
Obesity-Associated Non-T2 Mechanisms in Obese Asthmatic Individuals

Abstract
:1. Introduction

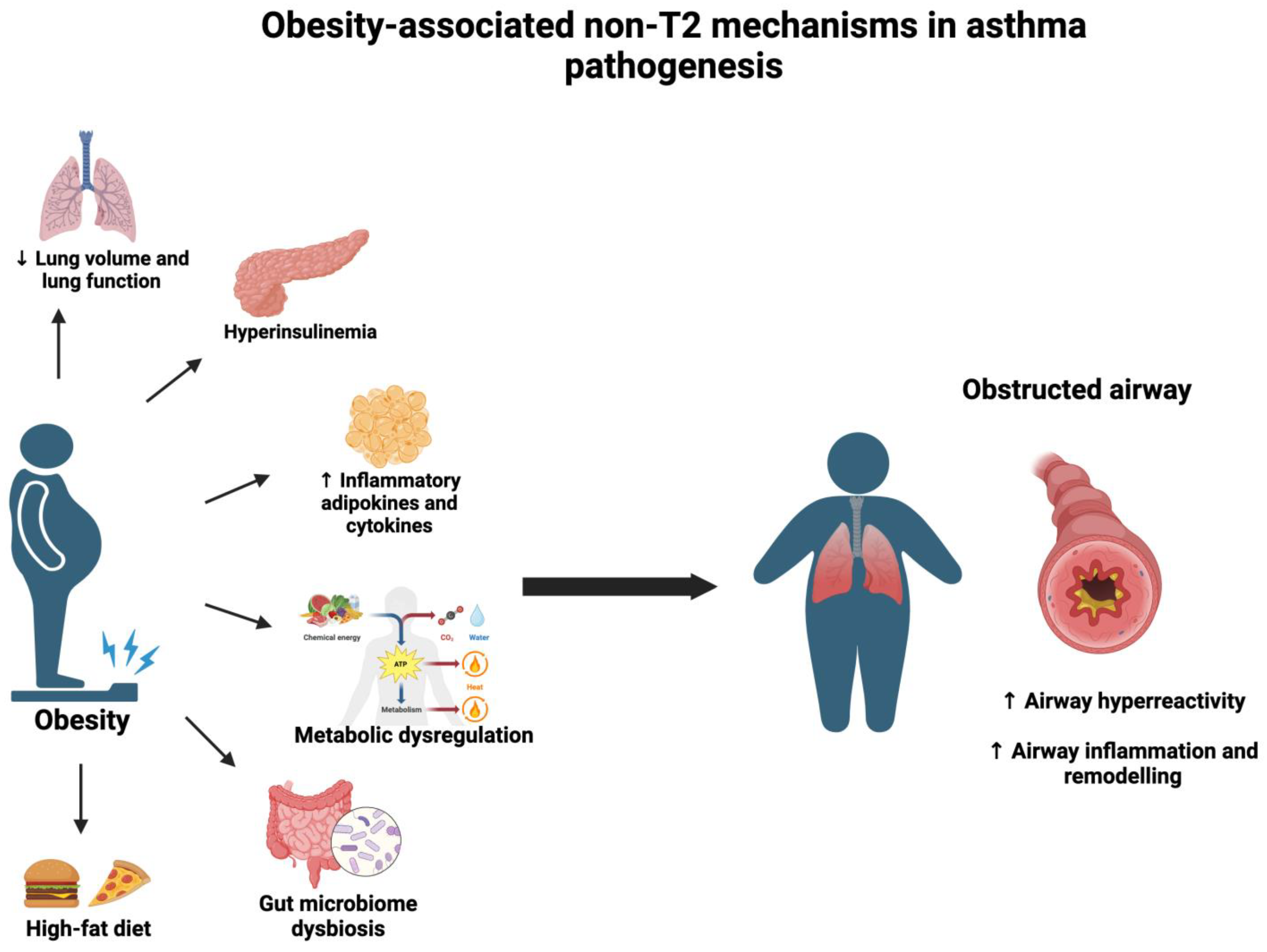{kind=link}
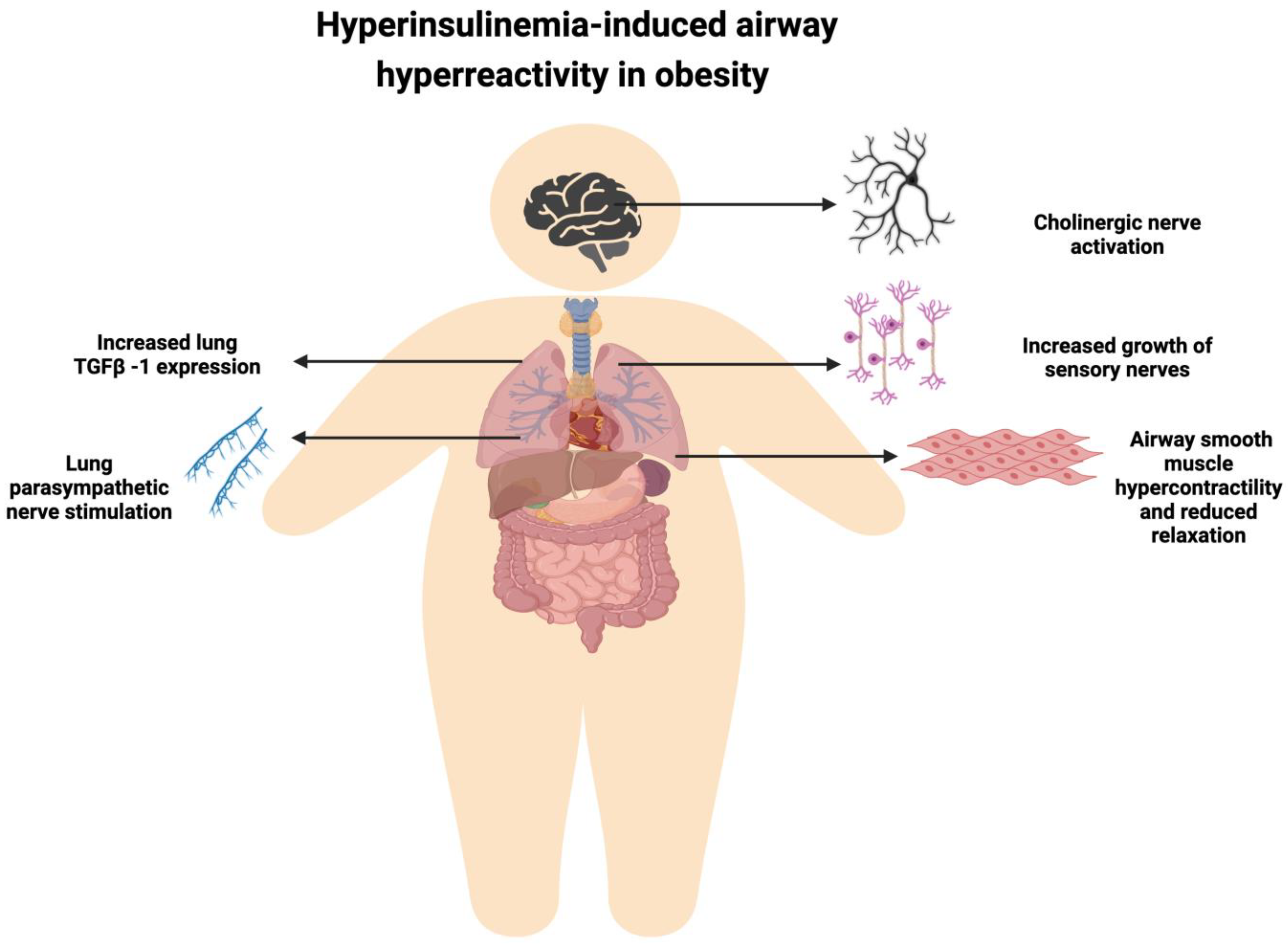{kind=link}
| Obesity-Associated Non-T2 Factors | Type of Study | Source | Key Findings | Authors |
|---|---|---|---|---|
| Adiposity-associated changes in lung mechanics | In humans | Retrospective study of 373 adults with different BMI | ↑ BMI is associated with exponential reduction in FRC and ERV | Jones et al., 2006 [18] |
| In humans | Cross-sectional study of healthy adults | ↑ WC and WHR are associated with noticeable changes with volume, capacity, and mechanics of the lung | Shanmugasundaram et al., 2023 [19] | |
| In humans | Retrospective study from mild to moderate obese asthmatic children of Childhood Asthma Management Program (CAMP) | ↑ BMI is associated with increase in FEV1 and FVC and decrease in FEV1/FVC ratio | Tantisira et al., 2003 [20] | |
| Hyperinsulinemia | Ex vivo | SMCs from bovine trachea | Hypercontractilibility of SMCs | Gosens et al., 2003 [21] |
| Ex vivo | SMCs from bovine trachea | ↑ Accumulation of smooth muscle myosin and laminin | Dekkers et al., 2009 [22] | |
| In vitro Ex vivo | Human ASMs Trachea of high-fat-diet-fed mice | ↓ β-AR agonist-mediated relaxation of airway smooth muscle cells | Xu et al., 2020 [23] | |
| In vivo Ex vivo | Diet-induced obese mice Human tracheal smooth muscle strips | ↑ Vagus nerve-stimulated airway hyper-responsiveness by inhibiting M2 muscarinic receptors ↑ Contraction of trachial smooth muscle cells in response to electric field stimulation by inhibiting M2 muscarinic receptors | Nei et al., 2014 [24] | |
| In vivo | Diet-induced obese mice | Reducing insulin levels decreases vagus nerve-stimulated airway hyper-responsiveness by restoring M2 muscarinic receptor function | Proskocil et al., 2021 [25] | |
| In vivo | Diet-induced obese mice | ↑ cholinergic nerve activation and airway hyper-responsiveness | Leiria et al., 2014 [12] | |
| In vivo | Diet-induced obese mice | ↑ sensory nerve population in the lung | Calco et al., 2022 [26] | |
| In vivo | Diet-induced obese mice | ↑ TGF-β1 expression in the lung, fibrosis of the lung, and airway hyper-responsiveness | Park et al., 2019 [27] | |
| In humans | Retrospective data from obese children | ↑ increased risk of allergic asthma in obese children | Sánchez Jiménez et al., 2014 [28] | |
| In humans | Retrospective data from National Health and Nutrition Examination Survey (NHANES), United States | Insulin resistance increases the risk of asthma incidence in obese individuals in a dose-dependent manner, independent of hypertension, systemic inflammation, hyperglycemia, and hypertriglyceridemia | Cardet et al., 2016 [29] | |
| In human | Retrospective cohort of US adults with obesity and asthma | Prediabetes and diabetes states increase asthma exacerbation rate by 27% and 33%, respectively | Wu et al., 2019 [30] | |
| Inflammation | In humans | Obese asthmatic women vs. obese women (in adults) | ↑ leptin increases systemic inflammation and airway reactivity without lung inflammation in obese asthma | Sideleva et al., 2012 [31] |
| In vitro | Normal human lung fibroblasts | leptin promotes the expression of MCP-1, eotaxin, IP-10, IL-6, and IL-8 | Watanabe et al., 2019 [32] | |
| In vitro | Human primary bronchial epithelial cells | Leptin promotes the expression of ICAM-1, CCL11, VEGF, IL-6, and G-CSF | Suzukawa et al., 2015 [33] | |
| Metabolic dysregulation | In vivo | Diet-induced obese mice | Changes in metabolomic profile of lung with altered concentration of 1C and TCA cycle metabolites | Rautureau et al., 2021 [34] |
