
The Human Extracellular Matrix Diseasome Reveals Genotype–Phenotype Associations with Clinical Implications for Age-Related Diseases



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
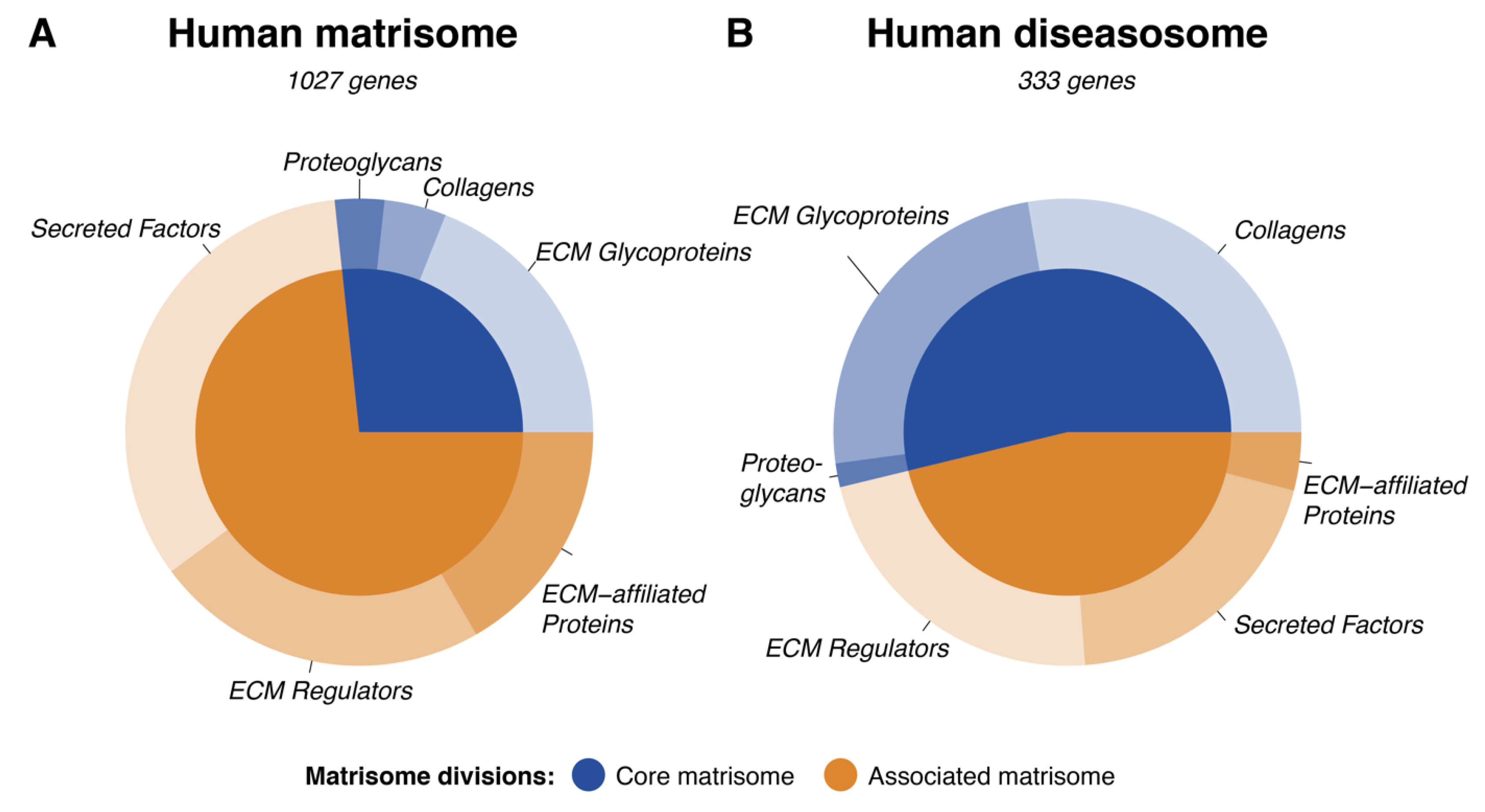
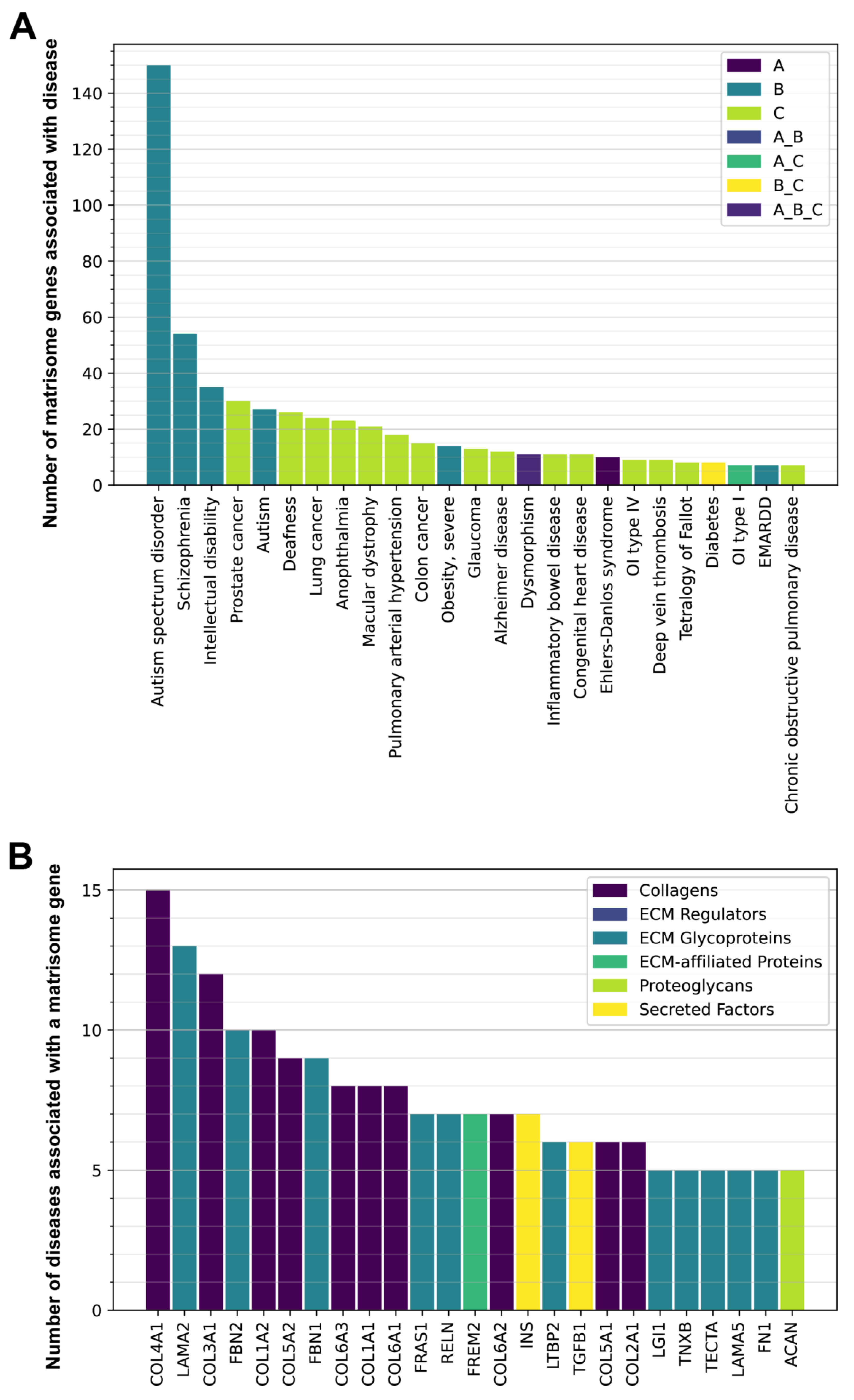
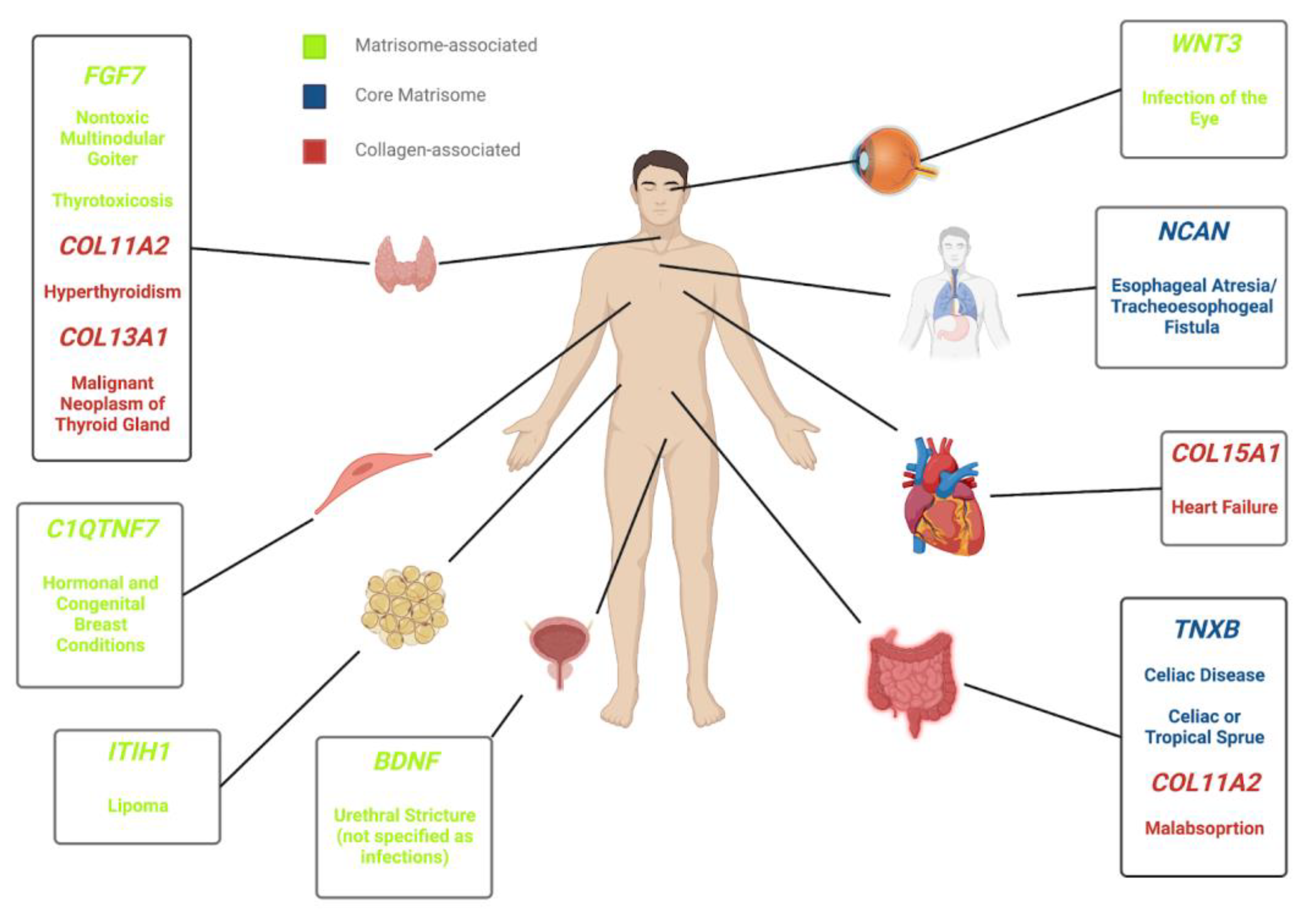
3.1. Clinical Implications of the Human Matrisome
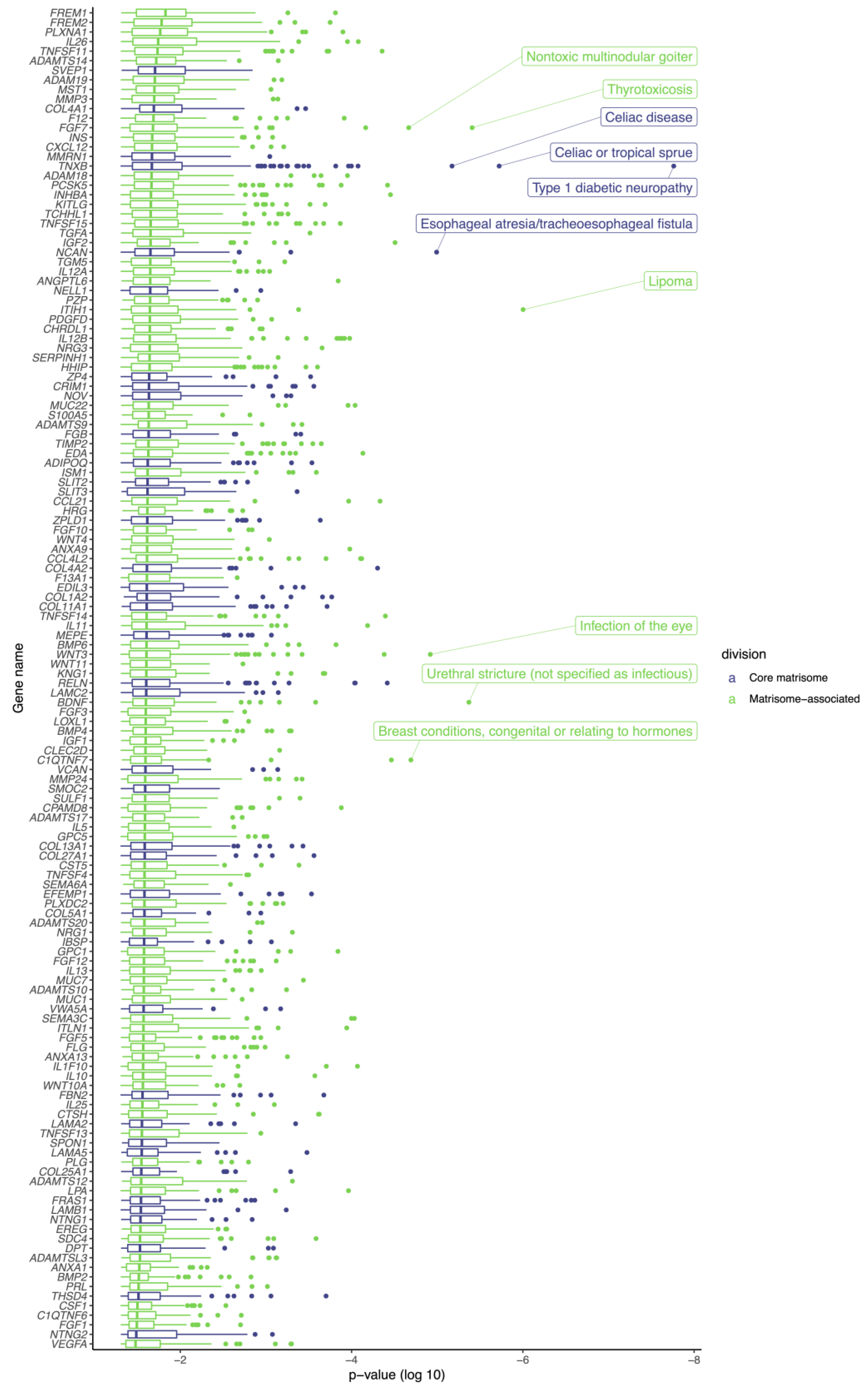
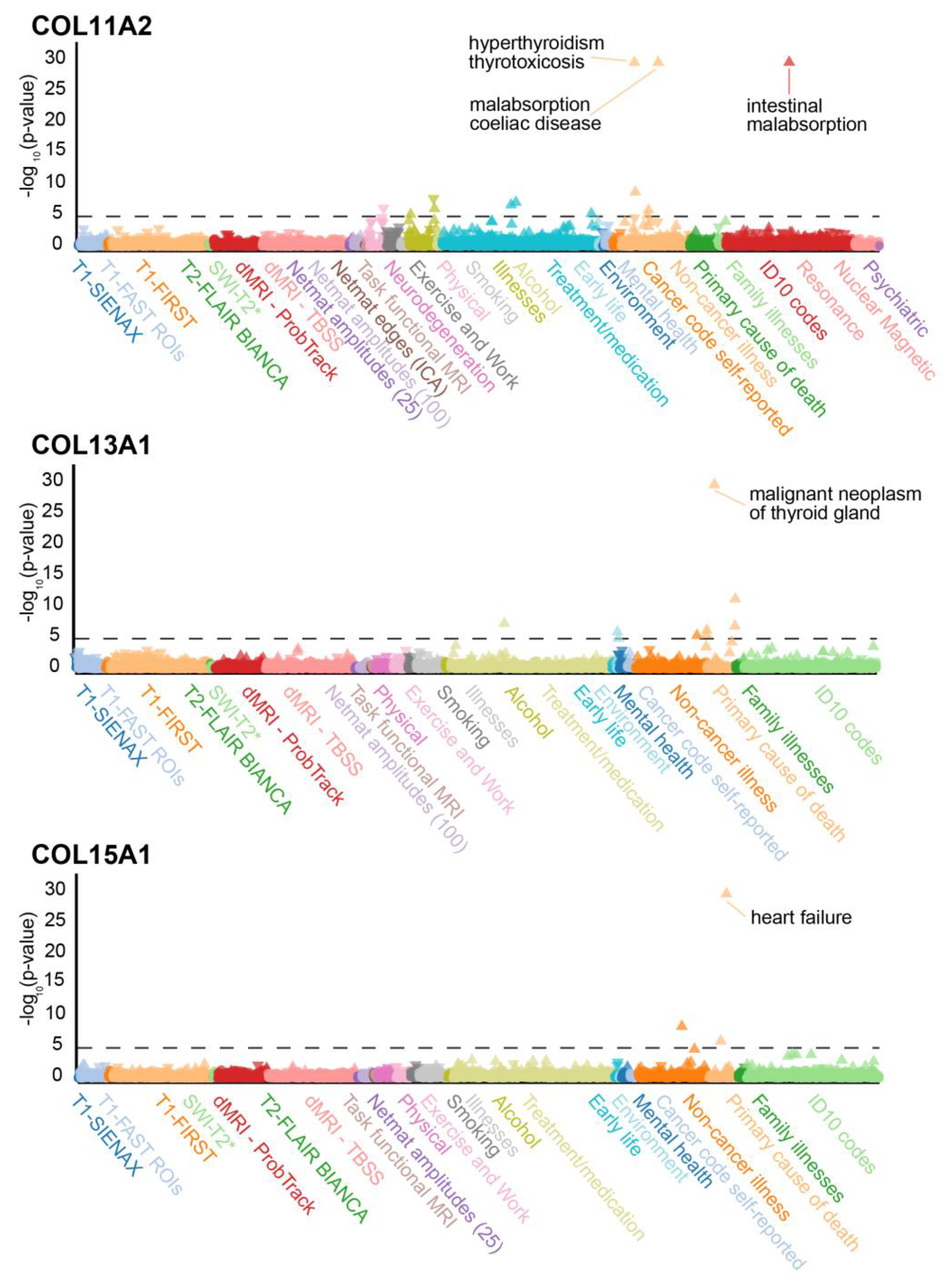
3.2. Genetic Variants in Matrisome Genes Associated with Diseases
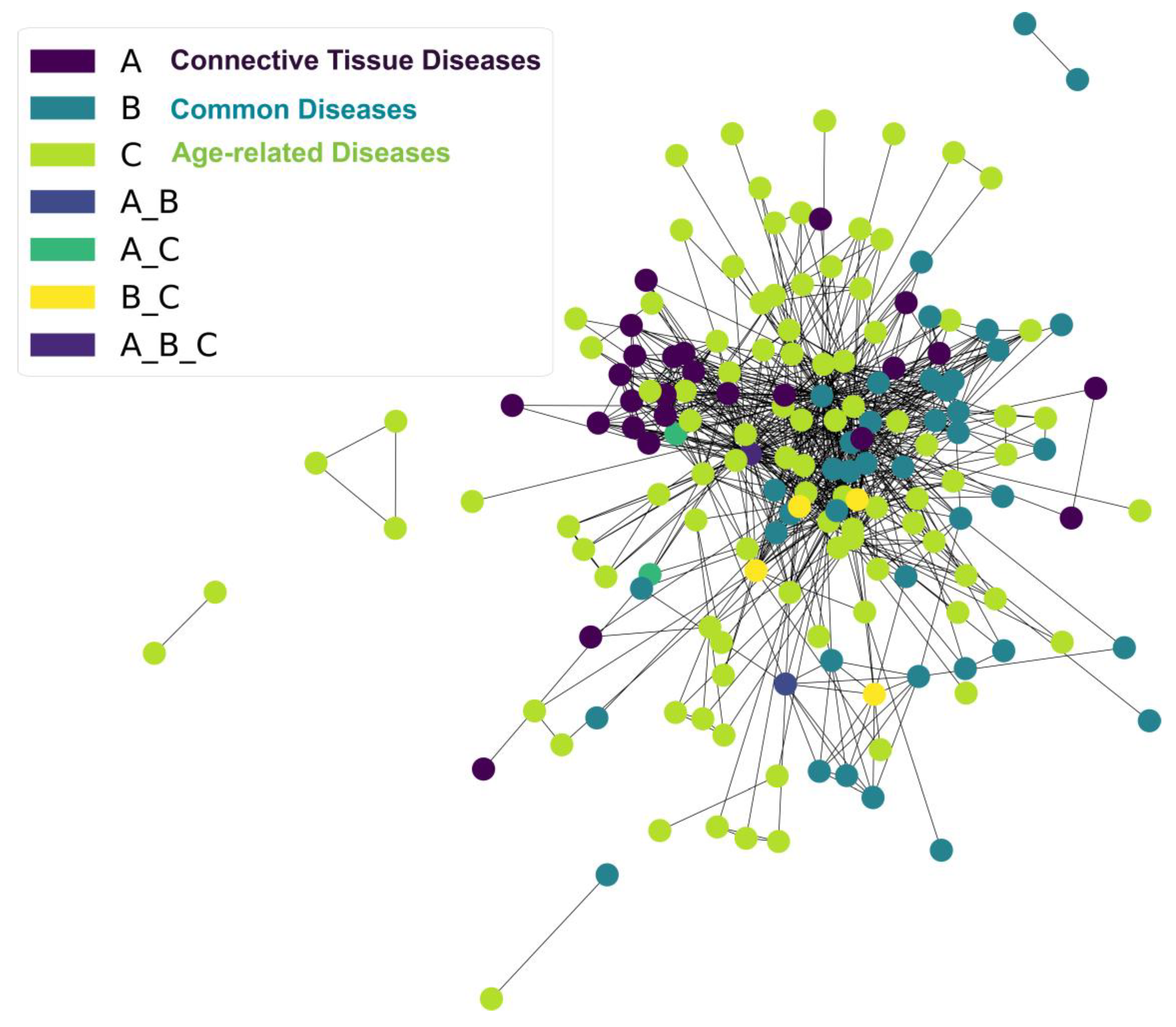
3.3. Matrisome in Age-Related Diseases
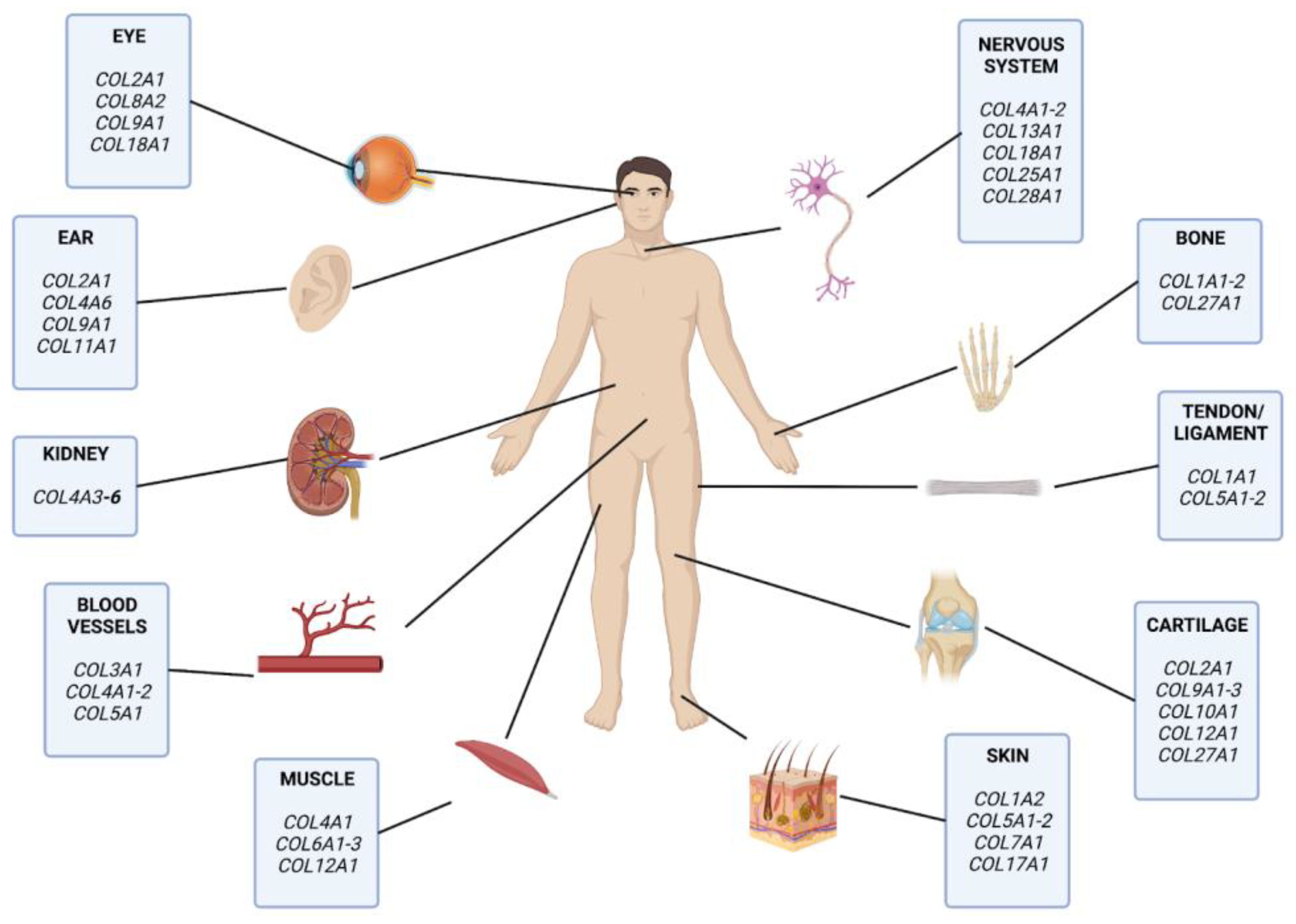
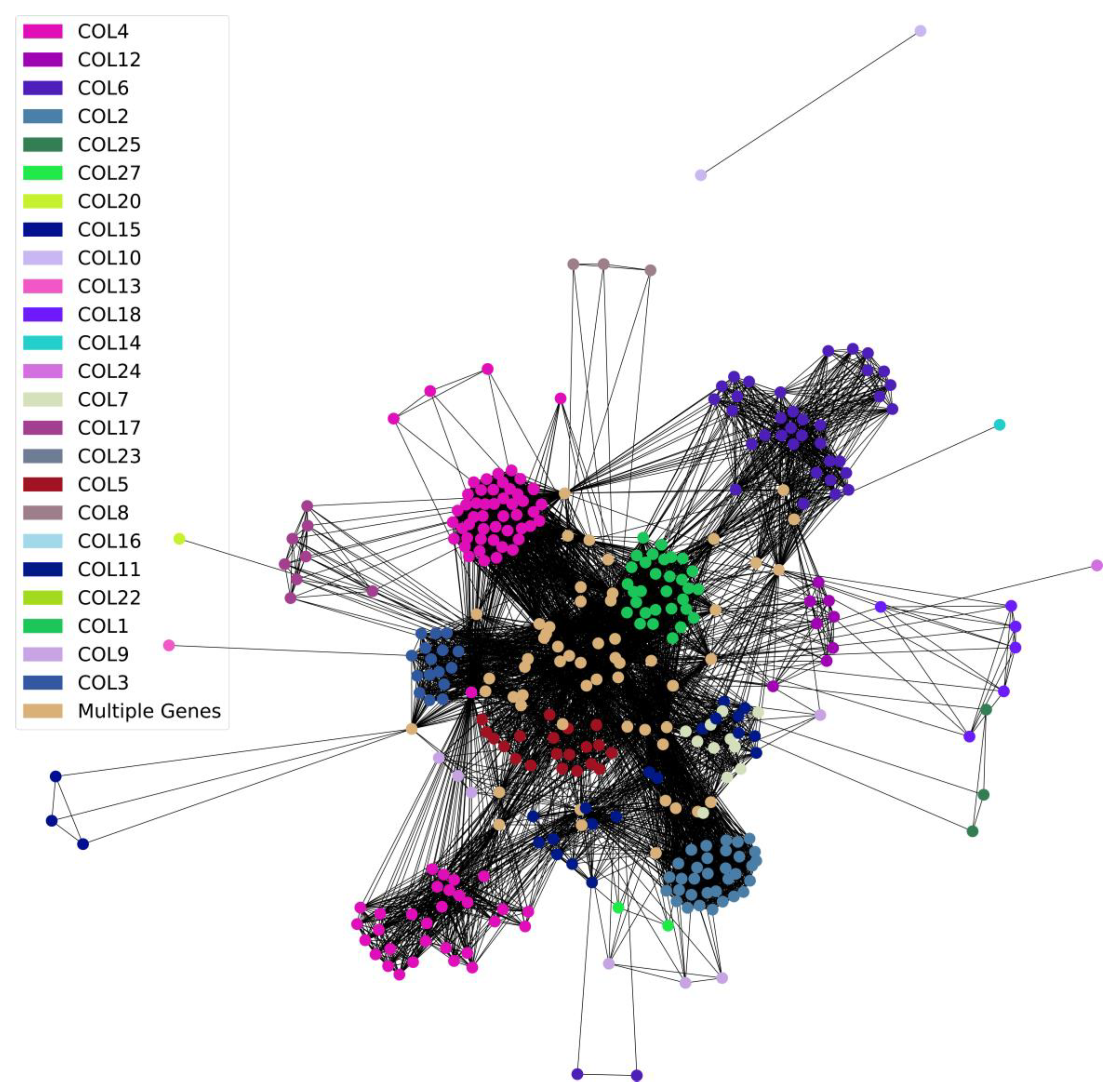
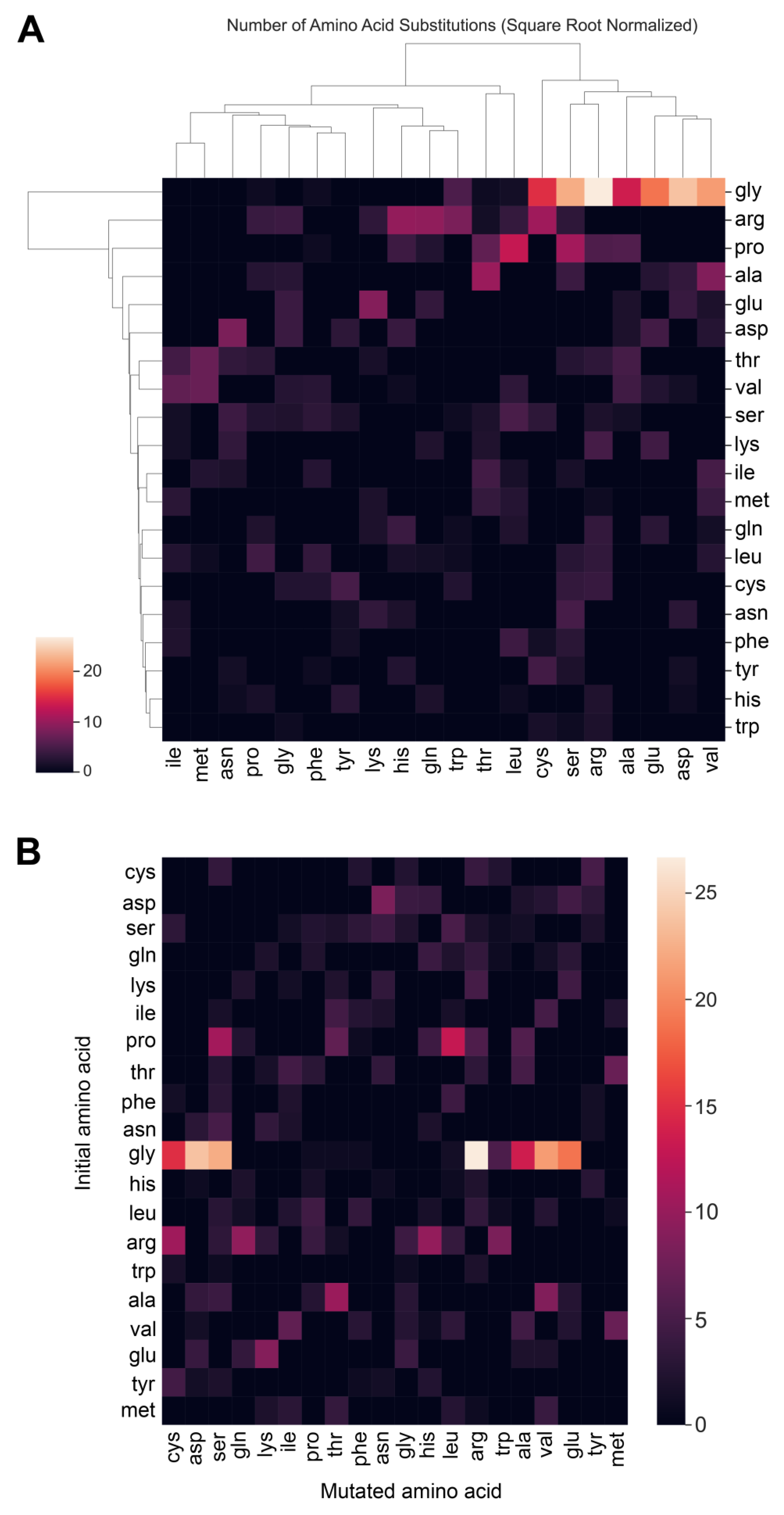
3.4. Collagens and Diseases
3.5. Potential Strategies Using Matrisome for Drug Repurposing
3.6. Targeting Matrisome Proteins in Rare Diseases
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bush, W.S.; Oetjens, M.T.; Crawford, D.C. Unravelling the Human Genome–Phenome Relationship Using Phenome-Wide Association Studies. Nat. Rev. Genet. 2016, 17, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Denny, J.C.; Bastarache, L.; Roden, D.M. Phenome-Wide Association Studies as a Tool to Advance Precision Medicine. Annu. Rev. Genom. Hum. Genet. 2016, 17, 353–373. [Google Scholar] [CrossRef]
