
Epigenome-Wide Association Studies of the Fractional Exhaled Nitric Oxide and Bronchodilator Drug Response in Moderate-to-Severe Pediatric Asthma

 , ,
, ,  , , , , ,
, , , , ,  , ,
, ,  , , , , and add
Show full author list
, , , , and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Measurement of FeNO and BDR
2.3. Genome-Wide Methylation Assessment and Quality Control
2.4. Estimation of Cell-Type Heterogeneity and Potential Confounders
2.5. Epigenome-Wide Association Study
2.6. Cross-Tissue Validation
2.7. Differentially Methylated Regions
2.8. Enrichment Analyses
3. Results
3.1. Quality Control of DNAm Data and Assessment of Global DNAm Patterns
3.2. Study Population
3.3. Epigenome-Wide Association Study
3.4. Differentially Methylated Regions
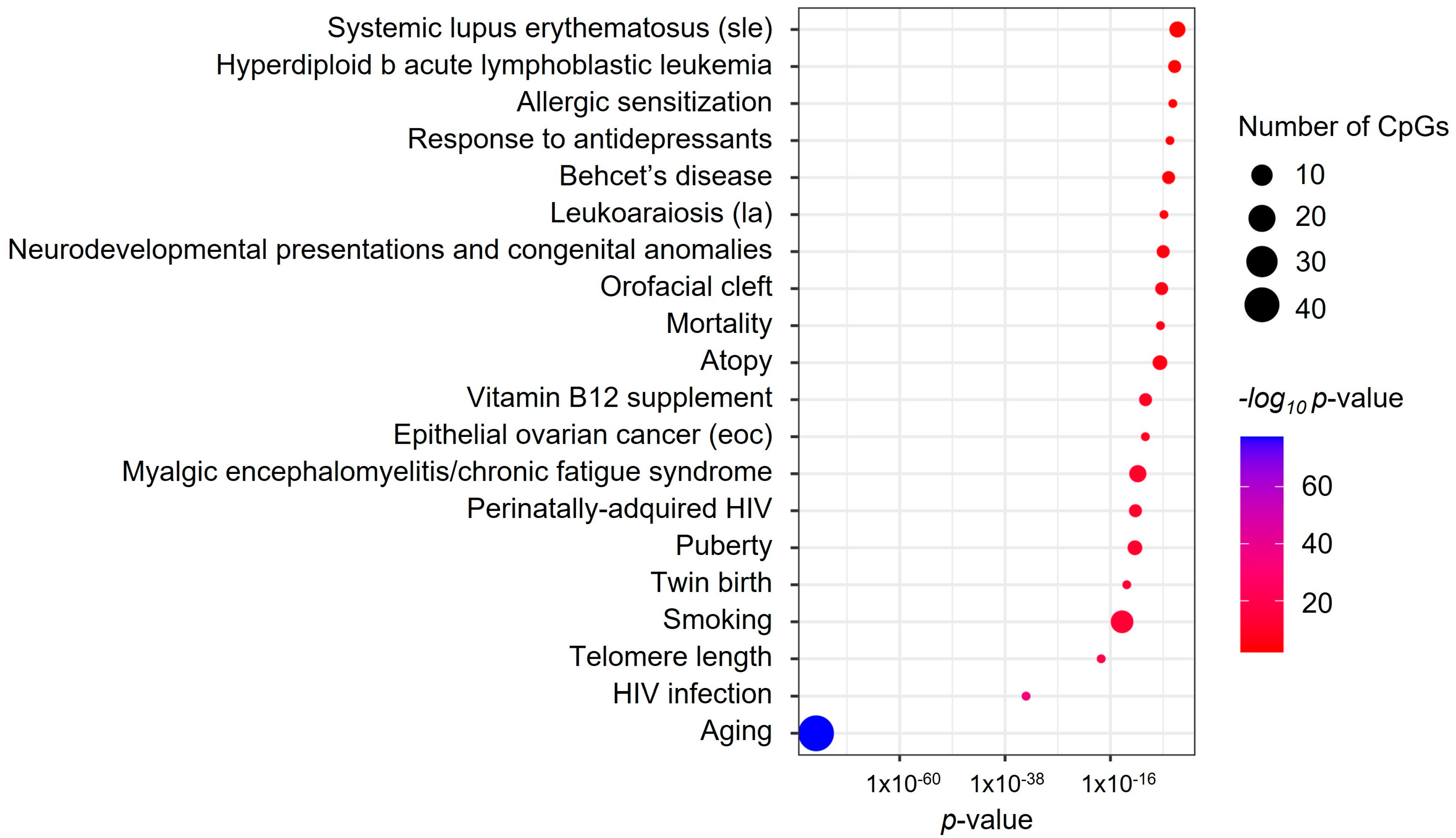
3.5. Enrichment Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Masoli, M.; Fabian, D.; Holt, S.; Beasley, R. The Global Burden of Asthma: Executive Summary of the GINA Dissemination Committee Report. Allergy 2004, 59, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Global Strategy for Asthma Management and Prevention; Global Initiative for Asthma: Fontana, WI, USA, 2022.
