
Effects of Atherogenic Factors on Endothelial Cells: Bioinformatics Analysis of Differentially Expressed Genes and Signaling Pathways


Abstract
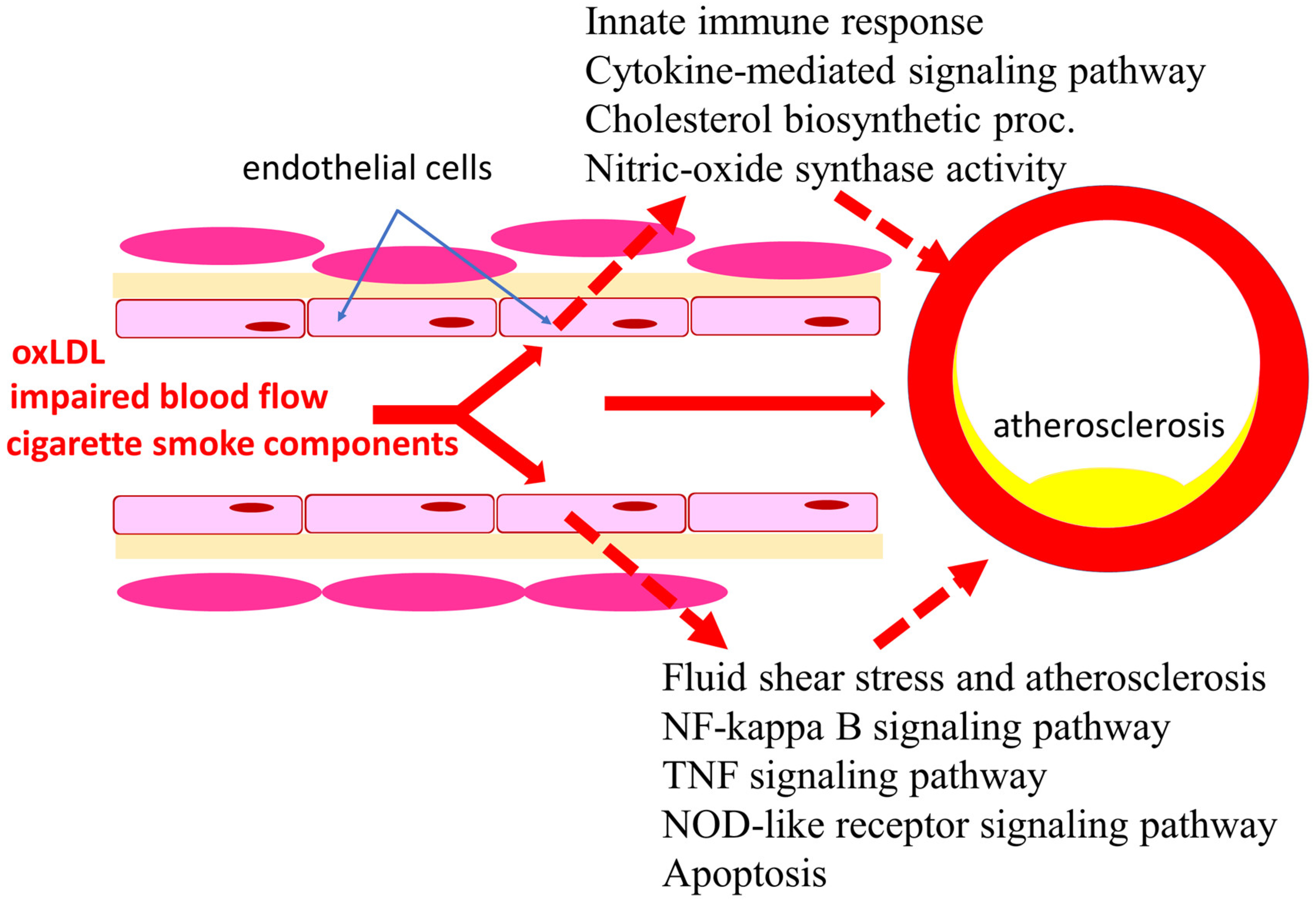
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Differential Expression Analysis
2.3. Gene Ontology Analysis
3. Results
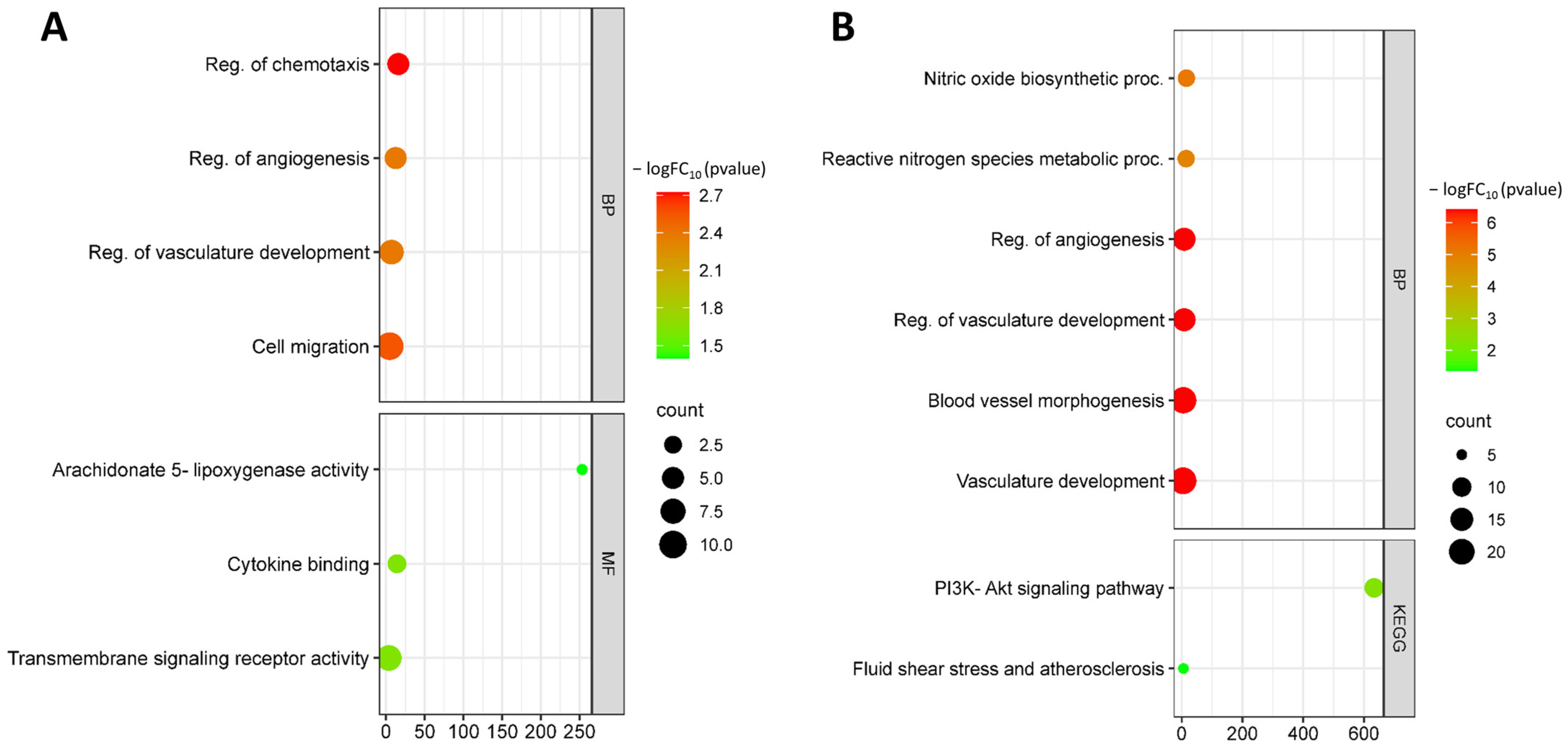
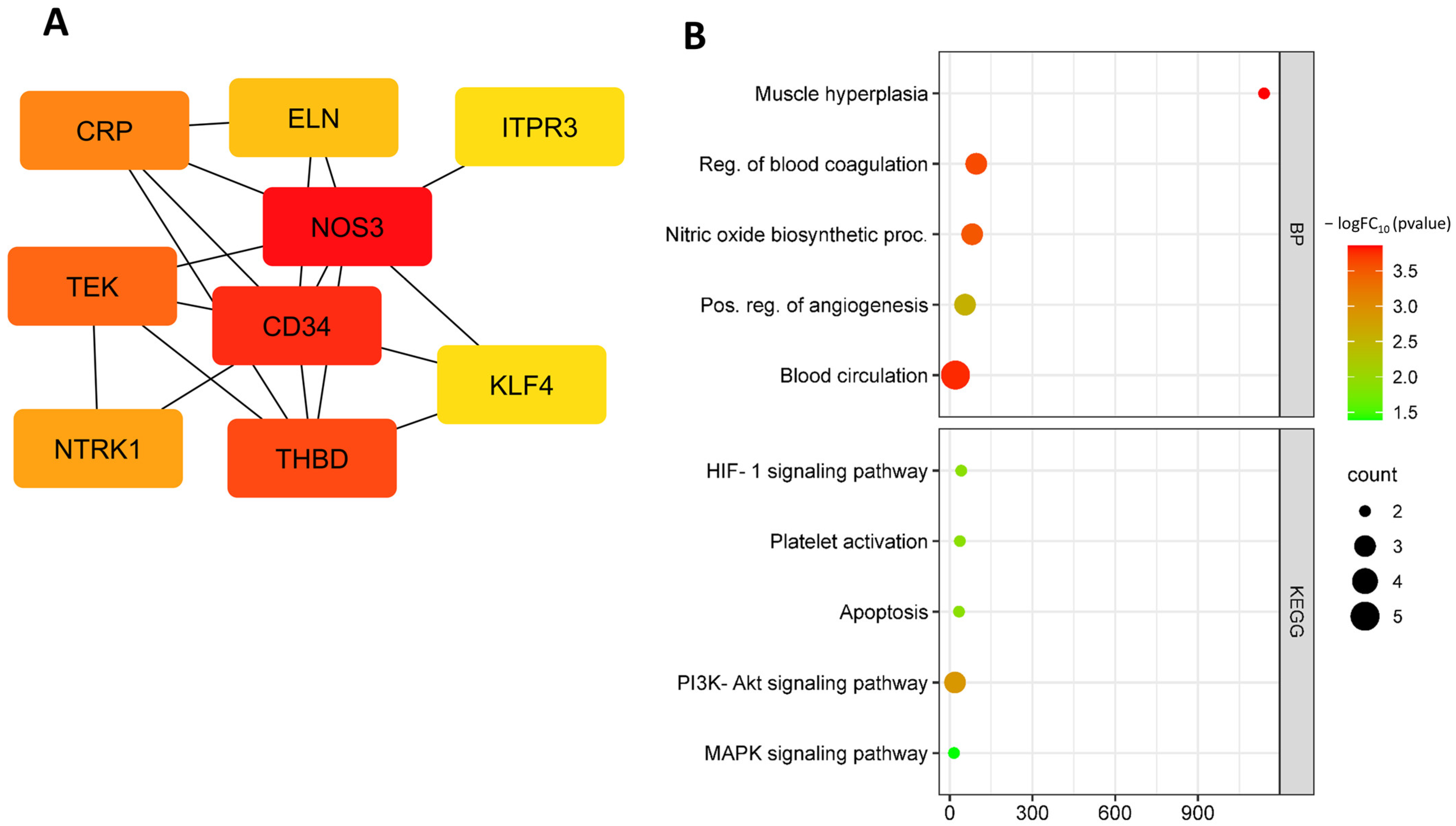
3.1. Identification and Functional Annotation of DEGs in Endothelial Cells during Flow Changes
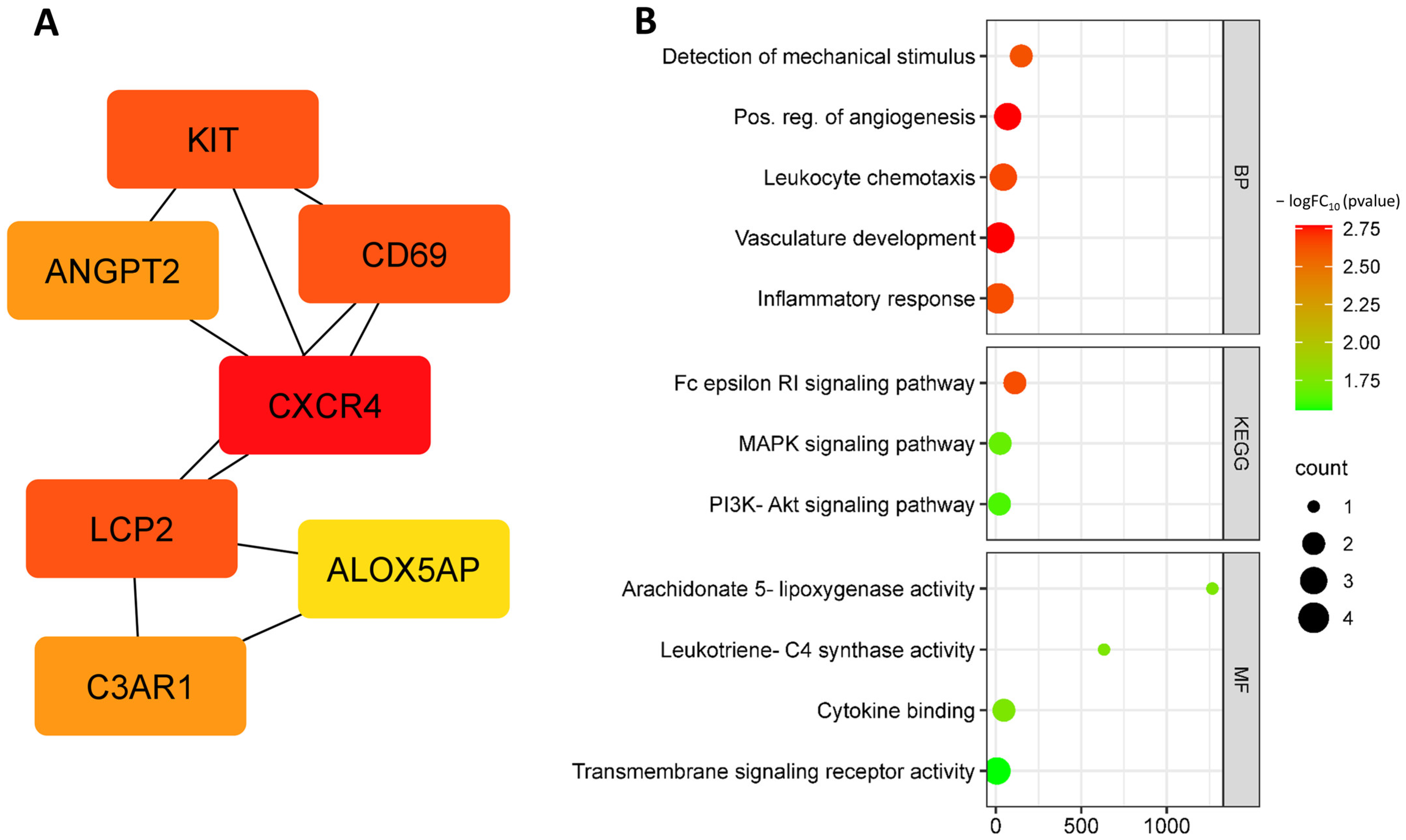
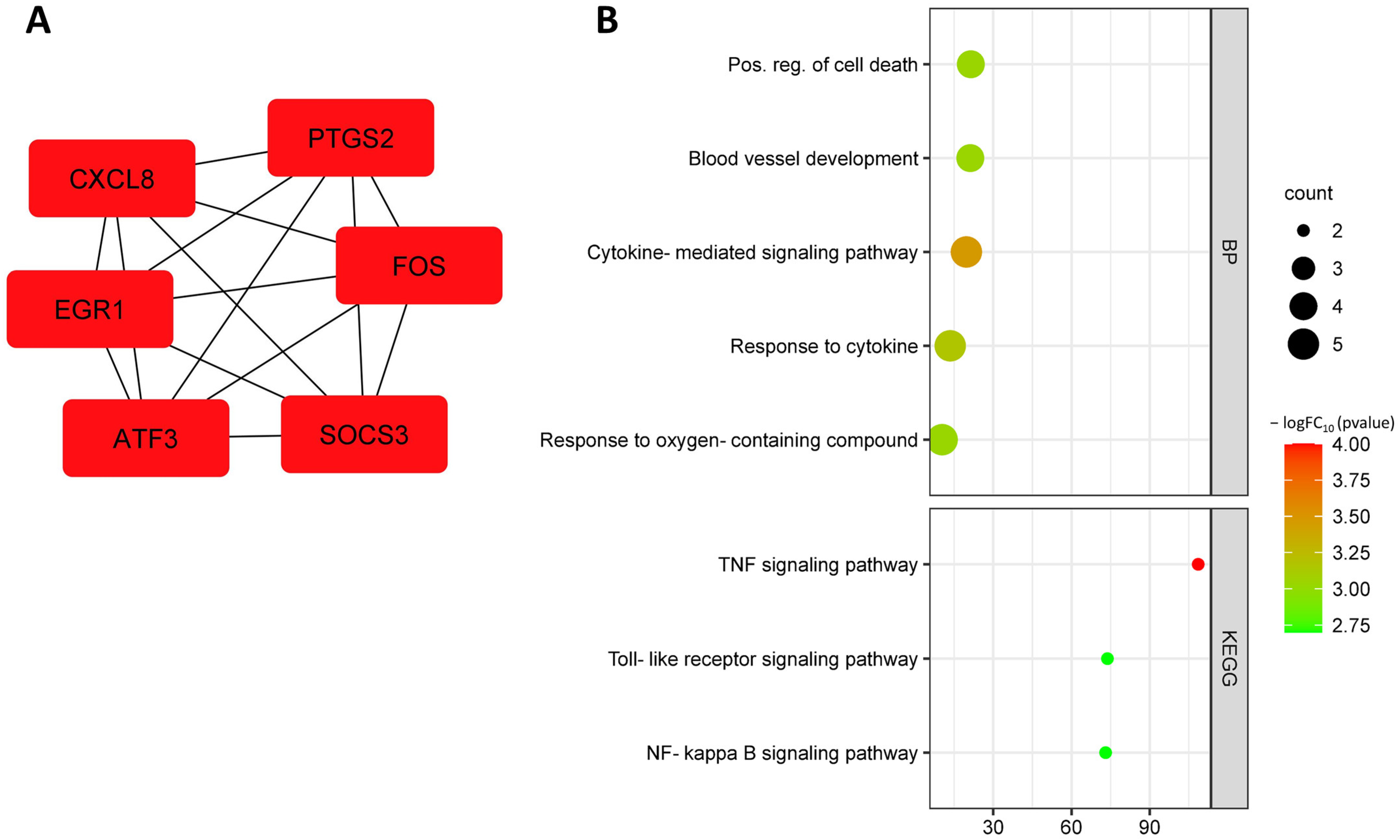
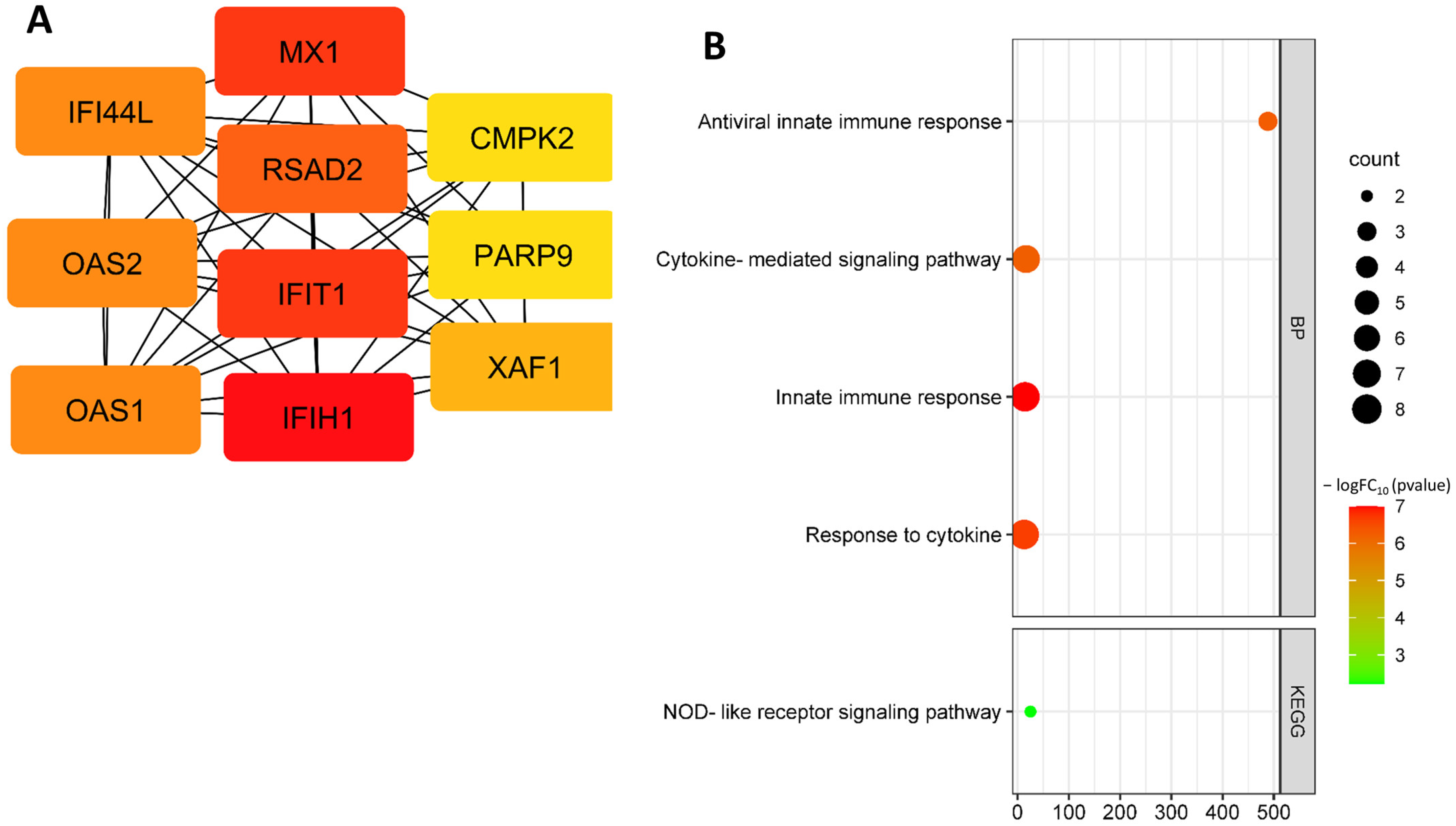
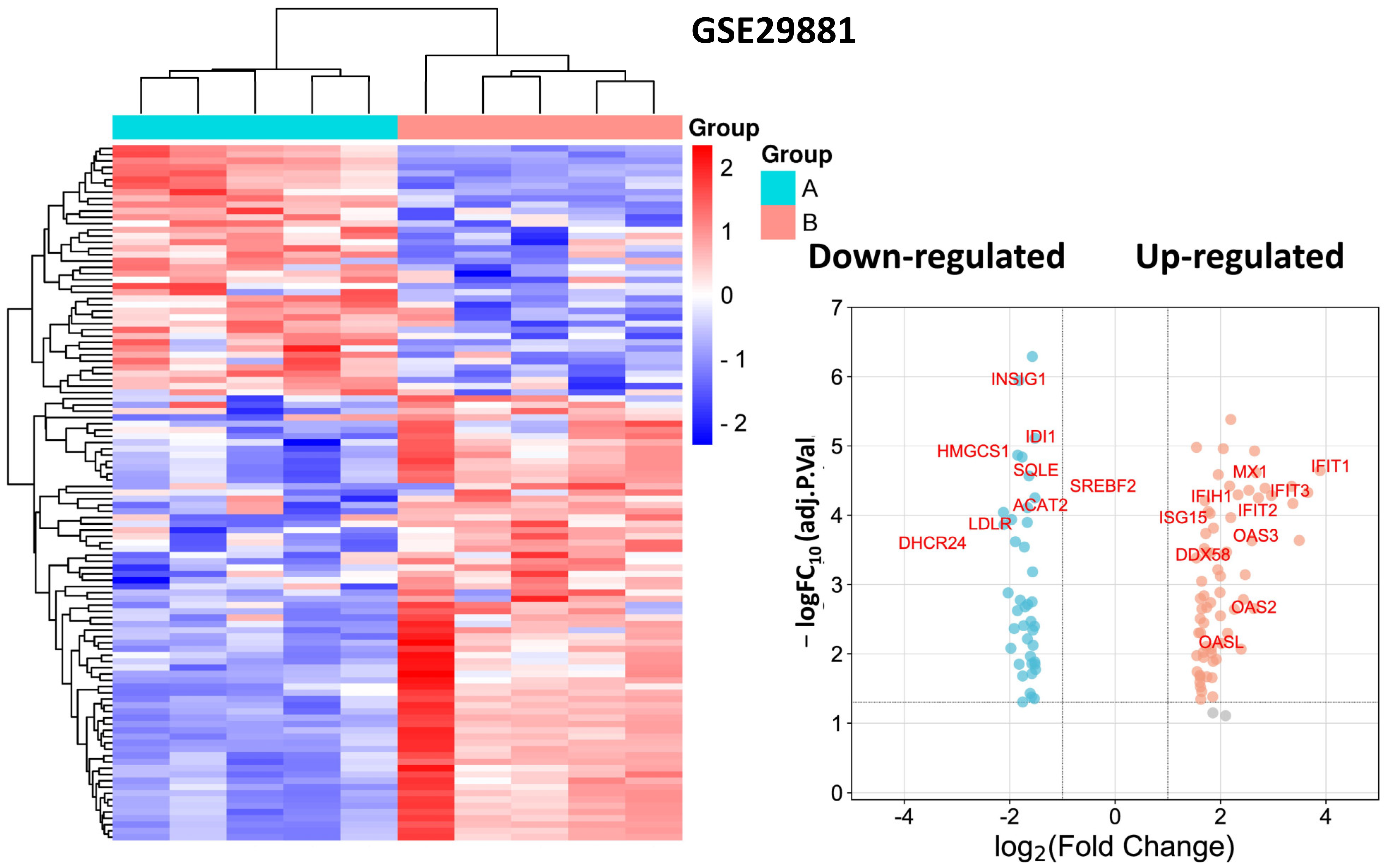
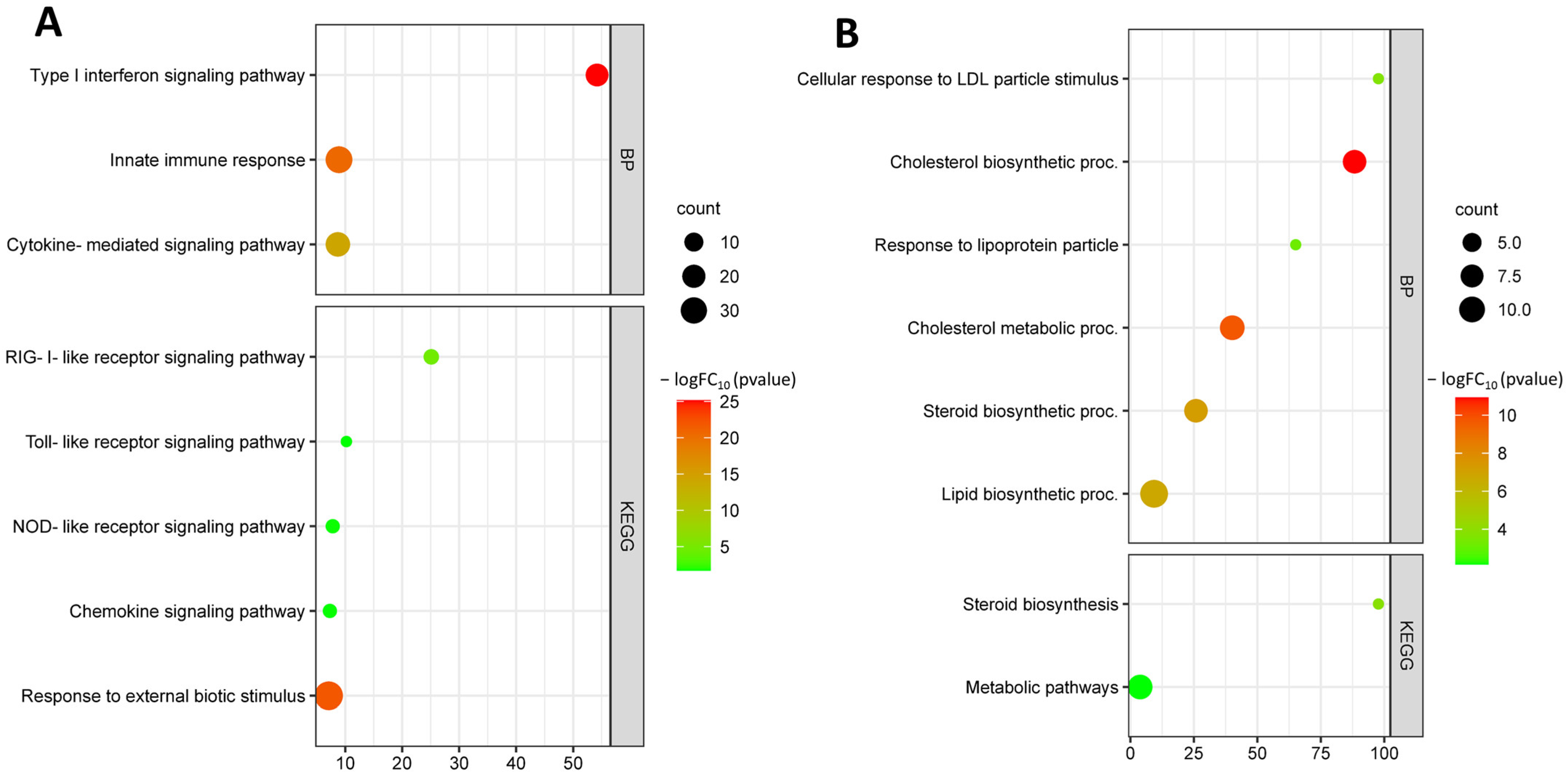
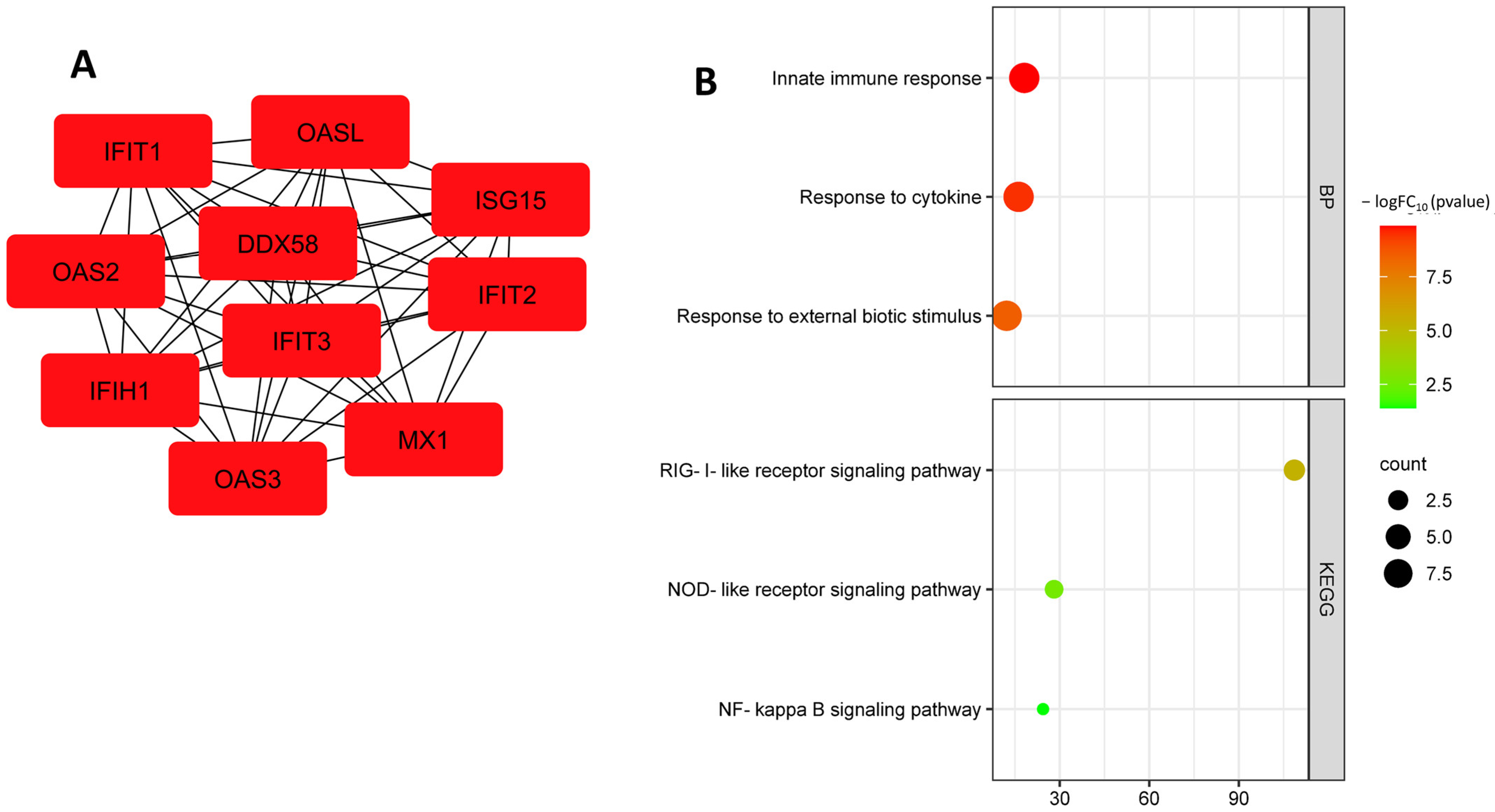
3.2. Identification and Functional Annotation of DEGs in Endothelial Cells in Smoking
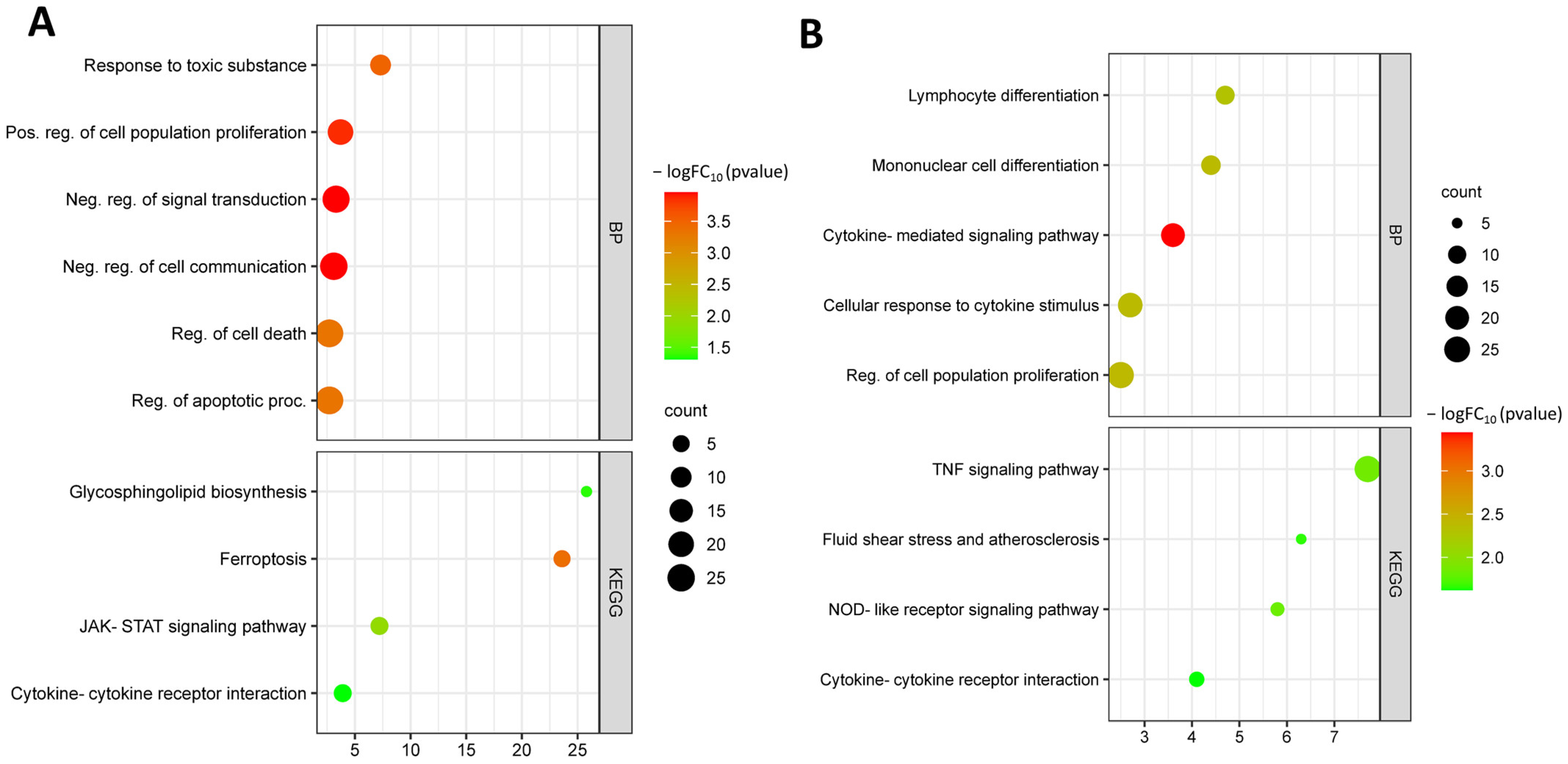
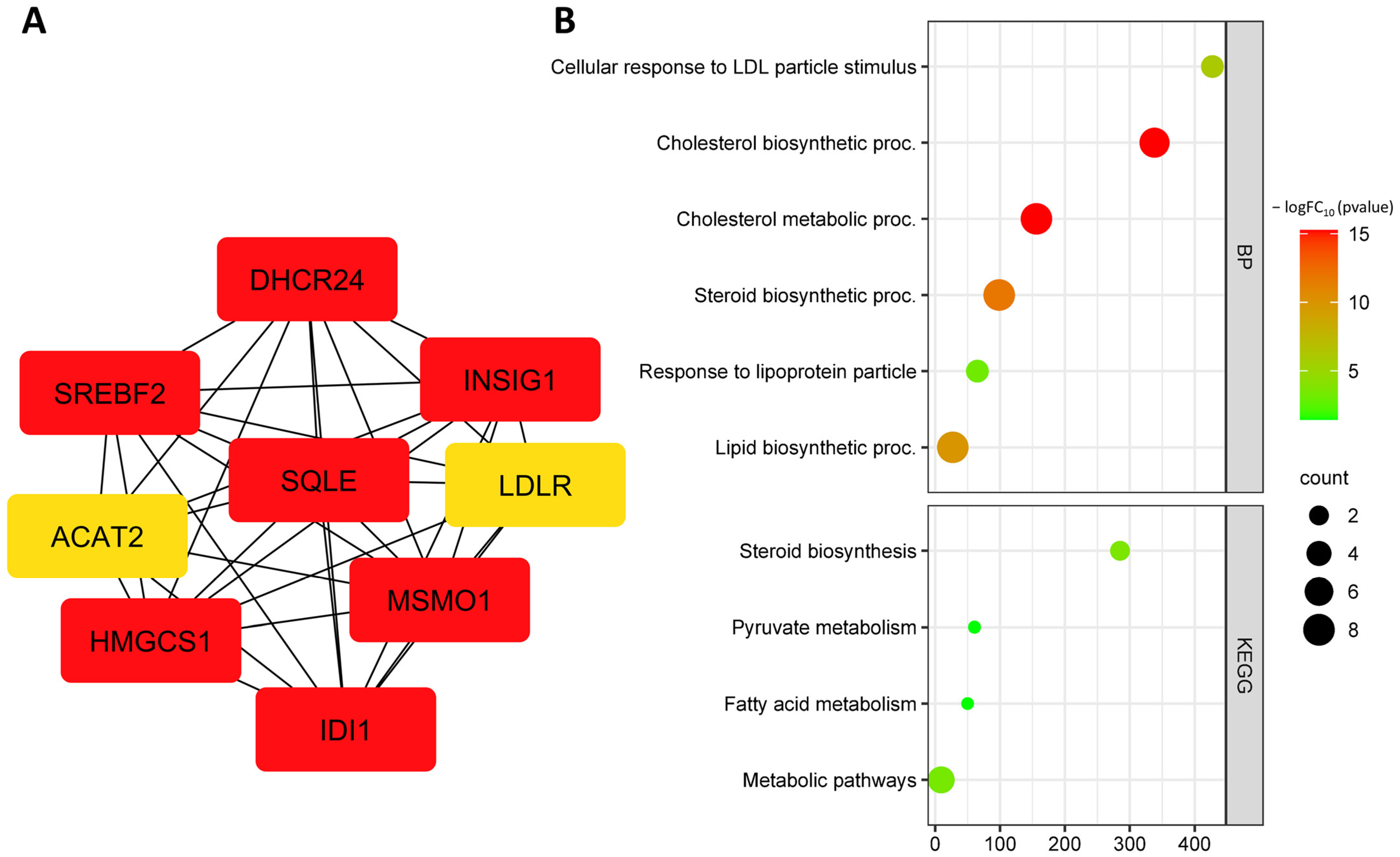
3.3. Identification and Functional Annotation of DEGs in Endothelial Cells upon Exposure to Oxidized LDL
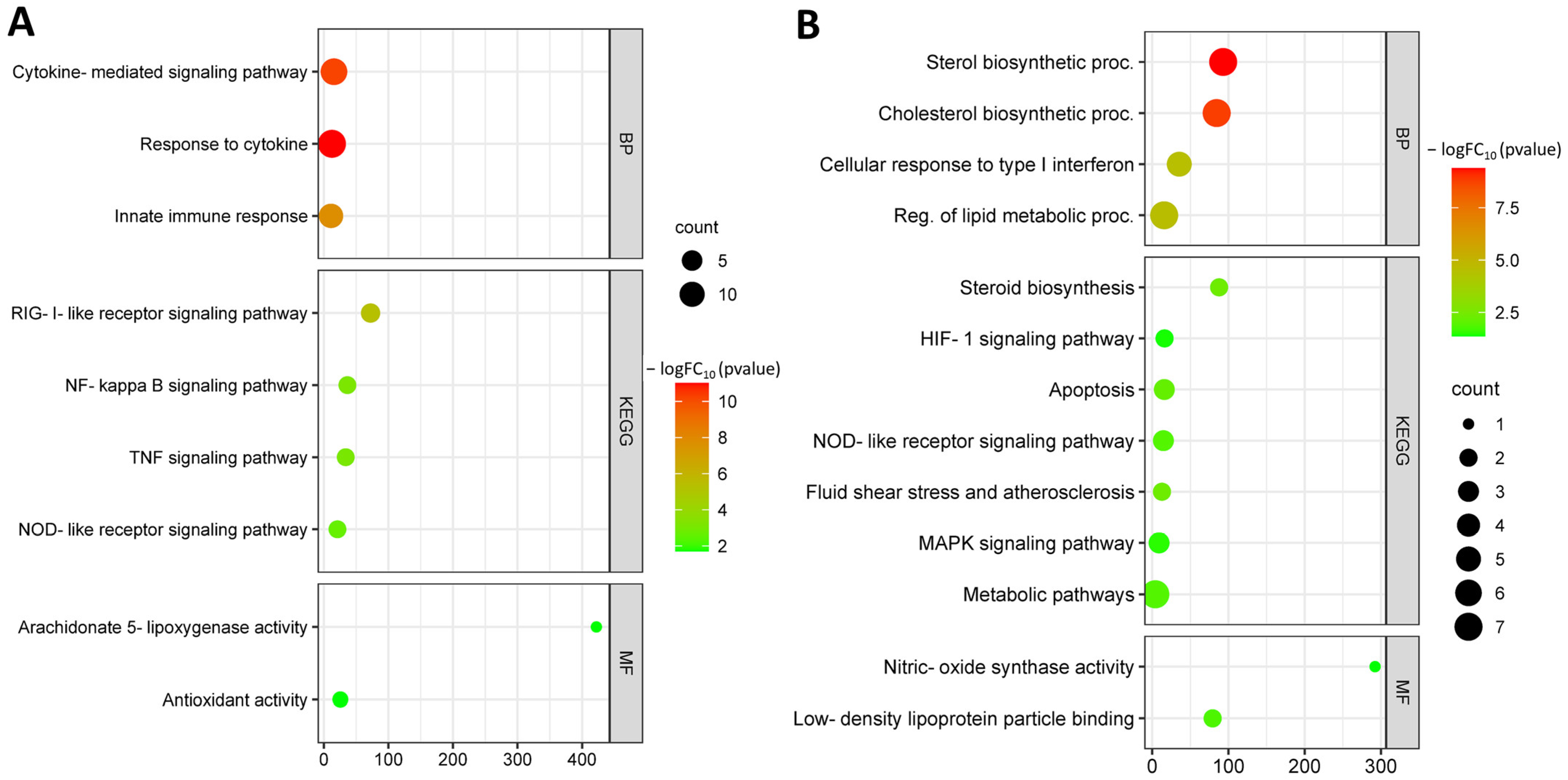
3.4. Identification and Functional Annotation of Common DEGs in Endothelial Cells under Different Risk Factors
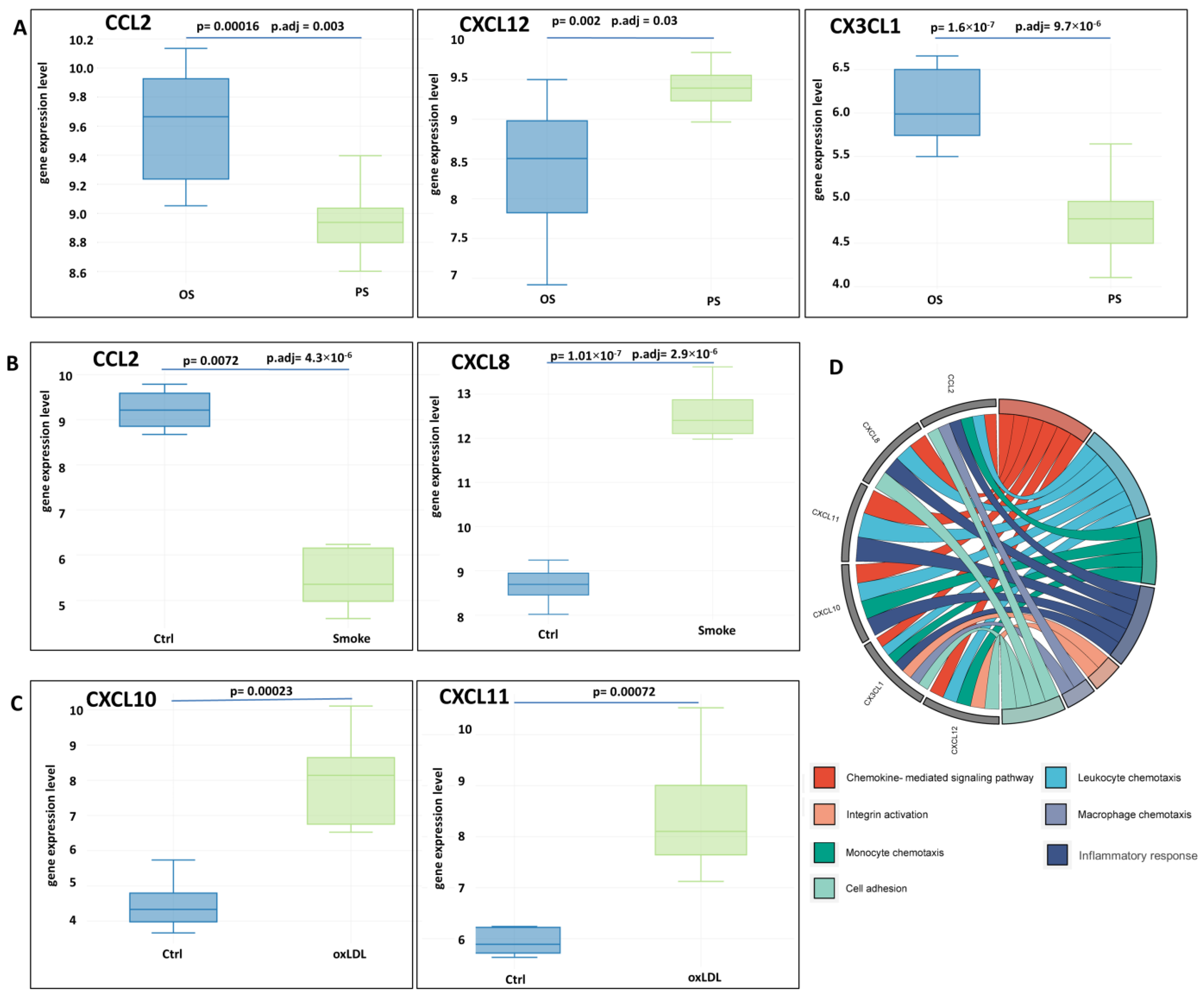
3.5. Identification of Differentially Expressed Genes of Chemokines
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bauersachs, R.; Zeymer, U.; Brière, J.-B.; Marre, C.; Bowrin, K.; Huelsebeck, M. Burden of Coronary Artery Disease and Peripheral Artery Disease: A Literature Review. Cardiovasc. Ther. 2019, 2019, 8295054. [Google Scholar] [CrossRef] [PubMed]
- Aursulesei Onofrei, V.; Ceasovschih, A.; Marcu, D.T.M.; Adam, C.A.; Mitu, O.; Mitu, F. Mortality Risk Assessment in Peripheral Arterial Disease—The Burden of Cardiovascular Risk Factors over the Years: A Single Center’s Experience. Diagnostics 2022, 12, 2499. [Google Scholar] [CrossRef]
- Kalinin, R.E.; Suchkov, I.A.; Chobanyan, A.A.; Nikiforov, A.A.; Shumskaya, E.I. Genetic Predictors of an Unfavorable Course of Obliterating Atherosclerosis of Lower Limb Arteries. IP Pavlov Russ. Med. Biol. Her. 2021, 29, 251–256. [Google Scholar] [CrossRef]
- Kotlyarov, S. Immune Function of Endothelial Cells: Evolutionary Aspects, Molecular Biology and Role in Atherogenesis. Int. J. Mol. Sci. 2022, 23, 9770. [Google Scholar] [CrossRef]
- Kotlyarov, S. Diversity of Lipid Function in Atherogenesis: A Focus on Endothelial Mechanobiology. Int. J. Mol. Sci. 2021, 22, 11545. [Google Scholar] [CrossRef] [PubMed]
- Poredos, P. Endothelial Dysfunction in the Pathogenesis of Atherosclerosis. Int. Angiol. J. Int. Union Angiol. 2002, 21, 109–116. [Google Scholar]
- Golbidi, S.; Edvinsson, L.; Laher, I. Smoking and Endothelial Dysfunction. Curr. Vasc. Pharmacol. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Messner, B.; Bernhard, D. Smoking and Cardiovascular Disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 509–515. [Google Scholar] [CrossRef]
- Jebari-Benslaiman, S.; Galicia-García, U.; Larrea-Sebal, A.; Olaetxea, J.R.; Alloza, I.; Vandenbroeck, K.; Benito-Vicente, A.; Martín, C. Pathophysiology of Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 3346. [Google Scholar] [CrossRef]
- Cunningham, K.S.; Gotlieb, A.I. The Role of Shear Stress in the Pathogenesis of Atherosclerosis. Lab. Investig. 2005, 85, 9–23. [Google Scholar] [CrossRef]
- Warboys, C.M.; Amini, N.; de Luca, A.; Evans, P.C. The Role of Blood Flow in Determining the Sites of Atherosclerotic Plaques. F1000 Med. Rep. 2011, 3, 5. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef]
- Chu, P.-H.; Chen, G.; Kuo, D.; Braisted, J.; Huang, R.; Wang, Y.; Simeonov, A.; Boehm, M.; Gerhold, D.L. Stem Cell-Derived Endothelial Cell Model That Responds to Tobacco Smoke Like Primary Endothelial Cells. Chem. Res. Toxicol. 2020, 33, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Ajami, N.E.; Gupta, S.; Maurya, M.R.; Nguyen, P.; Li, J.Y.-S.; Shyy, J.Y.-J.; Chen, Z.; Chien, S.; Subramaniam, S. Systems Biology Analysis of Longitudinal Functional Response of Endothelial Cells to Shear Stress. Proc. Natl. Acad. Sci. USA 2017, 114, 10990–10995. [Google Scholar] [CrossRef] [PubMed]
- GEO Accession Viewer. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/geo/query/acc.cgi?acc=GSE29881 (accessed on 7 March 2023).
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Assenov, Y.; Ramírez, F.; Schelhorn, S.-E.; Lengauer, T.; Albrecht, M. Computing Topological Parameters of Biological Networks. Bioinform. Oxf. Engl. 2008, 24, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Bader, G.D.; Hogue, C.W.V. An Automated Method for Finding Molecular Complexes in Large Protein Interaction Networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. CytoHubba: Identifying Hub Objects and Sub-Networks from Complex Interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef]
