
The Emergence of TRP Channels Interactome as a Potential Therapeutic Target in Pancreatic Ductal Adenocarcinoma


 ,
,
Abstract
:1. Introduction
2. Pancreatic Ductal Adenocarcinoma (PDAC)
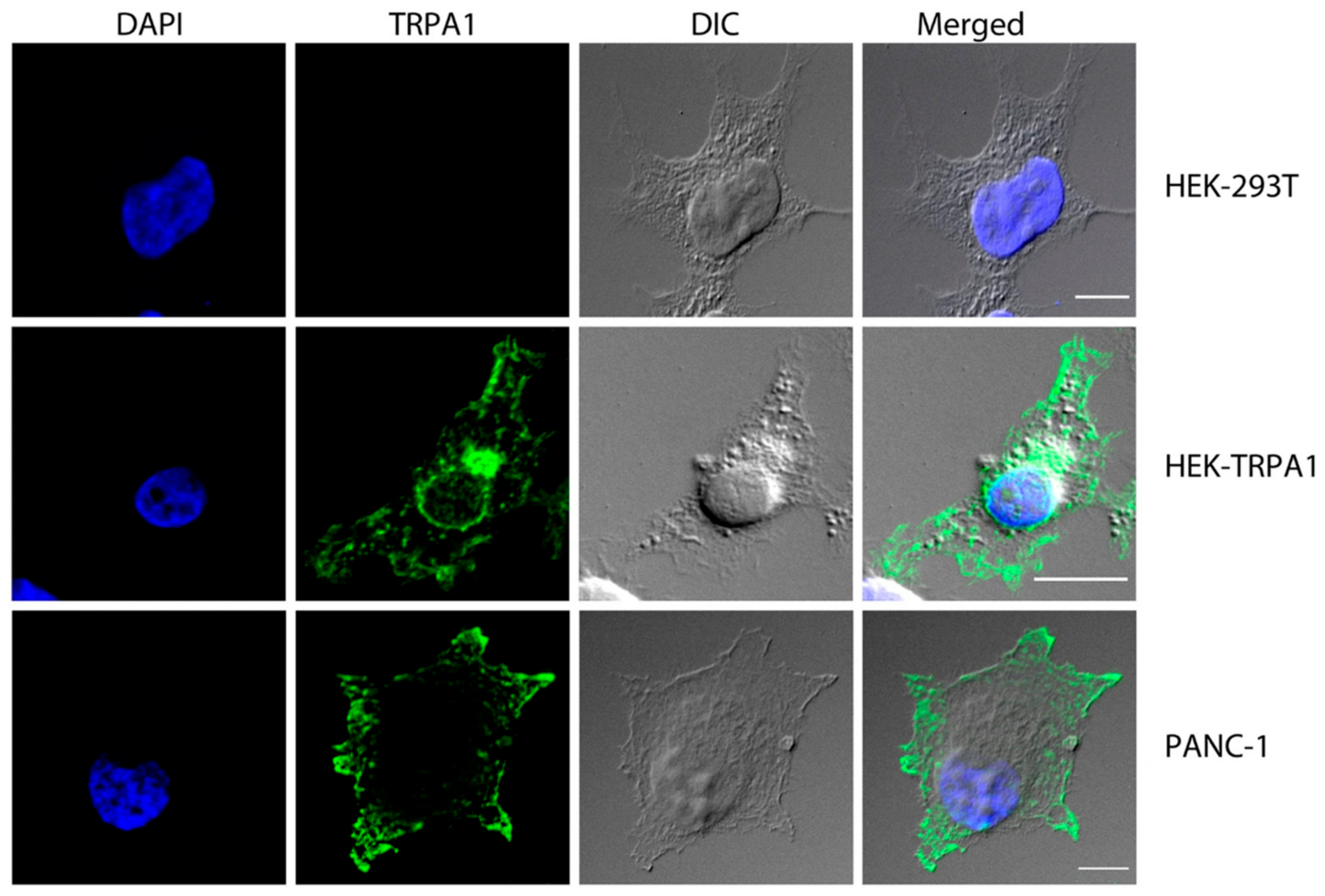
3. TRP Channels in Pancreatic Ductal Adenocarcinoma (PDAC)

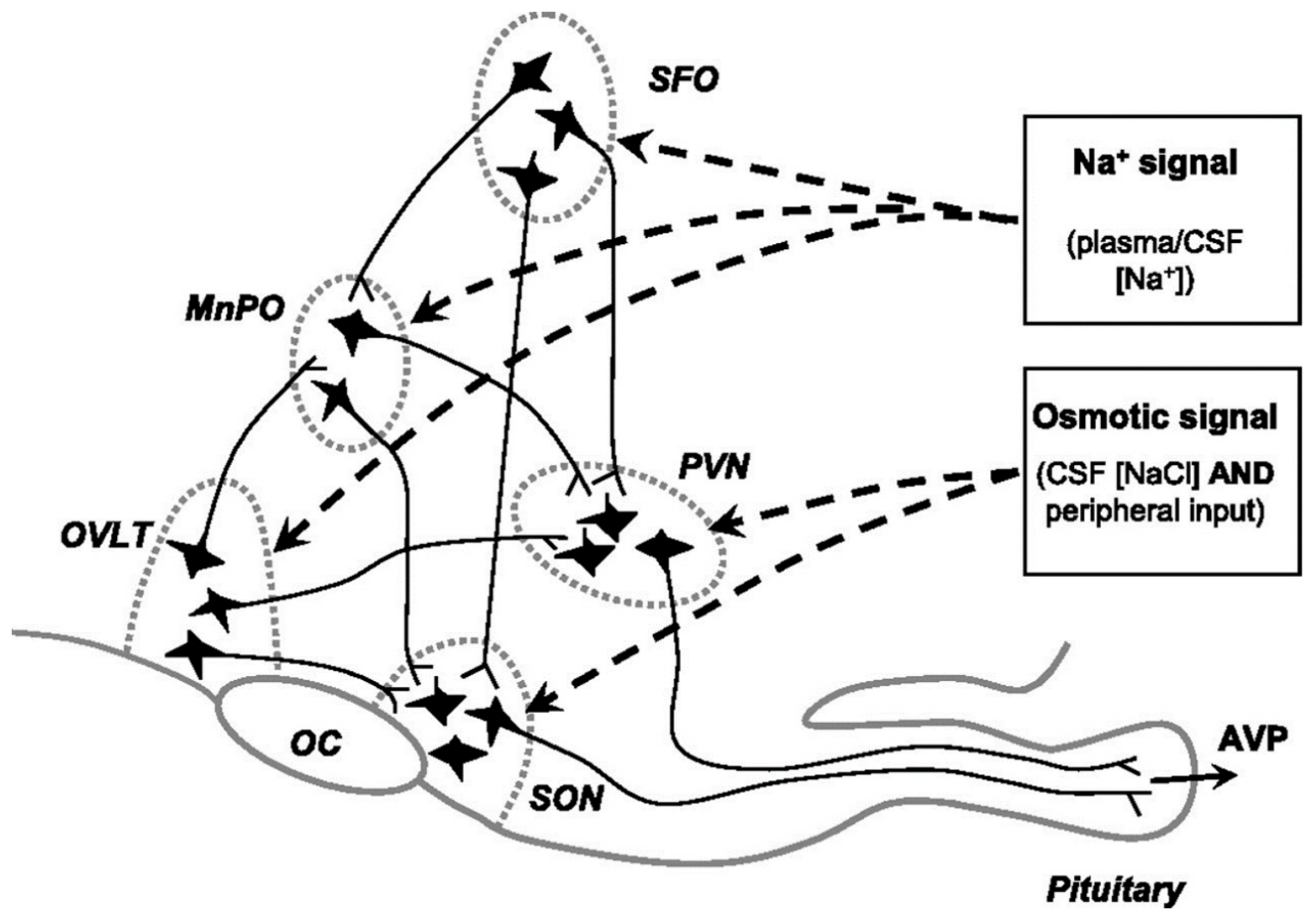
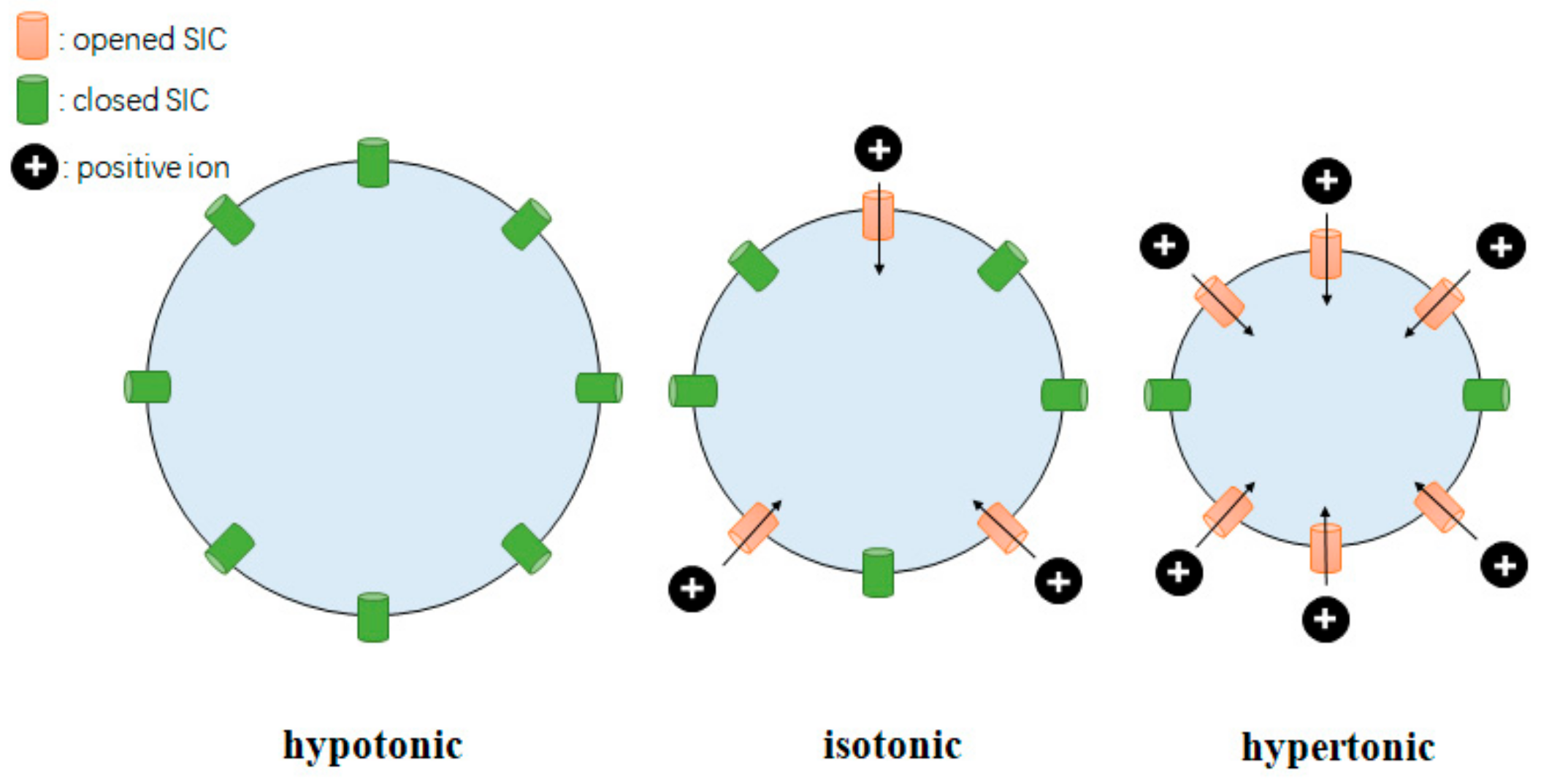
3.1. The Distribution of Osmoreceptors
3.2. Osmosensitive Neurons Are Associated with TRPV1
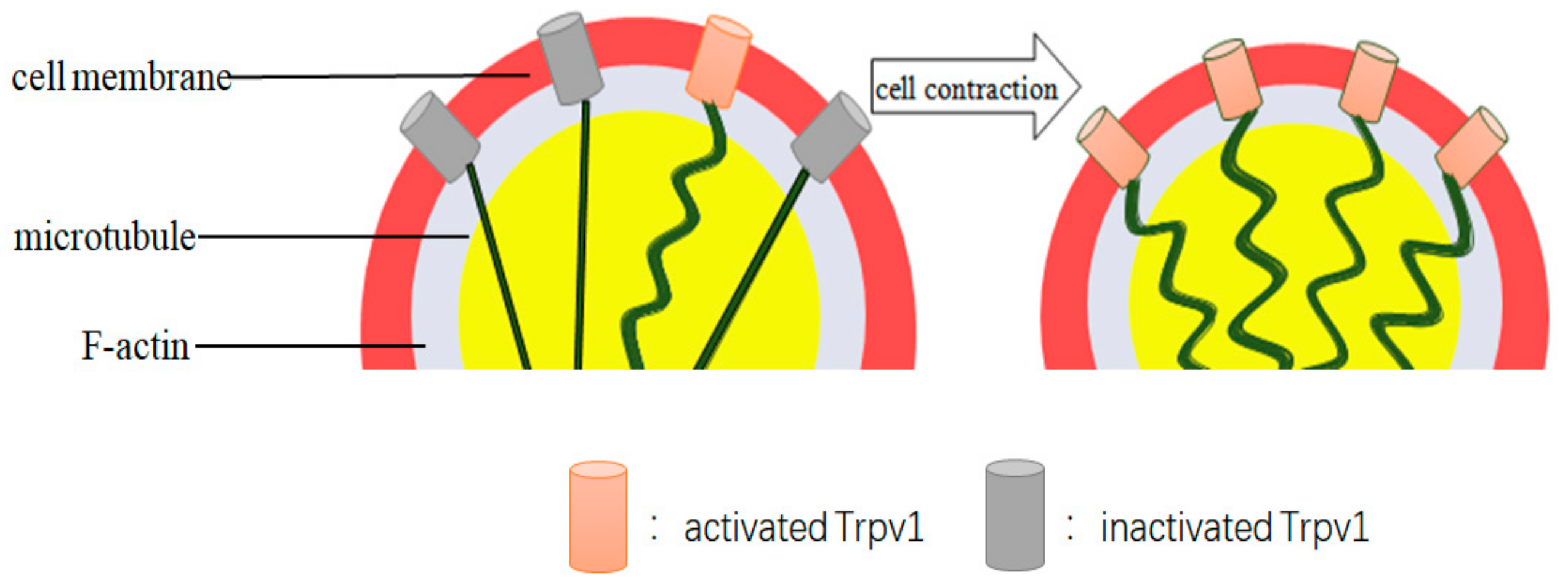
3.3. ΔThe n-TRPV1 Channel Senses Cell Volume Changes Induced by Changes in Osmotic Pressure
3.4. Nax/TRPV 4 Pathway
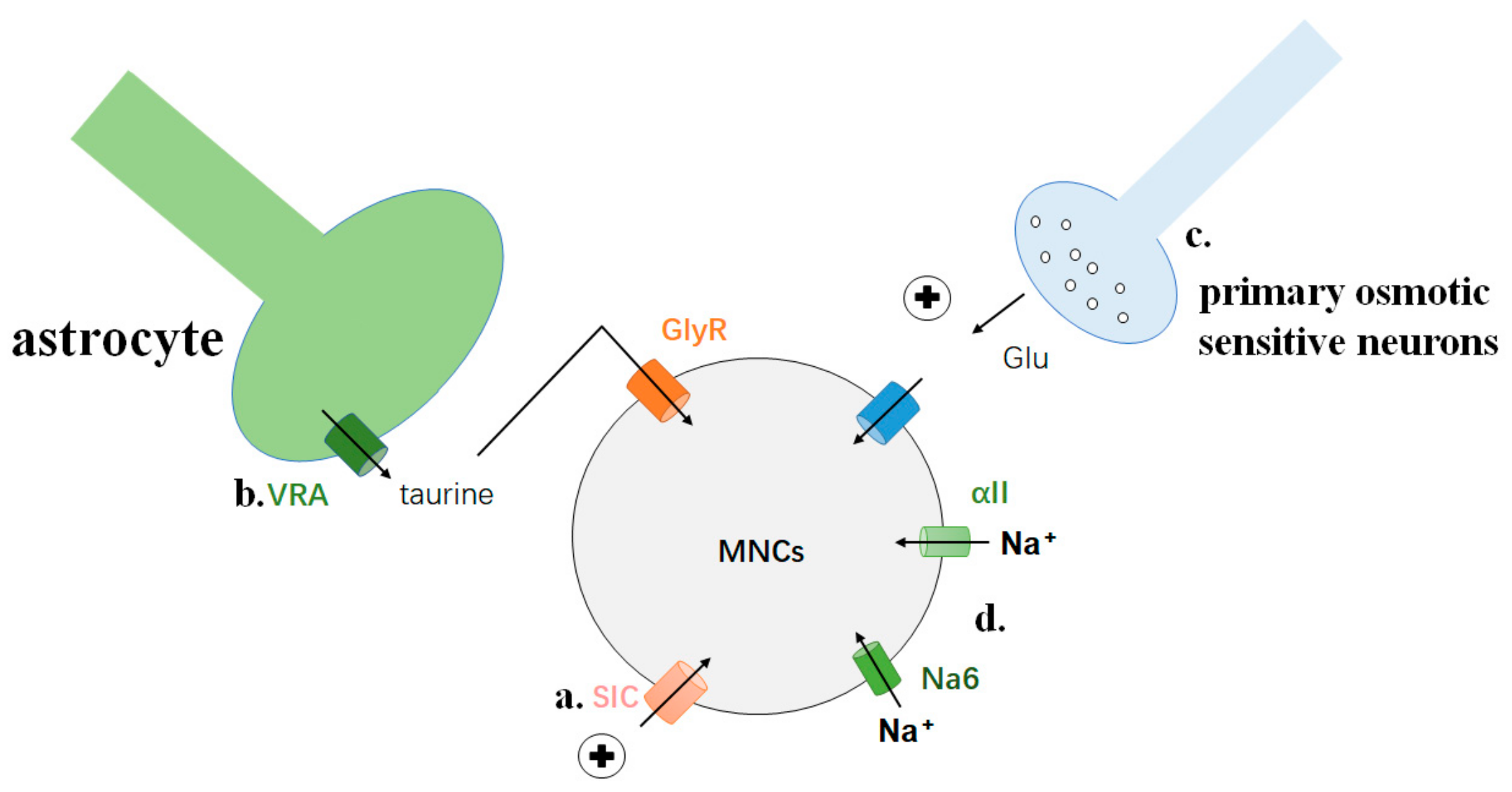
3.5. SLC9A4 Is Sensitive to [Na+] Changes
3.6. AngII Regulates the Excitatory Response of MNCs to Hypertonic Stimulation
3.7. The Interaction between TRPV1 and BCL2
4. Outcome and Implications
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rahib, L.; Wehner, M.R.; Matrisian, L.M.; Nead, K.T. Estimated projection of US cancer incidence and death to 2040. JAMA Netw. Open 2021, 4, e214708. [Google Scholar] [CrossRef]
- Tonini, V.; Zanni, M. Pancreatic cancer in 2021: What you need to know to win. World J. Gastroenterol. 2021, 27, 5851. [Google Scholar] [CrossRef]
- Hu, J.X.; Zhao, C.F.; Chen, W.B.; Liu, Q.C.; Li, Q.W.; Lin, Y.Y.; Gao, F. Pancreatic cancer: A review of epidemiology, trend, and risk factors. World J. Gastroenterol. 2021, 27, 4298. [Google Scholar] [CrossRef] [PubMed]
- Prevarskaya, N.; Zhang, L.; Barritt, G. TRP channels in cancer. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2007, 1772, 937–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokłosa, P.; Borgström, A.; Kappel, S.; Peinelt, C. TRP channels in digestive tract cancers. Int. J. Mol. Sci. 2020, 21, 1877. [Google Scholar] [CrossRef] [Green Version]
- Mesquita, G.; Prevarskaya, N.; Schwab, A.; Lehen’kyi, V.Y. Role of the TRP channels in pancreatic ductal adenocarcinoma development and progression. Cells 2021, 10, 1021. [Google Scholar] [CrossRef] [PubMed]
- Chelaru, N.R.; Chiosa, A.; Sorop, A.; Spiridon, A.; Cojocaru, F.; Domocos, D.; Dima, S.O. The association between TRP channels expression and Clinicopathological characteristics of patients with pancreatic adenocarcinoma. Int. J. Mol. Sci. 2022, 23, 9045. [Google Scholar] [CrossRef]
- Luo, D.; Liu, Y.; Li, Z.; Zhu, H.; Yu, X. NR2F1-AS1 Promotes Pancreatic Ductal Adenocarcinoma Progression Through Competing Endogenous RNA Regulatory Network Constructed by Sponging miRNA-146a-5p/miRNA-877-5p. Front. Cell Dev. Biol. 2021, 9, 736980. [Google Scholar] [CrossRef]
- Adamska, A.; Domenichini, A.; Falasca, M. Pancreatic ductal adenocarcinoma: Current and evolving therapies. Int. J. Mol. Sci. 2017, 18, 1338. [Google Scholar] [CrossRef] [PubMed]
- Hezel, A.F.; Kimmelman, A.C.; Stanger, B.Z.; Bardeesy, N.; DePinho, R.A. Genetics and biology of pancreatic ductal adenocarcinoma. Genes Dev. 2006, 20, 1218–1249. [Google Scholar] [CrossRef] [Green Version]