| In humans | Obese asthma vs. lean asthma (in adults) | Changes in metabolomic profile of serum, sputum, and PBMCs. Altered metabolomics are associated with airway inflammation | Liu et al., 2018 [35] | |
| Microbiome dysregulation | In humans | Obese asthma, obese non-asthma, non-obese asthma, and healthy control (in adults) | Obesity and asthma synergistically contribute to inflammation and microbiome alteration | Michalovich et al., 2019 [36] |
| High-fat diet | In vivo | Diet-induced obese mice vs. normal-diet-fed mice | high-fat diet increases AHR, serum IgE, and lung TNF-α upon OVA challenge | Kim et al., 2015 [37] |
| In humans In vivo | Obese asthmatics vs. lean asthmatics (in adults) High-fat diet induced obese mice vs. normal-diet-fed mice | Increased levels of palmitic acid and DPP4 in the BALF of obese asthmatics as compared to lean asthmatics High-fat-diet-associated increase in palmitic acid increases eosinophils, neutrophils, and eotaxin -2 in BALF upon allergic trigger | Dimasuay et al., 2023 [38] |
2. Methodology
3. Obesity and Lung Mechanics
4. Obesity-Associated Hyperinsulinemia and Airway Hyper-Responsiveness
5. Obesity, Inflammation, and Asthma
6. Metabolic Dysregulation in Obesity and Asthma
7. Obesity, Asthma, and Microbiome
8. Diet in Obesity-Associated Asthma
9. Current Management and Potential Future Therapies for Obesity-Associated Asthma
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Global Asthma Network. The Global Asthma Report, New Zealand. 2018. Available online: http://globalasthmanetwork.org/Global%20asthma%20Report%202018%20Embargo.pdf (accessed on 29 August 2023).
- Papi, A.; Brightling, C.; Pedersen, S.E.; Reddel, H.K. Asthma. Lancet 2018, 391, 783–800. [Google Scholar] [CrossRef]
- Ahmadiafshar, A.; Tabbekhha, S.; Mousavinasab, N.; Khoshnevis, P. Relation between asthma and body mass index in 6–15 years old children. Acta Med. Iran 2013, 51, 615–619. [Google Scholar] [PubMed]
- Black, M.H.; Zhou, H.; Takayanagi, M.; Jacobsen, S.J.; Koebnick, C. Increased asthma risk and asthma-related health care complications associated with childhood obesity. Am. J. Epidemiol. 2013, 178, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Akinbami, L.J.; Rossen, L.M.; Fakhouri, T.H.I.; Simon, A.E.; Kit, B.K. Contribution of weight status to asthma prevalence racial disparities, 2–19 year olds, 1988–2014. Ann. Epidemiol. 2017, 27, 472–478.e473. [Google Scholar] [CrossRef] [PubMed]
- Amra, B.; Rahmani, A.; Salimi, S.; Mohammadzadeh, Z.; Golshan, M. Association between Asthma and Body Mass Index in Children. Iran J. Allergy Asthma Immunol. 2005, 4, 33–37. [Google Scholar]
- Malden, S.; Gillespie, J.; Hughes, A.; Gibson, A.M.; Farooq, A.; Martin, A.; Summerbell, C.; Reilly, J.J. Obesity in young children and its relationship with diagnosis of asthma, vitamin D deficiency, iron deficiency, specific allergies and flat-footedness: A systematic review and meta-analysis. Obes. Rev. 2021, 22, e13129. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Kuo, H.-P.; Hsia, S.-M.; Wu, H.-T.; Pan, W.-H.; Lee, Y.L. Life course body mass index through childhood and young adulthood and risks of asthma and pulmonary function impairment. Pediatr. Pulmonol. 2021, 56, 849–857. [Google Scholar] [CrossRef]
- Peters, U.; Dixon, A.E.; Forno, E. Obesity and asthma. J. Allergy Clin. Immunol. 2018, 141, 1169–1179. [Google Scholar] [CrossRef]
- Lessard, A.; Turcotte, H.; Cormier, Y.; Boulet, L.P. Obesity and asthma: A specific phenotype? Chest 2008, 134, 317–323. [Google Scholar] [CrossRef]