- Zenkova, D.; Kamenev, V.; Sablina, R.; Artyomov, M.; Sergushichev, A. Visual and Interactive Gene Expression Analysis. Phantasus 2018. [Google Scholar] [CrossRef]
- Kanehisa, M. Toward Understanding the Origin and Evolution of Cellular Organisms. Protein Sci. Publ. Protein Soc. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for Taxonomy-Based Analysis of Pathways and Genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, G.W.; Jones, M.R.; Rouillard, A.D.; Kou, Y.; Monteiro, C.D.; Feldmann, A.S.; Hu, K.S.; Ma’ayan, A. GEO2Enrichr: Browser Extension and Server App to Extract Gene Sets from GEO and Analyze Them for Biological Functions. Bioinform. Oxf. Engl. 2015, 31, 3060–3062. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed]
- Carbon, S.; Ireland, A.; Mungall, C.J.; Shu, S.; Marshall, B.; Lewis, S.; AmiGO Hub; Web Presence Working Group. AmiGO: Online Access to Ontology and Annotation Data. Bioinform. Oxf. Engl. 2009, 25, 288–289. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER Version 14: More Genomes, a New PANTHER GO-Slim and Improvements in Enrichment Analysis Tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Gene Ontology Consortium The Gene Ontology Resource: Enriching a GOld Mine. Nucleic Acids Res. 2021, 49, D325–D334. [CrossRef] [PubMed]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The Reactome Pathway Knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef]
- Ansari, R.; Khosravi, A.; Bahonar, A.; Shirani, S.; Kelishadi, R.; Khosravi, Z. Risk Factors of Atherosclerosis in Male Smokers, Passive Smokers, and Hypertensive Nonsmokers in Central Iran. ARYA Atheroscler. 2012, 8, 90–95. [Google Scholar]
- Prakash, V.; Jaker, S.; Burgan, A.; Jacques, A.; Fluck, D.; Sharma, P.; Fry, C.H.; Han, T.S. The Smoking-Dyslipidaemia Dyad: A Potent Synergistic Risk for Atherosclerotic Coronary Artery Disease. JRSM Cardiovasc. Dis. 2021, 10, 2048004020980945. [Google Scholar] [CrossRef]
- Blagov, A.V.; Markin, A.M.; Bogatyreva, A.I.; Tolstik, T.V.; Sukhorukov, V.N.; Orekhov, A.N. The Role of Macrophages in the Pathogenesis of Atherosclerosis. Cells 2023, 12, 522. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Zhong, W.; Wang, L.; Zhang, Q.; Ma, X.; Wang, Y.; Wang, S.; He, C.; Wei, Q.; Fu, C. Effects of Shear Stress on Vascular Endothelial Functions in Atherosclerosis and Potential Therapeutic Approaches. Biomed. Pharmacother. Biomed. Pharmacother. 2023, 158, 114198. [Google Scholar] [CrossRef]
- Frangos, S.G.; Gahtan, V.; Sumpio, B. Localization of Atherosclerosis: Role of Hemodynamics. Arch. Surg. 1999, 134, 1142–1149. [Google Scholar] [CrossRef]
- Heiss, C.; Rodriguez-Mateos, A.; Kelm, M. Central Role of ENOS in the Maintenance of Endothelial Homeostasis. Antioxid. Redox Signal. 2015, 22, 1230–1242. [Google Scholar] [CrossRef]
- Tran, N.; Garcia, T.; Aniqa, M.; Ali, S.; Ally, A.; Nauli, S. Endothelial Nitric Oxide Synthase (ENOS) and the Cardiovascular System: In Physiology and in Disease States. Am. J. Biomed. Sci. Res. 2022, 15, 153–177. [Google Scholar] [PubMed]
- Mun, G.I.; Kim, I.-S.; Lee, B.-H.; Boo, Y.C. Endothelial Argininosuccinate Synthetase 1 Regulates Nitric Oxide Production and Monocyte Adhesion under Static and Laminar Shear Stress Conditions*. J. Biol. Chem. 2011, 286, 2536–2542. [Google Scholar] [CrossRef]
- Stitham, J.; Midgett, C.; Martin, K.; Hwa, J. Prostacyclin: An Inflammatory Paradox. Front. Pharmacol. 2011, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Hamik, A.; Nayak, L.; Tian, H.; Shi, H.; Lu, Y.; Sharma, N.; Liao, X.; Hale, A.; Boerboom, L.; et al. Endothelial Kruppel-like Factor 4 Protects against Atherothrombosis in Mice. J. Clin. Investig. 2012, 122, 4727–4731. [Google Scholar] [CrossRef] [PubMed]
- Hamik, A.; Lin, Z.; Kumar, A.; Balcells, M.; Sinha, S.; Katz, J.; Feinberg, M.W.; Gerzsten, R.E.; Edelman, E.R.; Jain, M.K. Kruppel-like Factor 4 Regulates Endothelial Inflammation. J. Biol. Chem. 2007, 282, 13769–13779. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-F.; Wu, K.-J. Endothelial Transdifferentiation of Tumor Cells Triggered by the Twist1-Jagged1-KLF4 Axis: Relationship between Cancer Stemness and Angiogenesis. Stem Cells Int. 2015, 2016, e6439864. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, C.; Gu, Q.; Sims, M.; Gu, W.; Pfeffer, L.M.; Yue, J. KLF4 Promotes Angiogenesis by Activating VEGF Signaling in Human Retinal Microvascular Endothelial Cells. PLoS ONE 2015, 10, e0130341. [Google Scholar] [CrossRef]
