
The Secretome of Human Dental Pulp Stem Cells and Its Components GDF15 and HB-EGF Protect Amyotrophic Lateral Sclerosis Motoneurons against Death

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Conditioned Media
2.1.1. DPSC-Conditioned Media
2.1.2. Adipose Derived Stems Cells (ASCs) and Fibroblast-Conditioned Media
2.2. Animals
2.3. Motoneurons Immunopurification and Culture
2.4. Motoneurons Survival
2.5. Axon Length Measurements
2.6. Electrophysiology
2.7. Immunocytochemistry
2.8. Immunohistochemistry
2.9. Quantitative Reverse Transcription Polymerase Chain Reaction
2.10. Statistical Analysis
3. Results
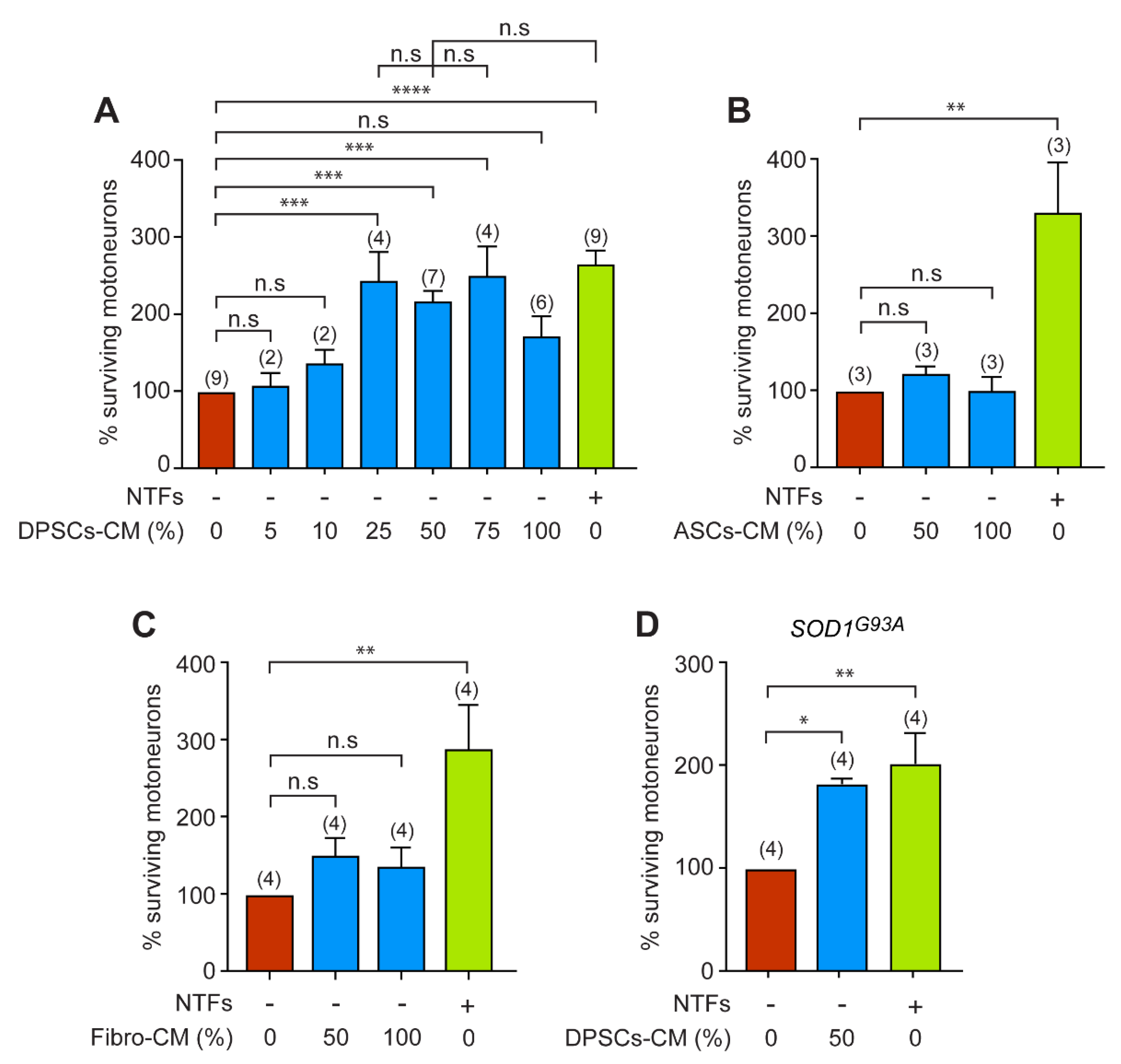
3.1. DPSCs-CM Promotes the Survival of Motoneurons
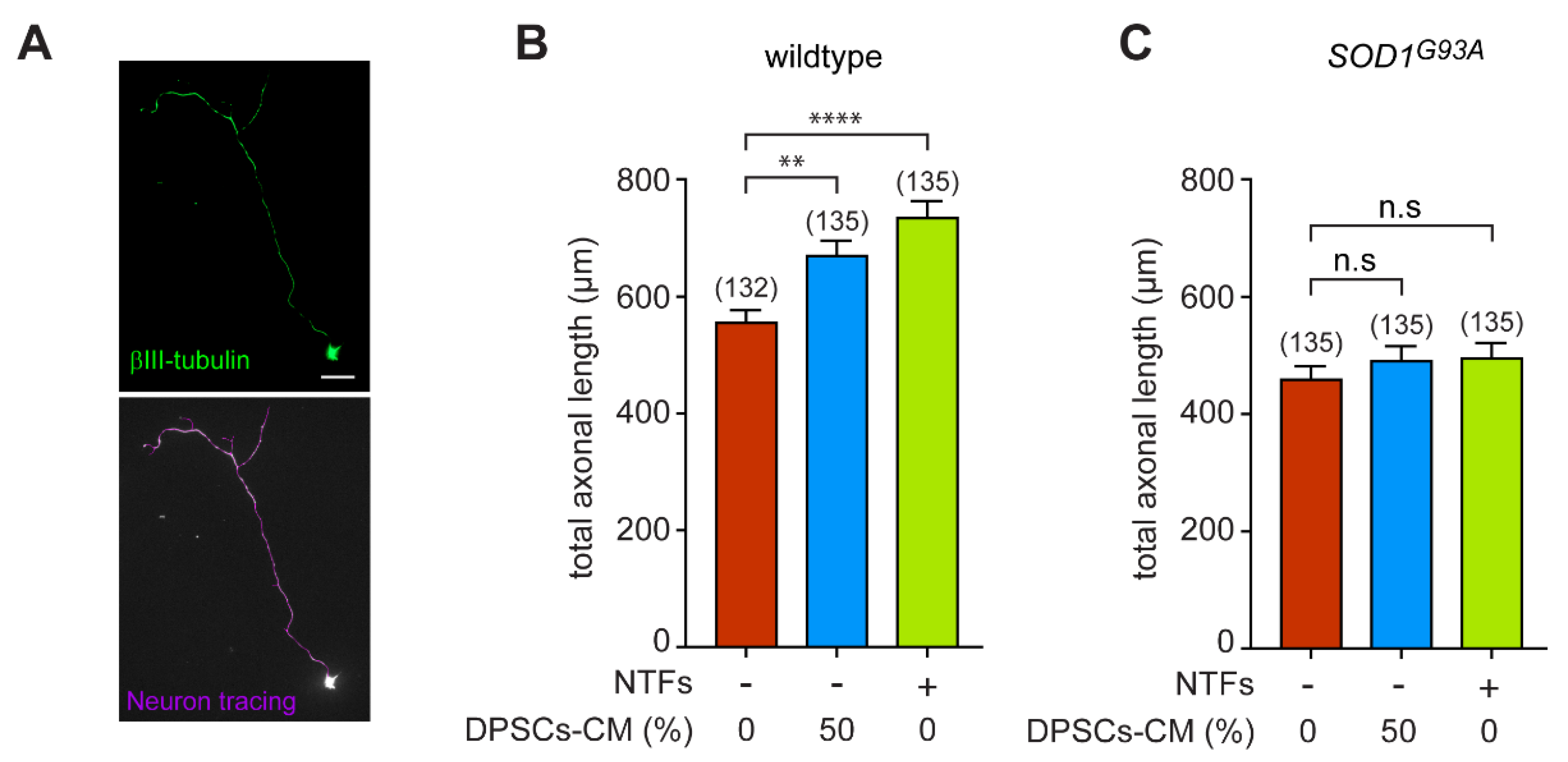
3.2. DPSCs-CM Promotes Axon Outgrowth of Wildtype, but Not SOD1G93A Motoneurons
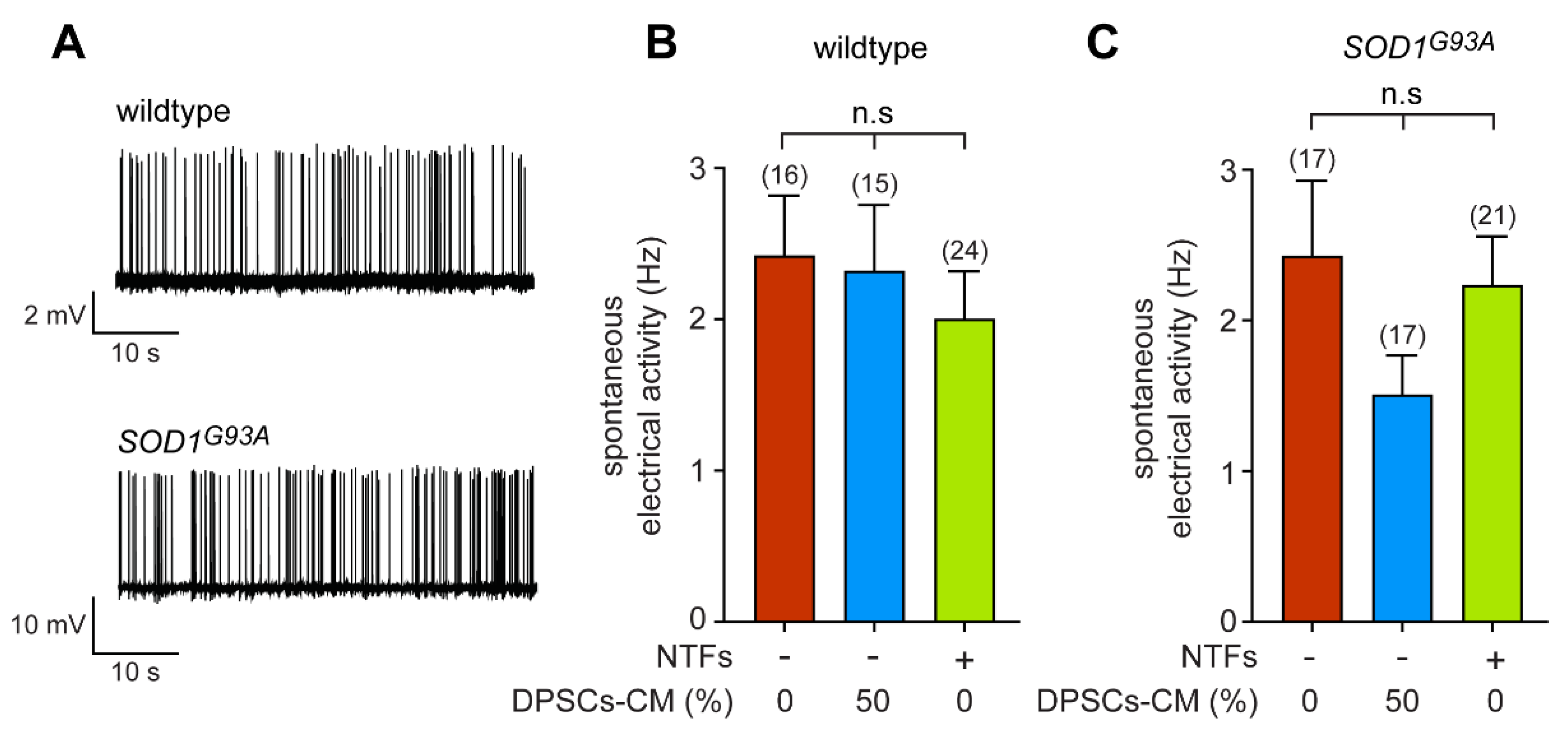
3.3. DPSCs-CM Does Not Modify the Synaptically Driven Activity of Wildtype and SOD1G93A Motoneurons
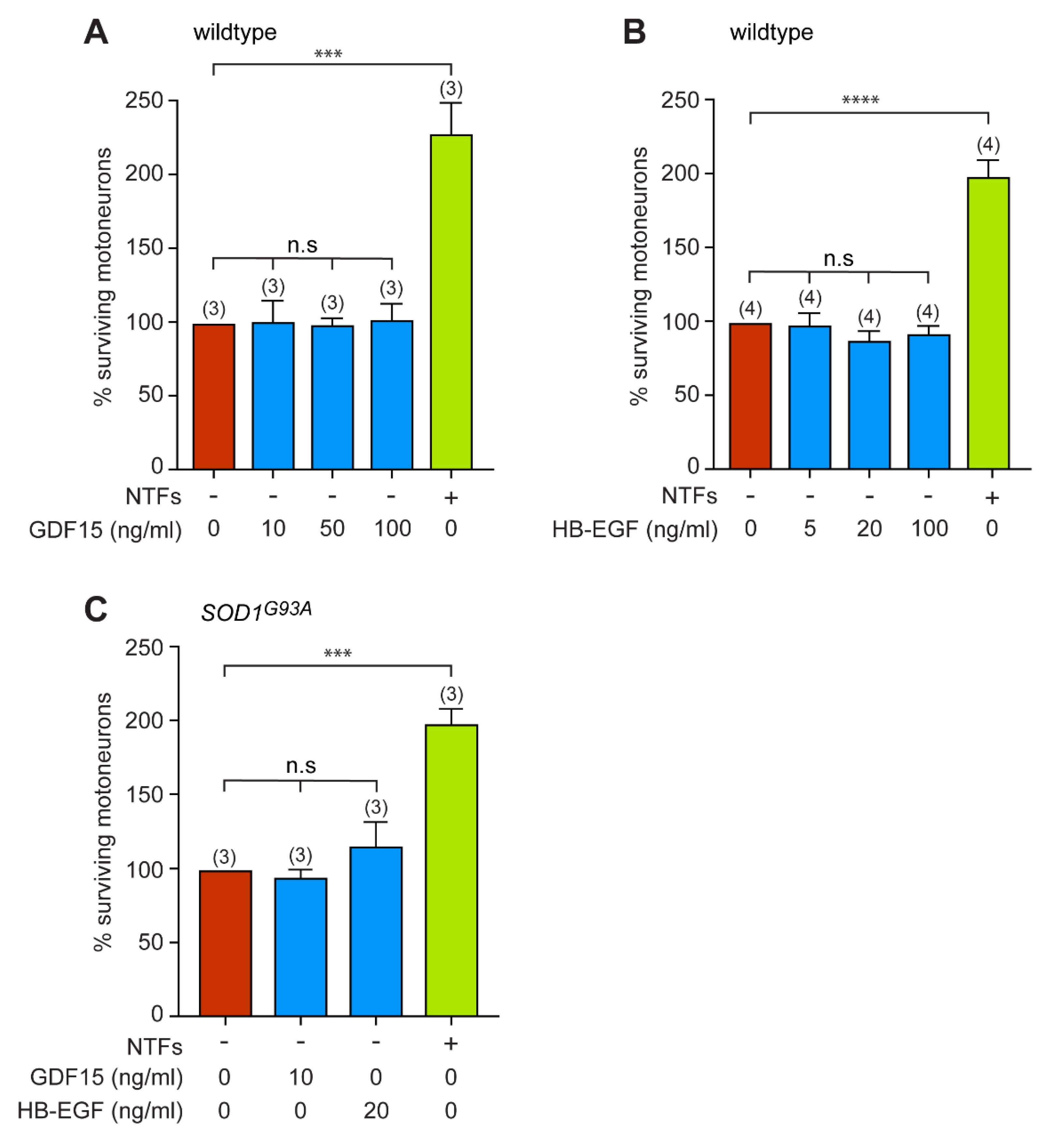
3.4. GDF15 and HB-EGF Do Not Provide Any Neutrotrophic Support to Motoneurons
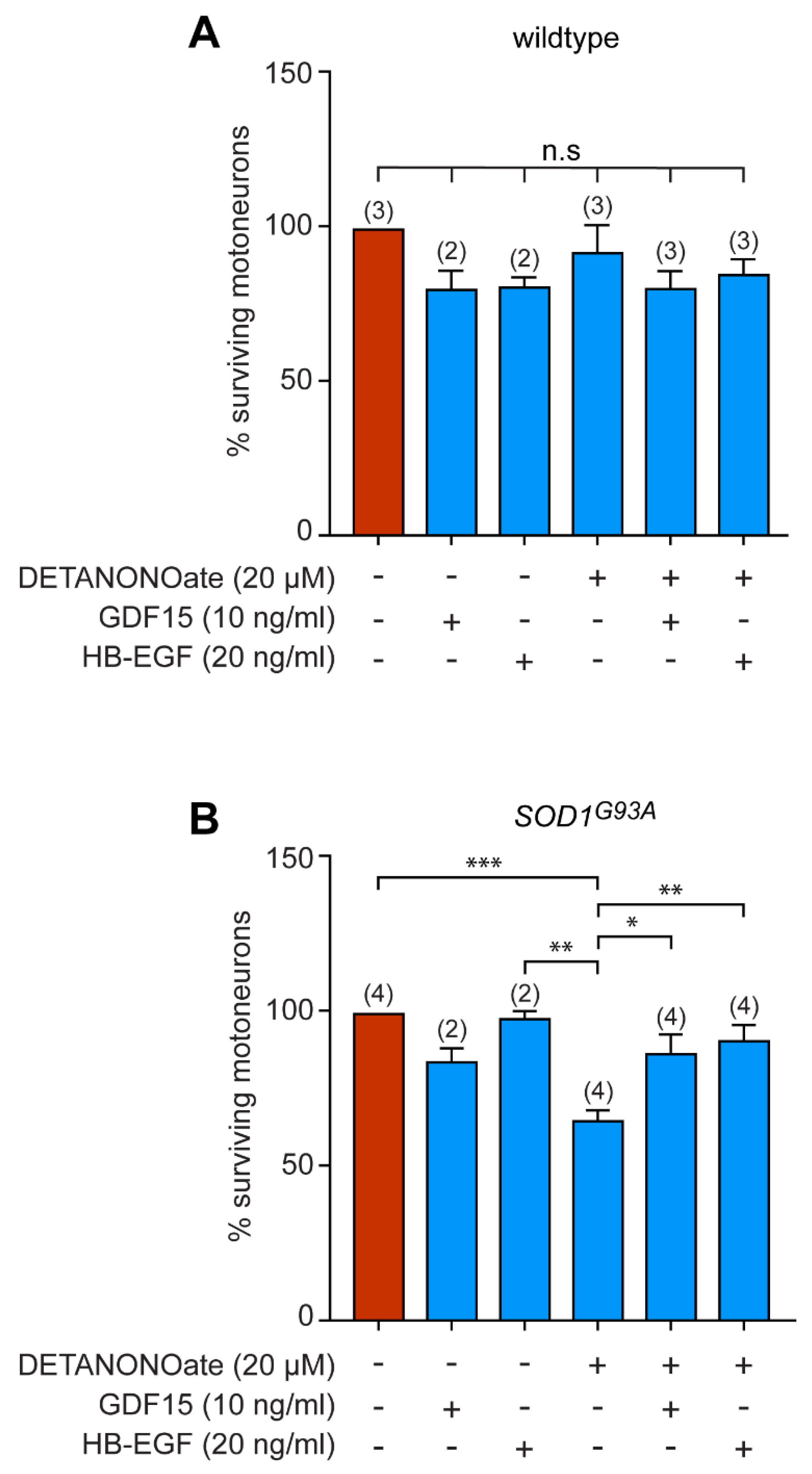
3.5. GDF15 and HB-EGF Prevent Motoneuron Death from Oxidative Insult
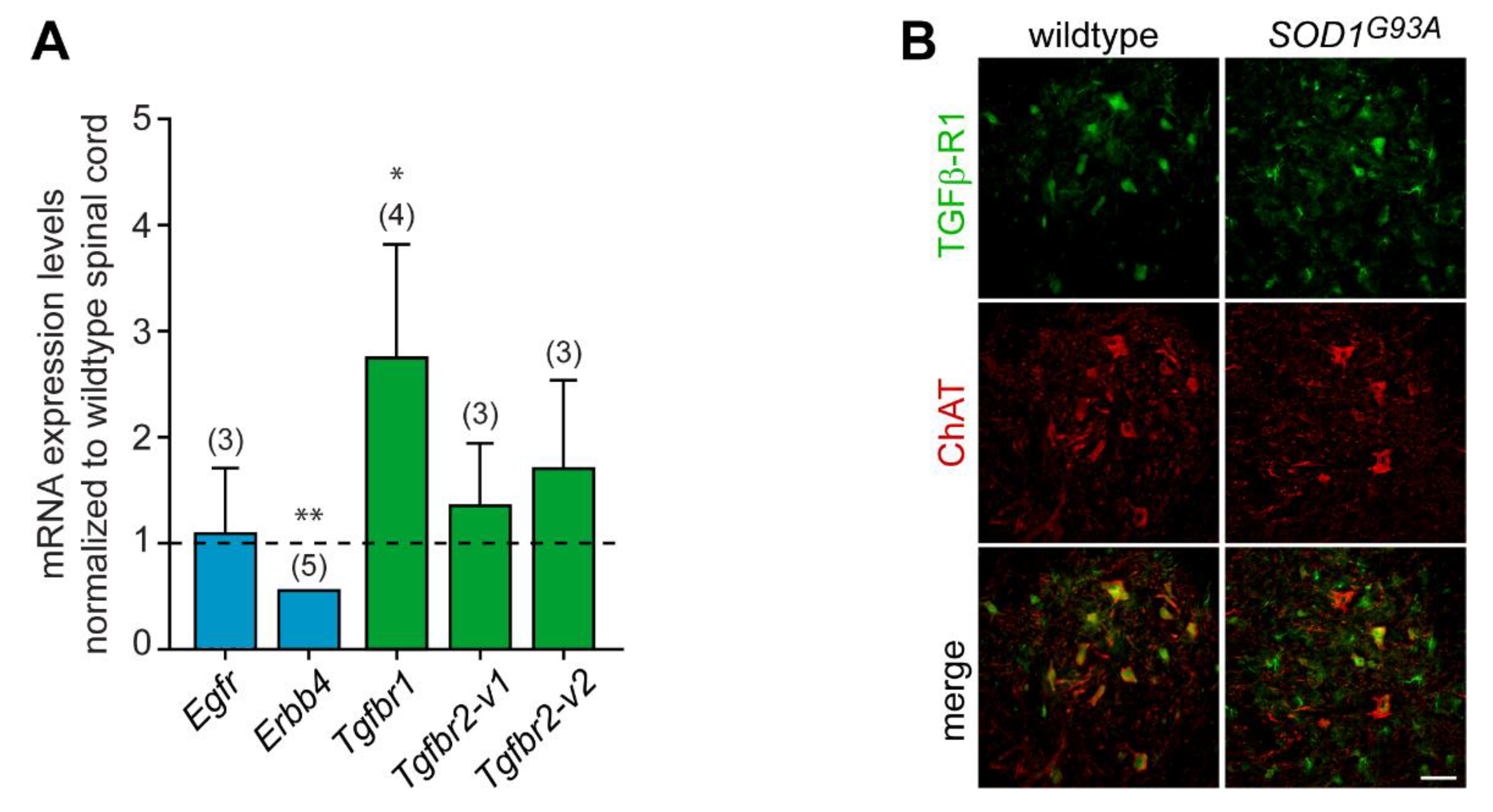
3.6. Expression of GDF15 and HB-EGF in Adult Spinal Cord
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, J.P.; Brown, R.H.; Cleveland, D.W. Decoding ALS: From Genes to Mechanism. Nature 2016, 539, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robberecht, W.; Philips, T. The Changing Scene of Amyotrophic Lateral Sclerosis. Nat. Rev. Neurosci. 2013, 14, 248–264. [Google Scholar] [CrossRef] [PubMed]
- Gosset, P.; Camu, W.; Raoul, C.; Mezghrani, A. Prionoids in Amyotrophic Lateral Sclerosis. Brain Commun. 2022, 4, fcac145. [Google Scholar] [CrossRef] [PubMed]
- Medinas, D.B.; Valenzuela, V.; Hetz, C. Proteostasis Disturbance in Amyotrophic Lateral Sclerosis. Hum. Mol. Genet. 2017, 26, R91–R104. [Google Scholar] [CrossRef]
- Crabé, R.; Aimond, F.; Gosset, P.; Scamps, F.; Raoul, C. How Degeneration of Cells Surrounding Motoneurons Contributes to Amyotrophic Lateral Sclerosis. Cells 2020, 9, 2550. [Google Scholar] [CrossRef]
- López-Pingarrón, L.; Almeida, H.; Soria-Aznar, M.; Reyes-Gonzales, M.C.; Terrón, M.P.; García, J.J. Role of Oxidative Stress on the Etiology and Pathophysiology of Amyotrophic Lateral Sclerosis (ALS) and Its Relation with the Enteric Nervous System. CIMB 2023, 45, 3315–3332. [Google Scholar] [CrossRef]
- Rothstein, J.D. Edaravone: A New Drug Approved for ALS. Cell 2017, 171, 725. [Google Scholar] [CrossRef]
- Finegan, E.; Hi Shing, S.L.; Chipika, R.H.; McKenna, M.C.; Doherty, M.A.; Hengeveld, J.C.; Vajda, A.; Donaghy, C.; McLaughlin, R.L.; Hutchinson, S.; et al. Thalamic, Hippocampal and Basal Ganglia Pathology in Primary Lateral Sclerosis and Amyotrophic Lateral Sclerosis: Evidence from Quantitative Imaging Data. Data Brief 2020, 29, 105115. [Google Scholar] [CrossRef]
