

CircAFF1 Is a Circular RNA with a Role in Alveolar Rhabdomyosarcoma Cell Migration

 , , , , and
, , , , and 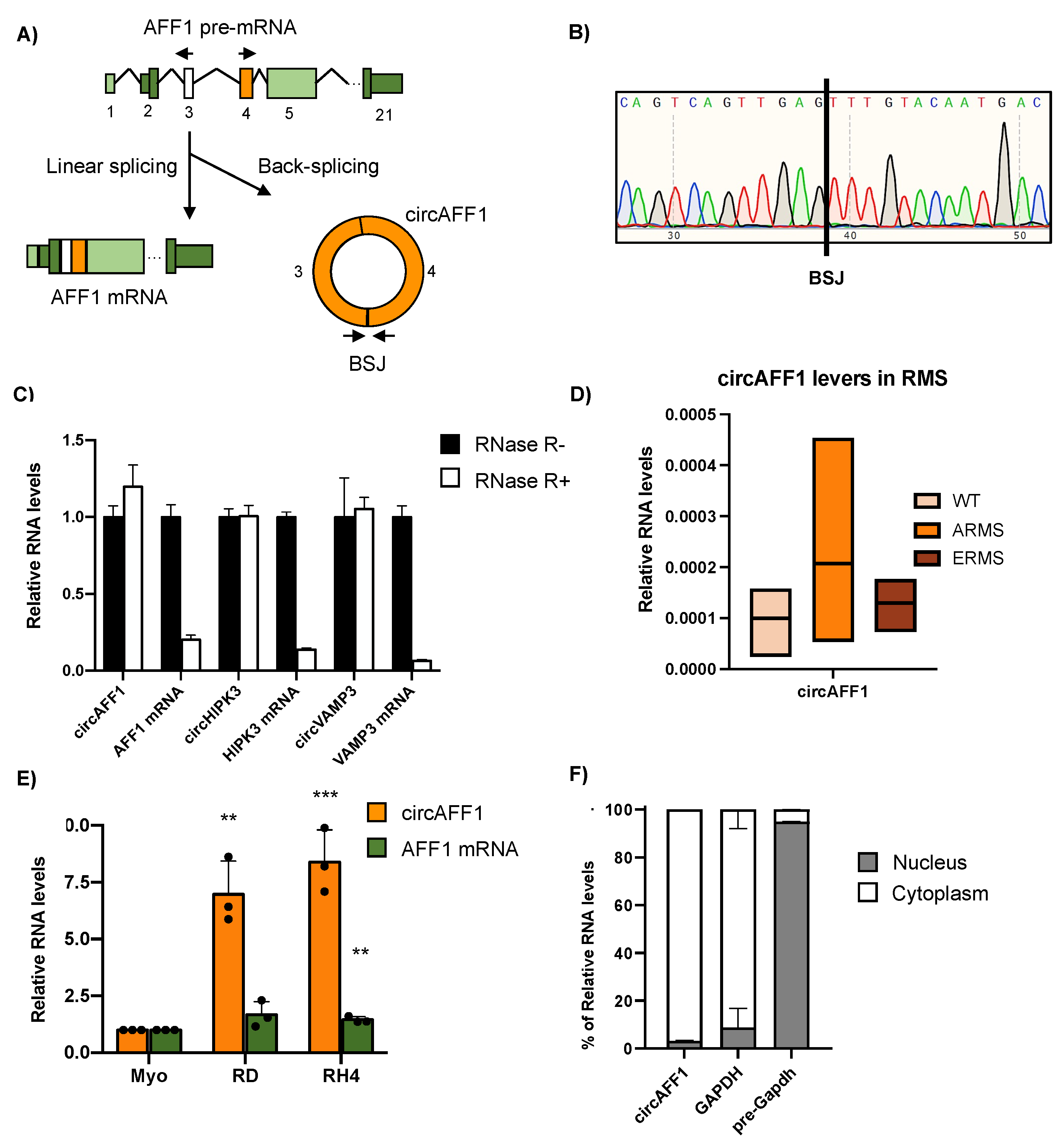{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures, Transfections and Flow Cytometric Analysis of Cell Cycle
2.2. RNA Purification and RNase-R Digestion
2.3. Patient Biopsies
2.4. Subcellular Fractionation
2.5. Growth Curve Assay
2.6. Sucrose Gradient Fractionation
2.7. RNA Sequencing and Bioinformatic Analyses
2.8. Migration and Wound Healing Experiments
2.9. Western Blot
3. Results
3.1. CircAFF1 Is Abundantly Expressed in RMS-Derived Cell Lines
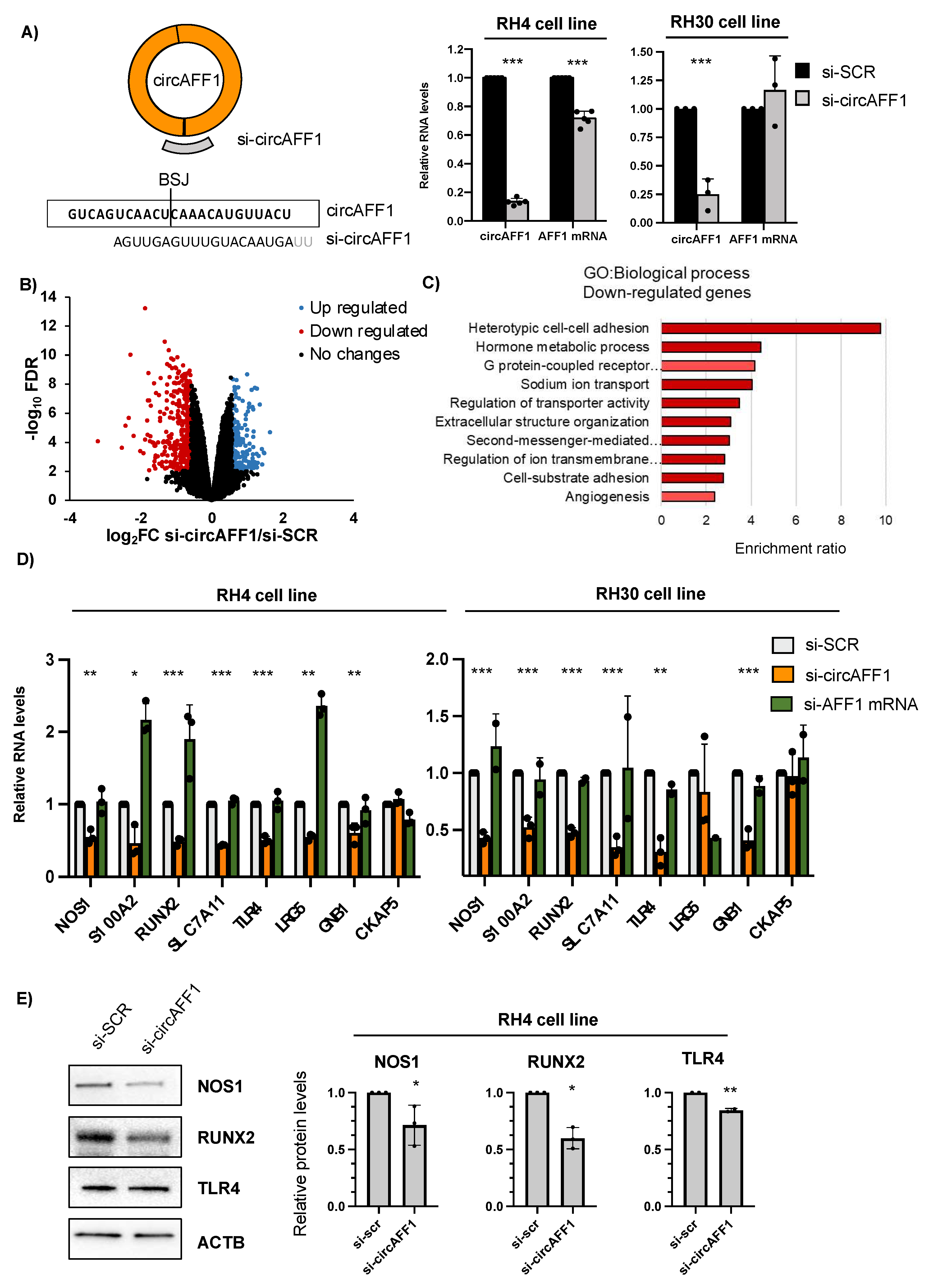
3.2. CircAFF1 Knockdown Affects Adhesion-Related Pathways in ARMS Cells
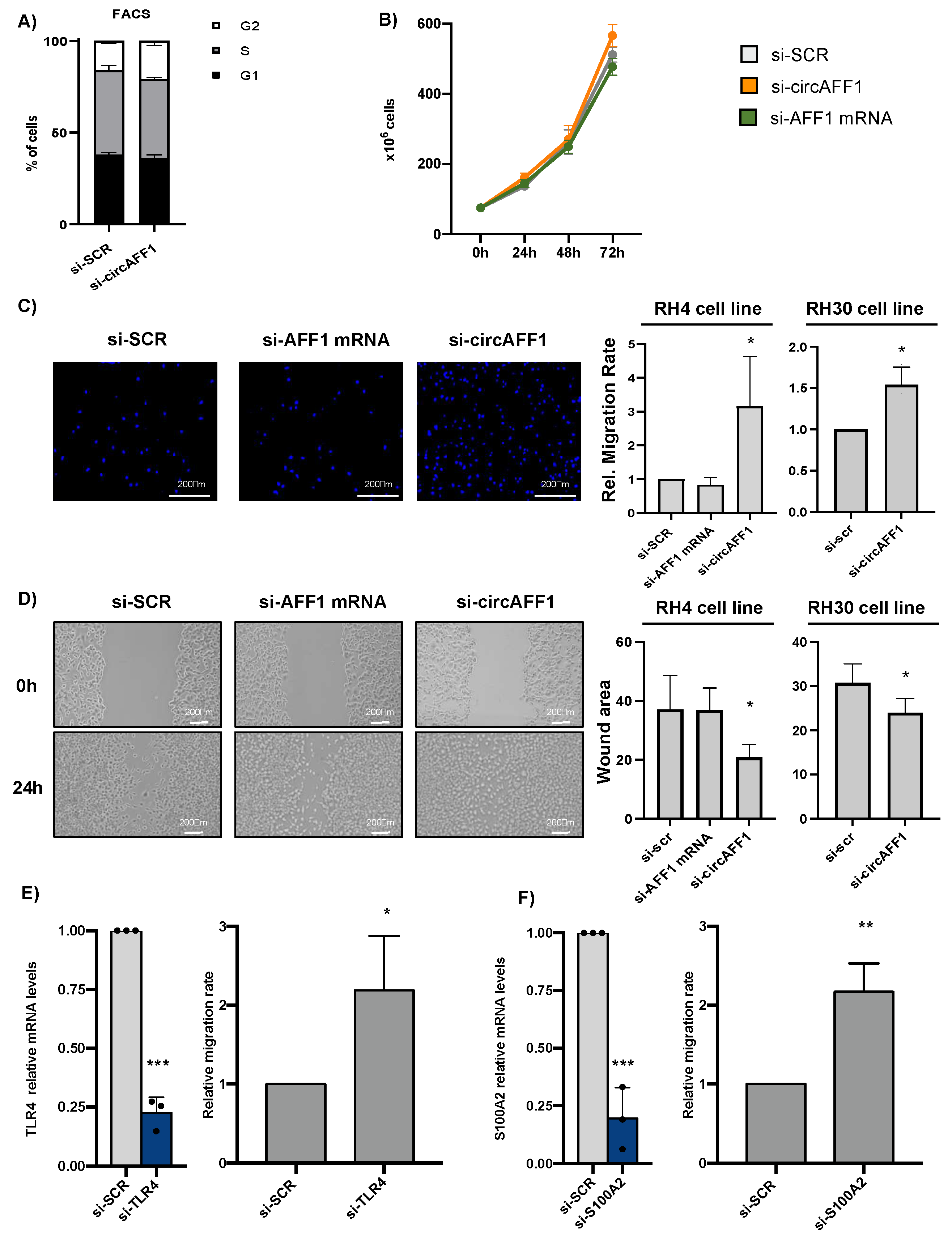
3.3. CircAFF1 Inhibits RH4 Cells Migration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Starke, S.; Jost, I.; Rossbach, O.; Schneider, T.; Schreiner, S.; Hung, L.H.; Bindereif, A. Exon Circularization Requires Canonical Splice Signals. Cell Rep. 2015, 10, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.L.; Bao, Y.; Yee, M.C.; Barrett, S.P.; Hogan, G.J.; Olsen, M.N.; Dinneny, J.R.; Brown, P.O.; Salzman, J. Circular RNA Is Expressed across the Eukaryotic Tree of Life. PLoS ONE 2014, 9, 90859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; Herzog, M.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2014, 58, 870–885. [Google Scholar] [CrossRef] [Green Version]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs Are the Predominant Transcript Isoform from Hundreds of Human Genes in Diverse Cell Types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA Circles Function as Efficient MicroRNA Sponges. Nature 2013, 495, 11993. [Google Scholar] [CrossRef]
- Yu, H.; Chen, Y.; Jiang, P. Circular RNA HIPK3 Exerts Oncogenic Properties through Suppression of MiR-124 in Lung Cancer. Biochem. Biophys. Res. Commun. 2018, 506, 455–462. [Google Scholar] [CrossRef]