- Licari, A.; Brambilla, I.; Marseglia, A.; De Filippo, M.; Paganelli, V.; Marseglia, L.L. Difficult vs. Severe Asthma: Definition and Limits of Asthma Control in the Pediatric Population. Front. Pediatr. 2018, 1, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sordillo, J.E.; Kelly, R.S.; Lutz, S.M.; Lasky-Su, J.; Wu, A.C. Pharmacogenetics of Bronchodilator Response: Future Directions. Curr. Allergy Asthma Rep. 2021, 21, 47. [Google Scholar] [CrossRef] [PubMed]
- Burchard, E.G.; Avila, P.C.; Nazario, S.; Casal, J.; Torres, A.; Rodriguez-Santana, J.R.; Toscano, M.; Sylvia, J.S.; Alioto, M.E.; Salazar, M.; et al. Lower Bronchodilator Responsiveness in Puerto Rican than in Mexican Subjects with Asthma. Am. J. Respir. Crit. Care Med. 2004, 169, 386–392. [Google Scholar] [CrossRef]
- Lund, M.B.; Kongerud, J.; Nystad, W.; Boe, J.; Harris, J.R. Genetic and Environmental Effects on Exhaled Nitric Oxide and Airway Responsiveness in a Population-Based Sample of Twins. Eur. Respir. J. 2007, 29, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Pacheco, N.; Pino-Yanes, M.; Flores, C. Genomic Predictors of Asthma Phenotypes and Treatment Response. Front. Pediatr. 2019, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Ober, C. Asthma Genetics in the Post-GWAS Era. Ann. Am. Thorac. Soc. 2016, 13 (Suppl. 1), S85–S90. [Google Scholar] [CrossRef]
- Cardenas, A.; Sordillo, J.E.; Rifas-Shiman, S.L.; Chung, W.; Liang, L.; Coull, B.A.; Hivert, M.F.; Lai, P.S.; Forno, E.; Celedón, J.C.; et al. The Nasal Methylome as a Biomarker of Asthma and Airway Inflammation in Children. Nat. Commun. 2019, 10, 3095. [Google Scholar] [CrossRef] [Green Version]
- Edris, A.; den Dekker, H.T.; Melén, E.; Lahousse, L. Epigenome-Wide Association Studies in Asthma: A Systematic Review. Clin. Exp. Allergy 2019, 49, 953–968. [Google Scholar] [CrossRef]
- Breton, C.V.; Byun, H.M.; Wang, X.; Salam, M.T.; Siegmund, K.; Gilliland, F.D. DNA Methylation in the Arginase-Nitric Oxide Synthase Pathway Is Associated with Exhaled Nitric Oxide in Children with Asthma. Am. J. Respir. Crit. Care Med. 2011, 184, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Ji, N.; Fang, M.; Baptista, A.; Cepeda, C.; Greenberg, M.; Mincey, I.C.; Ohman-Strickland, P.; Haynes, F.; Fiedler, N.; Kipen, H.M.; et al. Exposure to Traffic-Related Air Pollution and Changes in Exhaled Nitric Oxide and DNA Methylation in Arginase and Nitric Oxide Synthase in Children with Asthma. Environ. Health 2021, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, M.I.; Neerincx, A.H.; Vijverberg, S.J.H.; Hashimoto, S.; Brinkman, P.; Gorenjak, M.; Toncheva, A.A.; Harner, S.; Brandstetter, S.; Wolff, C.; et al. A System Pharmacology Multi-Omics Approach toward Uncontrolled Pediatric Asthma. J. Pers. Med. 2021, 11, 484. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Child Growth Standards. Dev. Med. Child Neurol. 2009, 51, 1002. [Google Scholar] [CrossRef]