- Ying, H.; Dey, P.; Yao, W.; Kimmelman, A.C.; Draetta, G.F.; Maitra, A.; DePinho, R.A. Genetics and biology of pancreatic ductal adenocarcinoma. Genes Dev. 2016, 30, 355–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, Y.; Hu, X.; Fan, X.; Li, Y. The potential of TRP channels as therapeutic targets in pancreatic cancer. Oncotarget 2018, 9, 14775–14786. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, X.; Sun, Y.; Wang, Q.; Chen, W. The role of TRP channels in pancreatic cancer progression and therapeutic implications. J. Cell. Physiol. 2018, 233, 8113–8120. [Google Scholar] [CrossRef]
- Li, H. TRP Channel Classification. In Transient Receptor Potential Canonical Channels and Brain Diseases; Springer: Dordrecht, The Netherlands, 2017; pp. 1–8. [Google Scholar]
- Samanta, A.; Hughes, T.E.; Moiseenkova-Bell, V.Y. Transient Receptor Potential (TRP) Channels. In Membrane Protein Complexes: Structure and Function; Springer: Singapore, 2018; pp. 141–165. [Google Scholar]
- Mulier, M.; Vriens, J.; Voets, T. TRP channel pores and local calcium signals. Cell Calcium 2017, 66, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Shah SM, A.; Ou, Y.Y. TRP-BERT: Discrimination of transient receptor potential (TRP) channels using contextual representations from deep bidirectional transformer based on BERT. Comput. Biol. Med. 2021, 137, 104821. [Google Scholar] [CrossRef] [PubMed]
- Kiss, F.; Pohóczky, K.; Szállási, A.; Helyes, Z. Transient receptor potential (TRP) channels in head-and-neck squamous cell carcinomas: Diagnostic, prognostic, and therapeutic potentials. Int. J. Mol. Sci. 2020, 21, 6374. [Google Scholar] [CrossRef]
- Vafaee, F.; Shirzad, S.; Shamsi, F.; Boskabady, M.H. Neuroscience and treatment of asthma, new therapeutic strategies and future aspects. Life Sci. 2021, 292, 120175. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, A.S.; Rigo, F.K.; De Prá, S.D.-T.; Milioli, A.M.; Pereira, G.C.; Lückemeyer, D.D.; Antoniazzi, C.T.; Kudsi, S.Q.; Araújo, D.M.P.A.; Oliveira, S.M.; et al. Role of transient receptor potential ankyrin 1 (TRPA1) on nociception caused by a murine model of breast carcinoma. Pharmacol. Res. 2020, 152, 104576. [Google Scholar] [CrossRef]
- Liu, J.J.; Li, L.Z.; Xu, P. Upregulation of TRPM8 can promote the colon cancer liver metastasis through mediating Akt/GSK-3 signal pathway. Biotechnol. Appl. Biochem. 2021, 69, 230–239. [Google Scholar] [CrossRef]
- Verney, E.B. The antidiuretic hormone and the factors which determine its release. Proc. R. Soc. London. Ser. B Boil. Sci. 1947, 135, 25–106. [Google Scholar]
- Ramsay, D.J.; Thrasher, T.N.; Keil, L.C. The organum vasculosum laminae terminalis: A critical area for osmoreception. Prog. Brain Res. 1983, 60, 91–98. [Google Scholar] [PubMed]
- McKinley, M.J.; Mathai, M.L.; McAllen, R.M.; McClear, R.C.; Miselis, R.R.; Pennington, G.L.; Vivas, L.; Wade, J.D.; Oldfield, B.J. Vasopressin secretion: Osmotic and hormonal regulation by the lamina terminalis. J. Neuroendocrinol. 2004, 16, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Hussy, N.; Deleuze, C.; Desarménien, M.G.; Moos, F.C. Osmotic regulation of neuronal activity: A new role for taurine and glial cells in a hypothalamic neuroendocrine structure. Prog. Neurobiol. 2000, 62, 113–134. [Google Scholar] [CrossRef]