- Baffi, C.W.; Winnica, D.E.; Holguin, F. Asthma and obesity: Mechanisms and clinical implications. Asthma Res. Pract. 2015, 1, 1. [Google Scholar] [CrossRef]
- Leiria, L.O.S.; Arantes-Costa, F.M.; Calixto, M.C.; Alexandre, E.C.; Moura, R.F.; Folli, F.; Prado, C.M.; Prado, M.A.; Prado, V.F.; Velloso, L.A.; et al. Increased Airway Reactivity and Hyperinsulinemia in Obese Mice Are Linked by ERK Signaling in Brain Stem Cholinergic Neurons. Cell Rep. 2015, 11, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Cowan, D.C. Obesity, Inflammation, and Severe Asthma: An Update. Curr. Allergy Asthma Rep. 2021, 21, 46. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.C.; Schiebler, M.L.; Cardet, J.C.; Johansson, M.W.; Sorkness, R.; DeBoer, M.D.; Bleecker, E.R.; Meyers, D.A.; Castro, M.; Sumino, K.; et al. The Impact of Insulin Resistance on Loss of Lung Function and Response to Treatment in Asthma. Am. J. Respir. Crit. Care Med. 2022, 206, 1096–1106. [Google Scholar] [CrossRef] [PubMed]
- Shailesh, H.; Janahi, I.A. Role of Obesity in Inflammation and Remodeling of Asthmatic Airway. Life 2022, 12, 948. [Google Scholar] [CrossRef]
- Kunøe, A.; Sevelsted, A.; Chawes, B.L.; Stokholm, J.; Eliasen, A.; Krakauer, M.; Bønnelykke, K.; Bisgaard, H. Associations between Inhaled Corticosteroid Use in the First 6 Years of Life and Obesity-related Traits. Am. J. Respir. Crit. Care Med. 2021, 204, 642–650. [Google Scholar] [CrossRef]
- Glazebrook, C.; McPherson, A.C.; Macdonald, I.A.; Swift, J.A.; Ramsay, C.; Newbould, R.; Smyth, A. Asthma as a barrier to children’s physical activity: Implications for body mass index and mental health. Pediatrics 2006, 118, 2443–2449. [Google Scholar] [CrossRef]
- Jones, R.L.; Nzekwu, M.-M.U. The Effects of Body Mass Index on Lung Volumes. Chest 2006, 130, 827–833. [Google Scholar] [CrossRef]
- Shanmugasundaram, K.; Bade, G.; Sampath, M.; Talwar, A. Effect of Obesity on Airway Mechanics. Indian J. Endocrinol. Metab. 2023, 27, 161–166. [Google Scholar] [CrossRef]
- Tantisira, K.G.; Litonjua, A.A.; Weiss, S.T.; Fuhlbrigge, A.L. Association of body mass with pulmonary function in the Childhood Asthma Management Program (CAMP). Thorax 2003, 58, 1036–1041. [Google Scholar] [CrossRef]
- Gosens, R.; Nelemans, S.A.; Hiemstra, M.; Grootte Bromhaar, M.M.; Meurs, H.; Zaagsma, J. Insulin induces a hypercontractile airway smooth muscle phenotype. Eur. J. Pharmacol. 2003, 481, 125–131. [Google Scholar] [CrossRef]
- Dekkers, B.G.; Schaafsma, D.; Tran, T.; Zaagsma, J.; Meurs, H. Insulin-induced laminin expression promotes a hypercontractile airway smooth muscle phenotype. Am. J. Respir. Cell Mol. Biol. 2009, 41, 494–504. [Google Scholar] [CrossRef]
- Xu, R.; Gopireddy, R.R.; Wu, Y.; Wu, L.; Tao, X.; Shao, J.; Wang, W.; Li, L.; Jovanovic, A.; Xu, B.; et al. Hyperinsulinemia promotes heterologous desensitization of β(2) adrenergic receptor in airway smooth muscle in obesity. FASEB J. 2020, 34, 3996–4008. [Google Scholar] [CrossRef]
- Nie, Z.; Jacoby, D.B.; Fryer, A.D. Hyperinsulinemia potentiates airway responsiveness to parasympathetic nerve stimulation in obese rats. Am. J. Respir. Cell Mol. Biol. 2014, 51, 251–261. [Google Scholar] [CrossRef]
- Proskocil, B.J.; Fryer, A.D.; Jacoby, D.B.; Nie, Z. Pioglitazone prevents obesity-related airway hyperreactivity and neuronal M2 receptor dysfunction. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 321, L236–L247. [Google Scholar] [CrossRef] [PubMed]
