
The Role of the Microbiome in the Pathogenesis and Treatment of Asthma
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Human Microbiome
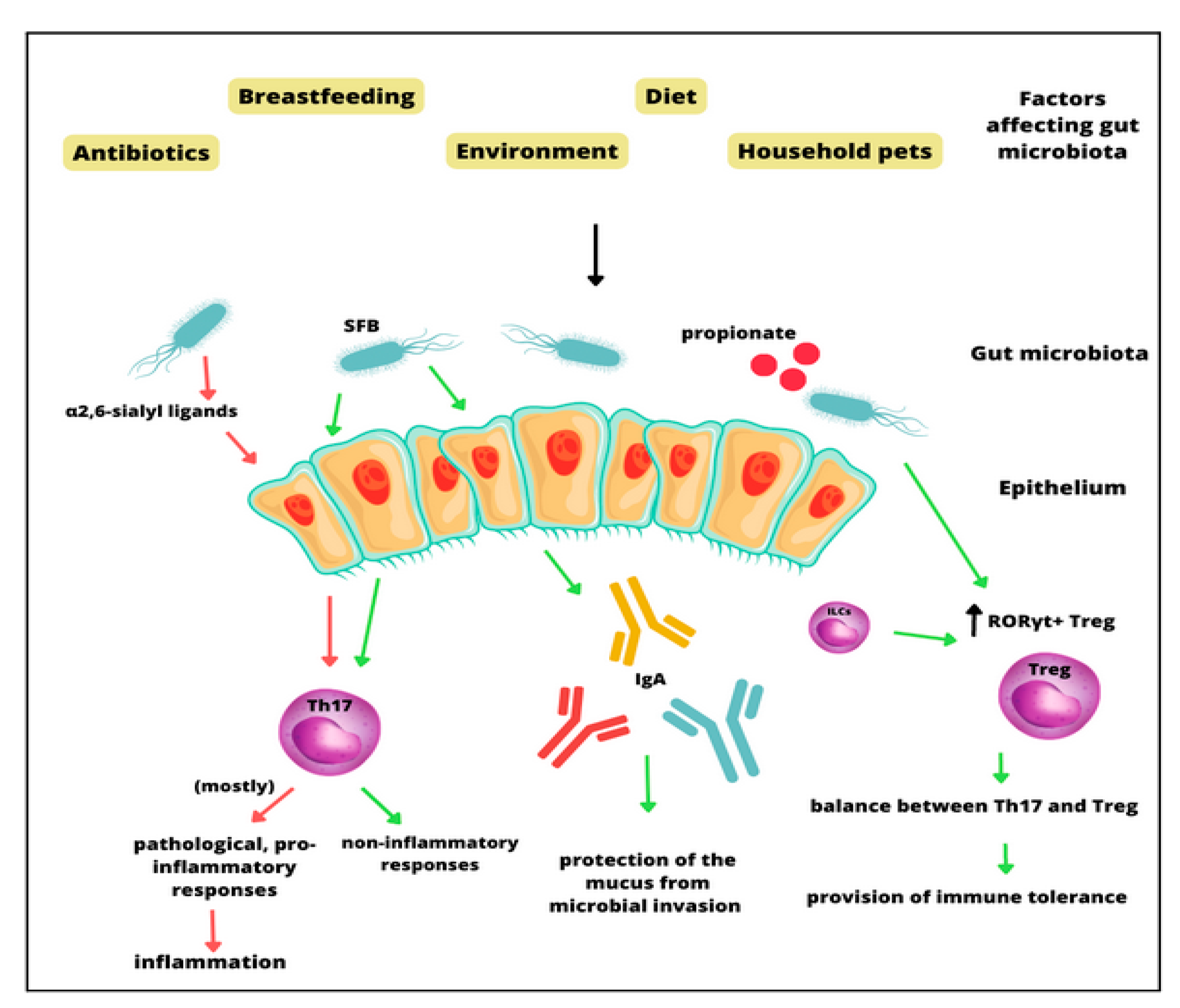
3. The Molecular Role of the Gut Microbiota in Asthma Pathogenesis
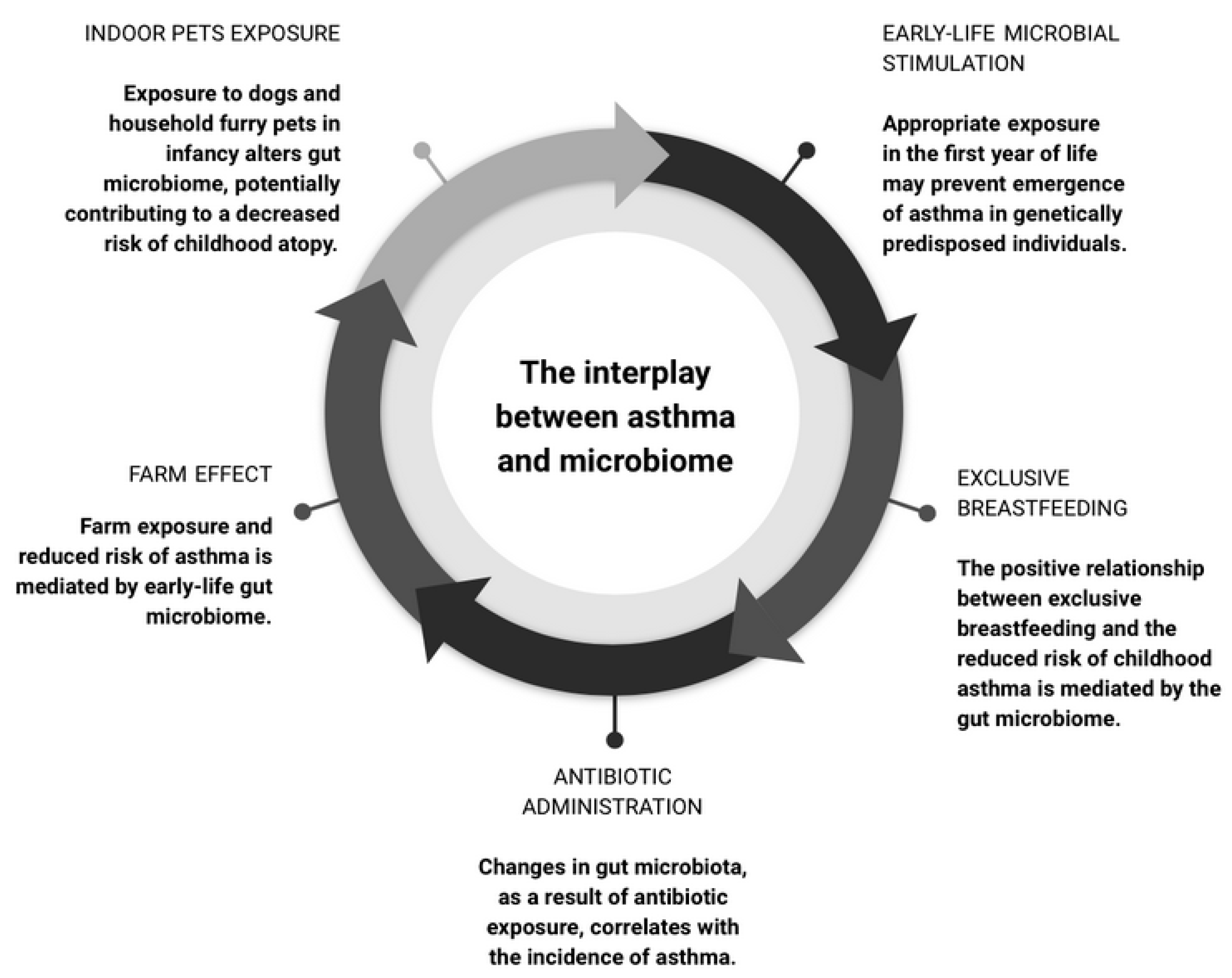
4. The Microbiome as a Moderator in Asthma Development
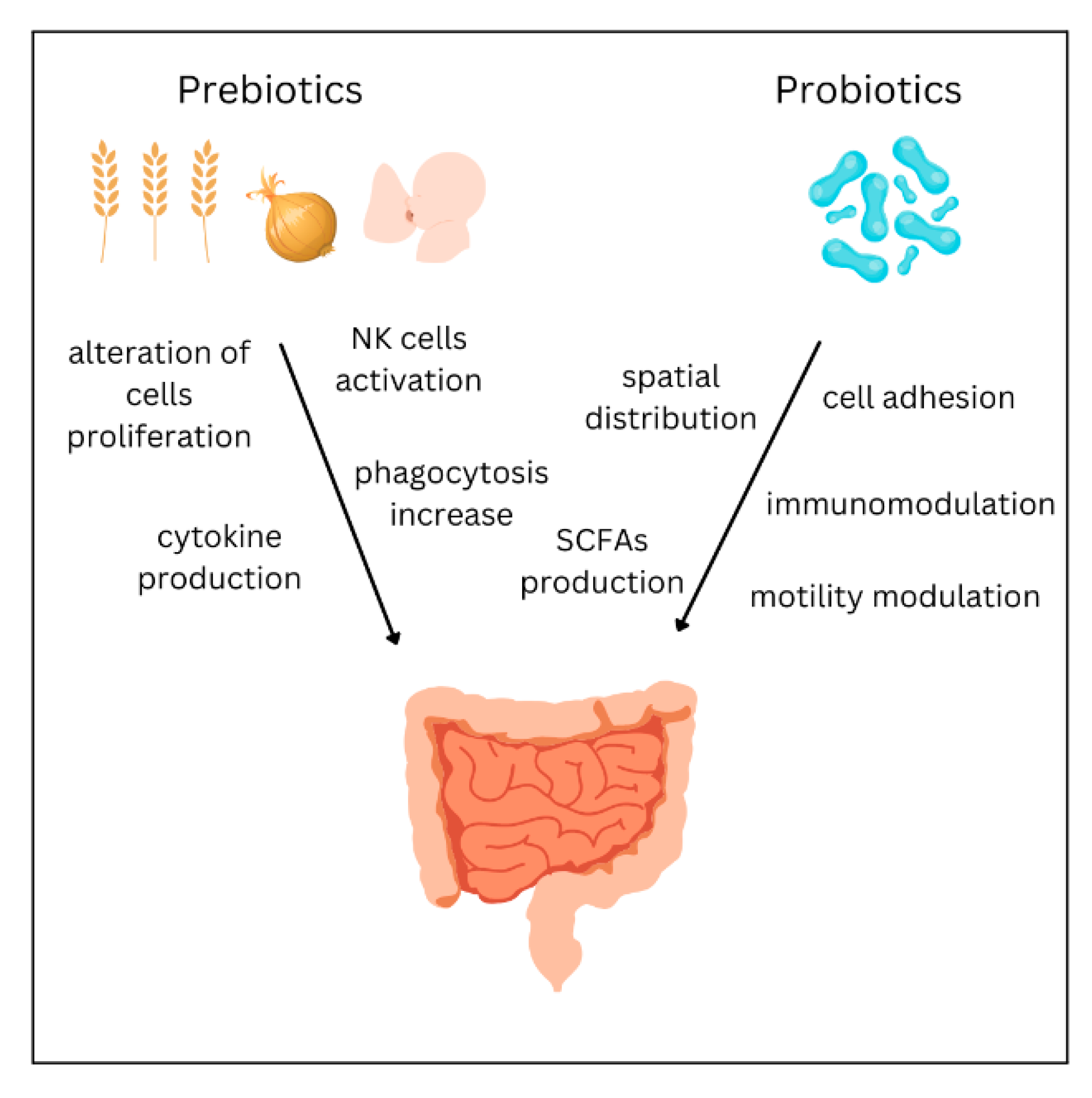
5. Treatment
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Porsbjerg, C.; Melén, E.; Lehtimäki, L.; Shaw, D. Asthma. Lancet 2023, 401, 858–873. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. The Basic Immunology of Asthma. Cell 2021, 184, 1469–1485. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.; Pier, J.; Litonjua, A.A. Asthma Epidemiology and Risk Factors. Semin. Immunopathol. 2020, 42, 5–15. [Google Scholar] [CrossRef]
- Gans, M.D.; Gavrilova, T. Understanding the Immunology of Asthma: Pathophysiology, Biomarkers, and Treatments for Asthma Endotypes. Paediatr. Respir. Rev. 2020, 36, 118–127. [Google Scholar] [CrossRef]
- Jeong, J.; Lee, H.K. The Role of CD4+ T Cells and Microbiota in the Pathogenesis of Asthma. Int. J. Mol. Sci. 2021, 22, 11822. [Google Scholar] [CrossRef]
- Rogliani, P.; Ora, J.; Calzetta, L.; Matera, M.G.; Cazzola, M. Asthma and Comorbidities: Recent Advances. Polish Arch. Intern. Med. 2022, 132, 16250. [Google Scholar] [CrossRef]
- Lommatzsch, M.; Buhl, R.; Korn, S. The Treatment of Mild and Moderate Asthma in Adults. Dtsch. Arztebl. Int. 2020, 117, 434–444. [Google Scholar] [CrossRef]
- De Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut Microbiome and Health: Mechanistic Insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human Gut Microbiota/Microbiome in Health and Diseases: A Review. Antonie Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Manos, J. The Human Microbiome in Disease and Pathology. Apmis 2022, 130, 690. [Google Scholar] [CrossRef]
- Welch, C.B.; Ryman, V.E.; Pringle, T.D.; Lourenco, J.M. Utilizing the Gastrointestinal Microbiota to Modulate Cattle Health through the Microbiome-Gut-Organ Axes. Microorganisms 2022, 10, 1391. [Google Scholar] [CrossRef]
- Brown, E.G.; Tanner, C.M.; Goldman, S.M. The Microbiome in Neurodegenerative Disease. Curr. Geriatr. Rep. 2018, 7, 81–91. [Google Scholar] [CrossRef]
- Ticinesi, A.; Tana, C.; Nouvenne, A.; Prati, B.; Lauretani, F.; Meschi, T. Gut Microbiota, Cognitive Frailty and Dementia in Older Individuals: A Systematic Review. Clin. Interv. Aging 2018, 13, 1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Munckhof, I.C.L.; Kurilshikov, A.; ter Horst, R.; Riksen, N.P.; Joosten, L.A.B.; Zhernakova, A.; Fu, J.; Keating, S.T.; Netea, M.G.; de Graaf, J.; et al. Role of Gut Microbiota in Chronic Low-Grade Inflammation as Potential Driver for Atherosclerotic Cardiovascular Disease: A Systematic Review of Human Studies. Obes. Rev. 2018, 19, 1719–1734. [Google Scholar] [CrossRef] [Green Version]
- Barcik, W.; Boutin, R.C.T.; Sokolowska, M.; Finlay, B.B. The Role of Lung and Gut Microbiota in the Pathology of Asthma. Immunity 2020, 52, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, J.R.; Ravel, J. The Vocabulary of Microbiome Research: A Proposal. Microbiome 2015, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- Barko, P.C.; McMichael, M.A.; Swanson, K.S.; Williams, D.A. The Gastrointestinal Microbiome: A Review. J. Vet. Intern. Med. 2018, 32, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Bezirtzoglou, E. The Intestinal Microflora During the First Weeks of Life. Anaerobe 1997, 3, 173–177. [Google Scholar] [CrossRef]
