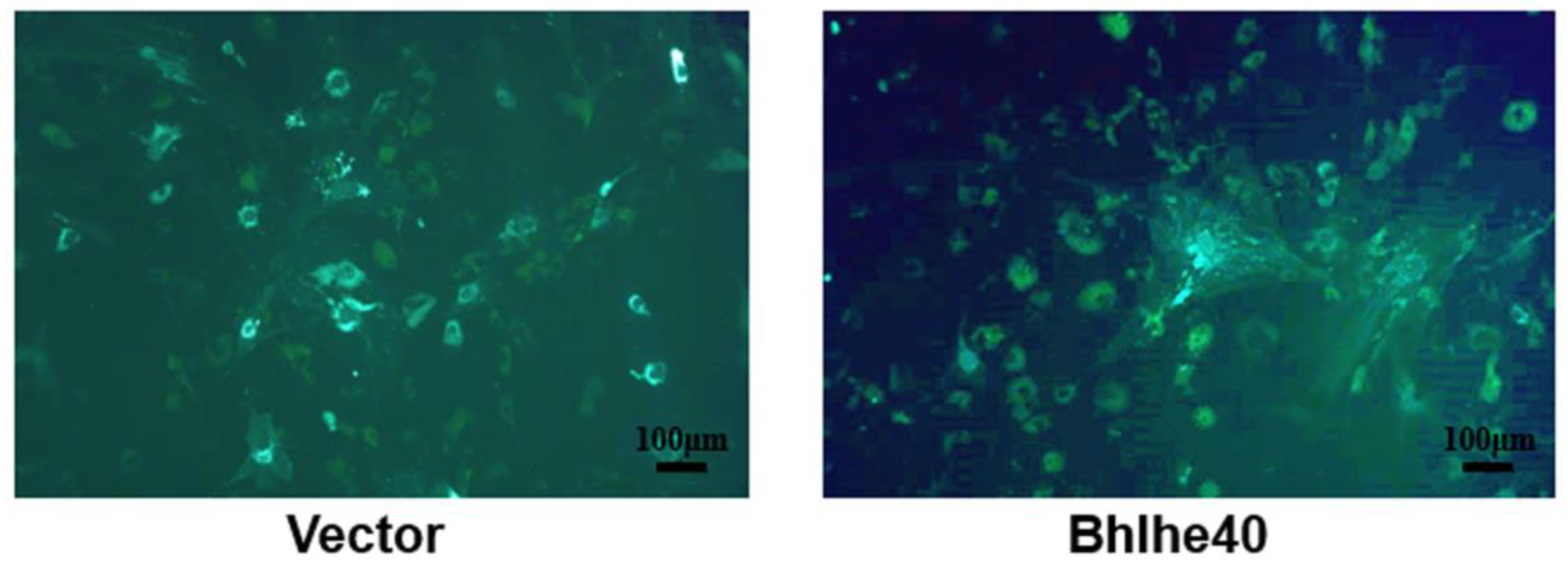
3.4. BHLHE40 Stimulated Proliferation, the Expression of Stemness Genes and Cell Surface Markers in PαS Cells
3.5. Overexpression of Bhlhe40 Stimulated Osteogenic and Chondrogenic Differentiation of PαS Cells
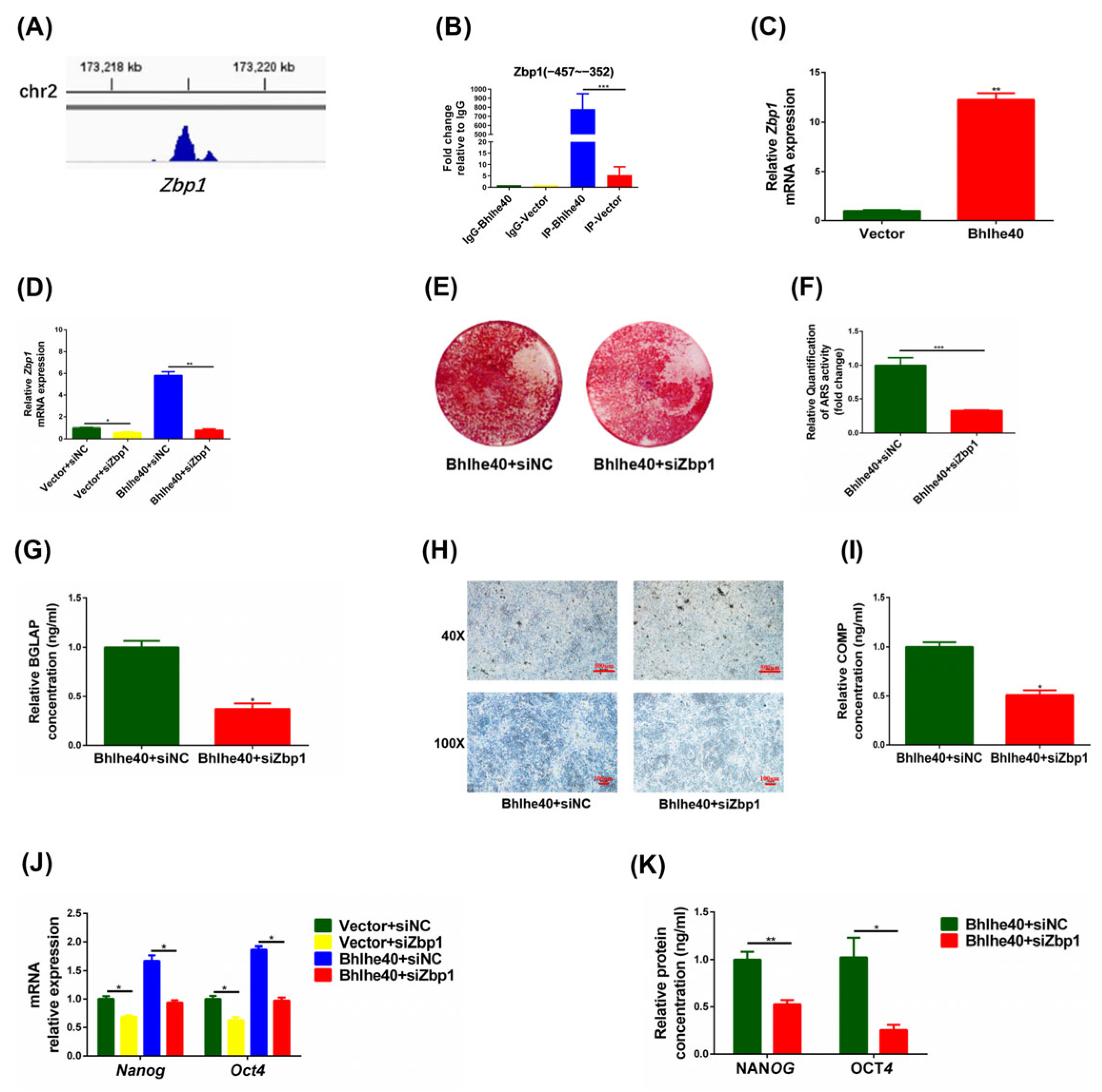
3.6. BHLHE40 Maintains the Stemness of PαS Cells by Activating Zbp1
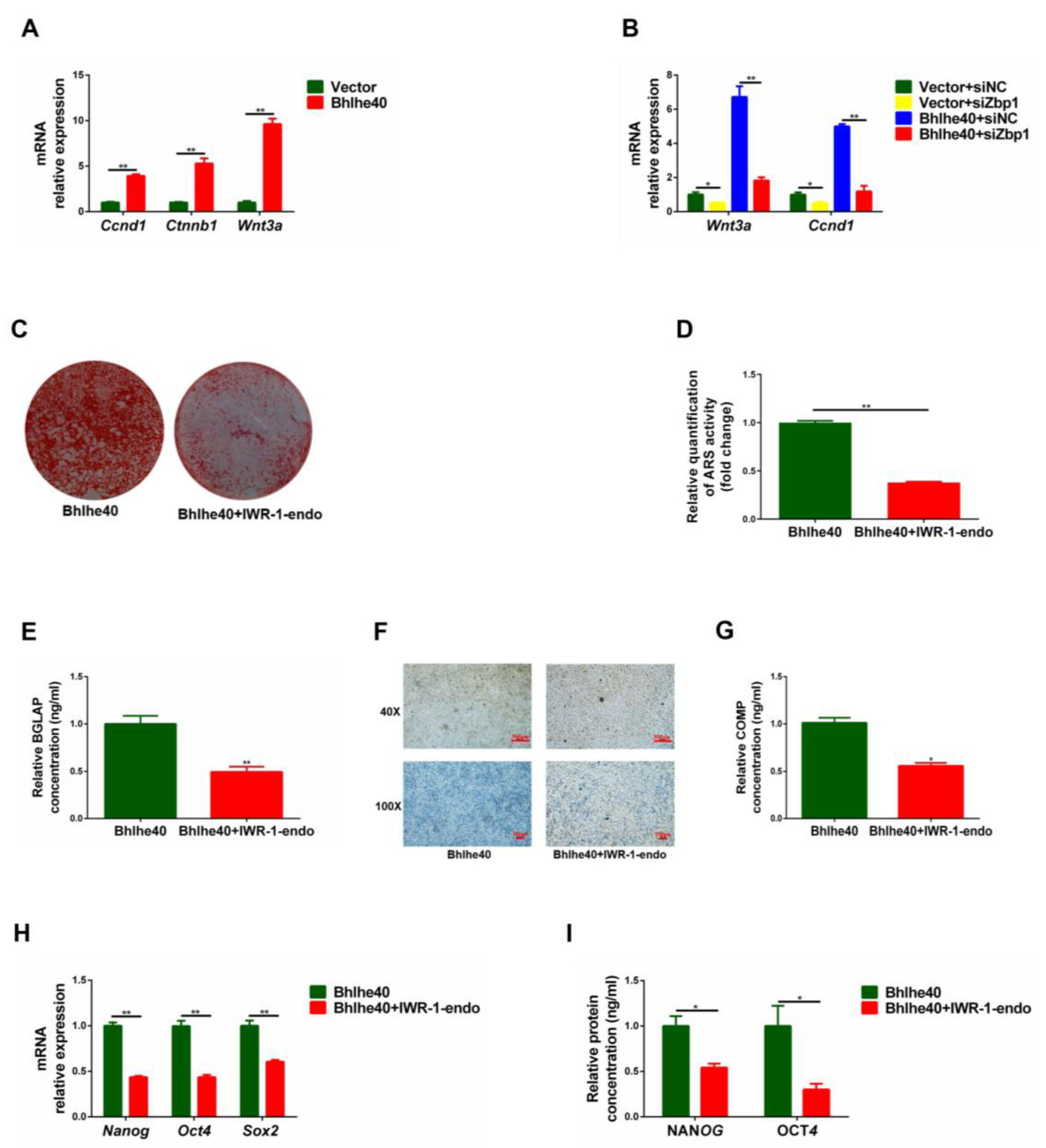
3.7. BHLHE40 Regulates the Stemness of PαS Cells through the Wnt/β-Catenin Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Bieback, K.; Hecker, A.; Kocaömer, A.; Lannert, H.; Schallmoser, K.; Strunk, D.; Klüter, H. Human Alternatives to Fetal Bovine Serum for the Expansion of Mesenchymal Stromal Cells from Bone Marrow. Stem Cells 2009, 27, 2331–2341. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.L.; Hankenson, K.D. Integration of BMP, Wnt, and notch signaling pathways in osteoblast differentiation. J. Cell. Biochem. 2011, 112, 3491–3501. [Google Scholar] [CrossRef] [Green Version]
- Leveque, X.; Hochane, M.; Geraldo, F.; Dumont, S.; Gratas, C.; Oliver, L.; Gaignier, C.; Trichet, V.; Layrolle, P.; Heymann, D.; et al. Low-Dose Pesticide Mixture Induces Accelerated Mesenchymal Stem Cell Aging In Vitro. Stem Cells 2019, 37, 1083–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasumarthy, K.K.; Jayavelu, N.D.; Kilpinen, L.; Andrus, C.; Battle, S.L.; Korhonen, M.; Lehenkari, P.; Lund, R.; Laitinen, S.; Hawkins, R.D. Methylome Analysis of Human Bone Marrow MSCs Reveals Extensive Age- and Culture-Induced Changes at Distal Regulatory Elements. Stem Cell Rep. 2017, 9, 999–1015. [Google Scholar] [CrossRef] [Green Version]
- Tokalov, S.V.; Grüner, S.; Schindler, S.; Wolf, G.; Baumann, M.; Abolmaali, N. Age-Related Changes in the Frequency of Mesenchymal Stem Cells in the Bone Marrow of Rats. Stem Cells Dev. 2007, 16, 439–446. [Google Scholar] [CrossRef]
- Bourillot, P.-Y.; Savatier, P. Krüppel-like transcription factors and control of pluripotency. BMC Biol. 2010, 8, 125–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rombouts, W.J.C.; E Ploemacher, R. Primary murine MSC show highly efficient homing to the bone marrow but lose homing ability following culture. Leukemia 2003, 17, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Abbuehl, J.-P.; Tatarova, Z.; Held, W.; Huelsken, J. Long-Term Engraftment of Primary Bone Marrow Stromal Cells Repairs Niche Damage and Improves Hematopoietic Stem Cell Transplantation. Cell Stem Cell 2017, 21, 241–255.e6. [Google Scholar] [CrossRef]