- Chipika, R.H.; Mulkerrin, G.; Pradat, P.-F.; Murad, A.; Ango, F.; Raoul, C.; Bede, P. Cerebellar Pathology in Motor Neuron Disease: Neuroplasticity and Neurodegeneration. Neural Regen. Res. 2022, 17, 2335–2341. [Google Scholar] [CrossRef]
- Chipika, R.H.; Finegan, E.; Li Hi Shing, S.; McKenna, M.C.; Christidi, F.; Chang, K.M.; Doherty, M.A.; Hengeveld, J.C.; Vajda, A.; Pender, N.; et al. “Switchboard” Malfunction in Motor Neuron Diseases: Selective Pathology of Thalamic Nuclei in Amyotrophic Lateral Sclerosis and Primary Lateral Sclerosis. Neuroi. Clin. 2020, 27, 102300. [Google Scholar] [CrossRef]
- Debye, B.; Schmülling, L.; Zhou, L.; Rune, G.; Beyer, C.; Johann, S. Neurodegeneration and NLRP3 Inflammasome Expression in the Anterior Thalamus of SOD1(G93A) ALS Mice. Brain Pathol. 2018, 28, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Fisher, E.M.C.; Greensmith, L.; Malaspina, A.; Fratta, P.; Hanna, M.G.; Schiavo, G.; Isaacs, A.M.; Orrell, R.W.; Cunningham, T.J.; Arozena, A.A. Opinion: More Mouse Models and More Translation Needed for ALS. Mol. Neurodegener. 2023, 18, 30. [Google Scholar] [CrossRef]
- Bonifacino, T.; Zerbo, R.A.; Balbi, M.; Torazza, C.; Frumento, G.; Fedele, E.; Bonanno, G.; Milanese, M. Nearly 30 Years of Animal Models to Study Amyotrophic Lateral Sclerosis: A Historical Overview and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 12236. [Google Scholar] [CrossRef]
- Bruijn, L.I.; Miller, T.M.; Cleveland, D.W. Unraveling the Mechanisms Involved in Motor Neuron Degeneration in ALS. Annu. Rev. Neurosci. 2004, 27, 723–749. [Google Scholar] [CrossRef] [Green Version]
- Lazarevic, V.; Yang, Y.; Ivanova, D.; Fejtova, A.; Svenningsson, P. Riluzole Attenuates the Efficacy of Glutamatergic Transmission by Interfering with the Size of the Readily Releasable Neurotransmitter Pool. Neuropharmacology 2018, 143, 38–48. [Google Scholar] [CrossRef]
- Abe, S.; Kirima, K.; Tsuchiya, K.; Okamoto, M.; Hasegawa, T.; Houchi, H.; Yoshizumi, M.; Tamaki, T. The Reaction Rate of Edaravone (3-Methyl-1-Phenyl-2-Pyrazolin-5-One (MCI-186)) with Hydroxyl Radical. Chem. Pharm. Bull. (Tokyo) 2004, 52, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, A.; Yamamoto, Y. Edaravone, a Potent Free Radical Scavenger, Reacts with Peroxynitrite to Produce Predominantly 4-NO-Edaravone. Redox Rep. 2016, 21, 98–103. [Google Scholar] [CrossRef]
- Takei, K.; Watanabe, K.; Yuki, S.; Akimoto, M.; Sakata, T.; Palumbo, J. Edaravone and Its Clinical Development for Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2017, 18, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Miller, T.M.; Cudkowicz, M.E.; Genge, A.; Shaw, P.J.; Sobue, G.; Bucelli, R.C.; Chiò, A.; Van Damme, P.; Ludolph, A.C.; Glass, J.D.; et al. Trial of Antisense Oligonucleotide Tofersen for SOD1 ALS. N. Engl. J. Med. 2022, 387, 1099–1110. [Google Scholar] [CrossRef]