- Lappalainen, I.; Almeida-King, J.; Kumanduri, V.; Senf, A.; Spalding, J.D.; ur-Rehman, S.; Saunders, G.; Kandasamy, J.; Caccamo, M.; Leinonen, R.; et al. The European Genome-Phenome Archive of Human Data Consented for Biomedical Research. Nat. Genet. 2015, 47, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Roden, D.M. Phenome-Wide Association Studies: A New Method for Functional Genomics in Humans. J. Physiol. 2017, 595, 4109–4115. [Google Scholar] [CrossRef] [PubMed]
- Shefchek, K.A.; Harris, N.L.; Gargano, M.; Matentzoglu, N.; Unni, D.; Brush, M.; Keith, D.; Conlin, T.; Vasilevsky, N.; Zhang, X.A.; et al. The Monarch Initiative in 2019: An Integrative Data and Analytic Platform Connecting Phenotypes to Genotypes across Species. Nucleic Acids Res. 2020, 48, D704–D715. [Google Scholar] [CrossRef]
- Ewald, C.Y. The Matrisome during Aging and Longevity: A Systems-Level Approach toward Defining Matreotypes Promoting Healthy Aging. Gerontology 2020, 66, 266–274. [Google Scholar] [CrossRef]
- Ewald, C.Y.; Landis, J.N.; Abate, J.P.; Murphy, C.T.; Blackwell, T.K. Dauer-Independent Insulin/IGF-1-Signalling Implicates Collagen Remodelling in Longevity. Nature 2015, 519, 97–101. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the Extracellular Matrix in Development and Disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Taha, I.N.; Naba, A. Exploring the Extracellular Matrix in Health and Disease Using Proteomics. Essays Biochem. 2019, 63, 417–432. [Google Scholar] [CrossRef]
- Lampi, M.C.; Reinhart-King, C.A. Targeting Extracellular Matrix Stiffness to Attenuate Disease: From Molecular Mechanisms to Clinical Trials. Sci. Transl. Med. 2018, 10, eaao0475. [Google Scholar] [CrossRef]