- Atkins, G.B.; Jain, M.K. Role of Krüppel-like Transcription Factors in Endothelial Biology. Circ. Res. 2007, 100, 1686–1695. [Google Scholar] [CrossRef]
- Yoshida, T.; Yamashita, M.; Horimai, C.; Hayashi, M. Deletion of Krüppel-Like Factor 4 in Endothelial and Hematopoietic Cells Enhances Neointimal Formation Following Vascular Injury. J. Am. Heart Assoc. 2014, 3, e000622. [Google Scholar] [CrossRef]
- Wang, C.; Han, M.; Zhao, X.-M.; Wen, J.-K. Kruppel-like Factor 4 Is Required for the Expression of Vascular Smooth Muscle Cell Differentiation Marker Genes Induced by All-Trans Retinoic Acid. J. Biochem. 2008, 144, 313–321. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, B.; Zhao, L.; Shen, J.; Yang, Z.; Zhang, Y.; Fan, R.; Zhang, M.; Ma, D.; Zheng, L.; et al. KLF4-PFKFB3-Driven Glycolysis Is Essential for Phenotypic Switching of Vascular Smooth Muscle Cells. Commun. Biol. 2022, 5, 1332. [Google Scholar] [CrossRef]
- Zheng, B.; Han, M.; Wen, J.-K. Role of Krüppel-like Factor 4 in Phenotypic Switching and Proliferation of Vascular Smooth Muscle Cells. IUBMB Life 2010, 62, 132–139. [Google Scholar] [CrossRef]
- Cowan, C.E.; Kohler, E.E.; Dugan, T.A.; Mirza, M.K.; Malik, A.B.; Wary, K.K. Kruppel-like Factor-4 Transcriptionally Regulates VE-Cadherin Expression and Endothelial Barrier Function. Circ. Res. 2010, 107, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Kotlyarov, S.; Kotlyarova, A. Participation of Krüppel-like Factors in Atherogenesis. Metabolites 2023, 13, 448. [Google Scholar] [CrossRef]
- De Bock, K.; Georgiadou, M.; Carmeliet, P. Role of Endothelial Cell Metabolism in Vessel Sprouting. Cell Metab. 2013, 18, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Theodorou, K.; Boon, R.A. Endothelial Cell Metabolism in Atherosclerosis. Front. Cell Dev. Biol. 2018, 6, 82. [Google Scholar] [CrossRef]
- Rohlenova, K.; Veys, K.; Miranda-Santos, I.; De Bock, K.; Carmeliet, P. Endothelial Cell Metabolism in Health and Disease. Trends Cell Biol. 2018, 28, 224–236. [Google Scholar] [CrossRef]
- Eelen, G.; de Zeeuw, P.; Simons, M.; Carmeliet, P. Endothelial Cell Metabolism in Normal and Diseased Vasculature. Circ. Res. 2015, 116, 1231–1244. [Google Scholar] [CrossRef]
- Fitzgerald, G.; Soro-Arnaiz, I.; De Bock, K. The Warburg Effect in Endothelial Cells and Its Potential as an Anti-Angiogenic Target in Cancer. Front. Cell Dev. Biol. 2018, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Van Schaftingen, E.; Lederer, B.; Bartrons, R.; Hers, H.-G. A Kinetic Study of Pyrophosphate: Fructose-6-Phosphate Phosphotransferase from Potato Tubers. Eur. J. Biochem. 1982, 129, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Loscalzo, J. Abstract 9817: Krüppel-Like Factor 4 Integrates Immunometabolism of Human Arterial Endothelial Cells. Circulation 2021, 144, A9817. [Google Scholar] [CrossRef]
- Doddaballapur, A.; Michalik, K.M.; Manavski, Y.; Lucas, T.; Houtkooper, R.H.; You, X.; Chen, W.; Zeiher, A.M.; Potente, M.; Dimmeler, S.; et al. Laminar Shear Stress Inhibits Endothelial Cell Metabolism via KLF2-Mediated Repression of PFKFB3. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 137–145. [Google Scholar] [CrossRef]
- Vieceli Dalla Sega, F.; Aquila, G.; Fortini, F.; Vaccarezza, M.; Secchiero, P.; Rizzo, P.; Campo, G. Context-Dependent Function of ROS in the Vascular Endothelium: The Role of the Notch Pathway and Shear Stress. BioFactors Oxf. Engl. 2017, 43, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.J.; Mosqueiro, T.S.; Archer, B.J.; Jones, W.M.; Sunshine, H.; Faas, G.C.; Briot, A.; Aragón, R.L.; Su, T.; Romay, M.C.; et al. NOTCH1 Is a Mechanosensor in Adult Arteries. Nat. Commun. 2017, 8, 1620. [Google Scholar] [CrossRef] [PubMed]
- Shirakura, K.; Baluk, P.; Nottebaum, A.F.; Ipe, U.; Peters, K.G.; McDonald, D.M.; Vestweber, D. Shear Stress Control of Vascular Leaks and Atheromas through Tie2 Activation by VE-PTP Sequestration. EMBO Mol. Med. 2023, 15, e16128. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, A.; Fang, S.; Hemanthakumar, K.A.; Örd, T.; van Avondt, K.; Chevre, R.; Toropainen, A.; Singha, P.; Gilani, H.; Nguyen, S.D.; et al. The Angiopoietin Receptor Tie2 Is Atheroprotective in Arterial Endothelium. Nat. Cardiovasc. Res. 2023, 2, 307–321. [Google Scholar] [CrossRef]
- Dixon, R.A.F.; Diehl, R.E.; Opas, E.; Rands, E.; Vickers, P.J.; Evans, J.F.; Gillard, J.W.; Miller, D.K. Requirement of a 5-Lipoxygenase-Activating Protein for Leukotriene Synthesis. Nature 1990, 343, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, X.; Zhang, D.; Cheng, W.; Zhang, Y.; Yu, B.; Zhang, Y. Genetic Variation in the Leukotriene Pathway Is Associated with Myocardial Infarction in the Chinese Population. Lipids Health Dis. 2019, 18, 25. [Google Scholar] [CrossRef]