- Kanning, K.C.; Kaplan, A.; Henderson, C.E. Motor Neuron Diversity in Development and Disease. Annu. Rev. Neurosci. 2010, 33, 409–440. [Google Scholar] [CrossRef]
- Tovar-y-Romo, L.B.; Ramírez-Jarquín, U.N.; Lazo-Gómez, R.; Tapia, R. Trophic Factors as Modulators of Motor Neuron Physiology and Survival: Implications for ALS Therapy. Front. Cell. Neurosci. 2014, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartus, R.T.; Johnson, E.M. Clinical Tests of Neurotrophic Factors for Human Neurodegenerative Diseases, Part 1: Where Have We Been and What Have We Learned? Neurobiol. Dis. 2017, 97, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Cappella, M.; Pradat, P.-F.; Querin, G.; Biferi, M.G. Beyond the Traditional Clinical Trials for Amyotrophic Lateral Sclerosis and The Future Impact of Gene Therapy. J. Neuromuscul. Dis. 2021, 8, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Nosrat, I.V.; Widenfalk, J.; Olson, L.; Nosrat, C.A. Dental Pulp Cells Produce Neurotrophic Factors, Interact with Trigeminal Neurons in Vitro, and Rescue Motoneurons after Spinal Cord Injury. Dev. Biol. 2001, 238, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Nosrat, I.V.; Smith, C.A.; Mullally, P.; Olson, L.; Nosrat, C.A. Dental Pulp Cells Provide Neurotrophic Support for Dopaminergic Neurons and Differentiate into Neurons in Vitro; Implications for Tissue Engineering and Repair in the Nervous System. Eur. J. Neurosci. 2004, 19, 2388–2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolar, M.K.; Itte, V.N.; Kingham, P.J.; Novikov, L.N.; Wiberg, M.; Kelk, P. The Neurotrophic Effects of Different Human Dental Mesenchymal Stem Cells. Sci. Rep. 2017, 7, 12605. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Gan, Q.F.; Kathivaloo, P.; Mohamad, N.A.; Dhamodharan, J.; Krishnan, A.; Sengodan, B.; Palanimuthu, V.R.; Marimuthu, K.; Rajandas, H.; et al. Deciduous DPSCs Ameliorate MPTP-Mediated Neurotoxicity, Sensorimotor Coordination and Olfactory Function in Parkinsonian Mice. Int. J. Mol. Sci. 2019, 20, 568. [Google Scholar] [CrossRef] [Green Version]
- Ueda, T.; Inden, M.; Ito, T.; Kurita, H.; Hozumi, I. Characteristics and Therapeutic Potential of Dental Pulp Stem Cells on Neurodegenerative Diseases. Front. Neurosci. 2020, 14, 407. [Google Scholar] [CrossRef]
- Wang, J.; Zuzzio, K.; Walker, C.L. Systemic Dental Pulp Stem Cell Secretome Therapy in a Mouse Model of Amyotrophic Lateral Sclerosis. Brain Sci. 2019, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-M.; Ouyang, Y.-J.; Yu, B.-Q.; Li, W.; Yu, M.-Y.; Li, J.-Y.; Jiao, Z.-M.; Yang, D.; Li, N.; Shi, Y.; et al. Therapeutic Potential of Dental Pulp Stem Cell Transplantation in a Rat Model of Alzheimer’s Disease. Neural Regen. Res. 2021, 16, 893–898. [Google Scholar] [CrossRef]