- Miller, M.R.; Hankinson, J.; Brusasco, V.; Burgos, F.; Casaburi, R.; Coates, A.; Crapo, R.; Enright, P.; Van Der Grinten, C.P.M.; Gustafsson, P.; et al. Standardisation of Spirometry. Eur. Respir. J. 2005, 26, 319–338. [Google Scholar] [CrossRef] [Green Version]
- American Thoracic Society; European Respiratory Society. ATS/ERS Recommendations for Standardized Procedures for the Online and Offline Measurement of Exhaled Lower Respiratory Nitric Oxide and Nasal Nitric Oxide. Am. J. Respir. Crit. Care Med. 2005, 171, 912–930. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Niu, L.; Li, L.; Taylor, J.A. ENmix: A Novel Background Correction Method for Illumina HumanMethylation450 BeadChip. Nucleic Acids Res. 2016, 44, e20. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.r-project.org/ (accessed on 10 May 2022).
- Niu, L.; Xu, Z.; Taylor, J.A. RCP: A Novel Probe Design Bias Correction Method for Illumina Methylation BeadChip. Bioinformatics 2016, 32, 2659–2663. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Langie, S.A.S.; De Boever, P.; Taylor, J.A.; Niu, L. RELIC: A Novel Dye-Bias Correction Method for Illumina Methylation BeadChip. BMC Genom. 2017, 18, 4. [Google Scholar] [CrossRef] [Green Version]
- Heiss, J.A.; Just, A.C. Identifying Mislabeled and Contaminated DNA Methylation Microarray Data: An Extended Quality Control Toolset with Examples from GEO. Clin. Epigenetics 2018, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Pidsley, R.; Zotenko, E.; Peters, T.J.; Lawrence, M.G.; Risbridger, G.P.; Molloy, P.; Van Djik, S.; Muhlhausler, B.; Stirzaker, C.; Clark, S.J. Critical Evaluation of the Illumina MethylationEPIC BeadChip Microarray for Whole-Genome DNA Methylation Profiling. Genome Biol. 2016, 17, 208. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.C.; Kuan, P.F. A Guide to Illumina BeadChip Data Analysis. Methods Mol. Biol. 2018, 1708, 303–330. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, E.; Zaitlen, N.; Baran, Y.; Eng, C.; Hu, D.; Galanter, J.; Oh, S.; Burchard, E.G.; Eskin, E.; Zou, J.; et al. Sparse PCA Corrects for Cell Type Heterogeneity in Epigenome-Wide Association Studies. Nat. Methods 2016, 13, 443–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Mansell, G.; Gorrie-Stone, T.J.; Bao, Y.; Kumari, M.; Schalkwyk, L.S.; Mill, J.; Hannon, E. Guidance for DNA Methylation Studies: Statistical Insights from the Illumina EPIC Array. BMC Genom. 2019, 20, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, K.D. IlluminaHumanMethylationEPICanno.Ilm10b4.Hg19: Annotation for Illumina’s EPIC Methylation Arrays. R Packag. Version 0.6.0 2017. [Google Scholar] [CrossRef]