- Wilson, Y.; Nag, N.; Davern, P.; Oldfield, B.J.; McKinley, M.J.; Greferath, U.; Murphy, M. Visualization of functionally activated circuitry in the brain. Proc. Natl. Acad. Sci. USA 2002, 99, 3252–3257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, K.; Kolmodin, R. Dependence of basic secretion of antidiuretic hormone on cerebrospinal fluid (Na+). Acta Physiol Scand. 1974, 91, 286–288. [Google Scholar] [CrossRef]
- Osborne, P.G.; Denton, D.A.; Weisinger, R.S. Effect of variation of the composition of CSF in the rat upon drinking of water and hypertonic NaCl solutions. Behav. Neurosci. 1987, 101, 371–377. [Google Scholar] [CrossRef]
- Mckinley, M.; Congiu, M.; Denton, D.; Lichardus, B.; Weisinger, R. Cerebrospinal Fluid Composition and Homeostatic Responses to Dehydration; Raven Press: New York, NY, USA, 1985; pp. 299–309. [Google Scholar]
- Zimmerman, C.A.; Leib, D.E.; Knight, Z.A. Neural circuits underlying thirst and fluid homeostasis. Nat. Rev. Neurosci. 2017, 18, 459–469. [Google Scholar] [CrossRef]
- McKinley, M.J.; Blaine, E.H.; Denton, D.A. Brain osmoreceptors, cerebrospinal fluid electrolyte composition and thirst. Brain Res. 1974, 70, 532–537. [Google Scholar] [CrossRef]
- McKinley, M.J.; Denton, D.A.; Weisinger, R.S. Sensors for antidiuresis and thirst--osmoreceptors or CSF sodium detectors? Brain Res. 1978, 141, 89–103. [Google Scholar] [CrossRef]
- Sakuta, H.; Nishihara, E.; Hiyama, T.Y.; Lin, C.H.; Noda, M. Nax signaling evoked by an increase in [Na+] in CSF induces water intake via EET-mediated TRPV4 activation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R299–R306. [Google Scholar] [CrossRef] [Green Version]
- Bourque, C.W.; Oliet, S.H.; Richard, D. Osmoreceptors, osmoreception, and osmoregulation. Front. Neuroendocrinol. 1994, 15, 231–274. [Google Scholar] [CrossRef]
- Zhang, Z.; Kindrat, A.N.; Sharif-Naeini, R.; Bourque, C.W. Actin filaments mediate mechanical gating during osmosensory transduction in rat supraoptic nucleus neurons. J. Neurosci. 2007, 27, 4008–4013. [Google Scholar] [CrossRef] [Green Version]
- Prager-Khoutorsky, M.; Bourque, C.W. Mechanical basis of osmosensory transduction in magnocellular neurosecretory neurones of the rat supraoptic nucleus. J. Neuroendocrinol. 2015, 27, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Prager-Khoutorsky, M.; Khoutorsky, A.; Bourque, C.W. Unique interweaved microtubule scaffold mediates osmosensory transduction via physical interaction with TRPV1. Neuron 2014, 83, 866–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciura, S.; Bourque, C.W. Transient receptor potential vanilloid 1 is required for intrinsic osmoreception in organum vasculosum lamina terminalis neurons and for normal thirst responses to systemic hyperosmolality. J. Neurosci. 2006, 26, 9069–9075. [Google Scholar] [CrossRef] [Green Version]
- Cojocaru, F.; Şelescu, T.; Domocoş, D.; Măruţescu, L.; Chiritoiu, G.; Chelaru, N.R.; Dima, S.; Mihăilescu, D.; Babes, A.; Cucu, D. Functional expression of the transient receptor potential ankyrin type 1 channel in pancreatic adenocarcinoma cells. Sci. Rep. 2021, 11, 2018. [Google Scholar] [CrossRef]