- Sandonà, M.; Di Pietro, L.; Esposito, F.; Ventura, A.; Silini, A.R.; Parolini, O.; Saccone, V. Mesenchymal Stromal Cells and Their Secretome: New Therapeutic Perspectives for Skeletal Muscle Regeneration. Front. Bioeng. Biotechnol. 2021, 9, 652970. [Google Scholar] [CrossRef]
- Frisbie, L.; Buckanovich, R.J.; Coffman, L. Carcinoma-Associated Mesenchymal Stem/Stromal Cells: Architects of the Pro-Tumorigenic Tumor Microenvironment. Stem Cells 2022, 40, 705–715. [Google Scholar] [CrossRef]
- Luo, L.; Albashari, A.A.; Wang, X.; Jin, L.; Zhang, Y.; Zheng, L.; Xia, J.; Xu, H.; Zhao, Y.; Xiao, J.; et al. Effects of Transplanted Heparin-Poloxamer Hydrogel Combining Dental Pulp Stem Cells and BFGF on Spinal Cord Injury Repair. Stem Cells Int. 2018, 2018, 2398521. [Google Scholar] [CrossRef] [Green Version]
- Király, M.; Kádár, K.; Horváthy, D.B.; Nardai, P.; Rácz, G.Z.; Lacza, Z.; Varga, G.; Gerber, G. Integration of Neuronally Predifferentiated Human Dental Pulp Stem Cells into Rat Brain In Vivo. Neurochem. Int. 2011, 59, 371–381. [Google Scholar] [CrossRef]
- Winderlich, J.N.; Kremer, K.L.; Koblar, S.A. Adult Human Dental Pulp Stem Cells Promote Blood-Brain Barrier Permeability through Vascular Endothelial Growth Factor-a Expression. J. Cereb. Blood Flow Metab. 2016, 36, 1087–1097. [Google Scholar] [CrossRef] [Green Version]
- Nito, C.; Sowa, K.; Nakajima, M.; Sakamoto, Y.; Suda, S.; Nishiyama, Y.; Nakamura-Takahashi, A.; Nitahara-Kasahara, Y.; Ueda, M.; Okada, T.; et al. Transplantation of Human Dental Pulp Stem Cells Ameliorates Brain Damage Following Acute Cerebral Ischemia. Biomed. Pharmacother. 2018, 108, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-F.; Chen, K.-W.; Chien, Y.; Lai, Y.-H.; Hsieh, S.-T.; Ma, H.-Y.; Wang, K.-C.; Shiau, C.-Y. Dental Pulp Stem Cell-Derived Factors Alleviate Subarachnoid Hemorrhage-Induced Neuroinflammation and Ischemic Neurological Deficits. Int. J. Mol. Sci. 2019, 20, 3747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, A.F.; Cevallos, R.R.; Gazarian, K.; Lamas, M. Human Dental Pulp Stem Cells Respond to Cues from the Rat Retina and Differentiate to Express the Retinal Neuronal Marker Rhodopsin. Neuroscience 2014, 280, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Chouaib, B.; Collart-Dutilleul, P.-Y.; Blanc-Sylvestre, N.; Younes, R.; Gergely, C.; Raoul, C.; Scamps, F.; Cuisinier, F.; Romieu, O. Identification of Secreted Factors in Dental Pulp Cell-Conditioned Medium Optimized for Neuronal Growth. Neurochem. Int. 2021, 144, 104961. [Google Scholar] [CrossRef]
- Chouaib, B.; Cuisinier, F.; Collart-Dutilleul, P.-Y. Dental Stem Cell-Conditioned Medium for Tissue Regeneration: Optimization of Production and Storage. World J. Stem Cells 2022, 14, 287–302. [Google Scholar] [CrossRef]
- Collart-Dutilleul, P.-Y.; Secret, E.; Panayotov, I.; Deville De Périère, D.; Martín-Palma, R.J.; Torres-Costa, V.; Martin, M.; Gergely, C.; Durand, J.-O.; Cunin, F.; et al. Adhesion and Proliferation of Human Mesenchymal Stem Cells from Dental Pulp on Porous Silicon Scaffolds. ACS Appl. Mater. Interfaces 2014, 6, 1719–1728. [Google Scholar] [CrossRef]