- Mo, C.; Guo, J.; Qin, J.; Zhang, X.; Sun, Y.; Wei, H.; Cao, D.; Zhang, Y.; Zhao, C.; Xiong, Y.; et al. Single-cell transcriptomics of LepR-positive skeletal cells reveals heterogeneous stress-dependent stem and progenitor pools. EMBO J. 2022, 41, e108415. [Google Scholar] [CrossRef]
- Peister, A.; Mellad, J.A.; Larson, B.L.; Hall, B.M.; Gibson, L.F.; Prockop, D.J. Adult stem cells from bone marrow (MSCs) isolated from different strains of inbred mice vary in surface epitopes, rates of proliferation, and differentiation potential. Blood 2004, 103, 1662–1668. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, S.; Mabuchi, Y.; Kubota, Y.; Nagai, Y.; Niibe, K.; Hiratsu, E.; Suzuki, S.; Miyauchi-Hara, C.; Nagoshi, N.; Sunabori, T.; et al. Prospective identification, isolation, and systemic transplantation of multipotent mesenchymal stem cells in murine bone marrow. J. Exp. Med. 2009, 206, 2483–2496. [Google Scholar] [CrossRef]
- Ogawa, Y.; Morikawa, S.; Okano, H.; Mabuchi, Y.; Suzuki, S.; Yaguchi, T.; Sato, Y.; Mukai, S.; Yaguchi, S.; Inaba, T.; et al. MHC-compatible bone marrow stromal/stem cells trigger fibrosis by activating host T cells in a scleroderma mouse model. Elife 2016, 5, e09394. [Google Scholar] [CrossRef] [PubMed]
- Ow, J.R.; Tan, Y.H.; Jin, Y.; Bahirvani, A.G.; Taneja, R. Stra13 and Sharp-1, the Non-Grouchy Regulators of Development and Disease. Curr. Top. Dev. Biol. 2014, 110, 317–338. [Google Scholar] [PubMed]
- Zhu, Y.; Xu, L.; Zhang, J.; Hu, X.; Liu, Y.; Yin, H.; Lv, T.; Zhang, H.; Liu, L.; An, H.; et al. Sunitinib induces cellular senescence via p53/Dec1 activation in renal cell carcinoma cells. Cancer Sci. 2013, 104, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, K.; Horikawa, Y. Regulation of lipogenesis via BHLHB2/DEC1 and ChREBP feedback looping. Biochem. Biophys. Res. Commun. 2008, 374, 95–100. [Google Scholar] [CrossRef]