- McLean, C.Y.; Bristor, D.; Hiller, M.; Clarke, S.L.; Schaar, B.T.; Lowe, C.B.; Wenger, A.M.; Bejerano, G. GREAT Improves Functional Interpretation of Cis-Regulatory Regions. Nat. Biotechnol. 2010, 28, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.S.; Schwartz, D.A.; Yang, I.V.; Kechris, K.J. Comb-p: Software for Combining, Analyzing, Grouping and Correcting Spatially Correlated P-Values. Bioinformatics 2012, 28, 2986–2988. [Google Scholar] [CrossRef] [Green Version]
- Peters, T.J.; Buckley, M.J.; Statham, A.L.; Pidsley, R.; Samaras, K.; Lord, R.V.; Clark, S.J.; Molloy, P.L. De Novo Identification of Differentially Methylated Regions in the Human Genome. Epigenetics Chromatin 2015, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.; Yang, F.; Li, M.; Ma, Y.; Zhao, W.; Wang, G.; Li, Z.; Zheng, X.; Zou, D.; Zong, W.; et al. EWAS Open Platform: Integrated Data, Knowledge and Toolkit for Epigenome-Wide Association Study. Nucleic Acids Res. 2022, 50, D1004–D1009. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, N.U.; Guntur, V.P.; Newcomb, D.C.; Wechsler, M.E. Sex and Gender in Asthma. Eur. Respir. Rev. 2021, 30, 210067. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.R.; Boulet, L.P.; Lavoie, K.L.; Raherison-Semjen, C.; Singh, D. Personalized Treatment of Asthma: The Importance of Sex and Gender Differences. J. Allergy Clin. Immunol. Pract. 2022, 10, 963–971.e3. [Google Scholar] [CrossRef] [PubMed]
- Nolin, J.D.; Lai, Y.; Ogden, H.L.; Manicone, A.M.; Murphy, R.C.; An, D.; Frevert, C.W.; Ghomashchi, F.; Naika, G.S.; Gelb, M.H.; et al. Secreted PLA2 Group X Orchestrates Innate and Adaptive Immune Responses to Inhaled Allergen. JCI Insight 2017, 2, e94929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallstrand, T.S.; Chi, E.Y.; Singer, A.G.; Gelb, M.H.; Henderson, W.R. Secreted Phospholipase A 2 Group X Overexpression in Asthma and Bronchial Hyperresponsiveness. Am. J. Respir. Crit. Care Med. 2017, 176, 1072–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouault, M.; Le Calvez, C.; Boilard, E.; Surrel, F.; Singer, A.; Ghomashchi, F.; Bezzine, S.; Scarzello, S.; Bollinger, J.; Gelb, M.H.; et al. Recombinant Production and Properties of Binding of the Full Set of Mouse Secreted Phospholipases A 2 to the Mouse M-Type Receptor. Biochemistry 2007, 46, 1647–1662. [Google Scholar] [CrossRef] [PubMed]