- Panayotov, I.V.; Collart-Dutilleul, P.-Y.; Salehi, H.; Martin, M.; Végh, A.; Yachouh, J.; Vladimirov, B.; Sipos, P.; Szalontai, B.; Gergely, C.; et al. Sprayed Cells and Polyelectrolyte Films for Biomaterial Functionalization: The Influence of Physical PLL-PGA Film Treatments on Dental Pulp Cell Behavior. Macromol. Biosci. 2014, 14, 1771–1782. [Google Scholar] [CrossRef] [Green Version]
- Soulard, C.; Salsac, C.; Mouzat, K.; Hilaire, C.; Roussel, J.; Mezghrani, A.; Lumbroso, S.; Raoul, C.; Scamps, F. Spinal Motoneuron TMEM16F Acts at C-Boutons to Modulate Motor Resistance and Contributes to ALS Pathogenesis. Cell Rep. 2020, 30, 2581–2593.e7. [Google Scholar] [CrossRef] [Green Version]
- Raoul, C.; Estévez, A.G.; Nishimune, H.; Cleveland, D.W.; deLapeyrière, O.; Henderson, C.E.; Haase, G.; Pettmann, B. Motoneuron Death Triggered by a Specific Pathway Downstream of Fas. Potentiation by ALS-Linked SOD1 Mutations. Neuron 2002, 35, 1067–1083. [Google Scholar] [CrossRef] [Green Version]
- Benlefki, S.; Sanchez-Vicente, A.; Milla, V.; Lucas, O.; Soulard, C.; Younes, R.; Gergely, C.; Bowerman, M.; Raoul, C.; Scamps, F.; et al. Expression of ALS-Linked SOD1 Mutation in Motoneurons or Myotubes Induces Differential Effects on Neuromuscular Function In Vitro. Neuroscience 2020, 435, 33–43. [Google Scholar] [CrossRef]
- Benlefki, S.; Younes, R.; Challuau, D.; Bernard-Marissal, N.; Hilaire, C.; Scamps, F.; Bowerman, M.; Kothary, R.; Schneider, B.L.; Raoul, C. Differential Effect of Fas Activation on Spinal Muscular Atrophy Motoneuron Death and Induction of Axonal Growth. Cell. Mol. Biol. 2023; in press. [Google Scholar]
- Otsmane, B.; Moumen, A.; Aebischer, J.; Coque, E.; Sar, C.; Sunyach, C.; Salsac, C.; Valmier, J.; Salinas, S.; Bowerman, M.; et al. Somatic and Axonal LIGHT Signaling Elicit Degenerative and Regenerative Responses in Motoneurons, Respectively. EMBO Rep. 2014, 15, 540–547. [Google Scholar] [CrossRef] [Green Version]
- Bowerman, M.; Salsac, C.; Bernard, V.; Soulard, C.; Dionne, A.; Coque, E.; Benlefki, S.; Hince, P.; Dion, P.A.; Butler-Browne, G.; et al. KCC3 Loss-of-Function Contributes to Andermann Syndrome by Inducing Activity-Dependent Neuromuscular Junction Defects. Neurobiol. Dis. 2017, 106, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Gancheva, M.R.; Kremer, K.L.; Gronthos, S.; Koblar, S.A. Using Dental Pulp Stem Cells for Stroke Therapy. Front. Neurol. 2019, 10, 422. [Google Scholar] [CrossRef]
- Zhou, Y.; Besner, G.E. Heparin-Binding Epidermal Growth Factor-like Growth Factor Is a Potent Neurotrophic Factor for PC12 Cells. Neurosignals 2010, 18, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Strelau, J.; Strzelczyk, A.; Rusu, P.; Bendner, G.; Wiese, S.; Diella, F.; Altick, A.L.; von Bartheld, C.S.; Klein, R.; Sendtner, M.; et al. Progressive Postnatal Motoneuron Loss in Mice Lacking GDF-15. J. Neurosci. 2009, 29, 13640–13648. [Google Scholar] [CrossRef] [Green Version]