- Palmer, C.; Bik, E.M.; DiGiulio, D.B.; Relman, D.A.; Brown, P.O. Development of the Human Infant Intestinal Microbiota. PLoS Biol. 2007, 5, 1556–1573. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Bello, M.G.; De Jesus-Laboy, K.M.; Shen, N.; Cox, L.M.; Amir, A.; Gonzalez, A.; Bokulich, N.A.; Song, S.J.; Hoashi, M.; Rivera-Vinas, J.I.; et al. Partial Restoration of the Microbiota of Cesarean-Born Infants via Vaginal Microbial Transfer. Nat. Med. 2016, 22, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Biasucci, G.; Rubini, M.; Riboni, S.; Morelli, L.; Bessi, E.; Retetangos, C. Mode of Delivery Affects the Bacterial Community in the Newborn Gut. Early Hum. Dev. 2010, 86 (Suppl. S1), 13–15. [Google Scholar] [CrossRef] [PubMed]
- Martín-Peláez, S.; Cano-Ibáñez, N.; Pinto-Gallardo, M.; Amezcua-Prieto, C. The Impact of Probiotics, Prebiotics, and Synbiotics during Pregnancy or Lactation on the Intestinal Microbiota of Children Born by Cesarean Section: A Systematic Review. Nutrients 2022, 14, 341. [Google Scholar] [CrossRef] [PubMed]
- Mohajeri, M.H.; Brummer, R.J.M.; Rastall, R.A.; Weersma, R.K.; Harmsen, H.J.M.; Faas, M.; Eggersdorfer, M. The Role of the Microbiome for Human Health: From Basic Science to Clinical Applications. Eur. J. Nutr. 2018, 57, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Gritz, E.C.; Bhandari, V. The Human Neonatal Gut Microbiome: A Brief Review. Front. Pediatr. 2015, 3, 17. [Google Scholar]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the Human Intestinal Microbial Flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Guarner, F.; Malagelada, J.R. Gut Flora in Health and Disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Cui, L.; Morris, A.; Ghedin, E. The Human Mycobiome in Health and Disease. Genome Med. 2013, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Scarpellini, E.; Ianiro, G.; Attili, F.; Bassanelli, C.; De Santis, A.; Gasbarrini, A. The Human Gut Microbiota and Virome: Potential Therapeutic Implications. Dig. Liver Dis. 2015, 47, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.M.; Wells, J.M. Regulation of Human Epithelial Tight Junction Proteins by Lactobacillus Plantarum in Vivo and Protective Effects on the Epithelial Barrier. Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 298, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Ohno, H. Intestinal M Cells. J. Biochem. 2016, 159, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Štěpánková, R.; Kovářů, F.; Kruml, J. Lymphatic Tissue of the Intestinal Tract of Germfree and Conventional Rabbits. Folia Microbiol. 1980, 25, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, S.A.; McGinniss, J.E.; Collman, R.G. The Lung Microbiome: Progress and Promise. J. Clin. Investig. 2021, 131, e150473. [Google Scholar] [CrossRef]
- Charlson, E.S.; Bittinger, K.; Haas, A.R.; Fitzgerald, A.S.; Frank, I.; Yadav, A.; Bushman, F.D.; Collman, R.G. Topographical Continuity of Bacterial Populations in the Healthy Human Respiratory Tract. Am. J. Respir. Crit. Care Med. 2011, 184, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Gleeson, K.; Eggli, D.F.; Maxwell, S.L. Quantitative Aspiration during Sleep in Normal Subjects. Chest 1997, 111, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Yu, Y.; Du, W.; Liu, Y.; Dai, R.; Tang, W.; Wang, P.; Zhang, C.; Shi, G. Fungal and Bacterial Microbiome Dysbiosis and Imbalance of Trans-Kingdom Network in Asthma. Clin. Transl. Allergy 2020, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Jeong, K.I.; Itoh, K.; Doi, K. Regional Variations in the Distributions of Small Intestinal Intraepithelial Lymphocytes in Germ-Free and Specific Pathogen-Free Mice. Exp. Mol. Pathol. 2002, 72, 230–235. [Google Scholar] [CrossRef]
- Campbell, C.; Kandalgaonkar, M.R.; Golonka, R.M.; Yeoh, B.S.; Vijay-Kumar, M.; Saha, P. Crosstalk between Gut Microbiota and Host Immunity: Impact on Inflammation and Immunotherapy. Biomedicines 2023, 11, 294. [Google Scholar] [CrossRef]