- Calco, G.N.; Maung, J.N.; Jacoby, D.B.; Fryer, A.D.; Nie, Z. Insulin increases sensory nerve density and reflex bronchoconstriction in obese mice. JCI Insight 2022, 7, e161898. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.H.; Oh, E.Y.; Han, H.; Yang, M.; Park, H.J.; Park, K.H.; Lee, J.H.; Park, J.W. Insulin resistance mediates high-fat diet-induced pulmonary fibrosis and airway hyperresponsiveness through the TGF-β1 pathway. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Jiménez, J.; Herrero Espinet, F.J.; Mengibar Garrido, J.M.; Roca Antonio, J.; Peños Mayor, S.; Peñas Boira Mdel, M.; Roca Comas, A.; Ballester Martínez, A. Asthma and insulin resistance in obese children and adolescents. Pediatr Allergy Immunol. 2014, 25, 699–705. [Google Scholar] [CrossRef]
- Cardet, J.C.; Ash, S.; Kusa, T.; Camargo, C.A., Jr.; Israel, E. Insulin resistance modifies the association between obesity and current asthma in adults. Eur. Respir. J. 2016, 48, 403–410. [Google Scholar] [CrossRef]
- Wu, T.D.; Brigham, E.P.; Keet, C.A.; Brown, T.T.; Hansel, N.N.; McCormack, M.C. Association Between Prediabetes/Diabetes and Asthma Exacerbations in a Claims-Based Obese Asthma Cohort. J. Allergy Clin. Immunol. Pract. 2019, 7, 1868–1873.e1865. [Google Scholar] [CrossRef]
- Sideleva, O.; Suratt, B.T.; Black, K.E.; Tharp, W.G.; Pratley, R.E.; Forgione, P.; Dienz, O.; Irvin, C.G.; Dixon, A.E. Obesity and asthma: An inflammatory disease of adipose tissue not the airway. Am. J. Respir. Crit. Care Med. 2012, 186, 598–605. [Google Scholar] [CrossRef]
- Watanabe, K.; Suzukawa, M.; Arakawa, S.; Kobayashi, K.; Igarashi, S.; Tashimo, H.; Nagai, H.; Tohma, S.; Nagase, T.; Ohta, K. Leptin enhances cytokine/chemokine production by normal lung fibroblasts by binding to leptin receptor. Allergol. Int. 2019, 68, S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Suzukawa, M.; Koketsu, R.; Baba, S.; Igarashi, S.; Nagase, H.; Yamaguchi, M.; Matsutani, N.; Kawamura, M.; Shoji, S.; Hebisawa, A.; et al. Leptin enhances ICAM-1 expression, induces migration and cytokine synthesis, and prolongs survival of human airway epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L801–L811. [Google Scholar] [CrossRef] [PubMed]
- Rautureau, G.J.P.; Morio, B.; Guibert, S.; Lefevre, C.; Perrier, J.; Alves, A.; Chauvin, M.A.; Pinteur, C.; Monet, M.A.; Godet, M.; et al. Dietary obesity in mice is associated with lipid deposition and metabolic shifts in the lungs sharing features with the liver. Sci. Rep. 2021, 11, 8712. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, J.; Zhang, H.P.; Zhang, X.; Wang, L.; Wood, L.; Wang, G. Obesity-Associated Metabolic Signatures Correlate to Clinical and Inflammatory Profiles of Asthma: A Pilot Study. Allergy Asthma Immunol. Res. 2018, 10, 628–647. [Google Scholar] [CrossRef] [PubMed]
- Michalovich, D.; Rodriguez-Perez, N.; Smolinska, S.; Pirozynski, M.; Mayhew, D.; Uddin, S.; Van Horn, S.; Sokolowska, M.; Altunbulakli, C.; Eljaszewicz, A.; et al. Obesity and disease severity magnify disturbed microbiome-immune interactions in asthma patients. Nat. Commun. 2019, 10, 5711. [Google Scholar] [CrossRef]
- Kim, J.Y.; Sohn, J.H.; Lee, J.H.; Park, J.W. Obesity increases airway hyperresponsiveness via the TNF-α pathway and treating obesity induces recovery. PLoS ONE 2015, 10, e0116540. [Google Scholar] [CrossRef]
- Dimasuay, K.G.; Berg, B.; Schaunaman, N.; Holguin, F.; Winnica, D.; Chu, H.W. High-fat diet and palmitic acid amplify airway type 2 inflammation. Front. Allergy 2023, 4, 1193480. [Google Scholar] [CrossRef]