- Gosselin, D.; Link, V.M.; Romanoski, C.E.; Fonseca, G.J.; Eichenfield, D.Z.; Spann, N.J.; Stender, J.D.; Chun, H.B.; Garner, H.; Geissmann, F.; et al. Environment Drives Selection and Function of Enhancers Controlling Tissue-Specific Macrophage Identities. Cell 2014, 159, 1327–1340. [Google Scholar] [CrossRef]
- Zafar, A.; Ng, H.P.; Kim, G.; Chan, E.R.; Mahabeleshwar, G.H. BHLHE40 promotes macrophage pro-inflammatory gene expression and functions. FASEB J. 2021, 35, e21940. [Google Scholar] [CrossRef]
- Rauschmeier, R.; Gustafsson, C.; Reinhardt, A.; A-Gonzalez, N.; Tortola, L.; Cansever, D.; Subramanian, S.; Taneja, R.; Rossner, M.J.; Sieweke, M.H.; et al. Bhlhe40 and Bhlhe41 transcription factors regulate alveolar macrophage self-renewal and identity. EMBO J. 2019, 38, e101233. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Zhao, Y.; Li, M.; Liu, Y.; Cheng, P.; Lv, Y.; Mao, F.; Chen, W.; Yang, S.; Hao, C.; et al. Upexpression of BHLHE40 in gastric epithelial cells increases CXCL12 production through interaction with p-STAT3 in Helicobacter pylori -associated gastritis. FASEB J. 2020, 34, 1169–1181. [Google Scholar] [CrossRef] [Green Version]
- Kiss, Z.; Mudryj, M.; Ghosh, P.M. Non-circadian aspects of BHLHE40 cellular function in cancer. Genes Cancer 2020, 11, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Jarjour, N.N.; Schwarzkopf, E.A.; Bradstreet, T.R.; Shchukina, I.; Lin, C.-C.; Huang, S.C.-C.; Lai, C.-W.; Cook, M.E.; Taneja, R.; Stappenbeck, T.S.; et al. Bhlhe40 mediates tissue-specific control of macrophage proliferation in homeostasis and type 2 immunity. Nat. Immunol. 2019, 20, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Yun, Z.; Maecker, H.L.; Johnson, R.S.; Giaccia, A.J. Inhibition of PPAR gamma 2 gene expression by the HIF-1-regulated gene DEC1/Stra13: A mechanism for regulation of adipogenesis by hypoxia. Dev. Cell 2002, 2, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, N.; Noshiro, M.; Kawamoto, T.; Nakashima, A.; Honda, K.; Fukuzaki-Dohi, U.; Honma, S.; Fujimoto, K.; Tanimoto, K.; Tanne, K.; et al. Regulation of basic helix-loop-helix transcription factors Dec1 and Dec2 by RORα and their roles in ad-ipogenesis. Genes Cells 2012, 17, 109–121. [Google Scholar] [CrossRef]