- Irons, E.E.; Cortes Gomez, E.; Andersen, V.L.; Lau, J.T.Y. Bacterial Colonization and TH17 Immunity Are Shaped by Intestinal Sialylation in Neonatal Mice. Glycobiology 2022, 32, 414–428. [Google Scholar] [CrossRef]
- Lyu, M.; Suzuki, H.; Kang, L.; Gaspal, F.; Zhou, W.; Goc, J.; Zhou, L.; Zhou, J.; Zhang, W.; Artis, D.; et al. ILC3s Select Microbiota-Specific Regulatory T Cells to Establish Tolerance in the Gut. Nature 2022, 610, 744–751. [Google Scholar] [CrossRef]
- Su, X.; Yin, X.; Liu, Y.; Yan, X.; Zhang, S.; Wang, X.; Lin, Z.; Zhou, X.; Gao, J.; Wang, Z.; et al. Gut Dysbiosis Contributes to the Imbalance of Treg and Th17 Cells in Graves’ Disease Patients by Propionic Acid. J. Clin. Endocrinol. Metab. 2020, 105, 3526–3547. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Ohno, H. Reciprocal Regulation of IgA and the Gut Microbiota: A Key Mutualism in the Intestine. Int. Immunol. 2021, 33, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Wolf, N.; Lavelle, E.C. Innate Immune Receptors. Methods Mol. Biol. 2016, 1417, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.B. Early Innate Immunity to Bacterial Infection in the Lung Is Regulated Systemically by the Commensal Microbiota via Nod-Like Receptor Ligands. Infect. Immun. 2014, 82, 4596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, C.; Yang, Y.; Zhu, W. Crosstalk Between The Immune Receptors and Gut Microbiota. Curr. Protein Pept. Sci. 2015, 16, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Guryanova, S.V.; Gigani, O.B.; Gudima, G.O.; Kataeva, A.M.; Kolesnikova, N.V. Dual Effect of Low-Molecular-Weight Bioregulators of Bacterial Origin in Experimental Model of Asthma. Life 2022, 12, 192. [Google Scholar] [CrossRef]
- Gu, B.H.; Rim, C.Y.; Lee, S.; Kim, T.Y.; Joo, S.S.; Lee, S.J.; Park, H.K.; Kim, M. Alteration of Gut Immunity and Microbiome in Mixed Granulocytic Asthma. Biomedicines 2022, 10, 2946. [Google Scholar] [CrossRef]
- Abdel-Aziz, M.I.; Vijverberg, S.J.H.; Neerincx, A.H.; Kraneveld, A.D.; Maitland-van der Zee, A.H. The Crosstalk between Microbiome and Asthma: Exploring Associations and Challenges. Clin. Exp. Allergy 2019, 49, 1067–1086. [Google Scholar] [CrossRef] [Green Version]
- Mandaliya, D.K.; Patel, S.; Seshadri, S. The Combinatorial Effect of Acetate and Propionate on High-Fat Diet Induced Diabetic Inflammation or Metaflammation and T Cell Polarization. Inflammation 2021, 44, 68–79. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Adivitiya; Kaushik, M.S.; Chakraborty, S.; Veleri, S.; Kateriya, S. Mucociliary Respiratory Epithelium Integrity in Molecular Defense and Susceptibility to Pulmonary Viral Infections. Biology 2021, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Strachan, D.P. Hay Fever, Hygiene, and Household Size. Br. Med. J. 1989, 299, 1259–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haspeslagh, E.; Heyndrickx, I.; Hammad, H.; Lambrecht, B.N. The Hygiene Hypothesis: Immunological Mechanisms of Airway Tolerance. Curr. Opin. Immunol. 2018, 54, 102. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A.W. 99th Dahlem Conference on Infection, Inflammation and Chronic Inflammatory Disorders: Darwinian Medicine and the ‘Hygiene’ or ‘Old Friends’ Hypothesis. Clin. Exp. Immunol. 2010, 160, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Stokholm, J.; Blaser, M.J.; Thorsen, J.; Rasmussen, M.A.; Waage, J.; Vinding, R.K.; Schoos, A.M.M.; Kunøe, A.; Fink, N.R.; Chawes, B.L.; et al. Maturation of the Gut Microbiome and Risk of Asthma in Childhood. Nat. Commun. 2018, 9, 141. [Google Scholar] [CrossRef] [Green Version]
- Christensen, E.D.; Hjelmsø, M.H.; Thorsen, J.; Shah, S.; Redgwell, T.; Poulsen, C.E.; Trivedi, U.; Russel, J.; Gupta, S.; Chawes, B.L.; et al. The Developing Airway and Gut Microbiota in Early Life Is Influenced by Age of Older Siblings. Microbiome 2022, 10, 106. [Google Scholar] [CrossRef]