- Hendry, W.H. Kidney Preservation. In Scientific Foundation of Urology; Williams, D.I., Chisholm, G.D., Eds.; William Heinemann Medical Books: London, UK, 1976; pp. 142–150. [Google Scholar]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif-Naeini, R.; Ciura, S.; Zhang, Z.; Bourque, C.W. Contribution of TRPV channels to osmosensory transduction, thirst, and vasopressin release. Kidney Int. 2008, 73, 811–815. [Google Scholar] [CrossRef] [Green Version]
- Ciura, S.; Liedtke, W.; Bourque, C.W. Hypertonicity sensing in organum vasculosum lamina terminalis neurons: A mechanical process involving TRPV1 but not TRPV4. J. Neurosci. 2011, 31, 14669–14676. [Google Scholar] [CrossRef] [Green Version]
- Sharif Naeini, R.; Witty, M.F.; Séguéla, P.; Bourque, C.W. An N-terminal variant of Trpv1 channel is required for osmosensory transduction. Nat. Neurosci. 2006, 9, 93–98. [Google Scholar] [CrossRef]
- Zaelzer, C.; Hua, P.; Prager-Khoutorsky, M.; Ciura, S.; Voisin, D.L.; Liedtke, W.; Bourque, C.W. ΔN-TRPV1: A Molecular Co-detector of Body Temperature and Osmotic Stress. Cell Rep. 2015, 13, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Chakfe, Y.; Bourque, C.W. Excitatory peptides and osmotic pressure modulate mechanosensitive cation channels in concert. Nat. Neurosci. 2000, 3, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Sudbury, J.R.; Ciura, S.; Sharif-Naeini, R.; Bourque, C.W. Osmotic and thermal control of magnocellular neurosecretory neurons–role of an N-terminal variant of trpv1. Eur. J. Neurosci. 2010, 32, 2022–2030. [Google Scholar] [CrossRef]
- Le-Corronc, H.; Rigo, J.M.; Branchereau, P.; Legendre, P. GABA A receptor and glycine receptor activation by paracrine/autocrine release of endogenous agonists: More than a simple communication pathway. Mol. Neurobiol. 2011, 44, 28–52. [Google Scholar] [CrossRef] [PubMed]
- Ciura, S.; Prager-Khoutorsky, M.; Thirouin, Z.S.; Wyrosdic, J.C.; Olson, J.E.; Liedtke, W.; Bourque, C.W. Trpv4 mediates hypotonic inhibition of central osmosensory neurons via taurine gliotransmission. Cell Rep. 2018, 23, 2245–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dib-Hajj, S.D.; Black, J.A.; Waxman, S.G. NaV1. 9: A sodium channel linked to human pain. Nat. Rev. Neurosci. 2015, 16, 511–519. [Google Scholar] [CrossRef]
- Jiao, R.; Cui, D.; Wang, S.C.; Li, D.; Wang, Y.F. Interactions of the Mechanosensitive Channels with Extracellular Matrix, Integrins, and Cytoskeletal Network in Osmosensation. Front Mol. Neurosci. 2017, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.F.; Parpura, V. Astroglial Modulation of Hydromineral Balance and Cerebral Edema. Front Mol. Neurosci. 2018, 11, 204. [Google Scholar] [CrossRef] [Green Version]
- Prager-Khoutorsky, M. Mechanosensing in Hypothalamic Osmosensory Neurons. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2017; Volume 71, pp. 13–21. [Google Scholar]
- Park, S.J.; Haan, K.D.; Nakamura, Y.; Fukami, K.; Fisher, T.E. PLCδ1 Plays Central Roles in the Osmotic Activation of ΔN-TRPV1 Channels in Mouse Supraoptic Neurons and in Murine Osmoregulation. J. Neurosci. 2021, 41, 3579–3587. [Google Scholar] [CrossRef]
- Bichet, D.G. Physiopathology of hereditary polyuric states: A molecular view of renal function. Swiss Med. Wkly. 2012, 142, w13613. [Google Scholar] [CrossRef]