- Sethuraman, A.; Brown, M.; Krutilina, R.; Wu, Z.-H.; Seagroves, T.N.; Pfeffer, L.M.; Fan, M. BHLHE40 confers a pro-survival and pro-metastatic phenotype to breast cancer cells by modulating HBEGF secretion. Breast Cancer Res. 2018, 20, 117–133. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Henson, E.S.; Xiao, W.; Huang, D.; McMillan-Ward, E.M.; Israels, S.J.; Gibson, S.B. Tyrosine kinase receptor EGFR regulates the switch in cancer cells between cell survival and cell death induced by autophagy in hypoxia. Autophagy 2016, 12, 1029–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Hu, R.; Li, P.; Zheng, Y.; Wang, Y.; Ma, X. DEC1 is required for anti-apoptotic activity of gastric cancer cells under hypoxia by promoting Survivin expression. Gastric Cancer 2018, 21, 632–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.-F.; Xiao, D.-J.; Ma, X.-L.; Song, Y.-Y.; Hu, R.; Kong, Y.; Zheng, Y.; Han, S.-Y.; Hong, R.-L.; Wang, Y.-S. Differentiated embryonic chondrocyte-expressed gene 1 is associated with hypoxia-inducible factor 1α and Ki67 in human gastric cancer. Diagn. Pathol. 2013, 8, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Yang, H.; Luo, W.; Shan, E.; Liu, J.; Zhang, F.; Xi, T.; Yang, J. The anti-metastatic effect of 8-MOP on hepatocellular carcinoma is potentiated by the down-regulation of bHLH transcription factor DEC1. Pharmacol. Res. 2016, 105, 121–133. [Google Scholar] [CrossRef]
- Murakami, K.; Wu, Y.; Imaizumi, T.; Aoki, Y.; Liu, Q.; Yan, X.; Seino, H.; Yoshizawa, T.; Morohashi, S.; Kato, Y.; et al. DEC1 promotes hypoxia-induced epithelial-mesenchymal transition (EMT) in human hepatocellular carcinoma cells. Biomed. Res. 2017, 38, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Nusspaumer, G.; Jaiswal, S.; Barbero, A.; Reinhardt, R.; Ronen, D.I.; Haumer, A.; Lufkin, T.; Martin, I.; Zeller, R. Ontogenic Identification and Analysis of Mesenchymal Stromal Cell Populations during Mouse Limb and Long Bone Development. Stem Cell Rep. 2017, 9, 1124–1138. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Dong, R.; Diao, S.; Yu, G.; Wang, L.; Li, J.; Fan, Z. SFRP2 enhanced the adipogenic and neuronal differentiation potentials of stem cells from apical papilla. Cell Biol. Int. 2017, 41, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-J.; Dong, R.; Wang, L.-P.; Wang, J.-S.; Du, J.; Wang, S.-L.; Shan, Z.-C.; Fan, Z.-P. Histone demethylase KDM2B inhibits the chondrogenic differentiation potentials of stem cells from apical papilla. Int. J. Clin. Exp. Med. 2015, 8, 2165–2173. [Google Scholar] [PubMed]