- Panda, A.C. Circular RNAs Act as MiRNA Sponges. Adv. Exp. Med. Biol. 2018, 1087, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. CircRNA Biogenesis Competes with Pre-MRNA Splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Holdt, L.M.; Stahringer, A.; Sass, K.; Pichler, G.; Kulak, N.A.; Wilfert, W.; Kohlmaier, A.; Herbst, A.; Northoff, B.H.; Nicolaou, A.; et al. Circular Non-Coding RNA ANRIL Modulates Ribosomal RNA Maturation and Atherosclerosis in Humans. Nat. Commun. 2016, 7, 12429. [Google Scholar] [CrossRef] [Green Version]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 Circular RNA Retards Cell Cycle Progression via Forming Ternary Complexes with P21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [Green Version]
- Begum, S.; Yiu, A.; Stebbing, J.; Castellano, L. Novel Tumour Suppressive Protein Encoded by Circular RNA, Circ-SHPRH, in Glioblastomas. Oncogene 2018, 37, 4055–4057. [Google Scholar] [CrossRef]
- Legnini, I.; di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA That Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37.e9. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.-C.; Wong, C.-W.; Liang, P.-P.; Shi, M.; Cao, Y.; Rao, S.-T.; Tsui, S.K.-W.; Waye, M.M.-Y.; Zhang, Q.; Fu, W.-M.; et al. Translation of the Circular RNA Circβ-Catenin Promotes Liver Cancer Cell Growth through Activation of the Wnt Pathway. Genome Biol. 2019, 20, 84. [Google Scholar] [CrossRef]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21.e7. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.; Beltran, M.; Damizia, M.; Grelloni, C.; Colantoni, A.; Setti, A.; Di Timoteo, G.; Dattilo, D.; Centrón-Broco, A.; Nicoletti, C.; et al. Circular RNA ZNF609/CKAP5 MRNA Interaction Regulates Microtubule Dynamics and Tumorigenicity. Mol. Cell 2022, 82, 75–89.e9. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-Intron Circular RNAs Regulate Transcription in the Nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Bach, D.H.; Lee, S.K.; Sood, A.K. Circular RNAs in Cancer. Mol. Ther. Nucleic Acids 2019, 16, 118–129. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Wang, Y.; Fan, Y.; Fang, N.; Wang, T.; Xu, T.; Shu, Y. CircRNAs in Cancer Metabolism: A Review. J. Hematol. Oncol. 2019, 12, 90. [Google Scholar] [CrossRef] [Green Version]
- Guarnerio, J.; Bezzi, M.; Jeong, J.C.; Paffenholz, S.V.; Berry, K.; Naldini, M.M.; Lo-Coco, F.; Tay, Y.; Beck, A.H.; Pandolfi, P.P. Oncogenic Role of Fusion-CircRNAs Derived from Cancer-Associated Chromosomal Translocations. Cell 2016, 165, 289–302. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.L.; Wang, Y.; et al. Extensive Translation of Circular RNAs Driven by N 6 -Methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [Green Version]
- di Timoteo, G.; Dattilo, D.; Centrón-Broco, A.; Colantoni, A.; Guarnacci, M.; Rossi, F.; Incarnato, D.; Oliviero, S.; Fatica, A.; Morlando, M.; et al. Modulation of CircRNA Metabolism by M6A Modification. Cell Rep. 2020, 31, 107641. [Google Scholar] [CrossRef]
- Huh, W.; Egas Bejar, D. Rhabdomyosarcoma in Adolescent and Young Adult Patients: Current Perspectives. Adolesc. Health Med. Ther. 2014, 5, 115. [Google Scholar] [CrossRef] [Green Version]
- Soleimani, V.D.; Rudnicki, M.A. New Insights into the Origin and the Genetic Basis of Rhabdomyosarcomas. Cancer Cell 2011, 19, 157–159. [Google Scholar] [CrossRef] [Green Version]
- Kashi, V.P.; Hatley, M.E.; Galindo, R.L. Probing for a Deeper Understanding of Rhabdomyosarcoma: Insights from Complementary Model Systems. Nat. Rev. Cancer 2015, 15, 426–439. [Google Scholar] [CrossRef] [Green Version]
- Skapek, S.X.; Ferrari, A.; Gupta, A.A.; Lupo, P.J.; Butler, E.; Shipley, J.; Barr, F.G.; Hawkins, D.S. Rhabdomyosarcoma. Nat. Rev. Dis. Prim. 2019, 5, 1. [Google Scholar] [CrossRef]