- Kotlyarov, S. Genetic and Epigenetic Regulation of Lipoxygenase Pathways and Reverse Cholesterol Transport in Atherogenesis. Genes 2022, 13, 1474. [Google Scholar] [CrossRef]
- van der Net, J.B.; Versmissen, J.; Oosterveer, D.M.; Defesche, J.C.; Yazdanpanah, M.; Aouizerat, B.E.; Steyerberg, E.W.; Malloy, M.J.; Pullinger, C.R.; Kane, J.P.; et al. Arachidonate 5-Lipoxygenase-Activating Protein (ALOX5AP) Gene and Coronary Heart Disease Risk in Familial Hypercholesterolemia. Atherosclerosis 2009, 203, 472–478. [Google Scholar] [CrossRef]
- Lehmann, C.; Homann, J.; Ball, A.-K.; Blöcher, R.; Kleinschmidt, T.K.; Basavarajappa, D.; Angioni, C.; Ferreirós, N.; Häfner, A.-K.; Rådmark, O.; et al. Lipoxin and Resolvin Biosynthesis Is Dependent on 5-Lipoxygenase Activating Protein. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 5029–5043. [Google Scholar] [CrossRef]
- Kim, C.W.; Oh, E.-T.; Park, H.J. A Strategy to Prevent Atherosclerosis via TNF Receptor Regulation. FASEB J. 2021, 35, e21391. [Google Scholar] [CrossRef]
- Qian, Y.; Mao, M.; Nian, F. The Effect of TNF-α on CHD and the Relationship between TNF-α Antagonist and CHD in Rheumatoid Arthritis: A Systematic Review. Cardiol. Res. Pract. 2022, 2022, e6192053. [Google Scholar] [CrossRef] [PubMed]
- Persson, J.; Nilsson, J.; Lindholm, M.W. Interleukin-1beta and Tumour Necrosis Factor-Alpha Impede Neutral Lipid Turnover in Macrophage-Derived Foam Cells. BMC Immunol. 2008, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liang, J.; Castrillon, D.H.; DePinho, R.A.; Olson, E.N.; Liu, Z.-P. FoxO4 Regulates Tumor Necrosis Factor Alpha-Directed Smooth Muscle Cell Migration by Activating Matrix Metalloproteinase 9 Gene Transcription. Mol. Cell. Biol. 2007, 27, 2676–2686. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.E.; Kandalam, V.; Chakrabarti, S.; Wang, X.; Penninger, J.M.; Davidge, S.T.; Oudit, G.Y.; Kassiri, Z. Tumor Necrosis Factor Induces Matrix Metalloproteinases in Cardiomyocytes and Cardiofibroblasts Differentially via Superoxide Production in a PI3Kgamma-Dependent Manner. Am. J. Physiol. Cell Physiol. 2010, 298, C679–C692. [Google Scholar] [CrossRef]
- Aukrust, P.; Sandberg, W.J.; Otterdal, K.; Vinge, L.E.; Gullestad, L.; Yndestad, A.; Halvorsen, B.; Ueland, T. Tumor Necrosis Factor Superfamily Molecules in Acute Coronary Syndromes. Ann. Med. 2011, 43, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.K. Cyclooxygenase-2 Induction in Congestive Heart Failure. Circulation 1998, 98, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Eligini, S.; Colli, S.; Habib, A.; Aldini, G.; Altomare, A.; Banfi, C. Cyclooxygenase-2 Glycosylation Is Affected by Peroxynitrite in Endothelial Cells: Impact on Enzyme Activity and Degradation. Antioxidants 2021, 10, 496. [Google Scholar] [CrossRef]
- Kirkby, N.S.; Lundberg, M.H.; Wright, W.R.; Warner, T.D.; Paul-Clark, M.J.; Mitchell, J.A. COX-2 Protects against Atherosclerosis Independently of Local Vascular Prostacyclin: Identification of COX-2 Associated Pathways Implicate Rgl1 and Lymphocyte Networks. PLoS ONE 2014, 9, e98165. [Google Scholar] [CrossRef]
- Belton, O.; Byrne, D.; Kearney, D.; Leahy, A.; Fitzgerald, D.J. Cyclooxygenase-1 and -2-Dependent Prostacyclin Formation in Patients with Atherosclerosis. Circulation 2000, 102, 840–845. [Google Scholar] [CrossRef]
- Yu, Z.; Crichton, I.; Tang, S.Y.; Hui, Y.; Ricciotti, E.; Levin, M.D.; Lawson, J.A.; Puré, E.; FitzGerald, G.A. Disruption of the 5-Lipoxygenase Pathway Attenuates Atherogenesis Consequent to COX-2 Deletion in Mice. Proc. Natl. Acad. Sci. USA 2012, 109, 6727–6732. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, M.; Yu, Y.; Lawson, J.; Funk, C.D.; Fitzgerald, G.A. Cyclooxygenases, Microsomal Prostaglandin E Synthase-1, and Cardiovascular Function. J. Clin. Investig. 2006, 116, 1391–1399. [Google Scholar] [CrossRef]
- Barbieri, S.S.; Amadio, P.; Gianellini, S.; Tarantino, E.; Zacchi, E.; Veglia, F.; Howe, L.R.; Weksler, B.B.; Mussoni, L.; Tremoli, E. Cyclooxygenase-2-Derived Prostacyclin Regulates Arterial Thrombus Formation by Suppressing Tissue Factor in a Sirtuin-1-Dependent-Manner. Circulation 2012, 126, 1373–1384. [Google Scholar] [CrossRef]
- Felizola, S.J.A.; Nakamura, Y.; Ozawa, Y.; Ono, Y.; Morimoto, R.; Midorikawa, S.; Suzuki, S.; Satoh, F.; Sasano, H. Activating Transcription Factor 3 (ATF3) in the Human Adrenal Cortex: Its Possible Involvement in Aldosterone Biosynthesis. Tohoku J. Exp. Med. 2014, 234, 249–254. [Google Scholar] [CrossRef]
- Wu, J.; Huang, Y.; Zhou, X.; Xiang, Z.; Yang, Z.; Meng, D.; Wu, D.; Zhang, J.; Yang, J. ATF3 and Its Emerging Role in Atherosclerosis: A Narrative Review. Cardiovasc. Diagn. Ther. 2022, 12, 926. [Google Scholar] [CrossRef] [PubMed]