- Thorsen, J.; Rasmussen, M.A.; Waage, J.; Mortensen, M.; Brejnrod, A.; Bønnelykke, K.; Chawes, B.L.; Brix, S.; Sørensen, S.J.; Stokholm, J.; et al. Infant Airway Microbiota and Topical Immune Perturbations in the Origins of Childhood Asthma. Nat. Commun. 2019, 10, 5001. [Google Scholar] [CrossRef] [Green Version]
- Rosas-Salazar, C.; Shilts, M.H.; Tang, Z.Z.; Hong, Q.; Turi, K.N.; Snyder, B.M.; Wiggins, D.A.; Lynch, C.E.; Gebretsadik, T.; Peebles, R.S.; et al. Exclusive Breast-Feeding, the Early-Life Microbiome and Immune Response, and Common Childhood Respiratory Illnesses. J. Allergy Clin. Immunol. 2022, 150, 612–621. [Google Scholar] [CrossRef]
- Biagi, E.; Quercia, S.; Aceti, A.; Beghetti, I.; Rampelli, S.; Turroni, S.; Faldella, G.; Candela, M.; Brigidi, P.; Corvaglia, L. The Bacterial Ecosystem of Mother’s Milk and Infant’s Mouth and Gut. Front. Microbiol. 2017, 8, 1214. [Google Scholar] [CrossRef] [Green Version]
- Laursen, M.F.; Sakanaka, M.; von Burg, N.; Mörbe, U.; Andersen, D.; Moll, J.M.; Pekmez, C.T.; Rivollier, A.; Michaelsen, K.F.; Mølgaard, C.; et al. Bifidobacterium Species Associated with Breastfeeding Produce Aromatic Lactic Acids in the Infant Gut. Nat. Microbiol. 2021, 6, 1367–1382. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Stiemsma, L.T.; Dimitriu, P.A.; Thorson, L.; Russell, S.; Yurist-Doutsch, S.; Kuzeljevic, B.; Gold, M.J.; Britton, H.M.; Lefebvre, D.L.; et al. Early Infancy Microbial and Metabolic Alterations Affect Risk of Childhood Asthma. Sci. Transl. Med. 2015, 7, 307ra152. [Google Scholar] [CrossRef] [PubMed]
- Stokholm, J.; Thorsen, J.; Blaser, M.J.; Rasmussen, M.A.; Hjelmsø, M.; Shah, S.; Christensen, E.D.; Chawes, B.L.; Bønnelykke, K.; Brix, S.; et al. Delivery Mode and Gut Microbial Changes Correlate with an Increased Risk of Childhood Asthma. Sci. Transl. Med. 2020, 12, eaax9929. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Friedman, H.; Boyd, B.C.; McGurn, A.; Babinski, P.; Markossian, T.; Dugas, L.R. Early Antibiotic Exposure and Development of Asthma and Allergic Rhinitis in Childhood. BMC Pediatr. 2019, 19, 225. [Google Scholar] [CrossRef] [PubMed]
- Patrick, D.M.; Sbihi, H.; Dai, D.L.Y.; Al Mamun, A.; Rasali, D.; Rose, C.; Marra, F.; Boutin, R.C.T.; Petersen, C.; Stiemsma, L.T.; et al. Decreasing Antibiotic Use, the Gut Microbiota, and Asthma Incidence in Children: Evidence from Population-Based and Prospective Cohort Studies. Lancet Respir. Med. 2020, 8, 1094–1105. [Google Scholar] [CrossRef]
- Von Ehrenstein, O.S.; Von Mutius, E.; Illi, S.; Baumann, L.; Böhm, O.; Von Kries, R. Reduced Risk of Hay Fever and Asthma among Children of Farmers. Clin. Exp. Allergy 2000, 30, 187–193. [Google Scholar] [CrossRef]
- Von Mutius, E.; Vercelli, D. Farm Living: Effects on Childhood Asthma and Allergy. Nat. Rev. Immunol. 2010, 10, 861–868. [Google Scholar] [CrossRef]
- Ege, M.J.; Mayer, M.; Normand, A.-C.; Genuneit, J.; Cookson, W.O.C.M.; Braun-Fahrländer, C.; Heederik, D.; Piarroux, R.; von Mutius, E. Exposure to Environmental Microorganisms and Childhood Asthma. N. Engl. J. Med. 2011, 364, 701–709. [Google Scholar] [CrossRef]
- Pivniouk, V.; Gimenes Junior, J.A.; Honeker, L.K.; Vercelli, D. The Role of Innate Immunity in Asthma Development and Protection: Lessons from the Environment. Clin. Exp. Allergy 2020, 50, 282–290. [Google Scholar] [CrossRef]
- Valkonen, M.; Wouters, I.M.; Täubel, M.; Rintala, H.; Lenters, V.; Vasara, R.; Genuneit, J.; Braun-Fahrländer, C.; Piarroux, R.; Von Mutius, E.; et al. Bacterial Exposures and Associations with Atopy and Asthma in Children. PLoS ONE 2015, 10, e0131594. [Google Scholar] [CrossRef] [Green Version]