- Hynes, R.O.; Naba, A. Overview of the Matrisome—An Inventory of Extracellular Matrix Constituents and Functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The Extracellular Matrix: Tools and Insights for the “Omics” Era. Matrix Biol. J. Int. Soc. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef]
- Cirincione, A.G.; Clark, K.L.; Kann, M.G. Pathway Networks Generated from Human Disease Phenome. BMC Med. Genom. 2018, 11, 75. [Google Scholar] [CrossRef]
- Goh, K.-I.; Cusick, M.E.; Valle, D.; Childs, B.; Vidal, M.; Barabási, A.-L. The Human Disease Network. Proc. Natl. Acad. Sci. USA 2007, 104, 8685–8690. [Google Scholar] [CrossRef]
- Arseni, L.; Lombardi, A.; Orioli, D. From Structure to Phenotype: Impact of Collagen Alterations on Human Health. Int. J. Mol. Sci. 2018, 19, 1407. [Google Scholar] [CrossRef]
- Myllyharju, J.; Kivirikko, K.I. Collagens and Collagen-Related Diseases. Ann. Med. 2001, 33, 7–21. [Google Scholar] [CrossRef]
- Denny, J.C.; Bastarache, L.; Ritchie, M.D.; Carroll, R.J.; Zink, R.; Mosley, J.D.; Field, J.R.; Pulley, J.M.; Ramirez, A.H.; Bowton, E.; et al. Systematic Comparison of Phenome-Wide Association Study of Electronic Medical Record Data and Genome-Wide Association Study Data. Nat. Biotechnol. 2013, 31, 1102–1111. [Google Scholar] [CrossRef]
- Lage, K.; Karlberg, E.O.; Størling, Z.M.; Ólason, P.Í.; Pedersen, A.G.; Rigina, O.; Hinsby, A.M.; Tümer, Z.; Pociot, F.; Tommerup, N.; et al. A Human Phenome-Interactome Network of Protein Complexes Implicated in Genetic Disorders. Nat. Biotechnol. 2007, 25, 309–316. [Google Scholar] [CrossRef]