- McDonald, A.I.; Shirali, A.S.; Aragón, R.; Ma, F.; Hernandez, G.; Vaughn, D.A.; Mack, J.J.; Lim, T.Y.; Sunshine, H.; Zhao, P.; et al. Endothelial Regeneration of Large Vessels Is a Biphasic Process Driven by Local Cells with Distinct Proliferative Capacities. Cell Stem Cell 2018, 23, 210–225. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, A.; Iwamoto, Y.; Maru, Y. Oxidative Stress-Responsive Transcription Factor ATF3 Potentially Mediates Diabetic Angiopathy. Mol. Cell. Biol. 2006, 26, 1087–1097. [Google Scholar] [CrossRef]
- Nawa, T.; Nawa, M.T.; Adachi, M.T.; Uchimura, I.; Shimokawa, R.; Fujisawa, K.; Tanaka, A.; Numano, F.; Kitajima, S. Expression of Transcriptional Repressor ATF3/LRF1 in Human Atherosclerosis: Colocalization and Possible Involvement in Cell Death of Vascular Endothelial Cells. Atherosclerosis 2002, 161, 281–291. [Google Scholar] [CrossRef]
- Yan, C.; Lu, D.; Hai, T.; Boyd, D.D. Activating Transcription Factor 3, a Stress Sensor, Activates P53 by Blocking Its Ubiquitination. EMBO J. 2005, 24, 2425–2435. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, J.; Zhang, C.; Nobori, K.; Hashimoto, Y.; Adachi, M.T.; Noda, A.; Sunamori, M.; Kitajima, S. Transcriptional Repressor Activating Transcription Factor 3 Protects Human Umbilical Vein Endothelial Cells from Tumor Necrosis Factor-Alpha-Induced Apoptosis through down-Regulation of P53 Transcription. J. Biol. Chem. 2002, 277, 39025–39034. [Google Scholar] [CrossRef]
- Luo, C.; Ruan, Y.; Sun, P.; Wang, H.; Yang, W.; Gong, Y.; Wang, D. The Role of Transcription Factors in Coronary Artery Disease and Myocardial Infarction. FBL 2022, 27, 329. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, Y.; Nabavi, S.M.; Sahebkar, A.; Little, P.J.; Xu, S.; Weng, J.; Ge, J. Mechanisms of Oxidized LDL-Mediated Endothelial Dysfunction and Its Consequences for the Development of Atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 925923. [Google Scholar] [CrossRef]
- Mundi, S.; Massaro, M.; Scoditti, E.; Carluccio, M.A.; van Hinsbergh, V.W.M.; Iruela-Arispe, M.L.; De Caterina, R. Endothelial Permeability, LDL Deposition, and Cardiovascular Risk Factors—A Review. Cardiovasc. Res. 2017, 114, 35–52. [Google Scholar] [CrossRef]
- Zhang, X.; Sessa, W.C.; Fernández-Hernando, C. Endothelial Transcytosis of Lipoproteins in Atherosclerosis. Front. Cardiovasc. Med. 2018, 5, 130. [Google Scholar] [CrossRef]
- van den Borne, P.; Quax, P.H.A.; Hoefer, I.E.; Pasterkamp, G. The Multifaceted Functions of CXCL10 in Cardiovascular Disease. BioMed Res. Int. 2014, 2014, 893106. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Wang, Z.; Ye, D.; Feng, Y.; Liu, M.; Xu, Y.; Wang, M.; Zhang, J.; Liu, J.; Zhao, M.; et al. The Role of CXC Chemokines in Cardiovascular Diseases. Front. Pharmacol. 2022, 12, 765768. [Google Scholar] [CrossRef]
- Gencer, S.; Evans, B.R.; van der Vorst, E.P.C.; Döring, Y.; Weber, C. Inflammatory Chemokines in Atherosclerosis. Cells 2021, 10, 226. [Google Scholar] [CrossRef]
- Allam, E.; Delacruz, K.; Ghoneima, A.; Sun, J.; Windsor, L.J. Effects of Tobacco on Cytokine Expression from Human Endothelial Cells. Oral Dis. 2013, 19, 660–665. [Google Scholar] [CrossRef]
- Döring, Y.; Noels, H.; van der Vorst, E.P.C.; Neideck, C.; Egea, V.; Drechsler, M.; Mandl, M.; Pawig, L.; Jansen, Y.; Schröder, K.; et al. Vascular CXCR4 Limits Atherosclerosis by Maintaining Arterial Integrity: Evidence from Mouse and Human Studies. Circulation 2017, 136, 388–403. [Google Scholar] [CrossRef] [PubMed]
- Weerackoon, N.; Gunawardhana, K.L.; Mani, A. Wnt Signaling Cascades and Their Role in Coronary Artery Health and Disease. J. Cell. Signal. 2021, 2, 52–62. [Google Scholar] [CrossRef]
- Du, J.; Li, J. The Role of Wnt Signaling Pathway in Atherosclerosis and Its Relationship with Angiogenesis. Exp. Ther. Med. 2018, 16, 1975–1981. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Neogi, A.; Mani, A. The Role of Wnt Signalling in Development of Coronary Artery Disease and Its Risk Factors. Open Biol. 2020, 10, 200128. [Google Scholar] [CrossRef] [PubMed]
- Wadey, K.S.; Somos, A.; Leyden, G.; Blythe, H.; Chan, J.; Hutchinson, L.; Poole, A.; Frankow, A.; Johnson, J.L.; George, S.J. Pro-Inflammatory Role of Wnt/β-Catenin Signaling in Endothelial Dysfunction. Front. Cardiovasc. Med. 2023, 9, 1059124. [Google Scholar] [CrossRef] [PubMed]