- Saxena, S.; Cabuy, E.; Caroni, P. A Role for Motoneuron Subtype-Selective ER Stress in Disease Manifestations of FALS Mice. Nat. Neurosci. 2009, 12, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, O.; Jedrychowski, M.P.; Moore, C.S.; Cialic, R.; Lanser, A.J.; Gabriely, G.; Koeglsperger, T.; Dake, B.; Wu, P.M.; Doykan, C.E.; et al. Identification of a Unique TGF-β-Dependent Molecular and Functional Signature in Microglia. Nat. Neurosci. 2014, 17, 131–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spittau, B.; Dokalis, N.; Prinz, M. The Role of TGFβ Signaling in Microglia Maturation and Activation. Trends Immunol. 2020, 41, 836–848. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Mazzon, E. Dental Mesenchymal Stem Cell Secretome: An Intriguing Approach for Neuroprotection and Neuroregeneration. Int. J. Mol. Sci. 2021, 23, 456. [Google Scholar] [CrossRef] [PubMed]
- Mead, B.; Logan, A.; Berry, M.; Leadbeater, W.; Scheven, B.A. Dental Pulp Stem Cells, a Paracrine-Mediated Therapy for the Retina. Neural Regen. Res. 2014, 9, 577–578. [Google Scholar] [CrossRef]
- Keefe, K.M.; Sheikh, I.S.; Smith, G.M. Targeting Neurotrophins to Specific Populations of Neurons: NGF, BDNF, and NT-3 and Their Relevance for Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef] [Green Version]
- Calvo, P.M.; Hernández, R.G.; de la Cruz, R.R.; Pastor, A.M. Role of Vascular Endothelial Growth Factor as a Critical Neurotrophic Factor for the Survival and Physiology of Motoneurons. Neural Regen. Res. 2023, 18, 1691–1696. [Google Scholar] [CrossRef]
- Scamps, F.; Aimond, F.; Hilaire, C.; Raoul, C. Synaptic Transmission and Motoneuron Excitability Defects in Amyotrophic Lateral Sclerosis. In Amyotrophic Lateral Sclerosis; Araki, T., Ed.; Exon Publications: Brisbane, Australia, 2021; ISBN 978-0-645-00177-8. [Google Scholar]
- Fontanilla, C.V.; Gu, H.; Liu, Q.; Zhu, T.Z.; Zhou, C.; Johnstone, B.H.; March, K.L.; Pascuzzi, R.M.; Farlow, M.R.; Du, Y. Adipose-Derived Stem Cell Conditioned Media Extends Survival Time of a Mouse Model of Amyotrophic Lateral Sclerosis. Sci. Rep. 2015, 5, 16953. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.L.; Meadows, R.M.; Merfeld-Clauss, S.; Du, Y.; March, K.L.; Jones, K.J. Adipose-Derived Stem Cell Conditioned Medium Impacts Asymptomatic Peripheral Neuromuscular Denervation in the Mutant Superoxide Dismutase (G93A) Transgenic Mouse Model of Amyotrophic Lateral Sclerosis. Restor. Neurol. Neurosci. 2018, 36, 621–627. [Google Scholar] [CrossRef]
- Ciervo, Y.; Gatto, N.; Allen, C.; Grierson, A.; Ferraiuolo, L.; Mead, R.J.; Shaw, P.J. Adipose-Derived Stem Cells Protect Motor Neurons and Reduce Glial Activation in Both in Vitro and in Vivo Models of ALS. Mol. Ther. Methods Clin. Dev. 2021, 21, 413–433. [Google Scholar] [CrossRef]