- Forno, E.; Weiner, D.J.; Mullen, J.; Sawicki, G.; Kurland, G.; Han, Y.Y.; Cloutier, M.M.; Canino, G.; Weiss, S.T.; Litonjua, A.A.; et al. Obesity and Airway Dysanapsis in Children with and without Asthma. Am. J. Respir. Crit. Care Med. 2017, 195, 314–323. [Google Scholar] [CrossRef]
- Sharp, J.T.; Henry, J.P.; Sweany, S.K.; Meadows, W.R.; Pietras, R.J. The total work of breathing in normal and obese men. J. Clin. Investig. 1964, 43, 728–739. [Google Scholar] [CrossRef]
- Mafort, T.T.; Rufino, R.; Costa, C.H.; Lopes, A.J. Obesity: Systemic and pulmonary complications, biochemical abnormalities, and impairment of lung function. Multidiscip. Respir. Med. 2016, 11, 28. [Google Scholar] [CrossRef]
- Hibbert, K.; Rice, M.; Malhotra, A. Obesity and ARDS. Chest 2012, 142, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Bates, J.H. Physiological Mechanisms of Airway Hyperresponsiveness in Obese Asthma. Am. J. Respir. Cell Mol. Biol. 2016, 54, 618–623. [Google Scholar] [CrossRef] [PubMed]
- van Huisstede, A.; Rudolphus, A.; Castro Cabezas, M.; Biter, L.U.; van de Geijn, G.J.; Taube, C.; Hiemstra, P.S.; Braunstahl, G.J. Effect of bariatric surgery on asthma control, lung function and bronchial and systemic inflammation in morbidly obese subjects with asthma. Thorax 2015, 70, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Pakhale, S.; Baron, J.; Dent, R.; Vandemheen, K.; Aaron, S.D. Effects of weight loss on airway responsiveness in obese adults with asthma: Does weight loss lead to reversibility of asthma? Chest 2015, 147, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Martinez, K.E.; Tucker, L.A.; Bailey, B.W.; LeCheminant, J.D. Expanded Normal Weight Obesity and Insulin Resistance in US Adults of the National Health and Nutrition Examination Survey. J. Diabetes Res. 2017, 2017, 9502643. [Google Scholar] [CrossRef]
- Thuesen, B.H.; Husemoen, L.L.N.; Hersoug, L.G.; Pisinger, C.; Linneberg, A. Insulin resistance as a predictor of incident asthma-like symptoms in adults. Clin. Exp. Allergy 2009, 39, 700–707. [Google Scholar] [CrossRef]
- Kim, K.M.; Kim, S.S.; Lee, S.H.; Song, W.J.; Chang, Y.S.; Min, K.U.; Cho, S.H. Association of insulin resistance with bronchial hyperreactivity. Asia Pac. Allergy 2014, 4, 99–105. [Google Scholar] [CrossRef]
- Spina, D. Airway sensory nerves: A burning issue in asthma? Thorax 1996, 51, 335–337. [Google Scholar] [CrossRef]
- Roffel, A.F.; Elzinga, C.R.; Zaagsma, J. Muscarinic M3 receptors mediate contraction of human central and peripheral airway smooth muscle. Pulm. Pharmacol. 1990, 3, 47–51. [Google Scholar] [CrossRef]
- Al-Shawwa, B.A.; Al-Huniti, N.H.; DeMattia, L.; Gershan, W. Asthma and insulin resistance in morbidly obese children and adolescents. J. Asthma 2007, 44, 469–473. [Google Scholar] [CrossRef]
- Rastogi, D.; Holguin, F. Metabolic Dysregulation, Systemic Inflammation, and Pediatric Obesity-related Asthma. Ann. Am. Thorac. Soc. 2017, 14, S363–S367. [Google Scholar] [CrossRef]
- Magrone, T.; Simone, M.; Altamura, M.; Munno, I. Characterization of the immune inflammatory profile in obese asthmatic children. Endocr. Metab. Immune Disord. Drug Targets 2014, 14, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Di Cicco, M.; Ghezzi, M.; Kantar, A.; Song, W.-J.; Bush, A.; Peroni, D.; D’Auria, E. Pediatric obesity and severe asthma: Targeting pathways driving inflammation. Pharmacol. Res. 2023, 188, 106658. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.E.; Poynter, M.E. Mechanisms of Asthma in Obesity. Pleiotropic Aspects of Obesity Produce Distinct Asthma Phenotypes. Am. J. Respir. Cell Mol. Biol. 2016, 54, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Divoux, A.; Moutel, S.; Poitou, C.; Lacasa, D.; Veyrie, N.; Aissat, A.; Arock, M.; Guerre-Millo, M.; Clement, K. Mast cells in human adipose tissue: Link with morbid obesity, inflammatory status, and diabetes. J. Clin. Endocrinol. Metab. 2012, 97, E1677–E1685. [Google Scholar] [CrossRef]