- Revez, J.N.M.A. The Role of the Interleukin-6 Pathway in Asthma. Ph.D. Thesis, School of Medicine, The University of Queensland, Brisbane, Australia, 2018. [Google Scholar]
- Rincon, M.; Irvin, C.G. Role of IL-6 in Asthma and Other Inflammatory Pulmonary Diseases. Int. J. Biol. Sci. 2012, 8, 1281–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilmarinen, P.; Tuomisto, L.E.; Niemela, O.; Danielsson, J.; Naanpaa, J.; Kankaanranta, T.; Kankaanranta, H. Comorbidities and Elevated IL-6 Associate with Negative Outcome in Adult-Onset Asthma. Eur. Respir. J. 2016, 48, 1052–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, E.S.; De Sanctis, G.T.; Boyce, J.; Maclean, J.A.; Jiao, A.; Green, F.H.Y.; Grasemann, H.; Faunce, D.; Fitzmaurice, G.; Shi, G.P.; et al. The Transcription Factor Early Growth-Response Factor 1 Modulates Tumor Necrosis Factor- α, Immunoglobulin E, and Airway Responsiveness in Mice. Am. J. Respir. Crit. Care Med. 2012, 163, 778–785. [Google Scholar] [CrossRef]
- Baron, V.T.; Pio, R.; Jia, Z.; Mercola, D. Early Growth Response 3 Regulates Genes of Inflammation and Directly Activates IL6 and IL8 Expression in Prostate Cancer. Br. J. Cancer 2015, 112, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Omodho, B.; Miao, T.; Symonds, A.L.J.; Singh, R.; Li, S.; Wang, P. Transcription Factors Early Growth Response Gene (Egr) 2 and 3 Control Inflammatory Responses of Tolerant T Cells. Immun. Inflamm. Dis. 2018, 6, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Jakiela, B.; Soja, J.; Sladek, K.; Przybyszowski, M.; Plutecka, H.; Gielicz, A.; Licholai, S.; Aab, A.; Rebane, A.; Bochenek, G. Bronchial Epithelial Cell Transcriptome Shows Endotype Heterogeneity of Asthma in Patients with NSAID-Exacerbated Respiratory Disease. J. Allergy Clin. Immunol. 2022; in press. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chen, D.; Liu, Y.; Yu, J.; Qiao, L.; Lin, S.; Chen, D.; Zhong, G.; Lu, X.; Wang, Y.; et al. Long Noncoding RNA HOXA-AS3 Integrates NF-ΚB Signaling To Regulate Endothelium Inflammation. Mol. Cell. Biol. 2019, 39, e00139-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, M.R.; Bartlett, N.W.; Clarke, D.; Birrell, M.; Belvisi, M.; Johnston, S.L. Targeting the NF-KappaB Pathway in Asthma and Chronic Obstructive Pulmonary Disease. Pharmacol. Ther. 2009, 121, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kambayashi, T.; Deshpande, D.A. The Role of Diacylglycerol Kinases in Allergic Airway Disease. Curr. Opin. Pharmacol. 