
Cancer Is Associated with the Emergence of Placenta-Reactive Autoantibodies
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Placental Samples
2.2. Protein Extraction and Quantification
2.3. Serum Samples
- (1)
- Normal males who were referred to the Farmand medical laboratory for a routine checkup.
- (2)
- Normal pregnant women in the first and second trimesters of pregnancy with normal screening tests who were referred to the department of Fetal and Maternal Health in the Hope Generation Foundation (HGF), Tehran, Iran.
- (3)
- Normal females in the third trimester of pregnancy who were referred to the cesarean section in Kamali hospital, Karaj, Iran (Table 2).
2.4. ELISA
- L2–L3 correction: the average OD of control wells containing only pooled serum (non-cancer controls and cancer patients separately) and conjugate (without coating layer) were subtracted from the OD of each sample.
- TP/FTP correction: the OD of control wells containing only the TP coating layer and conjugate (without serum layer) was divided by the OD of wells containing the FTP coating layer and conjugate to obtain the TP correction index. The OD of wells containing TP as a coating layer was then multiplied by this index.
- Internal control correction: the average OD of internal controls in all ELISA runs was divided by the average OD of internal controls in each ELISA run and the resulting value was multiplied by the OD of each test well.
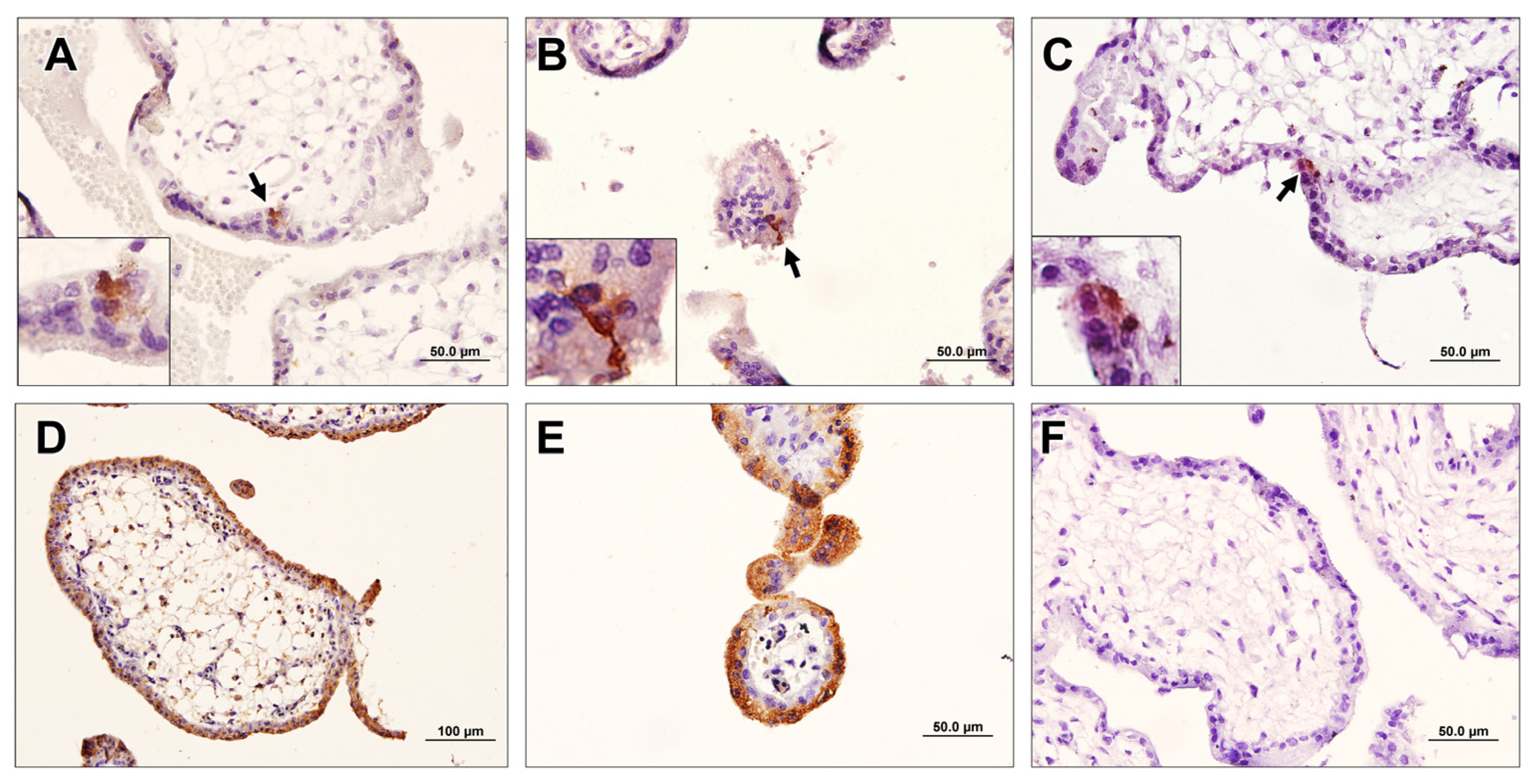
2.5. Immunohistochemistry (IHC)
2.6. Statistical Analysis
3. Results
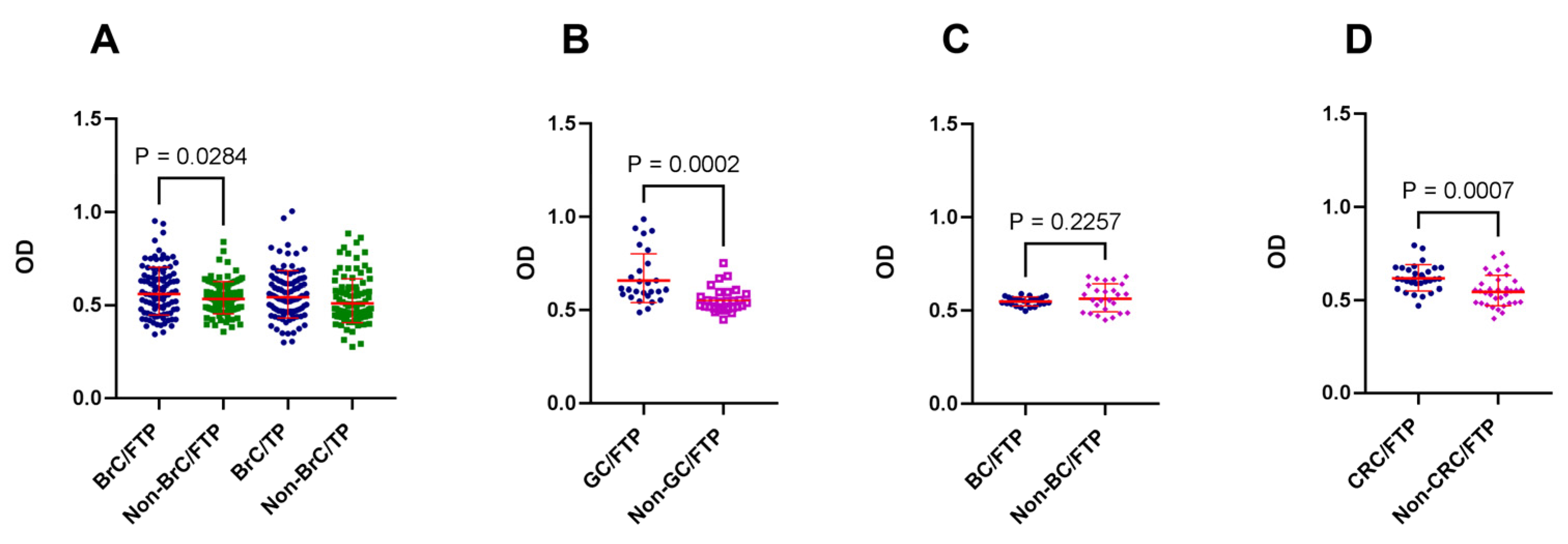
3.1. Cancer was Associated with the Generation of Placenta-Reactive Autoantibodies
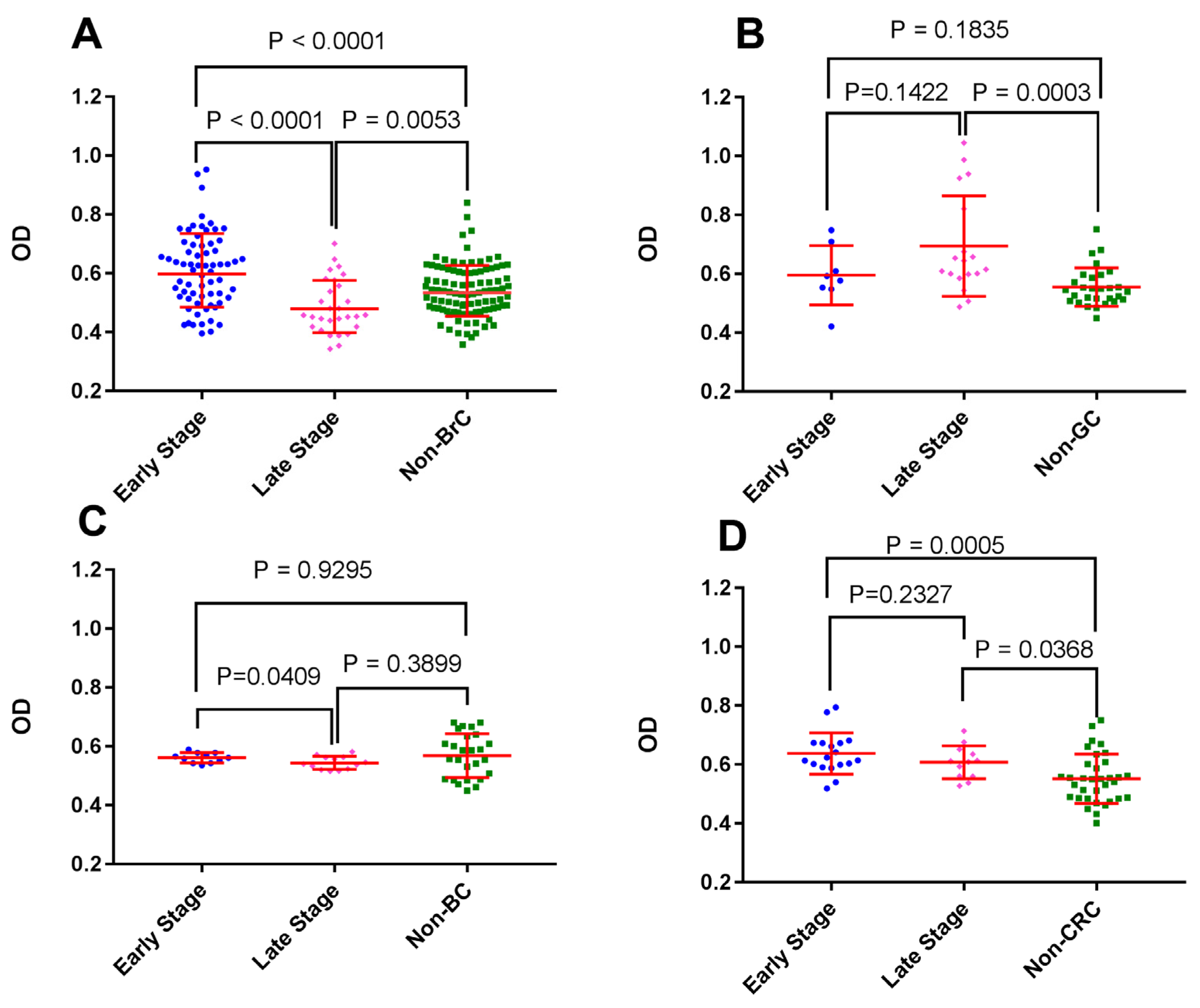
3.2. BrC at Early Stages Triggered Higher Autoantibody Reactivity against First-Trimester Placental Proteins
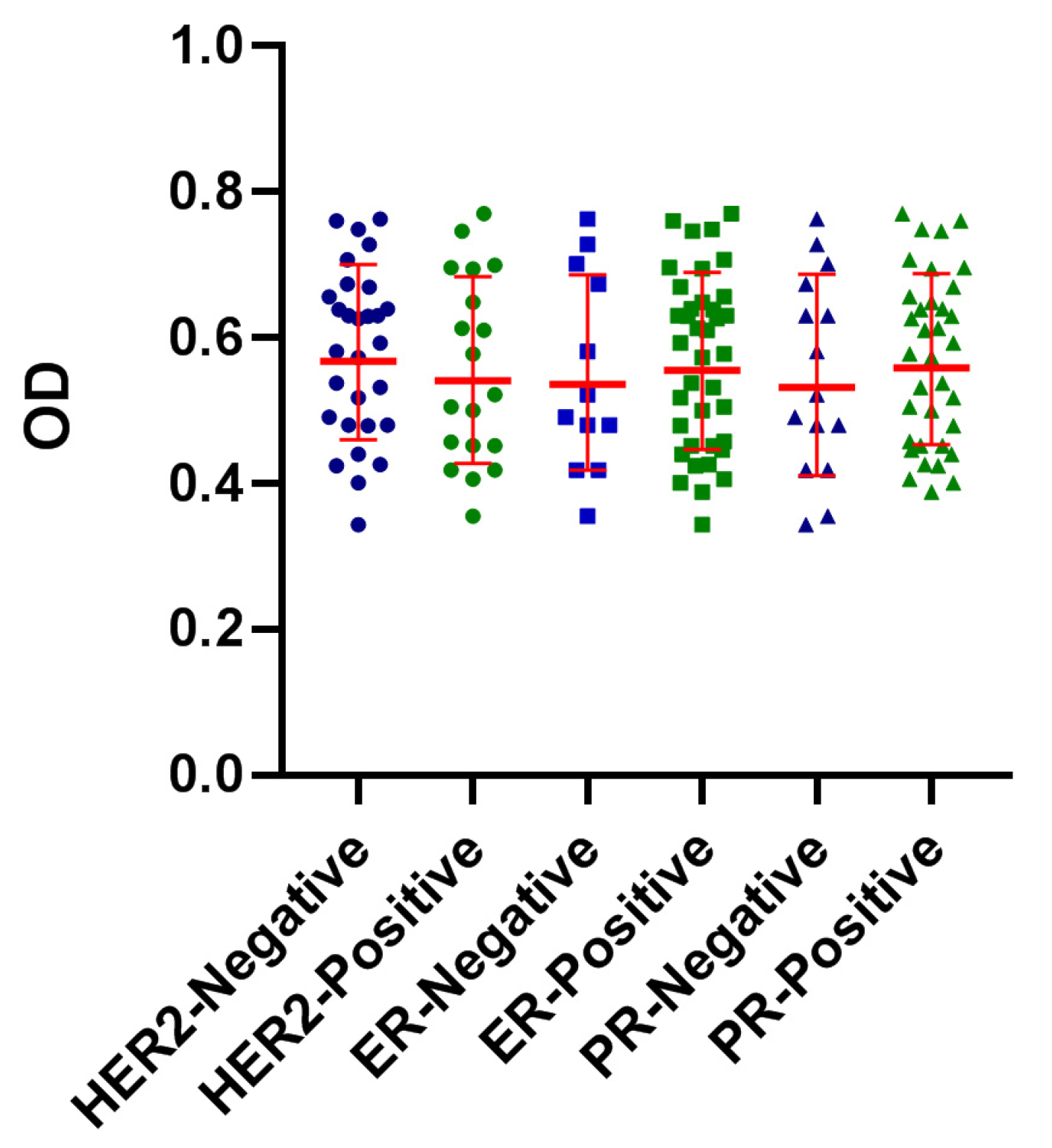
3.3. Reactivity of BrC Sera with FTP Did Not Show an Association with ER, PR, or HER2 Expression
3.4. Pregnancy in the Third Trimester was Associated with the Induction of TP-Reactive Autoantibodies
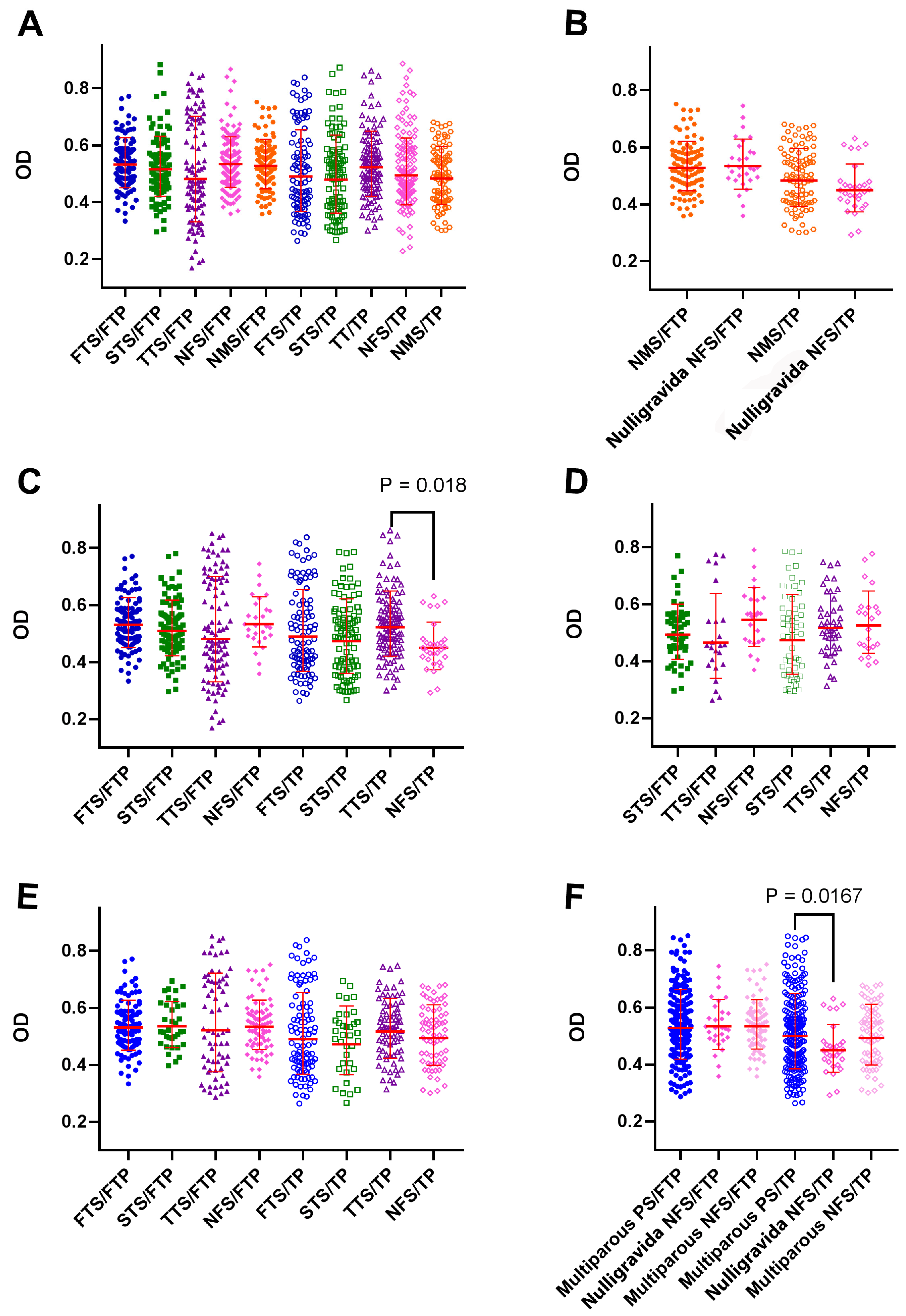
3.5. The Reactivity of BrC Sera with Placental Proteins Was Not Affected by Gravidity or Abortion
3.6. Breast-Cancer-Associated Autoantibodies Recognized Stem-Like Cells in the Placenta
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wagner, G.P.; Kshitiz; Dighe, A.; Levchenko, A. The Coevolution of Placentation and Cancer. Annu. Rev. Anim. Biosci. 2022, 10, 259–279. [Google Scholar] [CrossRef]