- Hinson, A.R.P.; Jones, R.; Lisa, L.E.; Belyea, B.C.; Barr, F.G.; Linardic, C.M. Human Rhabdomyosarcoma Cell Lines for Rhabdomyosarcoma Research: Utility and Pitfalls. Front. Oncol. 2013, 3, 183. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.; Centrón-Broco, A.; Dattilo, D.; di Timoteo, G.; Guarnacci, M.; Colantoni, A.; Beltran Nebot, M.; Bozzoni, I. CircVAMP3: A CircRNA with a Role in Alveolar Rhabdomyosarcoma Cell Cycle Progression. Genes 2021, 12, 985. [Google Scholar] [CrossRef]
- Megiorni, F.; Camero, S.; Ceccarelli, S.; Mcdowell, H.P.; Mannarino, O.; Marampon, F.; Pizer, B.; Shukla, R.; Marchese, C.; Clerico, A.; et al. DNMT3B in Vitro Knocking-down Is Able to Reverse Embryonal Rhabdomyosarcoma Cell Phenotype through Inhibition of Proliferation and Induction of Myogenic Differentiation. Oncotarget 2016, 7, 79342–79356. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.C.; Xiong, Q.C.; Zhu, X.X.; Du, W.; Deng, P.; Li, X.B.; Jiang, Y.Z.; Zou, S.J.; Wang, C.Y.; Yuan, Q. AFF1 and AFF4 Differentially Regulate the Osteogenic Differentiation of Human MSCs. Bone Res. 2017, 5, 17044. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Nakamura, T.; Alder, H.; Prasad, R.; Canaani, O.; Cimino, G.; Croce, C.M.; Canaani, E. The t(4;11) Chromosome Translocation of Human Acute Leukemias Fuses the ALL-1 Gene, Related to Drosophila Trithorax, to the AF-4 Gene. Cell 1992, 71, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Zhao, Y.; Zhou, Z.; Li, W.; Li, B.; Gu, G.; Luo, Y.; Shuai, X.; Fan, C.; Wu, L.; et al. Identifying Circular RNAs in HepG2 Expressing Genotype IV Swine Hepatitis E Virus ORF3 Via Whole Genome Sequencing. Cell Transplant. 2021, 30, 09636897211055042. [Google Scholar] [CrossRef] [PubMed]
- Glažar, P.; Papavasileiou, P.; Rajewsky, N. CircBase: A Database for Circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Yan, H.; Wang, C.; Li, M.; Lv, X.; Wu, H.; Fang, Z.; Mo, D.; Zhang, Z.; Liang, B.; et al. CircAFF1 Aggravates Vascular Endothelial Cell Dysfunction Mediated by MiR-516b/SAV1/YAP1 Axis. Front. Physiol. 2020, 11, 547434. [Google Scholar] [CrossRef]