- Tosolini, A.P.; Sleigh, J.N.; Surana, S.; Rhymes, E.R.; Cahalan, S.D.; Schiavo, G. BDNF-Dependent Modulation of Axonal Transport Is Selectively Impaired in ALS. Acta Neuropathol. Commun. 2022, 10, 121. [Google Scholar] [CrossRef]
- Higashiyama, S.; Abraham, J.A.; Miller, J.; Fiddes, J.C.; Klagsbrun, M. A Heparin-Binding Growth Factor Secreted by Macrophage-like Cells That Is Related to EGF. Science 1991, 251, 936–939. [Google Scholar] [CrossRef]
- Nakagawa, T.; Sasahara, M.; Hayase, Y.; Haneda, M.; Yasuda, H.; Kikkawa, R.; Higashiyama, S.; Hazama, F. Neuronal and Glial Expression of Heparin-Binding EGF-like Growth Factor in Central Nervous System of Prenatal and Early-Postnatal Rat. Brain Res. Dev. Brain Res. 1998, 108, 263–272. [Google Scholar] [CrossRef]
- Chouaib, B. Dental Pulp Stem Cell-Conditioned Medium for Tissue Regeneration. Ph.D. Thesis, Montpellier University, Montpellier, France, 2020. [Google Scholar]
- Opanashuk, L.A.; Mark, R.J.; Porter, J.; Damm, D.; Mattson, M.P.; Seroogy, K.B. Heparin-Binding Epidermal Growth Factor-like Growth Factor in Hippocampus: Modulation of Expression by Seizures and Anti-Excitotoxic Action. J. Neurosci. 1999, 19, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Maurya, S.K.; Mishra, J.; Abbas, S.; Bandyopadhyay, S. Cypermethrin Stimulates GSK3β-Dependent Aβ and p-Tau Proteins and Cognitive Loss in Young Rats: Reduced HB-EGF Signaling and Downstream Neuroinflammation as Critical Regulators. Mol. Neurobiol. 2016, 53, 968–982. [Google Scholar] [CrossRef]
- Wiese, S.; Pei, G.; Karch, C.; Troppmair, J.; Holtmann, B.; Rapp, U.R.; Sendtner, M. Specific Function of B-Raf in Mediating Survival of Embryonic Motoneurons and Sensory Neurons. Nat. Neurosci. 2001, 4, 137–142. [Google Scholar] [CrossRef]
- Lamballe, F.; Genestine, M.; Caruso, N.; Arce, V.; Richelme, S.; Helmbacher, F.; Maina, F. Pool-Specific Regulation of Motor Neuron Survival by Neurotrophic Support. J. Neurosci. 2011, 31, 11144–11158. [Google Scholar] [CrossRef]
- Xia, M.; Zhang, Q.; Zhang, Y.; Li, R.; Zhao, T.; Chen, L.; Liu, Q.; Zheng, S.; Li, H.; Qian, Z.; et al. Growth Differentiation Factor 15 Regulates Oxidative Stress-Dependent Ferroptosis Post Spinal Cord Injury by Stabilizing the P62-Keap1-Nrf2 Signaling Pathway. Front. Aging Neurosci. 2022, 14, 905115. [Google Scholar] [CrossRef]
- Nichterwitz, S.; Nijssen, J.; Storvall, H.; Schweingruber, C.; Comley, L.H.; Allodi, I.; van der Lee, M.; Deng, Q.; Sandberg, R.; Hedlund, E. LCM-Seq Reveals Unique Transcriptional Adaptation Mechanisms of Resistant Neurons and Identifies Protective Pathways in Spinal Muscular Atrophy. Genome Res. 2020, 30, 1083–1096. [Google Scholar] [CrossRef]
- Jennings, M.J.; Kagiava, A.; Vendredy, L.; Spaulding, E.L.; Stavrou, M.; Hathazi, D.; Grüneboom, A.; De Winter, V.; Gess, B.; Schara, U.; et al. NCAM1 and GDF15 Are Biomarkers of Charcot-Marie-Tooth Disease in Patients and Mice. Brain 2022, 145, 3999–4015. [Google Scholar] [CrossRef] [PubMed]