- Xu, C.; Jin, G.; Wu, H.; Cui, W.; Wang, U.H.; Manne, R.K.; Wang, G.; Zhang, W.; Zhang, X.; Han, F.; et al. SIRPγ-expressing cancer stem-like cells promote immune escape of lung cancer via Hippo signaling. J. Clin. Investig. 2022, 132, e141797. [Google Scholar] [CrossRef]
- Ruaro, B.; Salton, F. The History and Mystery of Alveolar Epithelial Type II Cells: Focus on Their Physiologic and Pathologic Role in Lung. Int. J. Mol. Sci. 2021, 22, 2566. [Google Scholar] [CrossRef]
- Choi, H.K.; Choi, B.C.; Lee, S.H.; Kim, J.W.; Cha, K.Y.; Baek, K.H. Expression of angiogenesis- and apoptosis-related genes in chorionic villi derived from recurrent pregnancy loss patients. Mol. Reprod. Dev. 2003, 66, 24–31. [Google Scholar] [CrossRef]
- Leach, L.; Babawale, M.O.; Anderson, M.; Lammiman, M. Vasculogenesis, angiogenesis and the molecular organisation of endothelial junctions in the early human placenta. J. Vasc. Res. 2002, 39, 246–259. [Google Scholar] [CrossRef]
- Brewer, B.G.; Mitchell, R.A.; Harandi, A.; Eaton, J.W. Embryonic vaccines against cancer: An early history. Exp. Mol. Pathol. 2009, 86, 192–197. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, Y.; Wu, B.; Liu, S.; Wu, E.; Fan, H.X.; Gui, M.M.; Chen, L.; Li, C.; Ju, Y.; et al. Placenta-derived gp96 as a multivalent prophylactic cancer vaccine. Sci. Rep. 2013, 3, 1947. [Google Scholar] [CrossRef] [Green Version]