- Depner, M.; Taft, D.H.; Kirjavainen, P.V.; Kalanetra, K.M.; Karvonen, A.M.; Peschel, S.; Schmausser-Hechfellner, E.; Roduit, C.; Frei, R.; Lauener, R.; et al. Maturation of the Gut Microbiome during the First Year of Life Contributes to the Protective Farm Effect on Childhood Asthma. Nat. Med. 2020, 26, 1766–1775. [Google Scholar] [CrossRef]
- Haahtela, T. Biodiversity for Resilience-What Is Needed for Allergic Children. Pediatr. Allergy Immunol. 2022, 33, e13779. [Google Scholar] [CrossRef]
- Hanski, I.; Von Hertzen, L.; Fyhrquist, N.; Koskinen, K.; Torppa, K.; Laatikainen, T.; Karisola, P.; Auvinen, P.; Paulin, L.; Mäkelä, M.J.; et al. Environmental Biodiversity, Human Microbiota, and Allergy Are Interrelated. Proc. Natl. Acad. Sci. USA 2012, 109, 8334–8339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtimäki, J.; Thorsen, J.; Rasmussen, M.A.; Hjelmsø, M.; Shah, S.; Mortensen, M.S.; Trivedi, U.; Vestergaard, G.; Bønnelykke, K.; Chawes, B.L.; et al. Urbanized Microbiota in Infants, Immune Constitution, and Later Risk of Atopic Diseases. J. Allergy Clin. Immunol. 2021, 148, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zhang, B.; Zhang, K.; Lv, X.; Wang, Q.; Bai, X. The Impact of Air Pollution on Intestinal Microbiome of Asthmatic Children: A Panel Study. BioMed Res. Int. 2020, 2020, 5753427. [Google Scholar] [CrossRef] [PubMed]
- Çolak, Y.; Afzal, S.; Nordestgaard, B.G.; Lange, P. Characteristics and Prognosis of Never-Smokers and Smokers with Asthma in the Copenhagen General Population Study. A Prospective Cohort Study. Am. J. Respir. Crit. Care Med. 2015, 192, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Panzer, A.R.; Sitarik, A.R.; Fadrosh, D.; Havstad, S.L.; Jones, K.; Davidson, B.; Finazzo, S.; Wegienka, G.R.; Woodcroft, K.; Lukacs, N.W.; et al. The Impact of Prenatal Dog Keeping on Infant Gut Microbiota Development. Clin. Exp. Allergy 2023, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tun, H.M.; Konya, T.; Takaro, T.K.; Brook, J.R.; Chari, R.; Field, C.J.; Guttman, D.S.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; et al. Exposure to Household Furry Pets Influences the Gut Microbiota of Infant at 3–4 Months Following Various Birth Scenarios. Microbiome 2017, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Shailesh, H.; Janahi, I.A. Role of Obesity in Inflammation and Remodeling of Asthmatic Airway. Life 2022, 12, 948. [Google Scholar] [CrossRef]
- Michalovich, D.; Rodriguez-Perez, N.; Smolinska, S.; Pirozynski, M.; Mayhew, D.; Uddin, S.; Van Horn, S.; Sokolowska, M.; Altunbulakli, C.; Eljaszewicz, A.; et al. Obesity and Disease Severity Magnify Disturbed Microbiome-Immune Interactions in Asthma Patients. Nat. Commun. 2019, 10, 5711. [Google Scholar] [CrossRef] [Green Version]
- Cloutier, M.M.; Baptist, A.P.; Blake, K.V.; Brooks, E.G.; Bryant-Stephens, T.; DiMango, E.; Dixon, A.E.; Elward, K.S.; Hartert, T.; Krishnan, J.A.; et al. 2020 Focused Updates to the Asthma Management Guidelines: A Report from the National Asthma Education and Prevention Program Coordinating Committee Expert Panel Working Group. J. Allergy Clin. Immunol. 2020, 146, 1217–1270. [Google Scholar] [CrossRef]
- Khan, L. Overview of the Updates for the Management of Asthma Guidelines. Pediatr. Ann. 2022, 51, e132–e135. [Google Scholar] [CrossRef]