- Liu, X.; Li, Z.; Liu, H.; Zhu, Y.; Xia, D.; Wang, S.; Gu, R.; Wu, W.; Zhang, P.; Liu, Y.; et al. Low concentration flufenamic acid enhances osteogenic differentiation of mesenchymal stem cells and suppresses bone loss by inhibition of the NF-κB signaling pathway. Stem Cell Res. Ther. 2019, 10, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, Y.; Jin, C.; Zhang, M.; Lv, L.; Zhang, X.; Liu, H.; Zhou, Y. Histone H3K9 Acetyltransferase PCAF Is Essential for Osteogenic Differentiation Through Bone Morphogenetic Protein Signaling and May Be Involved in Osteoporosis. Stem Cells 2016, 34, 2332–2341. [Google Scholar] [CrossRef]
- Min, Z.; Xiaomeng, L.; Zheng, L.; Yangge, D.; Xuejiao, L.; Longwei, L.; Xiao, Z.; Yunsong, L.; Ping, Z.; Yongsheng, Z. Asymmetrical methyltransferase PRMT3 regulates human mesenchymal stem cell osteogenesis via miR-3648. Cell Death Dis. 2019, 10, 581–597. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Liu, Y.; Wang, Y.; Zhang, M.; Lv, L.; Zhang, X.; Zhou, Y. SIRT6 promotes osteogenic differentiation of mesenchymal stem cells through BMP signaling. Sci. Rep. 2017, 7, 10229–10238. [Google Scholar] [CrossRef] [Green Version]
- Nagele, P. Misuse of standard error of the mean (SEM) when reporting variability of a sample. A critical evaluation of four anaesthesia journals. Br. J. Anaesth. 2003, 90, 514–516. [Google Scholar] [CrossRef] [Green Version]
- Barde, M.P.; Barde, P.J. What to use to express the variability of data: Standard deviation or standard error of mean? Perspect Clin. Res. 2012, 3, 113–116. [Google Scholar] [CrossRef]
- Jaykaran, J. “Mean ± SEM” or “Mean (SD)”? Indian J. Pharmacol. 2010, 42, 329. [Google Scholar] [CrossRef]
- Gomez-Larrauri, A.; Gangoiti, P.; Camacho, L.; Presa, N.; Martin, C.; Gomez-Muñoz, A. Phosphatidic Acid Stimulates Lung Cancer Cell Migration through Interaction with the LPA1 Receptor and Subsequent Activation of MAP Kinases and STAT3. Biomedicines 2023, 11, 1804–1821. [Google Scholar] [CrossRef]