- Home—Genetic and Rare Diseases Information Center. Available online: https://rarediseases.info.nih.gov/ (accessed on 11 February 2023).
- Elliott, L.T.; Sharp, K.; Alfaro-Almagro, F.; Shi, S.; Miller, K.L.; Douaud, G.; Marchini, J.; Smith, S.M. Genome-Wide Association Studies of Brain Imaging Phenotypes in UK Biobank. Nature 2018, 562, 210–216. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef]
- Annes, J.P.; Munger, J.S.; Rifkin, D.B. Making Sense of Latent TGFβ Activation. J. Cell Sci. 2003, 116, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Muise, E.S.; Iyengar, P.; Wang, Z.V.; Chandalia, M.; Abate, N.; Zhang, B.B.; Bonaldo, P.; Chua, S.; Scherer, P.E. Metabolic Dysregulation and Adipose Tissue Fibrosis: Role of Collagen VI. Mol. Cell. Biol. 2009, 29, 1575–1591. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, D.G.K.; Hansen, T.W.; von Scholten, B.J.; Nielsen, S.H.; Reinhard, H.; Parving, H.-H.; Tepel, M.; Karsdal, M.A.; Jacobsen, P.K.; Genovese, F.; et al. Higher Collagen VI Formation Is Associated With All-Cause Mortality in Patients With Type 2 Diabetes and Microalbuminuria. Diabetes Care 2018, 41, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Rastegar-Mojarad, M.; Ye, Z.; Kolesar, J.M.; Hebbring, S.J.; Lin, S.M. Opportunities for Drug Repositioning from Phenome-Wide Association Studies. Nat. Biotechnol. 