- Durack, J.; Lynch, S.V.; Nariya, S.; Bhakta, N.R.; Beigelman, A.; Castro, M.; Dyer, A.M.; Israel, E.; Kraft, M.; Martin, R.J.; et al. Features of the Bronchial Bacterial Microbiome Associated with Atopy, Asthma and Responsiveness to Inhaled Corticosteroid Treatment. J. Allergy Clin. Immunol. 2017, 140, 63. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.J.; Nariya, S.; Harris, J.M.; Lynch, S.V.; Choy, D.F.; Arron, J.R.; Boushey, H. The Airway Microbiome in Severe Asthma: Associations with Disease Features and Severity. J. Allergy Clin. Immunol. 2015, 136, 874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozturk, A.B.; Turturice, B.A.; Perkins, D.L.; Finn, P.W. The Potential for Emerging Microbiome-Mediated Therapeutics in Asthma. Curr. Allergy Asthma Rep. 2017, 17, 62. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary Modulation of the Human Colonic Microbiota: Introducing the Concept of Prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Enam, F.; Mansell, T.J. Prebiotics: Tools to Manipulate the Gut Microbiome and Metabolome. J. Ind. Microbiol. Biotechnol. 2019, 46, 1445–1459. [Google Scholar] [CrossRef] [PubMed]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- Carlson, J.L.; Erickson, J.M.; Lloyd, B.B.; Slavin, J.L. Health Effects and Sources of Prebiotic Dietary Fiber. Curr. Dev. Nutr. 2018, 2, nzy005. [Google Scholar] [CrossRef] [Green Version]
- Panesar, P.S.; Kumari, S.; Panesar, R. Biotechnological Approaches for the Production of Prebiotics and Their Potential Applications. Crit. Rev. Biotechnol. 2013, 33, 345–364. [Google Scholar] [CrossRef]
- Comstock, S.S.; Wang, M.; Hester, S.N.; Li, M.; Donovan, S.M. Select Human Milk Oligosaccharides Directly Modulate Peripheral Blood Mononuclear Cells Isolated from 10-d-Old Pigs. Br. J. Nutr. 2014, 111, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Coppa, G.V.; Zampini, L.; Galeazzi, T.; Gabrielli, O. Prebiotics in Human Milk: A Review. Dig. Liver Dis. 2006, 38 (Suppl. S2), S291–S294. [Google Scholar] [CrossRef] [PubMed]
- Vohra, Y.; Vasan, M.; Venot, A.; Boons, G.J. One-Pot Synthesis of Oligosaccharides by Combining Reductive Openings of Benzylidene Acetals and Glycosylations. Org. Lett. 2008, 10, 3247–3250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborn, D.A.; Sinn, J.K. Prebiotics in Infants for Prevention of Allergy. Cochrane Database Syst. Rev. 2013, 2013, CD006474. [Google Scholar] [CrossRef]
- Wawryk-Gawda, E.; Markut-Miotła, E.; Emeryk, A. Postnatal Probiotics Administration Does Not Prevent Asthma in Children, but Using Prebiotics or Synbiotics May Be the Effective Potential Strategies to Decrease the Frequency of Asthma in High-Risk Children—A Meta-Analysis of Clinical Trials. Allergol. Immunopathol. 2021, 49, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Cuello-Garcia, C.; Fiocchi, A.; Pawankar, R.; Yepes-Nuñez, J.J.; Morgano, G.P.; Zhang, Y.; Agarwal, A.; Gandhi, S.; Terracciano, L.; Schünemann, H.J.; et al. Prebiotics for the Prevention of Allergies: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Clin. Exp. Allergy 2017, 47, 1468–1477. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The Pros, Cons, and Many Unknowns of Probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- Williams, N.T. Probiotics. Am. J. Health Syst. Pharm. 2010, 67, 449–458. [Google Scholar] [CrossRef]
- Chapman, C.M.C.; Gibson, G.R.; Rowland, I. Health Benefits of Probiotics: Are Mixtures More Effective than Single Strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef]
- Chapman, C.M.C.; Gibson, G.R.; Rowland, I. In Vitro Evaluation of Single- and Multi-Strain Probiotics: Inter-Species Inhibition between Probiotic Strains, and Inhibition of Pathogens. Anaerobe 2012, 18, 405–413. [Google Scholar] [CrossRef]
- McFarland, L.V. Efficacy of Single-Strain Probiotics Versus Multi-Strain Mixtures: Systematic Review of Strain and Disease Specificity. Dig. Dis. Sci. 2021, 66, 694–704. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in Medicine: A Long Debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef] [PubMed]