- Barbur, I.; Opris, H.; Colosi, H.A.; Baciut, M.; Opris, D.; Cuc, S.; Petean, I.; Moldovan, M.; Dinu, C.M.; Baciut, G. Improving the Mechanical Properties of Orthodontic Occlusal Splints Using Nanoparticles: Silver and Zinc Oxide. Biomedicines 2023, 11, 1965–1979. [Google Scholar] [CrossRef]
- Yue, F.; Cheng, Y.; Breschi, A.; Vierstra, J.; Wu, W.; Ryba, T.; Sandstrom, R.; Ma, Z.; Davis, C.; Pope, B.D.; et al. A comparative encyclopedia of DNA elements in the mouse genome. Nature 2014, 515, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Xie, L.; Wang, Z.; Wang, J.; Xu, H.; Han, X.; Bai, D.; Deng, P. ZBP1 (DAI/DLM-1) promotes osteogenic differentiation while inhibiting adipogenic differentiation in mesenchymal stem cells through a positive feedback loop of Wnt/β-catenin signaling. Bone Res. 2020, 8, 12–21. [Google Scholar] [CrossRef]
- D’amico, R.; Cordaro, M.; Siracusa, R.; Impellizzeri, D.; Salinaro, A.T.; Scuto, M.; Ontario, M.L.; Crea, R.; Cuzzocrea, S.; Di Paola, R.; et al. Wnt/β-Catenin Pathway in Experimental Model of Fibromyalgia: Role of Hidrox®. Biomedicines 2021, 9, 1683–1693. [Google Scholar] [CrossRef]
- Chua, K.-H.; Safwani, W.K.Z.W.; Hamid, A.A.; Shuhup, S.K.; Haflah, N.H.M.; Yahaya, N.H.M. Retropatellar fat pad–derived stem cells from older osteoarthritic patients have lesser differentiation capacity and expression of stemness genes. Cytotherapy 2014, 16, 599–611. [Google Scholar] [CrossRef]
- Chen, Q.; Zhou, H.; Hu, P. Stemness distinctions between the ectomesenchymal stem cells from neonatal and adult mice. Acta Histochem. 2017, 119, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Tratwal, J.; Rojas-Sutterlin, S.; Bataclan, C.; Blum, S.; Naveiras, O. Bone marrow adiposity and the hematopoietic niche: A historical perspective of reciprocity, heterogeneity, and lineage commitment. Best Pr. Res. Clin. Endocrinol. Metab. 2021, 35, 101564–101582. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulos, I.; Brizuela, C.; Khoury, M. Gingival Mesenchymal Stem Cells Outperform Haploidentical Dental Pulp-derived Mesenchymal Stem Cells in Proliferation Rate, Migration Ability, and Angiogenic Potential. Cell Transplant. 