2015, 33, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Griggs, R.C.; Batshaw, M.; Dunkle, M.; Gopal-Srivastava, R.; Kaye, E.; Krischer, J.; Nguyen, T.; Paulus, K.; Merkel, P.A. Rare Diseases Clinical Research Network Clinical Research for Rare Disease: Opportunities, Challenges, and Solutions. Mol. Genet. Metab. 2009, 96, 20–26. [Google Scholar] [CrossRef]
- Shourick, J.; Wack, M.; Jannot, A.-S. Assessing Rare Diseases Prevalence Using Literature Quantification. Orphanet J. Rare Dis. 2021, 16, 139. [Google Scholar] [CrossRef]
- Nature Genetics. Rare Diseases, Common Challenges. Nat. Genet. 2022, 54, 215. [Google Scholar] [CrossRef]
- Nam, Y.; Jung, S.-H.; Yun, J.-S.; Sriram, V.; Singhal, P.; Byrska-Bishop, M.; Verma, A.; Shin, H.; Park, W.-Y.; Won, H.-H.; et al. Discovering Comorbid Diseases Using an Inter-Disease Interactivity Network Based on Biobank-Scale PheWAS Data. Bioinformatics 2023, 39, btac822. [Google Scholar] [CrossRef]
- Robinson, J.R.; Denny, J.C.; Roden, D.M.; Van Driest, S.L. Genome-wide and Phenome-wide Approaches to Understand Variable Drug Actions in Electronic Health Records. Clin. Transl. Sci. 2018, 11, 112–122. [Google Scholar] [CrossRef]
- Sticht, J.; Álvaro-Benito, M.; Konigorski, S. Type 1 Diabetes and the HLA Region: Genetic Association Besides Classical HLA Class II Genes. Front. Genet. 2021, 12, 683946. [Google Scholar] [CrossRef]