- Dattilo, D.; Di Timoteo, G.; Setti, A.; Giuliani, A.; Peruzzi, G.; Beltran Nebot, M.; Centrón-Broco, A.; Mariani, D.; Mozzetta, C.; Bozzoni, I. The M6A Reader YTHDC1 and the RNA Helicase DDX5 Control the Production of Rhabdomyosarcoma-Enriched CircRNAs. Nat. Commun. 2023, 14, 1898. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sharpless, N.E. Detecting and Characterizing Circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef]
- Rossi, F.; Legnini, I.; Megiorni, F.; Colantoni, A.; Santini, T.; Morlando, M.; Di Timoteo, G.; Dattilo, D.; Dominici, C.; Bozzoni, I. Circ-ZNF609 Regulates G1-S Progression in Rhabdomyosarcoma. Oncogene 2019, 38, 3843–3854. [Google Scholar] [CrossRef] [Green Version]
- Bersani, F.; Lingua, M.F.; Morena, D.; Foglizzo, V.; Miretti, S.; Lanzetti, L.; Carrà, G.; Morotti, A.; Ala, U.; Provero, P.; et al. Deep Sequencing Reveals a Novel MiR-22 Regulatory Network with Therapeutic Potential in Rhabdomyosarcoma. Cancer Res. 2016, 76, 6095–6106. [Google Scholar] [CrossRef] [Green Version]
- Yahiro, K.; Matsumoto, Y.; Yamada, H.; Endo, M.; Setsu, N.; Fujiwara, T.; Nakagawa, M.; Kimura, A.; Shimada, E.; Okada, S.; et al. Activation of TLR4 Signaling Inhibits Progression of Osteosarcoma by Stimulating CD8-Positive Cytotoxic Lymphocytes. Cancer Immunol. Immunother. 2020, 69, 745–758. [Google Scholar] [CrossRef]
- Nagy, N.; Brenner, C.; Markadieu, N.; Chaboteaux, C.; Camby, I.; Schäfer, B.W.; Pochet, R.; Heizmann, C.W.; Salmon, I.; Kiss, R.; et al. S100A2, a Putative Tumor Suppressor Gene, Regulates In Vitro Squamous Cell Carcinoma Migration. Lab. Investig. 2001, 81, 599–612. [Google Scholar] [CrossRef] [Green Version]
- Tu, C.; Du, T.; Shao, C.; Liu, Z.; Li, L.; Shen, Y. Evaluating the Potential of Housekeeping Genes, RRNAs, SnRNAs, MicroRNAs and CircRNAs as Reference Genes for the Estimation of PMI. Forensic Sci. Med. Pathol. 2018, 14, 194–201. [Google Scholar] [CrossRef]