2018, 27, 967–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanasekaran, M.; Indumathi, S.; Lissa, R.P.; Harikrishnan, R.; Rajkumar, J.S.; Sudarsanam, D. A comprehensive study on optimization of proliferation and differentiation potency of bone marrow derived mesenchymal stem cells under prolonged culture condition. Cytotechnology 2013, 65, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snippert, H.J.; Clevers, H. Tracking adult stem cells. EMBO Rep. 2011, 12, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Baustian, C.; Hanley, S.; Ceredig, R. Isolation, selection and culture methods to enhance clonogenicity of mouse bone marrow derived mesenchymal stromal cell precursors. Stem Cell Res. Ther. 2015, 6, 151–163. [Google Scholar] [CrossRef] [PubMed]








| Gene | Chr | Score of Significance | Region |
|---|---|---|---|
| Chd7 | chr4 | 406.97581 | genebody |
| Mettl3 | chr14 | 26.26893 | promoter |
| Foxo3 | chr10 | 352.03472 | promoter |
| Foxp1 | chr6 | 84.28053 | promoter |
| Kdm4c | chr4 | 28.58553 | genebody |
| Kdm3a | chr6 | 23.25266 | promoter |
| Kdm2b | chr5 | 27.44679 | promoter |
| Zbp1 | chr2 | 76.28664 | promoter |
| Kdm6a | chrX | 83.51532 | promoter |
| Kdm1a | chr4 | 2.56084 | promoter |
| Cdk6 | chr5 | 3.24094 | promoter |
| P2rx7 | chr5 | 26.09183 | promoter |
| Hoxc10 | chr15 | 29.97397 | promoter |
| Smad3 | chr9 | 105.85294 | promoter |
| Hdac4 | chr1 | 106.92581 | promoter |
| Tet2 | chr3 | 210.94673 | promoter |
| Sirt1 | chr10 | 164.0141 | promoter |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, M.; Tian, Y.; Liu, X.; Guo, Q.; Lu, D.; Wang, X.; Lv, L.; Zhang, X.; Liu, Y.; Zhou, Y.; et al. BHLHE40 Maintains the Stemness of PαS Cells In Vitro by Targeting Zbp1 through the Wnt/β-Catenin Signaling Pathway. Biomedicines 2023, 11, 2190. https://doi.org/10.3390/biomedicines11082190
Hu M, Tian Y, Liu X, Guo Q, Lu D, Wang X, Lv L, Zhang X, Liu Y, Zhou Y, et al. BHLHE40 Maintains the Stemness of PαS Cells In Vitro by Targeting Zbp1 through the Wnt/β-Catenin Signaling Pathway. Biomedicines. 2023; 11(8):2190. https://doi.org/10.3390/biomedicines11082190
Chicago/Turabian StyleHu, Menglong, Yueming Tian, Xuenan Liu, Qian Guo, Dazhuang Lu, Xu Wang, Longwei Lv, Xiao Zhang, Yunsong Liu, Yongsheng Zhou, and et al. 2023. "BHLHE40 Maintains the Stemness of PαS Cells In Vitro by Targeting Zbp1 through the Wnt/β-Catenin Signaling Pathway" Biomedicines 11, no. 8: 2190. https://doi.org/10.3390/biomedicines11082190