- Matsumoto, K.; Aoki, H. The Roles of Tenascins in Cardiovascular, Inflammatory, and Heritable Connective Tissue Diseases. Front. Immunol. 2020, 11, 3138. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, F.S.; Ghali, N.; Demirdas, S.; Baker, D. TNXB-Related Classical-Like Ehlers-Danlos Syndrome. In GeneReviews®; Adam, M.P., Everman, D.B., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Bao, Y.; Wang, L.; Shi, L.; Yun, F.; Liu, X.; Chen, Y.; Chen, C.; Ren, Y.; Jia, Y. Transcriptome Profiling Revealed Multiple Genes and ECM-Receptor Interaction Pathways That May Be Associated with Breast Cancer. Cell. Mol. Biol. Lett. 2019, 24, 38. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Gubbiotti, M.A. Extracellular Matrix: The Driving Force of Mammalian Diseases. Matrix Biol. J. Int. Soc. Matrix Biol. 2018, 71–72, 1–9. [Google Scholar] [CrossRef]
- Theocharis, A.D.; Manou, D.; Karamanos, N.K. The Extracellular Matrix as a Multitasking Player in Disease. FEBS J. 2019, 286, 2830–2869. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Zhang, Y.; Gu, H.; Chen, Z.; Ren, L.; Lu, X.; Chen, L.; Wang, F.; Liu, Y.; et al. X-Linked Alport Syndrome: Pathogenic Variant Features and Further Auditory Genotype-Phenotype Correlations in Males. Orphanet J. Rare Dis. 2018, 13, 229. [Google Scholar] [CrossRef]
- Dietz, H. FBN1-Related Marfan Syndrome. In GeneReviews®; Adam, M.P., Everman, D.B., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- What Causes Osteogenesis Imperfecta (OI)? Available online: https://www.nichd.nih.gov/health/topics/osteogenesisimp/conditioninfo/causes (accessed on 14 February 2023).