- Yin, M.; Chen, W.; Li, M.; Wang, K.; Hu, N.; Li, Z. CircAFF1 Enhances Intracerebral Hemorrhage Induced Neuronal Ferroptosis by Targeting MiR-140–5p to Regulate GSK-3β Mediated Wnt/β-Catenin Signal Pathway. Brain Res. Bull. 2022, 189, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Zhu, X.; Li, D.; Yang, F.; Meng, J.; Jiang, Y.; Ma, J.; Lu, Y. The Differential Expression of Circular RNAs and the Role of CircAFF1 in Lens Epithelial Cells of High-Myopic Cataract. J. Clin. Med. 2023, 12, 813. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekoninck, S.; Blanpain, C. Stem Cell Dynamics, Migration and Plasticity during Wound Healing. Nat. Cell Biol. 2019, 21, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, D.W.; Dinger, M.E. Endogenous MicroRNA Sponges: Evidence and Controversy. Nat. Rev. Genet. 2016, 17, 272–283. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Centrón-Broco, A.; Rossi, F.; Grelloni, C.; Garraffo, R.; Dattilo, D.; Giuliani, A.; Di Timoteo, G.; Colantoni, A.; Bozzoni, I.; Beltran Nebot, M. CircAFF1 Is a Circular RNA with a Role in Alveolar Rhabdomyosarcoma Cell Migration. Biomedicines 2023, 11, 1893. https://doi.org/10.3390/biomedicines11071893
Centrón-Broco A, Rossi F, Grelloni C, Garraffo R, Dattilo D, Giuliani A, Di Timoteo G, Colantoni A, Bozzoni I, Beltran Nebot M. CircAFF1 Is a Circular RNA with a Role in Alveolar Rhabdomyosarcoma Cell Migration. Biomedicines. 2023; 11(7):1893. https://doi.org/10.3390/biomedicines11071893
Chicago/Turabian StyleCentrón-Broco, Alvaro, Francesca Rossi, Chiara Grelloni, Raffaele Garraffo, Dario Dattilo, Andrea Giuliani, Gaia Di Timoteo, Alessio Colantoni, Irene Bozzoni, and Manuel Beltran Nebot. 2023. "CircAFF1 Is a Circular RNA with a Role in Alveolar Rhabdomyosarcoma Cell Migration" Biomedicines 11, no. 7: 1893. https://doi.org/10.3390/biomedicines11071893