- Kunz, H.E.; Hart, C.R.; Gries, K.J.; Parvizi, M.; Laurenti, M.; Dalla Man, C.; Moore, N.; Zhang, X.; Ryan, Z.; Polley, E.C.; et al. Adipose tissue macrophage populations and inflammation are associated with systemic inflammation and insulin resistance in obesity. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E105–E121. [Google Scholar]
- Wegner, C.D.; Gundel, R.H.; Reilly, P.; Haynes, N.; Letts, L.G.; Rothlein, R. Intercellular adhesion molecule-1 (ICAM-1) in the pathogenesis of asthma. Science 1990, 247, 456–459. [Google Scholar] [CrossRef]
- Reynoso-García, J.; Miranda-Santiago, A.E.; Meléndez-Vázquez, N.M.; Acosta-Pagán, K.; Sánchez-Rosado, M.; Díaz-Rivera, J.; Rosado-Quiñones, A.M.; Acevedo-Márquez, L.; Cruz-Roldán, L.; Tosado-Rodríguez, E.L.; et al. A complete guide to human microbiomes: Body niches, transmission, development, dysbiosis, and restoration. Front. Syst. Biol. 2022, 2, 951403. [Google Scholar] [CrossRef]
- Frati, F.; Salvatori, C.; Incorvaia, C.; Bellucci, A.; Di Cara, G.; Marcucci, F.; Esposito, S. The Role of the Microbiome in Asthma: The Gut–Lung Axis. Int. J. Mol. Sci. 2018, 20, 123. [Google Scholar] [CrossRef]
- Stokholm, J.; Blaser, M.J.; Thorsen, J.; Rasmussen, M.A.; Waage, J.; Vinding, R.K.; Schoos, A.M.; Kunøe, A.; Fink, N.R.; Chawes, B.L.; et al. Maturation of the gut microbiome and risk of asthma in childhood. Nat. Commun. 2018, 9, 141. [Google Scholar] [CrossRef]
- Arrieta, M.-C.; Stiemsma, L.T.; Dimitriu, P.A.; Thorson, L.; Russell, S.; Yurist-Doutsch, S.; Kuzeljevic, B.; Gold, M.J.; Britton, H.M.; Lefebvre, D.L.; et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci. Transl. Med. 2015, 7, 307ra152. [Google Scholar] [CrossRef] [PubMed]
- Galazzo, G.; van Best, N.; Bervoets, L.; Dapaah, I.O.; Savelkoul, P.H.; Hornef, M.W.; Lau, S.; Hamelmann, E.; Penders, J. Development of the Microbiota and Associations With Birth Mode, Diet, and Atopic Disorders in a Longitudinal Analysis of Stool Samples, Collected From Infancy Through Early Childhood. Gastroenterology 2020, 158, 1584–1596. [Google Scholar] [CrossRef] [PubMed]
- Menegati, L.M.; de Oliveira, E.E.; Oliveira, B.d.C.; Macedo, G.C.; de Castro e Silva, F.M. Asthma, obesity, and microbiota: A complex immunological interaction. Immunol. Lett. 2023, 255, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Bilotta, A.J.; Cong, Y. Gut microbiota metabolite regulation of host defenses at mucosal surfaces: Implication in precision medicine. Precis. Clin. Med. 2019, 2, 110–119. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Arévalo, A.; Stiemsma, L.; Dimitriu, P.; Chico, M.E.; Loor, S.; Vaca, M.; Boutin, R.C.T.; Morien, E.; Jin, M.; et al. Associations between infant fungal and bacterial dysbiosis and childhood atopic wheeze in a nonindustrialized setting. J. Allergy Clin. Immunol. 2018, 142, 424–434.e410. [Google Scholar] [CrossRef]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef]
- Folkerts, J.; Redegeld, F.; Folkerts, G.; Blokhuis, B.; van den Berg, M.P.M.; de Bruijn, M.J.W.; van Ijcken, W.F.J.; Junt, T.; Tam, S.Y.; Galli, S.J.; et al. Butyrate inhibits human mast cell activation via epigenetic regulation of FcεRI-mediated signaling. Allergy 2020, 75, 1966–1978. [Google Scholar] [CrossRef]