- Long, K.R.; Huttner, W.B. The Role of the Extracellular Matrix in Neural Progenitor Cell Proliferation and Cortical Folding During Human Neocortex Development. Front. Cell. Neurosci. 2022, 15, 563. [Google Scholar] [CrossRef]
- Coban, N.; Gokcen, C.; Akbayram, S.; Calisgan, B. Evaluation of Platelet Parameters in Children with Autism Spectrum Disorder: Elongated Collagen-Adenosine Diphosphate and Collagen-Epinephrine Closure Times. Autism Res. 2019, 12, 1069–1076. [Google Scholar] [CrossRef]
- Casanova, E.L.; Baeza-Velasco, C.; Buchanan, C.B.; Casanova, M.F. The Relationship between Autism and Ehlers-Danlos Syndromes/Hypermobility Spectrum Disorders. J. Pers. Med. 2020, 10, 260. [Google Scholar] [CrossRef]
- Glans, M.R.; Thelin, N.; Humble, M.B.; Elwin, M.; Bejerot, S. The Relationship Between Generalised Joint Hypermobility and Autism Spectrum Disorder in Adults: A Large, Cross-Sectional, Case Control Comparison. Front. Psychiatry 2022, 12, 2622. [Google Scholar] [CrossRef]
- Dwyer, C.A.; Esko, J.D. Glycan Susceptibility Factors in Autism Spectrum Disorders. Mol. Aspects Med. 2016, 51, 104–114. [Google Scholar] [CrossRef]
- Berretta, S. Extracellular Matrix Abnormalities in Schizophrenia. Neuropharmacology 2012, 62, 1584–1597. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, B.R.; Smit, A.B.; Spijker, S.; van den Oever, M.C. Chapter 12—Neural ECM in Addiction, Schizophrenia, and Mood Disorder. In Progress in Brain Research; Brain Extracellular Matrix in Health and Disease; Dityatev, A., Wehrle-Haller, B., Pitkänen, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 214, pp. 263–284. [Google Scholar]
- Pantazopoulos, H.; Katsel, P.; Haroutunian, V.; Chelini, G.; Klengel, T.; Berretta, S. Molecular Signature of Extracellular Matrix Pathology in Schizophrenia. Eur. J. Neurosci. 2021, 53, 3960–3987. [Google Scholar] [CrossRef]
- Pantazopoulos, H.; Woo, T.-U.W.; Lim, M.P.; Lange, N.; Berretta, S. Extracellular Matrix-Glial Abnormalities in the Amygdala and Entorhinal Cortex of Subjects Diagnosed With Schizophrenia. Arch. Gen. Psychiatry 2010, 67, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Amorim, D.; Rivera-Baltanás, T.; Fernández-Palleiro, P.; Iglesias-Martínez-Almeida, M.; Freiría-Martínez, L.; Jarmardo-Rodriguez, C.; del Carmen Vallejo-Curto, M.; Álvarez-Ariza, M.; López-García, M.; de las Heras, E.; et al. Changes in the Brain Extracellular Matrix Composition in Schizophrenia: A Pathophysiological Dysregulation and a Potential Therapeutic Target. Cell. Mol. Neurobiol. 2022, 42, 1921–1932. [Google Scholar] [CrossRef]
- Sethi, M.K.; Zaia, J. Extracellular Matrix Proteomics in Schizophrenia and Alzheimer’s Disease. Anal. Bioanal. Chem. 2017, 409, 379–394. [Google Scholar] [CrossRef]
- Downs, M.; Sethi, M.K.; Raghunathan, R.; Layne, M.D.; Zaia, J. Matrisome Changes in Parkinson’s Disease. Anal. Bioanal. Chem. 2022, 414, 3005–3015. [Google Scholar] [CrossRef]
- Gregorio, I.; Braghetta, P.; Bonaldo, P.; Cescon, M. Collagen VI in Healthy and Diseased Nervous System. Dis. Model. Mech. 2018, 11, dmm032946. [Google Scholar] [CrossRef]
- Hicks, D.; Farsani, G.T.; Laval, S.; Collins, J.; Sarkozy, A.; Martoni, E.; Shah, A.; Zou, Y.; Koch, M.; Bonnemann, C.G.; et al. Mutations in the Collagen XII Gene Define a New Form of Extracellular Matrix-Related Myopathy. Hum. Mol. Genet. 2014, 23, 2353–2363. [Google Scholar] [CrossRef]
- Sarbacher, C.A.; Halper, J.T. Connective Tissue and Age-Related Diseases. In Biochemistry and Cell Biology of Ageing: Part II Clinical Science; Subcellular Biochemistry; Harris, J.R., Korolchuk, V.I., Eds.; Springer: Singapore, 2019; pp. 281–310. ISBN 9789811336812. [Google Scholar]
- Romero-Ortuno, R.; Kenny, R.A.; McManus, R. Collagens and Elastin Genetic Variations and Their Potential Role in Aging-Related Diseases and Longevity in Humans. Exp. Gerontol. 2020, 129, 110781. [Google Scholar] [CrossRef]
- Richards, A.; Narcisi, P.; Lloyd, J.; Ferguson, C.; Pope, F.M. The Substitution of Glycine 661 by Arginine in Type III Collagen Produces Mutant Molecules with Different Thermal Stabilities and Causes Ehlers-Danlos Syndrome Type IV. J. Med. Genet. 1993, 30, 690–693. [Google Scholar] [CrossRef]
- Kawai, S.; Nomura, S.; Harano, T.; Harano, K.; Fukushima, T.; Wago, M.; Shimizu, B.; Osawa, G. A Single-Base Mutation in Exon 31 Converting Glycine 852 to Arginine in the Collagenous Domain in an Alport Syndrome Patient. Nephron 1996, 74, 333–336. [Google Scholar] [CrossRef]
- Knebelmann, B.; Deschenes, G.; Gros, F.; Hors, M.C.; Grünfeld, J.P.; Zhou, J.; Tryggvason, K.; Gubler, M.C.; Antignac, C. Substitution of Arginine for Glycine 325 in the Collagen Alpha 5 (IV) Chain Associated with X-Linked Alport Syndrome: Characterization of the Mutation by Direct Sequencing of PCR-Amplified Lymphoblast CDNA Fragments. Am. J. Hum. Genet. 1992, 51, 135–142. [Google Scholar] [PubMed]