- Harandi, A. Immunoplacental therapy, a potential multi-epitope cancer vaccine. Med. Hypotheses 2006, 66, 1182–1187. [Google Scholar] [CrossRef]
- Corocleanu, M. A possible “universal” cancer vaccine that might cause an immune response against emerging cancer cells that originate from any tissue. Med. Hypotheses 2008, 70, 381–383. [Google Scholar] [CrossRef]
- O’Connor, M.L.; Xiang, D.; Shigdar, S.; Macdonald, J.; Li, J.; Wang, T.; Pu, C.; Wang, Z.; Qiao, L.; Duan, W. Cancer stem cells: A contentious hypothesis now moving forward. Cancer Lett. 2014, 344, 180–187. [Google Scholar] [CrossRef]
- Fox, E.; Amaral, D.; Van de Water, J. Maternal and fetal antibrain antibodies in development and disease. Dev. Neurobiol. 2012, 72, 1327–1334. [Google Scholar] [CrossRef]
- Jasti, S.; Farahbakhsh, M.; Nguyen, S.; Petroff, B.K.; Petroff, M.G. Immune response to a model shared placenta/tumor-associated antigen reduces cancer risk in parous mice. Biol. Reprod. 2017, 96, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Bijian, K.; Mlynarek, A.M.; Balys, R.L.; Jie, S.; Xu, Y.; Hier, M.P.; Black, M.J.; Di Falco, M.R.; LaBoissiere, S.; Alaoui-Jamali, M.A. Serum proteomic approach for the identification of serum biomarkers contributed by oral squamous cell carcinoma and host tissue microenvironment. J. Proteome Res. 2009, 8, 2173–2185. [Google Scholar] [CrossRef]