- Taibi, A.; Comelli, E.M. Practical Approaches to Probiotics Use. Appl. Physiol. Nutr. Metab. 2014, 39, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Krishna Rao, R.; Samak, G. Protection and Restitution of Gut Barrier by Probiotics: Nutritional and Clinical Implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Hertzberger, R.Y.; Knaus, U.G. Hydrogen Peroxide Production by Lactobacilli Promotes Epithelial Restitution during Colitis. Redox Biol. 2018, 16, 11–20. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial Effects on Host Energy Metabolism of Short-Chain Fatty Acids and Vitamins Produced by Commensal and Probiotic Bacteria. Microb. Cell Fact. 2017, 16, 79. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Ran, C.; Qin, C.; Li, S.; Zhang, H.; De Vos, W.M.; Ringø, E.; Zhou, Z. Anti-Infective Effect of Adhesive Probiotic Lactobacillus in Fish Is Correlated With Their Spatial Distribution in the Intestinal Tissue. Sci. Rep. 2017, 7, 13195. [Google Scholar] [CrossRef] [Green Version]
- Górska, A.; Przystupski, D.; Niemczura, M.J.; Kulbacka, J. Probiotic Bacteria: A Promising Tool in Cancer Prevention and Therapy. Curr. Microbiol. 2019, 76, 939–949. [Google Scholar] [CrossRef] [Green Version]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [Green Version]
- Hassanzad, M.; Maleki Mostashari, K.; Ghaffaripour, H.; Emami, H.; Limouei, S.R.; Velayati, A.A. Synbiotics and Treatment of Asthma: A Double-Blinded, Randomized, Placebo-Controlled Clinical Trial. Galen Med. J. 2019, 8, 1350. [Google Scholar] [CrossRef]
- Del Giudice, M.M.; Indolfi, C.; Capasso, M.; Maiello, N.; Decimo, F.; Ciprandi, G. Bifidobacterium Mixture (B Longum BB536, B Infantis M-63, B Breve M-16V) Treatment in Children with Seasonal Allergic Rhinitis and Intermittent Asthma. Ital. J. Pediatr. 2017, 43, 25. [Google Scholar] [CrossRef] [Green Version]
- Elazab, N.; Mendy, A.; Gasana, J.; Vieira, E.R.; Quizon, A.; Forno, E. Probiotic Administration in Early Life, Atopy, and Asthma: A Meta-Analysis of Clinical Trials. Pediatrics 2013, 132, e666–e676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Jiang, P.; Liu, J.; Sun, R.; Zhu, L. Association between Probiotic Supplementation and Asthma Incidence in Infants: A Meta-Analysis of Randomized Controlled Trials. J. Asthma 2020, 57, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhang, Y.; He, C.; Dai, J. Probiotics Supplementation in Children with Asthma: A Systematic Review and Meta-Analysis. J. Paediatr. Child Health 2018, 54, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Grandinetti, R.; Fainardi, V.; Caffarelli, C.; Capoferri, G.; Lazzara, A.; Tornesello, M.; Meoli, A.; Bergamini, B.M.; Bertelli, L.; Biserna, L.; et al. Risk Factors Affecting Development and Persistence of Preschool Wheezing: Consensus Document of the Emilia-Romagna Asthma (ERA) Study Group. J. Clin. Med. 2022, 11, 6558. [Google Scholar] [CrossRef] [PubMed]
- Ciprandi, G.; Tosca, M.A. Probiotics in Children with Asthma. Children 2022, 9, 978. [Google Scholar] [CrossRef] [PubMed]
- Sangkanjanavanich, S.; Pradubpongsa, P.; Mitthamsiri, W.; Sangasapaviliya, A.; Boonpiyathad, T. Bifidobacterium Infantis 35624 Efficacy in Patients with Uncontrolled Asthma: A Randomized Placebo-Controlled Trial. Ann. Allergy. Asthma Immunol. 2022, 129, 790–792. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Liu, F.; Gao, Q.; Wang, R.; Zhang, L.; Li, Y. A Meta-Analysis of Probiotics for the Treatment of Allergic Airway Diseases in Children and Adolescents. Am. J. Rhinol. Allergy 2022, 36, 480–490. [Google Scholar] [CrossRef]
- Wood, L.G. Diet, Obesity, and Asthma. Ann. Am. Thorac. Soc. 2017, 14, S332–S338. [Google Scholar] [CrossRef]
- Alwarith, J.; Kahleova, H.; Crosby, L.; Brooks, A.; Brandon, L.; Levin, S.M.; Barnard, N.D. The Role of Nutrition in Asthma Prevention and Treatment. Nutr. Rev. 2020, 78, 928–938. [Google Scholar] [CrossRef] [Green Version]