- Lasiene, J.; Komine, O.; Fujimori-Tonou, N.; Powers, B.; Endo, F.; Watanabe, S.; Shijie, J.; Ravits, J.; Horner, P.; Misawa, H.; et al. Neuregulin 1 Confers Neuroprotection in SOD1-Linked Amyotrophic Lateral Sclerosis Mice via Restoration of C-Boutons of Spinal Motor Neurons. Acta Neuropathol. Commun. 2016, 4, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Uchino, A.; Shioya, A.; Sano, T.; Matsumoto, C.; Numata-Uematsu, Y.; Nagano, S.; Araki, T.; Murayama, S.; Saito, Y. Altered Immunoreactivity of ErbB4, a Causative Gene Product for ALS19, in the Spinal Cord of Patients with Sporadic ALS. Neuropathology 2019, 39, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Genestine, M.; Caricati, E.; Fico, A.; Richelme, S.; Hassani, H.; Sunyach, C.; Lamballe, F.; Panzica, G.C.; Pettmann, B.; Helmbacher, F.; et al. Enhanced Neuronal Met Signalling Levels in ALS Mice Delay Disease Onset. Cell Death Dis. 2011, 2, e130. [Google Scholar] [CrossRef] [Green Version]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Insights Into Mechanisms of GDF15 and Receptor GFRAL: Therapeutic Targets. Trends Endocrinol. Metab. 2020, 31, 939–951. [Google Scholar] [CrossRef]
- Galbiati, M.; Crippa, V.; Rusmini, P.; Cristofani, R.; Messi, E.; Piccolella, M.; Tedesco, B.; Ferrari, V.; Casarotto, E.; Chierichetti, M.; et al. Multiple Roles of Transforming Growth Factor Beta in Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2020, 21, 4291. [Google Scholar] [CrossRef]
- Iglesias, C.; Sangari, S.; El Mendili, M.-M.; Benali, H.; Marchand-Pauvert, V.; Pradat, P.-F. Electrophysiological and Spinal Imaging Evidences for Sensory Dysfunction in Amyotrophic Lateral Sclerosis. BMJ Open 2015, 5, e007659. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Younes, R.; Issa, Y.; Jdaa, N.; Chouaib, B.; Brugioti, V.; Challuau, D.; Raoul, C.; Scamps, F.; Cuisinier, F.; Hilaire, C. The Secretome of Human Dental Pulp Stem Cells and Its Components GDF15 and HB-EGF Protect Amyotrophic Lateral Sclerosis Motoneurons against Death. Biomedicines 2023, 11, 2152. https://doi.org/10.3390/biomedicines11082152
Younes R, Issa Y, Jdaa N, Chouaib B, Brugioti V, Challuau D, Raoul C, Scamps F, Cuisinier F, Hilaire C. The Secretome of Human Dental Pulp Stem Cells and Its Components GDF15 and HB-EGF Protect Amyotrophic Lateral Sclerosis Motoneurons against Death. Biomedicines. 2023; 11(8):2152. https://doi.org/10.3390/biomedicines11082152
Chicago/Turabian StyleYounes, Richard, Youssef Issa, Nadia Jdaa, Batoul Chouaib, Véronique Brugioti, Désiré Challuau, Cédric Raoul, Frédérique Scamps, Frédéric Cuisinier, and Cécile Hilaire. 2023. "The Secretome of Human Dental Pulp Stem Cells and Its Components GDF15 and HB-EGF Protect Amyotrophic Lateral Sclerosis Motoneurons against Death" Biomedicines 11, no. 8: 2152. https://doi.org/10.3390/biomedicines11082152