- Anderson, J.W.; Smith, P.M.; Ferguson, A.V. Subfornical organ neurons projecting to paraventricular nucleus: Whole-cell properties. Brain Res. 2001, 921, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Grob, M.; Drolet, G.; Mouginot, D. Specific Na+ sensors are functionally expressed in a neuronal population of the median preoptic nucleus of the rat. J. Neurosci. 2004, 24, 3974–3984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, M.; Sakuta, H. Central regulation of body-fluid homeostasis. Trends Neurosci. 2013, 36, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Hiyama, T.Y.; Yoshida, M.; Matsumoto, M.; Suzuki, R.; Matsuda, T.; Watanabe, E.; Noda, M. Endothelin-3 expression in the subfornical organ enhances the sensitivity of Na(x), the brain sodium-level sensor, to suppress salt intake. Cell Metab. 2013, 17, 507–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, H.; Watanabe, E.; Hiyama, T.Y.; Nagakura, A.; Fujikawa, A.; Okado, H.; Yanagawa, Y.; Obata, K.; Noda, M. Glial Nax channels control lactate signaling to neurons for brain [Na+] sensing. Neuron 2007, 54, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Hiyama, T.Y.; Niimura, F.; Matsusaka, T.; Fukamizu, A.; Kobayashi, K.; Kobayashi, K.; Noda, M. Distinct neural mechanisms for the control of thirst and salt appetite in the subfornical organ. Nat. Neurosci. 2017, 20, 230–241. [Google Scholar] [CrossRef]
- Ohnou, T.; Yokai, M.; Kurihara, T.; Hasegawa-Moriyama, M.; Shimizu, T.; Inoue, K.; Kambe, Y.; Kanmura, Y.; Miyata, A. Pituitary adenylate cyclase-activating polypeptide type 1 receptor signaling evokes long-lasting nociceptive behaviors through the activation of spinal astrocytes in mice. J. Pharmacol. Sci. 2016, 130, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Orlov, S.N.; Hamet, P. Salt and gene expression: Evidence for [Na+]i/[K+]i-mediated signaling pathways. Pflügers Arch.-Eur. J. Physiol. 2015, 467, 489–498. [Google Scholar] [CrossRef]
- Sakuta, H.; Lin, C.H.; Hiyama, T.Y.; Matsuda, T.; Yamaguchi, K.; Shigenobu, S.; Kobayashi, K.; Noda, M. SLC9A4 in the organum vasculosum of the lamina terminalis is a [Na+] sensor for the control of water intake. Pflugers Arch. 2020, 472, 609–624. [Google Scholar] [CrossRef]
- Nomura, K.; Hiyama, T.Y.; Sakuta, H.; Matsuda, T.; Lin, C.H.; Kobayashi, K.; Kobayashi, K.; Kuwaki, T.; Takahashi, K.; Matsui, S.; et al. [Na+] Increases in Body Fluids Sensed by Central Nax Induce Sympathetically Mediated Blood Pressure Elevations via H+-Dependent Activation of ASIC1a. Neuron 2019, 101, 60–75. [Google Scholar] [CrossRef] [Green Version]
- Tasker, J.G.; Prager-Khoutorsky, M.; Teruyama, R.; Lemos, J.R.; Amstrong, W.E. Advances in the neurophysiology of magnocellular neuroendocrine cells. J. Neuroendocrinol. 2020, 32, e12826. [Google Scholar] [CrossRef] [PubMed]
- Saffran, A. Activation of TRPV1 by Capsaicin Regulates ENaC. Master’s Thesis, VCU University, Richmond, VA, USA, 2018. [Google Scholar]
- Osswald, I. Characterization of Calcium-Permeable AMPA Receptors in the Developing Retina and Cerebellum. Ph.D. Thesis, McGill University, Montreal, QC, Canada, 2011. [Google Scholar]