2020, 51, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Den Dekker, H.T.; Burrows, K.; Felix, J.F.; Salas, L.A.; Nedeljkovic, I.; Yao, J.; Rifas-Shiman, S.L.; Ruiz-Arenas, C.; Amin, N.; Bustamante, M.; et al. Newborn DNA-Methylation, Childhood Lung Function, and the Risks of Asthma and COPD across the Life Course. Eur. Respir. J. 2019, 53, 1801795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera-Luis, E.; Li, A.; Mak, A.C.Y.; Perez-Garcia, J.; Elhawary, J.R.; Oh, S.S.; Hu, D.; Eng, C.; Keys, K.L.; Huntsman, S.; et al. Epigenome-wide Association Study of Lung Function in Latino Children and Youth with Asthma. Clin. Epigenetics 2022, 14, 9. [Google Scholar] [CrossRef]
- Liao, Y.; Li, P.; Wang, Y.; Chen, H.; Ning, S.; Su, D. Construction of Asthma Related Competing Endogenous RNA Network Revealed Novel Long Non-Coding RNAs and Potential New Drugs. Respir. Res. 2020, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Howrylak, J.A.; Moll, M.; Weiss, S.T.; Raby, B.A.; Wu, W.; Xing, E.P. Gene Expression Profiling of Asthma Phenotypes Demonstrates Molecular Signatures of Atopy and Asthma Control. J. Allergy Clin. Immunol. 2016, 137, 1390–1397. [Google Scholar] [CrossRef] [Green Version]
- Zaffini, R.; Gotte, G.; Menegazzi, M. Asthma and Poly(ADP-Ribose) Polymerase Inhibition: A New Therapeutic Approach. Drug Des. Devel. Ther. 2018, 12, 281–293. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wetzel-Strong, S.E.; Hua, X.; Tilley, S.L.; Oswald, E.; Krummel, M.F.; Caron, K.M. Deficiency of RAMP1 Attenuates Antigen-Induced Airway Hyperresponsiveness in Mice. PLoS ONE 2014, 9, e102356. [Google Scholar] [CrossRef] [Green Version]
- Kytikova, O.Y.; Perelman, J.M.; Novgorodtseva, T.P.; Denisenko, Y.K.; Kolosov, V.P.; Antonyuk, M.V.; Gvozdenko, T.A.; Hsun, T.; Huang, W. Peroxisome Proliferator-Activated Receptors as a Therapeutic Target in Asthma. PPAR Res. 2020, 2020, 8906968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Principe, S.; Porsbjerg, C.; Bolm Ditlev, S.; Kjærsgaard Klein, D.; Golebski, K.; Dyhre-Petersen, N.; van Dijk, Y.E.; van Bragt, J.J.M.H.; Dankelman, L.L.H.; Dahlen, S.E.; et al. Treating Severe Asthma: Targeting the IL-5 Pathway. Clin. Exp. Allergy 2021, 51, 992–1005. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.C. Immunoglobulin E Receptor Signaling and Asthma. J. Biol. Chem. 2011, 286, 32891–32897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C.; Cardenas, A.; Rifas-Shiman, S.L.; Hivert, M.F.; Gold, D.R.; Platts-Mills, T.A.; Lin, X.; Oken, E.; Avila, L.; Celedón, J.C.; et al. Epigenetic Age Acceleration Is Associated with Allergy and Asthma in Children in Project Viva. J. Allergy Clin. Immunol. 2019, 143, 2263–2270.e14. [Google Scholar] [CrossRef] [Green Version]