- Okoniewski, W.; Lu, K.D.; Forno, E. Weight Loss for Children and Adults with Obesity and Asthma. A Systematic Review of Randomized Controlled Trials. Ann. Am. Thorac. Soc. 2019, 16, 613–625. [Google Scholar] [CrossRef]
- Nuzzi, G.; Di Cicco, M.; Trambusti, I.; Agosti, M.; Peroni, D.G.; Comberiati, P. Primary Prevention of Pediatric Asthma through Nutritional Interventions. Nutrients 2022, 14, 754. [Google Scholar] [CrossRef] [PubMed]
- Venter, C.; Agostoni, C.; Arshad, S.H.; Ben-Abdallah, M.; Du Toit, G.; Fleischer, D.M.; Greenhawt, M.; Glueck, D.H.; Groetch, M.; Lunjani, N.; et al. Dietary Factors during Pregnancy and Atopic Outcomes in Childhood: A Systematic Review from the European Academy of Allergy and Clinical Immunology. Pediatr. Allergy Immunol. 2020, 31, 889. [Google Scholar] [CrossRef] [PubMed]
- Best, K.P.; Gold, M.; Kennedy, D.; Martin, J.; Makrides, M. Omega-3 Long-Chain PUFA Intake during Pregnancy and Allergic Disease Outcomes in the Offspring: A Systematic Review and Meta-Analysis of Observational Studies and Randomized Controlled Trials. Am. J. Clin. Nutr. 2016, 103, 128–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, C.; Meyer, R.W.; Nwaru, B.I.; Roduit, C.; Untersmayr, E.; Adel-Patient, K.; Agache, I.; Agostoni, C.; Akdis, C.A.; Bischoff, S.C.; et al. EAACI Position Paper: Influence of Dietary Fatty Acids on Asthma, Food Allergy, and Atopic Dermatitis. Allergy 2019, 74, 1429–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, G.C.; da Silva, D.T.R.; Hennemann, M.L.; Sarmento, R.A.; Almeida, J.C.; de Tarso Roth Dalcin, P. Diet Effects in the Asthma Treatment: A Systematic Review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1878–1887. [Google Scholar] [CrossRef]
- Wang, J.W.; Kuo, C.H.; Kuo, F.C.; Wang, Y.K.; Hsu, W.H.; Yu, F.J.; Hu, H.M.; Hsu, P.I.; Wang, J.Y.; Wu, D.C. Fecal Microbiota Transplantation: Review and Update. J. Formos. Med. Assoc. 2019, 118 (Suppl. S1), S23–S31. [Google Scholar] [CrossRef]
- Xu, M.Q.; Cao, H.L.; Wang, W.Q.; Wang, S.; Cao, X.C.; Yan, F.; Wang, B.M. Fecal Microbiota Transplantation Broadening Its Application beyond Intestinal Disorders. World J. Gastroenterol. 2015, 21, 102. [Google Scholar] [CrossRef]
- Kang, Y.; Cai, Y. Future Prospect of Faecal Microbiota Transplantation as a Potential Therapy in Asthma. Allergol. Immunopathol. 2018, 46, 307–309. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Genera/Phyla | Fungal Genera/Fungus Taxa | Virus Genera | |
|---|---|---|---|
| Gastrointestinal microbiome | Bacteroides, Eubacterium, Peptococcus, Peptostreptococcus, Clostridium, Ruminococcus, Faecalibacterium, Escherichia, Lactobacillus | Candida, Saccharomyces, Aspergillus, Penicillium, Bullera, Pleospora, Rhodotorula, Trametes, Galactomyces | Bacteriophages |
| Respiratory microbiome | Prevotella, Streptococcus, Veillonella, Firmicutes, Bacteroidetes | Ascomycota, Basidiomycota, Candida, Saccharomyces, Penicillium, Cladosporium, Fusarium | Understudied |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Logoń, K.; Świrkosz, G.; Nowak, M.; Wrześniewska, M.; Szczygieł, A.; Gomułka, K. The Role of the Microbiome in the Pathogenesis and Treatment of Asthma. Biomedicines 2023, 11, 1618. https://doi.org/10.3390/biomedicines11061618
Logoń K, Świrkosz G, Nowak M, Wrześniewska M, Szczygieł A, Gomułka K. The Role of the Microbiome in the Pathogenesis and Treatment of Asthma. Biomedicines. 2023; 11(6):1618. https://doi.org/10.3390/biomedicines11061618
Chicago/Turabian StyleLogoń, Katarzyna, Gabriela Świrkosz, Monika Nowak, Martyna Wrześniewska, Aleksandra Szczygieł, and Krzysztof Gomułka. 2023. "The Role of the Microbiome in the Pathogenesis and Treatment of Asthma" Biomedicines 11, no. 6: 1618. https://doi.org/10.3390/biomedicines11061618