- Photowala, H. G Protein-Coupled Receptors Inhibit Neurotransmitter Release by Modifying Vesicle Fusion Properties. Ph.D. Thesis, University of Illinois at Chicago, Chicago, IL, USA, 2005. [Google Scholar]
- Billeter, A.T.; Hellmann, J.L.; Bhatnagar, A.; Polk, H.C., Jr. Transient receptor potential ion channels: Powerful regulators of cell function. Ann. Surg. 2014, 259, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Rizza, S.; Montagna, C.; Di Giacomo, G.; Cirotti, C.; Filomeni, G. S-nitrosation and ubiquitin-proteasome system interplay in neuromuscular disorders. Int. J. Cell Biol. 2014, 2014, 428764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Sukumaran, P.; Schaar, A.; Sun, Y.; Singh, B.B. Functional role of TRP channels in modulating ER stress and Autophagy. Cell Calcium 2016, 60, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Damaskos, C.; Karatzas, T.; Kostakis, I.D.; Nikolidakis, L.; Kostakis, A.; Kouraklis, G. Nuclear receptors in pancreatic tumor cells. Anticancer 2014, 34, 6897–6911. [Google Scholar]
- Li, L.; Xiao, Z.; He, P.; Zou, W.; Deng, Z.; Zhang, G.; Liu, R. Molecular subtyping based on TRP family and prognostic assessment for TRP-associated lncRNAs in pancreatic adenocarcinoma. BMC Gastroenterol. 2022, 22, 454. [Google Scholar] [CrossRef]
- Fallah, H.P.; Ahuja, E.; Lin, H.; Qi, J.; He, Q.; Gao, S.; Liang, D. A Review on the Role of TRP Channels and Their Potential as Drug Targets_An Insight Into the TRP Channel Drug Discovery Methodologies. Front. Pharmacol. 2022, 13, 1784. [Google Scholar] [CrossRef]
- Taha Khalaf, A.; Song, J.Q.; Gao, T.T.; Yu, X.P.; Lei, T.C. CTLA-4 gene polymorphism and the risk of systemic lupus erythematosus in the Chinese population. J. Biomed. Biotechnol. 2011, 2011. [Google Scholar]
- Arcangeli, A.; Becchetti, A. Novel perspectives in cancer therapy: Targeting ion channels. Drug Resist. Updates 2015, 21, 11–19. [Google Scholar] [CrossRef]
- Sakuta, H.; Lin, C.H.; Yamada, M.; Kita, Y.; Tokuoka, S.M.; Shimizu, T.; Noda, M. Nax-positive glial cells in the organum vasculosum laminae terminalis produce epoxyeicosatrienoic acids to induce water intake in response to increases in [Na+] in body fluids. Neurosci. Res. 2020, 154, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, G.; Jacob, J.; Buckendahl, A.C.; Grützmann, R.; Alldinger, I.; Sipos, B.; Pilarsky, C. Peroxisome proliferator-activated receptor γ is highly expressed in pancreatic cancer and is associated with shorter overall survival times. Clin. Cancer Res. 2006, 12, 6444–6451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partida-Sanchez, S.; Desai, B.N.; Schwab, A.; Zierler, S. TRP channels in inflammation and immunity. Front. Immunol. 2021, 12, 684172. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Smani, T.; Naziroglu, M. Involvements of TRP channels, oxidative stress and apoptosis in neurodegenerative diseases. Front. Physiol. 2021, 12, 649230. [Google Scholar] [CrossRef]
- Miller, B.A. The role of TRP channels in oxidative stress-induced cell death. J. Membr. Biol. 2006, 209, 31–41. [Google Scholar] [CrossRef]