- Stapleton, M.; Howard-Thompson, A.; George, C.; Hoover, R.M.; Self, T.H. Smoking and Asthma. J. Am. Board Fam. Med. 2011, 24, 313–322. [Google Scholar] [CrossRef]
- Teh, A.L.; Pan, H.; Lin, X.; Lim, Y.I.; Patro, C.P.K.; Cheong, C.Y.; Gong, M.; MacIsaac, J.L.; Kwoh, C.K.; Meaney, M.J.; et al. Comparison of Methyl-Capture Sequencing vs. Infinium 450K Methylation Array for Methylome Analysis in Clinical Samples. Epigenetics 2016, 11, 36–48. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Characteristics | n | EWAS of BDR | n | EWAS of FeNO |
|---|---|---|---|---|
| Sex (male) | 121 | 75 (62) | 109 | 67 (61.5) |
| Age (years) | 121 | 12.0 (9.8−14.0) | 109 | 12.1 (9.8−14.0) |
| Ancestry | 121 | 109 | ||
| African | 4 (3.3) | 2 (1.9) | ||
| Asian | 2 (1.7) | 1 (0.9) | ||
| European | 96 (79.3) | 88 (80.7) | ||
| Latin | 9 (7.4) | 8 (7.3) | ||
| Mixed/Other | 10 (8.3) | 10 (9.2) | ||
| Body mass index (z-score) | 121 | 0.5 (−0.3−4.0) | 108 | 0.4 (-0.4−1.3) |
| Uncontrolled asthma | 121 | 77 (63.6) | 109 | 71 (65.1) |
| pre-FEV1 (predicted %) | 121 | 93.6 (82.5−103.2) | 108 | 95.3 (82.7−103.2) |
| pre-FVC (predicted %) | 121 | 99.4 (91.2−108.1) | 108 | 100.3 (91.0−107.6) |
| BDR (%) | 121 | 4.2 (0.6−11.3) | 108 | 4.2 (0.6−10.5) |
| FeNO (ppb) | 120 | 16.0 (9.0−38.0) | 109 | 16.0 (9.0−41.0) |
| SABAs | 103 | 94 (91.3) | 92 | 84 (91.3) |
| LABAs | 103 | 96 (93.2) | 92 | 87 (94.6) |
| ICS | 103 | 103 (100) | 92 | 92 (100) |
| LTRA | 103 | 17 (16.5) | 92 | 15 (16.3) |
| OCS | 103 | 2 (1.9) | 92 | 1 (1.09) |
| Biological therapy † | 103 | 9 (8.7) | 92 | 8 (8.7) |
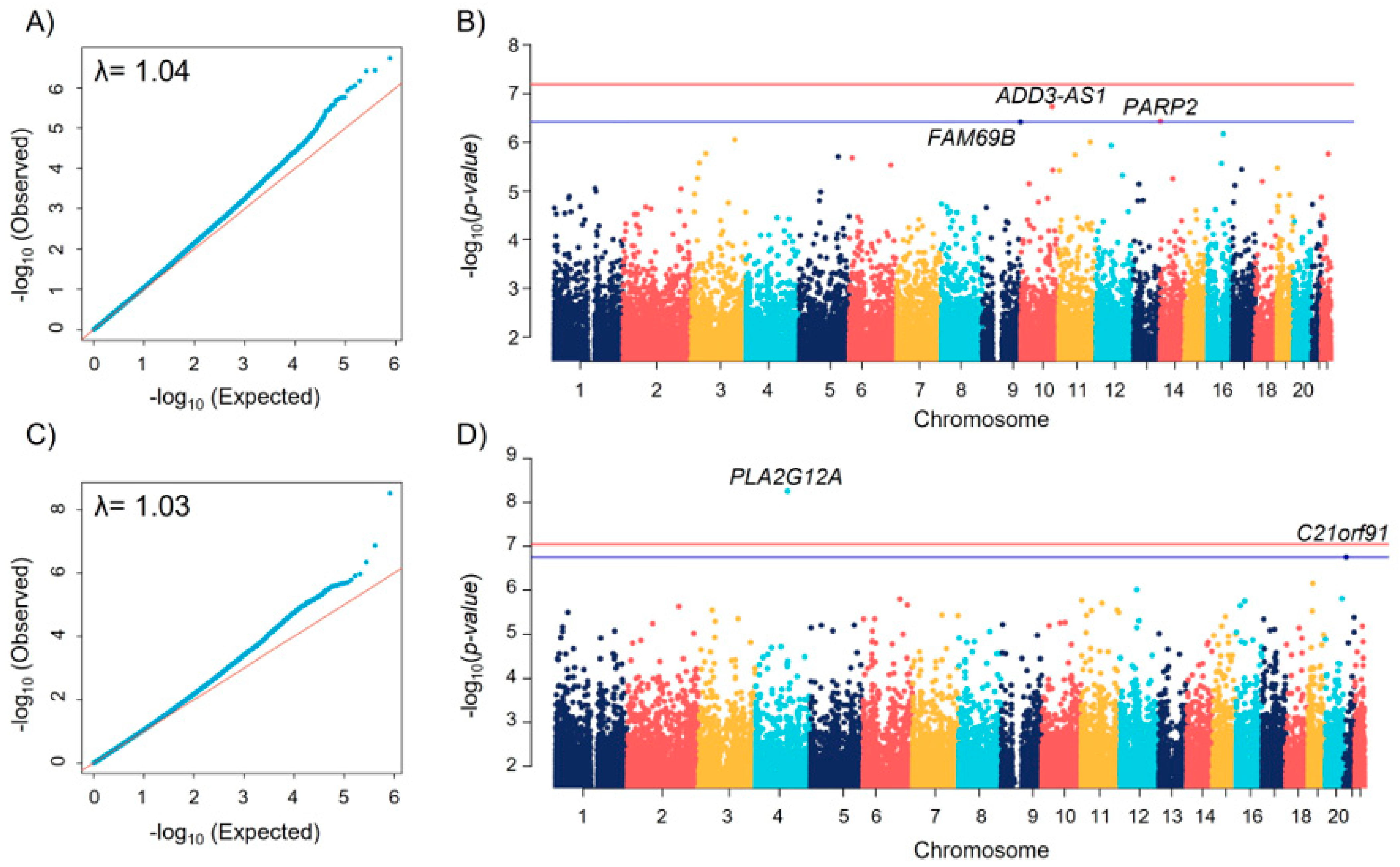
| CpG | Chr | Position † | Gene | Relative Position * | Coef | SE | p-Value | FDR |
|---|---|---|---|---|---|---|---|---|
| Bronchodilator drug response | ||||||||
| cg26203256 | 10 | 111756055 | ADD3-AS1 | First intron | −0.020 | 0.004 | 1.85 × 10−7 | 0.099 |
| cg14985321 | 14 | 20823915 | PARP2 | Tenth exon | −0.010 | 0.002 | 3.71 × 10−7 | 0.099 |
| cg06975120 | 9 | 139606856 | FAM69B | 165 bp upstream | −0.034 | 0.006 | 3.86 × 10−7 | 0.099 |
| Fractional exhaled nitric oxide | ||||||||
| cg12835256 | 4 | 110651671 | PLA2G12A | 439 bp upstream | −0.015 | 0.002 | 2.53 × 10−9 | 0.002 |
| cg19644580 | 21 | 19166676 | C21orf91 | Fourth intron | 0.006 | 0.001 | 1.29 × 10−7 | 0.050 |
| Chr | Start † | End † | Gene | Relative Position * | N° of CpGs | Coefficient | FDR |
|---|---|---|---|---|---|---|---|
| Bronchodilator drug response | |||||||
| 5 | 126408755 | 126409453 | C5orf63 | Promoter, first exon, and intron | 11 | −0.024 | 2.80 × 10−7 |