- Christiano, A.M.; Morricone, A.; Paradisi, M.; Angelo, C.; Mazzanti, C.; Cavalieri, R.; Uitto, J. A Glycine-to-Arginine Substitution in the Triple-Helical Domain of Type VII Collagen in a Family with Dominant Dystrophic Epidermolysis Bullosa. J. Investig. Dermatol. 1995, 104, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Deak, S.B.; Scholz, P.M.; Amenta, P.S.; Constantinou, C.D.; Levi-Minzi, S.A.; Gonzalez-Lavin, L.; Mackenzie, J.W. The Substitution of Arginine for Glycine 85 of the Alpha 1(I) Procollagen Chain Results in Mild Osteogenesis Imperfecta. The Mutation Provides Direct Evidence for Three Discrete Domains of Cooperative Melting of Intact Type I Collagen. J. Biol. Chem. 1991, 266, 21827–21832. [Google Scholar] [CrossRef] [PubMed]
- Wenstrup, R.J.; Cohn, D.H.; Cohen, T.; Byers, P.H. Arginine for Glycine Substitution in the Triple-Helical Domain of the Products of One Alpha 2(I) Collagen Allele (COL1A2) Produces the Osteogenesis Imperfecta Type IV Phenotype. J. Biol. Chem. 1988, 263, 7734–7740. [Google Scholar] [CrossRef]
- Katz, J.M.; Tadi, P. Physiology, Plasminogen Activation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- North, B.J.; Sinclair, D.A. The Intersection Between Aging and Cardiovascular Disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef]
- Wilkerson, W.R.; Sane, D.C. Aging and Thrombosis. Semin. Thromb. Hemost. 2002, 28, 555–568. [Google Scholar] [CrossRef]
- Ohuchi, K.; Amagai, R.; Ikawa, T.; Muto, Y.; Roh, Y.; Endo, J.; Maekawa, T.; Kambayashi, Y.; Asano, Y.; Fujimura, T. Plasminogen Activating Inhibitor-1 Promotes Angiogenesis in Cutaneous Angiosarcomas. Exp. Dermatol. 2023, 32, 50–59. [Google Scholar] [CrossRef]
- Kohler, H.P.; Grant, P.J. Plasminogen-Activator Inhibitor Type 1 and Coronary Artery Disease. N. Engl. J. Med. 2000, 342, 1792–1801. [Google Scholar] [CrossRef]
- Yamamoto, K.; Takeshita, K.; Kojima, T.; Takamatsu, J.; Saito, H. Aging and Plasminogen Activator Inhibitor-1 (PAI-1) Regulation: Implication in the Pathogenesis of Thrombotic Disorders in the Elderly. Cardiovasc. Res. 2005, 66, 276–285. [Google Scholar] [CrossRef]
- Gerckens, M.; Alsafadi, H.; Wagner, D.; Heinzelmann, K.; Schorpp, K.; Hadian, K.; Lindner, M.; Behr, J.; Königshoff, M.; Eickelberg, O.; et al. High-Throughput Drug Screening of ECM Deposition Inhibitors for Antifibrotic Drug Discovery. Pneumologie 2019, 73, 113–114. [Google Scholar]
- Khomtchouk, B.B.; Lee, Y.S.; Khan, M.L.; Sun, P.; Mero, D.; Davidson, M.H. Targeting the Cytoskeleton and Extracellular Matrix in Cardiovascular Disease Drug Discovery. Expert Opin. Drug Discov. 2022, 17, 443–460. [Google Scholar] [CrossRef]
- Parker, A.L.; Bowman, E.; Zingone, A.; Ryan, B.M.; Cooper, W.A.; Kohonen-Corish, M.; Harris, C.C.; Cox, T.R. Extracellular Matrix Profiles Determine Risk and Prognosis of the Squamous Cell Carcinoma Subtype of Non-Small Cell Lung Carcinoma. Genome Med. 2022, 14, 126. [Google Scholar] [CrossRef]
- Lamb, C.A.; Saifuddin, A.; Powell, N.; Rieder, F. The Future of Precision Medicine to Predict Outcomes and Control Tissue Remodeling in Inflammatory Bowel Disease. Gastroenterology 2022, 162, 1525–1542. [Google Scholar] [CrossRef]
- Moretti, L.; Bizzoca, D.; Geronimo, A.; Moretti, F.L.; Monaco, E.; Solarino, G.; Moretti, B. Towards Precision Medicine for Osteoarthritis: Focus on the Synovial Fluid Proteome. Int. J. Mol. Sci. 2022, 23, 9731. [Google Scholar] [CrossRef]
- Xu, R.; Zhou, X.; Wang, S.; Trinkle, C. Tumor Organoid Models in Precision Medicine and Investigating Cancer-Stromal Interactions. Pharmacol. Ther. 2021, 218, 107668. [Google Scholar] [CrossRef]
- Lumibao, J.C.; Okhovat, S.R.; Peck, K.; Lande, K.; Zou, J.; Engle, D.D. The Impact of Extracellular Matrix on the Precision Medicine Utility of Pancreatic Cancer Patient-Derived Organoids. bioRxiv 2023. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Statzer, C.; Luthria, K.; Sharma, A.; Kann, M.G.; Ewald, C.Y. The Human Extracellular Matrix Diseasome Reveals Genotype–Phenotype Associations with Clinical Implications for Age-Related Diseases. Biomedicines 2023, 11, 1212. https://doi.org/10.3390/biomedicines11041212
Statzer C, Luthria K, Sharma A, Kann MG, Ewald CY. The Human Extracellular Matrix Diseasome Reveals Genotype–Phenotype Associations with Clinical Implications for Age-Related Diseases. Biomedicines. 2023; 11(4):1212. https://doi.org/10.3390/biomedicines11041212
Chicago/Turabian StyleStatzer, Cyril, Karan Luthria, Arastu Sharma, Maricel G. Kann, and Collin Y. Ewald. 2023. "The Human Extracellular Matrix Diseasome Reveals Genotype–Phenotype Associations with Clinical Implications for Age-Related Diseases" Biomedicines 11, no. 4: 1212. https://doi.org/10.3390/biomedicines11041212