- Morelli, M.B.; Amantini, C. Transient Receptor Potential (TRP) Channels: Markers and Therapeutic Targets for Cancer? Biomolecules 2022, 12, 547. [Google Scholar] [CrossRef]
- Fels, B.; Bulk, E.; Pethő, Z.; Schwab, A. The role of TRP channels in the metastatic cascade. Pharmaceuticals 2018, 11, 48. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Luan, Y.; Yu, R.; Zhang, Z.; Zhang, J.; Wang, W. Transient receptor potential (TRP) channels, promising potential diagnostic and therapeutic tools for cancer. Biosci. Trends 2014, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Akl, H.; Vervloessem, T.; Kiviluoto, S.; Bittremieux, M.; Parys, J.B.; De Smedt, H.; Bultynck, G. A dual role for the anti-apoptotic Bcl-2 protein in cancer: Mitochondria versus endoplasmic reticulum. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2014, 1843, 2240–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, T.; Zhang, W.; Guo, H.; Pan, X.; Chen, X.; He, Q.; Ding, L. The regulatory and modulatory roles of TRP family channels in malignant tumors and relevant therapeutic strategies. Acta Pharm. Sin. B 2022, 12, 1761–1780. [Google Scholar] [CrossRef]
- Yang, D.; Kim, J. Emerging role of transient receptor potential (TRP) channels in cancer progression. BMB Rep. 2020, 53, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Rybarczyk, P.; Vanlaeys, A.; Brassart, B.; Dhennin-Duthille, I.; Chatelain, D.; Sevestre, H.; Gautier, M. The transient receptor potential melastatin 7 channel regulates pancreatic cancer cell invasion through the Hsp90α/uPA/MMP2 pathway. Neoplasia 2017, 19, 288–300. [Google Scholar] [CrossRef]
- Zhang, L.; Barritt, G.J. Evidence that TRPM8 is an androgen-dependent Ca2+ channel required for the survival of prostate cancer cells. Cancer Res. 2004, 64, 8365–8373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yee, N.S.; Zhou, W.; Lee, M.; Yee, R.K. Targeted silencing of TRPM7 ion channel induces replicative senescence and produces enhanced cytotoxicity with gemcitabine in pancreatic adenocarcinoma. Cancer Lett. 2012, 318, 99–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Iwadate, D.; Kato, H.; Nakai, Y.; Tateishi, K.; Fujishiro, M. Targeting autophagy as a therapeutic strategy against pancreatic cancer. J. Gastroenterol. 2022, 57, 603–618. [Google Scholar] [CrossRef]
- Bettaieb, L.; Brulé, M.; Chomy, A.; Diedro, M.; Fruit, M.; Happernegg, E.; Lehen’kyi, V.Y. Ca2+ Signaling and Its Potential Targeting in Pancreatic Ductal Carcinoma. Cancers 2021, 13, 3085. [Google Scholar] [CrossRef]
- Holzer, P.; Izzo, A.A. The pharmacology of TRP channels. Br. J. Pharmacol. 2014, 171, 2469–2473. [Google Scholar] [CrossRef] [Green Version]
- Ouadid-Ahidouch, H.; Dhennin-Duthille, I.; Gautier, M.; Sevestre, H.; Ahidouch, A. TRP channels: Diagnostic markers and therapeutic targets for breast cancer? Trends Mol. Med. 2013, 19, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Saldías, M.P.; Maureira, D.; Orellana-Serradell, O.; Silva, I.; Lavanderos, B.; Cruz, P.; Cerda, O. TRP channels interactome as a novel therapeutic target in breast cancer. Front. Oncol. 2021, 11, 621614. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Author and Year | Key Findings |
|---|---|---|