- Barcik, W.; Pugin, B.; Westermann, P.; Perez, N.R.; Ferstl, R.; Wawrzyniak, M.; Smolinska, S.; Jutel, M.; Hessel, E.M.; Michalovich, D.; et al. Histamine-secreting microbes are increased in the gut of adult asthma patients. J. Allergy Clin. Immunol. 2016, 138, 1491–1494.e1497. [Google Scholar] [CrossRef]
- Yamauchi, K.; Ogasawara, M. The Role of Histamine in the Pathophysiology of Asthma and the Clinical Efficacy of Antihistamines in Asthma Therapy. Int. J. Mol. Sci. 2019, 20, 1733. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Womble, J.T.; Gunsch, C.K.; Ingram, J.L. The Gut/Lung Microbiome Axis in Obesity, Asthma, and Bariatric Surgery: A Literature Review. Obesity 2021, 29, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Pinart, M.; Dötsch, A.; Schlicht, K.; Laudes, M.; Bouwman, J.; Forslund, S.K.; Pischon, T.; Nimptsch, K. Gut Microbiome Composition in Obese and Non-Obese Persons: A Systematic Review and Meta-Analysis. Nutrients 2021, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.G.; Li, Q.; Scott, H.A.; Rutting, S.; Berthon, B.S.; Gibson, P.G.; Hansbro, P.M.; Williams, E.; Horvat, J.; Simpson, J.L.; et al. Saturated fatty acids, obesity, and the nucleotide oligomerization domain-like receptor protein 3 (NLRP3) inflammasome in asthmatic patients. J. Allergy Clin. Immunol. 2019, 143, 305–315. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Wen, S.; Yuan, G.; Li, C.; Xiong, Y.; Zhong, X.; Li, X. High cellulose dietary intake relieves asthma inflammation through the intestinal microbiome in a mouse model. PLoS ONE 2022, 17, e0263762. [Google Scholar] [CrossRef]
- Forno, E.; Lescher, R.; Strunk, R.; Weiss, S.; Fuhlbrigge, A.; Celedón, J.C. Decreased response to inhaled steroids in overweight and obese asthmatic children. J. Allergy Clin. Immunol. 2011, 127, 741–749. [Google Scholar] [CrossRef]
- Park, C.S.; Bang, B.R.; Kwon, H.S.; Moon, K.A.; Kim, T.B.; Lee, K.Y.; Moon, H.B.; Cho, Y.S. Metformin reduces airway inflammation and remodeling via activation of AMP-activated protein kinase. Biochem. Pharmacol. 2012, 84, 1660–1670. [Google Scholar] [CrossRef]
- Ma, W.; Jin, Q.; Guo, H.; Han, X.; Xu, L.; Lu, S.; Wu, C. Metformin Ameliorates Inflammation and Airway Remodeling of Experimental Allergic Asthma in Mice by Restoring AMPKα Activity. Front. Pharmacol. 2022, 13, 780148. [Google Scholar] [CrossRef]
- McCreight, L.J.; Bailey, C.J.; Pearson, E.R. Metformin and the gastrointestinal tract. Diabetologia 2016, 59, 426–435. [Google Scholar] [CrossRef]
- Rodriguez, J.; Hiel, S.; Delzenne, N.M. Metformin: Old friend, new ways of action-implication of the gut microbiome? Curr. Opin Clin. Nutr Metab. Care 2018, 21, 294–301. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Metformin Is Associated With Higher Relative Abundance of Mucin-Degrading Akkermansia muciniphila and Several Short-Chain Fatty Acid-Producing Microbiota in the Gut. Diabetes Care 2017, 40, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.D.; Keet, C.A.; Fawzy, A.; Segal, J.B.; Brigham, E.P.; McCormack, M.C. Association of Metformin Initiation and Risk of Asthma Exacerbation. A Claims-based Cohort Study. Ann. Am. Thorac. Soc. 2019, 16, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Erickson, S.R.; Wu, C.H. Metformin use and asthma outcomes among patients with concurrent asthma and diabetes. Respirology 2016, 21, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Calixto, M.C.; Lintomen, L.; André, D.M.; Leiria, L.O.; Ferreira, D.; Lellis-Santos, C.; Anhê, G.F.; Bordin, S.; Landgraf, R.G.; Antunes, E. Metformin attenuates the exacerbation of the allergic eosinophilic inflammation in high fat-diet-induced obesity in mice. PLoS ONE 2013, 8, e76786. [Google Scholar] [CrossRef]