- Mary George, N.; Parida, V.; Lahti, T.; Wincent, J. A systematic literature review of entrepreneurial opportunity recognition: Insights on influencing factors. Int. Entrep. Manag. J. 2016, 12, 309–350. [Google Scholar] [CrossRef]
- Tan, Q.; Wang, D.; Yang, J.; Xing, P.; Yang, S.; Li, Y.; Qin, Y.; He, X.; Liu, Y.; Zhou, S.; et al. Autoantibody profiling identifies predictive biomarkers of response to anti-PD1 therapy in cancer patients. Theranostics 2020, 10, 6399. [Google Scholar] [CrossRef]
- Belousov, P.V. The Autoantibodies against Tumor-Associated Antigens as Potential Blood-Based Biomarkers in Thyroid Neoplasia: Rationales, Opportunities and Challenges. Biomedicines 2022, 10, 468. [Google Scholar] [CrossRef]
- Poletaev, A. The main principles of adaptive immune system function: Self-recognition, self-interaction, and self-maintenance. Physiol. Autoimmun. Prev. Med. 2013, 1, 3–20. [Google Scholar]
- Zaenker, P.; Ziman, M.R. Serologic Autoantibodies as Diagnostic Cancer Biomarkers—A ReviewSerologic Autoantibodies as Diagnostic Cancer Biomarkers. Cancer Epidemiol. Biomark. Prev. 2013, 22, 2161–2181. [Google Scholar] [CrossRef] [Green Version]