- Grigorescu, E.-D.; Sorodoc, V.; Floria, M.; Anisie, E.; Popa, A.D.; Onofriescu, A.; Ceasovschih, A.; Sorodoc, L. The Inflammatory Marker HSCRP as a Predictor of Increased Insulin Resistance in Type 2 Diabetics without Atherosclerotic Manifestations. Rev. Chim. 2019, 70, 1791–1794. [Google Scholar] [CrossRef]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuñiga, F.A. Association between Insulin Resistance and the Development of Cardiovascular Disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef]
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and Atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Other Names | Full Name | Protein | Biological Processes |
|---|---|---|---|---|
| CRP | PTX1 | C-reactive protein | C-reactive protein | Inflammatory response; innate immune response; negative regulation of macrophage-derived foam cell differentiation. |
| TEK | TIE2; VMCM; GLC3E; TIE-2; VMCM1; CD202B | TEK receptor tyrosine kinase | Angiopoietin-1 receptor | Angiogenesis; endothelial cell proliferation; cell–cell signaling; negative regulation of endothelial cell apoptotic process; negative regulation of apoptotic process; positive regulation of endothelial cell migration; positive regulation of MAPK cascade; positive regulation of actin cytoskeleton reorganization; regulation of vascular permeability. |
| NTRK1 | MTC; TRK; TRK1; TRKA; Trk-A; p140-TrkA | Neurotrophic receptor tyrosine kinase 1 | High-affinity nerve growth factor receptor | Negative regulation of apoptotic process; positive regulation of programmed cell death; mechanoreceptor differentiation; positive regulation of angiogenesis; positive regulation of ERK1 and ERK2 cascade; positive regulation of NF-kappaB transcription factor activity; transmembrane receptor protein tyrosine kinase signaling pathway. |
| CD34 | - | CD34 molecule | Hematopoietic progenitor cell antigen | Cell motility; cell–cell adhesion; cell–matrix adhesion; endothelial cell proliferation; hemopoiesis; leukocyte migration; negative regulation of blood coagulation; positive regulation of angiogenesis. |
| FGF18 | ZFGF5; FGF-18 | Fibroblast growth factor 18 | Fibroblast growth factor 18 | Angiogenesis; cell differentiation; cell–cell signaling; ERK1 and ERK2 cascade; positive regulation of blood vessel endothelial cell migration; positive regulation of endothelial cell chemotaxis to fibroblast growth factor. |
| NOS3 | eNOS; ECNOS | Nitric oxide synthase 3 | Nitric oxide synthase, endothelial | Nitric oxide biosynthetic process; angiogenesis; endothelial cell migration; negative regulation of blood pressure; regulation of platelet activation; negative regulation of smooth muscle cell proliferation. |
| KLF4 | EZF; GKLF | KLF transcription factor 4 | Krueppel-like factor 4 | Cellular response to laminar fluid shear stress; canonical Wnt signaling pathway; negative regulation of angiogenesis; negative regulation of inflammatory response; negative regulation of interleukin-8 production; negative regulation of leukocyte adhesion to arterial endothelial cell; negative regulation of NF-kappaB transcription factor activity; negative regulation of response to cytokine stimulus. |
| THBD | TM; THRM; AHUS6; BDCA3; CD141; BDCA-3; THPH12 | Thrombomodulin | Thrombomodulin | Blood coagulation; negative regulation of blood coagulation; negative regulation of platelet activation. |
| ITPR3 | IP3R; CMT1J; IP3R3 | Inositol 1,4,5-trisphosphate receptor type 3 | Inositol 1,4,5-trisphosphate receptor type 3 | G protein-coupled receptor signaling pathway; calcium transport. |
| ELN | WS; WBS; SVAS; ADCL1 | Elastin | Elastin | Extracellular matrix assembly; regulation of smooth muscle cell proliferation. |
| Gene Symbol | Other Names | Full Name | Protein | Biological Processes |
|---|---|---|---|---|
| ALOX5AP | FLAP | Arachidonate 5-lipoxygenase-activating protein | Arachidonate 5-lipoxygenase-activating protein | Leukotriene biosynthesis; synthesis of lipoxins (LX); synthesis of 5-eicosatetraenoic acids. |
| ANGPT2 | ANG2; AGPT2; LMPHM10 | Angiopoietin 2 | Angiopoietin-2 | Angiogenesis; response to glucose; response to mechanical stimulus; response to hypoxia. |
| KIT | PBT; SCFR; C-Kit; CD117; MASTC | KIT proto-oncogene, receptor tyrosine kinase | Mast/stem cell growth factor receptor kit | Regulation of cell survival and proliferation; mast cell development, migration, and function; actin cytoskeleton reorganization; hematopoiesis, stem cell maintenance; cytokine-mediated signaling pathway. |
| LCP2 | IMD81; SLP76; SLP-76 | Lymphocyte cytosolic protein 2 | Lymphocyte cytosolic protein 2 | Immune response; intracellular signal transduction; mast cell activation; positive regulation of protein kinase activity; transmembrane receptor protein tyrosine kinase signaling pathway. |