- Guo, Y.; Shi, J.; Wang, Q.; Hong, L.; Chen, M.; Liu, S.; Yuan, X.; Jiang, S. Metformin alleviates allergic airway inflammation and increases Treg cells in obese asthma. J. Cell. Mol. Med. 2021, 25, 2279–2284. [Google Scholar] [CrossRef]
- Gu, C.; Loube, J.; Lee, R.; Bevans-Fonti, S.; Wu, T.D.; Barmine, J.H.; Jun, J.C.; McCormack, M.C.; Hansel, N.N.; Mitzner, W.; et al. Metformin Alleviates Airway Hyperresponsiveness in a Mouse Model of Diet-Induced Obesity. Front. Physiol. 2022, 13, 883275. [Google Scholar] [CrossRef]
- Calco, G.N.; Proskocil, B.J.; Jacoby, D.B.; Fryer, A.D.; Nie, Z. Metformin prevents airway hyperreactivity in rats with dietary obesity. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2021, 321, L1105–L1118. [Google Scholar] [CrossRef]
- Toki, S.; Goleniewska, K.; Reiss, S.; Zhang, J.; Bloodworth, M.H.; Stier, M.T.; Zhou, W.; Newcomb, D.C.; Ware, L.B.; Stanwood, G.D.; et al. Glucagon-like peptide 1 signaling inhibits allergen-induced lung IL-33 release and reduces group 2 innate lymphoid cell cytokine production in vivo. J. Allergy Clin. Immunol. 2018, 142, 1515–1528.e1518. [Google Scholar] [CrossRef]
- Toki, S.; Newcomb, D.C.; Printz, R.L.; Cahill, K.N.; Boyd, K.L.; Niswender, K.D.; Peebles, R.S., Jr. Glucagon-like peptide-1 receptor agonist inhibits aeroallergen-induced activation of ILC2 and neutrophilic airway inflammation in obese mice. Allergy 2021, 76, 3433–3445. [Google Scholar] [CrossRef]
- Hur, J.; Kang, J.Y.; Kim, Y.K.; Lee, S.Y.; Lee, H.Y. Glucagon-like peptide 1 receptor (GLP-1R) agonist relieved asthmatic airway inflammation via suppression of NLRP3 inflammasome activation in obese asthma mice model. Pulm. Pharmacol. Ther. 2021, 67, 102003. [Google Scholar] [CrossRef] [PubMed]
- Foer, D.; Beeler, P.E.; Cui, J.; Karlson, E.W.; Bates, D.W.; Cahill, K.N. Asthma Exacerbations in Patients with Type 2 Diabetes and Asthma on Glucagon-like Peptide-1 Receptor Agonists. Am. J. Respir. Crit. Care Med. 2021, 203, 831–840. [Google Scholar] [CrossRef] [PubMed]


In Vivo (Asthmatic Murine Studies) | In Vivo (Obese Asthmatic Murine Studies) | Retrospective Studies (Concurrent Asthma and Diabetes) | |
|---|---|---|---|
| Metformin | ↓ Lung infiltration of eosinophils, peribronchial fibrosis, smooth muscle layer thickening, mucus secretion, cytokine secretion in BALF | ↓ TNF-α-induced inflammation and NF-κB-mediated iNOs expression in lungs ↓ Airway hyper-responsiveness, weight gain, fat gain, insulin levels data | ↓ Asthma exacerbation ↓ Asthma-related hospitalization and ED visit |
| GLP-1R agonists | ↓ IL-33 expression and secretion ↓ ILC2 activation ↓ Chemokines and cytokines ↓ Eosinophilia, mucus secretion, airway hyper-reactivity | ↓ IL-33 and TSLP secretion ↓ ILC2 activation ↓ Chemokines and cytokines ↓ Eosinophils and neutrophils ↓ Airway hyper-reactivity ↓ ICAM-1 expression in lung epithelial and endothelial cells | ↓ Asthma exacerbation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shailesh, H.; Bhat, A.A.; Janahi, I.A. Obesity-Associated Non-T2 Mechanisms in Obese Asthmatic Individuals. Biomedicines 2023, 11, 2797. https://doi.org/10.3390/biomedicines11102797
Shailesh H, Bhat AA, Janahi IA. Obesity-Associated Non-T2 Mechanisms in Obese Asthmatic Individuals. Biomedicines. 2023; 11(10):2797. https://doi.org/10.3390/biomedicines11102797
Chicago/Turabian StyleShailesh, Harshita, Ajaz A. Bhat, and Ibrahim A. Janahi. 2023. "Obesity-Associated Non-T2 Mechanisms in Obese Asthmatic Individuals" Biomedicines 11, no. 10: 2797. https://doi.org/10.3390/biomedicines11102797