| 11 | 7597813 | 7598150 | PPFIBP2 | Gene body (alternative transcripts) | 5 | −0.011 | 9.24 × 10−7 |
| 2 | 238767350 | 238767779 | RAMP1 | 671 bp upstream | 4 | −0.022 | 2.67 × 10−6 |
| 6 | 31650734 | 31650849 | LY6G5C | Gene body (alternative transcripts) | 3 | −0.023 | 8.96 × 10−6 |
| Fractional exhaled nitric oxide | |||||||
| 19 | 46998382 | 46999839 | PPP5D1 | Third intron | 16 | −0.004 | 1.48 × 10−18 |
| 8 | 22560921 | 22561950 | EGR3 | 10,622 bp upstream | 7 | −0.005 | 3.15 × 10−17 |
| 15 | 76016128 | 76016334 | ODF3L1 | Promoter and first exon | 6 | −0.003 | 6.66 × 10−14 |
| 7 | 27186553 | 27187559 | HOXA6 | Promoter and gene body | 11 | 0.005 | 2.01 × 10−13 |
| 7 | 27183273 | 27184736 | HOXA-AS3 | First intron | 33 | 0.004 | 2.53 × 10−13 |
| 7 | 27170240 | 27170831 | HOXA4 | Promoter and first exon | 10 | −0.006 | 1.00 × 10−10 |
| 19 | 57742111 | 57742443 | AURKC | Promoter and first exon | 10 | −0.008 | 3.95 × 10−10 |
| 7 | 100844059 | 100844444 | MOGAT3 | First exon | 7 | −0.002 | 4.21 × 10−10 |
| 7 | 27198188 | 27198428 | HOXA7 | 1857 bp upstream | 3 | 0.005 | 1.37 × 10−9 |
| 12 | 41581774 | 41582136 | PDZRN4 | Promoter and first exon | 6 | −0.004 | 1.37 × 10−9 |
| 7 | 79083996 | 79084165 | MAGI2-AS3 | Second intron | 7 | 0.003 | 2.59 × 10−9 |
| 9 | 125137543 | 125137593 | PTGS1 | Second intron | 3 | 0.003 | 7.61 × 10−9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Almeida, M.; Perez-Garcia, J.; Herrera-Luis, E.; Rosa-Baez, C.; Gorenjak, M.; Neerincx, A.H.; Sardón-Prado, O.; Toncheva, A.A.; Harner, S.; Wolff, C.; et al. Epigenome-Wide Association Studies of the Fractional Exhaled Nitric Oxide and Bronchodilator Drug Response in Moderate-to-Severe Pediatric Asthma. Biomedicines 2023, 11, 676. https://doi.org/10.3390/biomedicines11030676
Martin-Almeida M, Perez-Garcia J, Herrera-Luis E, Rosa-Baez C, Gorenjak M, Neerincx AH, Sardón-Prado O, Toncheva AA, Harner S, Wolff C, et al. Epigenome-Wide Association Studies of the Fractional Exhaled Nitric Oxide and Bronchodilator Drug Response in Moderate-to-Severe Pediatric Asthma. Biomedicines. 2023; 11(3):676. https://doi.org/10.3390/biomedicines11030676
Chicago/Turabian StyleMartin-Almeida, Mario, Javier Perez-Garcia, Esther Herrera-Luis, Carlos Rosa-Baez, Mario Gorenjak, Anne H. Neerincx, Olaia Sardón-Prado, Antoaneta A. Toncheva, Susanne Harner, Christine Wolff, and et al. 2023. "Epigenome-Wide Association Studies of the Fractional Exhaled Nitric Oxide and Bronchodilator Drug Response in Moderate-to-Severe Pediatric Asthma" Biomedicines 11, no. 3: 676. https://doi.org/10.3390/biomedicines11030676