| C3AR1 | AZ3B; C3AR; HNFAG09 | Complement C3a receptor 1 | C3a anaphylatoxin chemotactic receptor | Chemotaxis; positive regulation of macrophage chemotaxis; complement receptor-mediated signaling pathway; inflammatory response; positive regulation of angiogenesis; positive regulation of vascular endothelial growth factor production. |
| CD69 | AIM; EA1; MLR-3; CLEC2C; GP32/28; BL-AC/P26 | CD69 molecule | Early-activation antigen CD69 | Cellular response to xenobiotic stimulus. |
| CXCR4 | FB22; HM89; LAP3; LCR1; NPYR; WHIM; CD184; LAP-3; LESTR; NPY3R; NPYRL; WHIMS; HSY3RR; NPYY3R; WHIMS1; D2S201E | C-X-C motif chemokine receptor 4 | C-X-C chemokine receptor type 4 | Apoptotic process; cell chemotaxis; CXCL12-activated CXCR4 signaling pathway; endothelial cell differentiation; immune response; inflammatory response; positive regulation of cell migration. |
| Gene Symbol | Other Names | Full Name | Protein | Biological Processes |
|---|---|---|---|---|
| EGR1 | TIS8; AT225; G0S30; NGFI-A; ZNF225; KROX-24; ZIF-268 | Early-growth response 1 | Early-growth response protein 1 | Cellular response to interleukin-8; interleukin-1-mediated signaling pathway; negative regulation of canonical Wnt signaling pathway; positive regulation of chemokine production; regulation of apoptotic process; response to glucose; response to insulin; response to hypoxia. |
| ATF3 | - | Activating transcription factor 3 | Cyclic AMP-dependent transcription factor ATF-3 | Gluconeogenesis; positive regulation of cell population proliferation; positive regulation of transcription by RNA polymerase II. |
| CXCL8 | IL8; NAF; GCP1; LECT; LUCT; NAP1; GCP-1; LYNAP; MDNCF; MONAP; NAP-1; SCYB8 | C-X-C motif chemokine ligand 8 | Interleukin-8 | Chemotaxis; neutrophil chemotaxis; angiogenesis; cellular response to interleukin-1; cellular response to lipopolysaccharide; inflammatory response; regulation of cell adhesion. |
| SOCS3 | CIS3; SSI3; ATOD4; Cish3; SSI-3; SOCS-3 | Suppressor of cytokine signaling 3 | Suppressor of cytokine signaling 3 | Cell differentiation; negative regulation of apoptotic process; negative regulation of inflammatory response; negative regulation of insulin receptor signaling pathway. |
| FOS | p55; AP-1; C-FOS | Fos proto-oncogene, AP-1 transcription factor subunit | Protein c-Fos | Cellular response to hypoxia; response to insulin; cellular response to reactive oxygen species; cellular response to tumor necrosis factor; inflammatory response; response to toxic substance; response to xenobiotic stimulus. |
| PTGS2 | COX2; COX-2; PHS-2; PGG/HS; PGHS-2; hCox-2; GRIPGHS | Prostaglandin-endoperoxide synthase 2 | Prostaglandin G/H synthase 2 | Angiogenesis; cellular response to fluid shear stress; cellular response to hypoxia; inflammatory response; cyclooxygenase pathway; prostaglandin biosynthetic process; positive regulation of vasoconstriction; fatty acid metabolism. |
| Gene Symbol | Other Names | Full Name | Protein | Biological Processes |
|---|---|---|---|---|
| MX1 | MX; MxA; IFI78; IFI-78K; lncMX1-215 | MX dynamin like GTPase 1 | Interferon-induced GTP-binding protein Mx1 | Antiviral innate immune response; apoptotic process; innate immune response; response to Type I interferon. |
| CMPK2 | NDK; TYKi; TMPK2; UMP-CMPK2 | Cytidine/uridine monophosphate kinase 2 | UMP-CMP kinase 2, mitochondrial | Cellular response to lipopolysaccharide; pyrimidine biosynthesis. |
| OAS1 | OIAS; IFI-4; OIASI; IMD100; E18/E16 | 2′-5′-oligoadenylate synthetase 1 | 2′-5′-oligoadenylate synthase 1 | Cellular response to interferon-alpha; cellular response to interferon-beta; glucose homeostasis; antiviral defense. |
| OAS2 | - | 2′-5′-oligoadenylate synthetase 2 | 2′-5′-oligoadenylate synthase 2 | Antiviral defense; immunity; innate immunity. |
| IFI44L | GS3686; TLDC5B; C1orf29 | Interferon-induced protein 44 like | Interferon-induced protein 44-like | Antiviral defense. |
| IFIH1 | - | Interferon-induced with helicase C domain 1 | Interferon-induced helicase C domain-containing protein 1 | Antiviral defense; immunity; innate immunity. |
| PARP9 | BAL; BAL1; ARTD9; MGC:7868 | Poly(ADP-ribose) polymerase family member 9 | Protein mono-ADP-ribosyltransferase PARP9 | Antiviral defense; immunity; innate immunity; DNA damage; DNA repair. |
| RSAD2 | SAND; cig5; vig1; cig33 | Radical S-adenosyl methionine domain containing 2 | S-adenosylmethionine-dependent nucleotide dehydratase RSAD2 | Antiviral defense; immunity; innate immunity. |
| XAF1 | BIRC4BP; XIAPAF1; HSXIAPAF1 | XIAP-associated factor 1 | XIAP-associated factor 1 | Apoptotic process; response to interferon-beta. |
| IFIT1 | C56; P56; G10P1; IFI56; ISG56; IFI-56; IFIT-1; IFNAI1; RNM561; IFI-56K | Interferon-induced protein with tetratricopeptide repeats 1 | Interferon-induced protein with tetratricopeptide repeats 1 | Antiviral defense; immunity; innate immunity. |