- Casiano, C.A.; Mediavilla-Varela, M.; Tan, E.M. Tumor-associated antigen arrays for the serological diagnosis of cancer. Mol. Cell. Proteom. 2006, 5, 1745–1759. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.T.; Low, J.; Lim, S.G.; Chung MC, M. Serum autoantibodies as biomarkers for early cancer detection. FEBS J. 2009, 276, 6880–6904. [Google Scholar] [CrossRef]
- Zayakin, P.; Ancāns, G.; Siliņa, K.; Meistere, I.; Kalniņa, Z.; Andrejeva, D.; Endzeliņš, E.; Ivanova, L.; Pismennaja, P.; Ruskule, A.; et al. Tumor-associated autoantibody signature for the early detection of gastric cancer. Int. J. Cancer 2013, 132, 137–147. [Google Scholar] [CrossRef]
- Anagnostou, V.; Smith, K.; Forde, P.M.; Niknafs, N.; Bhattacharya, R.; White, J.; Zhang, T.; Adleff, V.; Phallen, J.; Wali, N.; et al. Evolution of Neoantigen Landscape during Immune Checkpoint Blockade in Non–Small Cell Lung CancerDynamics of Neoantigen Landscape during Immunotherapy. Cancer Discov. 2017, 7, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Lacombe, J.; Mangé, A.; Solassol, J. Use of autoantibodies to detect the onset of breast cancer. J. Immunol. Res. 2014, 2014, 574981. [Google Scholar] [CrossRef]