| [19] | Fallah H. P., et al., 2022 | TRP proteins are a large group of ion channels that control many physiological functions in the body and are considered potential therapeutic targets for various diseases, including cancers. |
| [20] | Chelaru N. R., et al., 2022 | Significantly higher expression levels of TRPA1, TRPM8, and TCAF1/F2 were found in tumoral tissues compared to normal tissues, but lower expression levels of TRPV6. The TRP channels have either tumor-suppressive or oncogenic roles. |
| [21] | Li L., et al., 2022 | Research has revealed altered expression of various TRP proteins in numerous cancer types, including PDAC. TRP ion channels are crucial in tumor formation, proliferation, and migration. TRP channel family members have been reported as good prognostic markers and potential targets for cancer drug therapy. |
| [6] | Mesquita G., et al., 2021 | Collected data indicating the TRP family has a potential role in the development and progression of PDAC. It has been found to affect both cancer and pancreatic stellate cells, impacting cell proliferation, migration, invasion, and death. The TRP family may offer new treatments and diagnostics tools for PDAC. |
| Research Direction | Challenges | Possible Solutions | References |
|---|---|---|---|
| Further investigation of TRP channel subfamilies as therapeutic targets | Lack of specificity of TRP channel modulators | Development of more selective TRP channel modulators | [84,94,95] |
| Evaluation of the role of TRP channels in pancreatic cancer progression and metastasis | Limited understanding of the mechanisms underlying TRP channel involvement in cancer | Use of genetic and pharmacological approaches to elucidate the signaling pathways involving TRP channels | [94,96] |
| Identification of TRP channel interactors in pancreatic cancer cells | Limited knowledge of TRP channel interactome in pancreatic cancer | Use of proteomic approaches to identify novel TRP channel interactors in pancreatic cancer cells | [94,97] |
| Development of TRP channel-targeted therapies for pancreatic cancer | Lack of clinical trials and approved TRP channel-targeted treatments for pancreatic cancer | Conducting preclinical studies and clinical trials to evaluate the efficacy and safety of TRP channel-targeted therapies in pancreatic cancer | [89,94] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Khalaf, A.T.; Rui, C.; Abdul Kadir, S.Y.; Zainol, J.; Oglah, Z. The Emergence of TRP Channels Interactome as a Potential Therapeutic Target in Pancreatic Ductal Adenocarcinoma. Biomedicines 2023, 11, 1164. https://doi.org/10.3390/biomedicines11041164
Wei Y, Khalaf AT, Rui C, Abdul Kadir SY, Zainol J, Oglah Z. The Emergence of TRP Channels Interactome as a Potential Therapeutic Target in Pancreatic Ductal Adenocarcinoma. Biomedicines. 2023; 11(4):1164. https://doi.org/10.3390/biomedicines11041164
Chicago/Turabian StyleWei, Yuanyuan, Ahmad Taha Khalaf, Cao Rui, Samiah Yasmin Abdul Kadir, Jamaludin Zainol, and Zahraa Oglah. 2023. "The Emergence of TRP Channels Interactome as a Potential Therapeutic Target in Pancreatic Ductal Adenocarcinoma" Biomedicines 11, no. 4: 1164. https://doi.org/10.3390/biomedicines11041164