| Gene Symbol | Other Names | Full Name | Protein | Biological Processes |
|---|---|---|---|---|
| MX1 | MX; MxA; IFI78; IFI-78K; lncMX1-215 | MX dynamin like GTPase 1 | Interferon-induced GTP-binding protein Mx1 | Antiviral innate immune response; apoptotic process; innate immune response; response to Type I interferon. |
| IFIH1 | - | Interferon-induced with helicase C domain 1 | Interferon-induced helicase C domain-containing protein 1 | Antiviral defense; immunity; innate immunity. |
| IFIT1 | C56; P56; G10P1; IFI56; ISG56; IFI-56; IFIT-1; IFNAI1; RNM561; IFI-56K | Interferon-induced protein with tetratricopeptide repeats 1 | Interferon-induced protein with tetratricopeptide repeats 1 | Antiviral defense; immunity; innate immunity. |
| OAS2 | - | 2′-5′-oligoadenylate synthetase 2 | 2′-5′-oligoadenylate synthase 2 | Antiviral defense; immunity; innate immunity. |
| OAS3 | p100; p100OAS | 2′-5′-oligoadenylate synthetase 3 | 2′-5′-oligoadenylate synthase 3 | Antiviral defense; immunity; innate immunity. |
| DDX58 | RIGI; RIG1; RIG-I; RLR-1; SGMRT2 | RNA sensor RIG-I | Antiviral innate immune response receptor RIG-I | Antiviral defense; immunity; innate immunity. |
| ISG15 | G1P2; IP17; UCRP; IFI15; IMD38; hUCRP | ISG15 ubiquitin-like modifier | Ubiquitin-like protein ISG15 | Antiviral defense; immunity; innate immunity. |
| OASL | OASL1; OASLd; TRIP14; TRIP-14; p59OASL; p59 OASL; p59-OASL | 2′-5′-oligoadenylate synthetase-like | 2′-5′-oligoadenylate synthase-like protein | Antiviral defense; immunity; innate immunity. |
| IFIT2 | P54; G10P2; IFI54; ISG54; cig42; IFI-54; IFIT-2; ISG-54; GARG-39; IFI-54K; ISG-54K; ISG-54 K | Interferon-induced protein with tetratricopeptide repeats 2 | Interferon-induced protein with tetratricopeptide repeats 2 | Antiviral defense; immunity; innate immunity. |
| Gene Symbol | Other Names | Full Name | Protein | Biological Processes |
|---|---|---|---|---|
| IDI1 | IPP1; IPPI1 | Isopentenyl-diphosphate delta isomerase 1 | Isopentenyl-diphosphate Delta-isomerase 1 | Cholesterol biosynthetic process isoprenoid biosynthetic process; lipid metabolism. |
| INSIG1 | CL6 | Insulin-induced gene 1 | Insulin-induced gene 1 protein | Cellular response to insulin stimulus; cellular response to sterol; cholesterol biosynthetic process; SREBP signaling pathway; triglyceride metabolic process. |
| LDLR | FH; FHC; FHCL1; LDLCQ2 | Low-density lipoprotein receptor | Low-density lipoprotein receptor | Artery morphogenesis; cellular response to fatty acid; cellular response to low-density lipoprotein particle stimulus; cholesterol homeostasis; endocytosis; lipid metabolic process; low-density lipoprotein particle clearance; phagocytosis; phospholipid transport; positive regulation of inflammatory response; receptor-mediated endocytosis. |
| SREBF2 | SREBP2; bHLHd2; SREBP-2 | Sterol regulatory element-binding transcription factor 2 | Sterol regulatory element-binding protein 2 | Cellular response to laminar fluid shear stress; cholesterol homeostasis; lipid metabolic process; sterol metabolism. |
| DHCR24 | DCE; SELADIN1; Nbla03646; seladin-1 | 24-dehydrocholesterol reductase | Delta(24)-sterol reductase | Cholesterol biosynthetic process; membrane organization; negative regulation of apoptotic process; response to oxidative stress; steroid biosynthesis. |
| HMGCS1 | HMGCS | 3-hydroxy-3-methylglutaryl-CoA synthase 1 | Hydroxymethylglutaryl-CoA synthase, cytoplasmic | Cholesterol biosynthetic process; lipid metabolic process. |
| ACAT2 | - | Acetyl-CoA acetyltransferase 2 | Acetyl-CoA acetyltransferase, cytosolic | Acetyl-CoA C-acetyltransferase activity; fatty acid beta-oxidation; lipid metabolic process. |
| SQLE | - | Squalene epoxidase | Squalene monooxygenase | Cholesterol metabolic process; sterol biosynthetic process; regulation of cell population proliferation. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotlyarov, S. Effects of Atherogenic Factors on Endothelial Cells: Bioinformatics Analysis of Differentially Expressed Genes and Signaling Pathways. Biomedicines 2023, 11, 1216. https://doi.org/10.3390/biomedicines11041216
Kotlyarov S. Effects of Atherogenic Factors on Endothelial Cells: Bioinformatics Analysis of Differentially Expressed Genes and Signaling Pathways. Biomedicines. 2023; 11(4):1216. https://doi.org/10.3390/biomedicines11041216
Chicago/Turabian StyleKotlyarov, Stanislav. 2023. "Effects of Atherogenic Factors on Endothelial Cells: Bioinformatics Analysis of Differentially Expressed Genes and Signaling Pathways" Biomedicines 11, no. 4: 1216. https://doi.org/10.3390/biomedicines11041216