- Füzéry, A.K.; Levin, J.; Chan, M.M.; Chan, D.W. Translation of proteomic biomarkers into FDA approved cancer diagnostics: Issues and challenges. Clin. Proteom. 2013, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudian, J.; Ghods, R.; Nazari, M.; Jeddi- Tehrani, M.; Ghahremani, M.H.; Ghaffari-Tabrizi-Wizsy, N.; Ostad, S.N.; Zarnani, A.H. PLAC1: Biology and potential application in cancer immunotherapy. Cancer Immunol. Immunother 2019, 68, 1039–1058. [Google Scholar] [CrossRef]
- Li, P.; Shi, J.X.; Xing, M.T.; Dai, L.P.; Li, J.T.; Zhang, J.Y. Evaluation of serum autoantibodies against tumor-associated antigens as biomarkers in lung cancer. Tumour Biol. 2017, 39, 1010428317711662. [Google Scholar] [CrossRef] [Green Version]
- Hurley, L.C.; Levin, N.K.; Chatterjee, M.; Coles, J.; Muszkat, S.; Howarth, Z.; Dyson, G.; Tainsky, M.A. Evaluation of paraneoplastic antigens reveals TRIM21 autoantibodies as biomarker for early detection of ovarian cancer in combination with autoantibodies to NY-ESO-1 and TP53. Cancer Biomark. 2020, 27, 407–421. [Google Scholar] [CrossRef]
- Khorami Sarvestani, S.; Shojaeian, S.; Vanaki, N.; Gharesi-Fard, B.; Amini, M.; Gilani, K.; Soltanghoraei, H.; Arefi, S.; Jeddi-Tehrani, M.; Zarnani, A.H. Proteome profiling of human placenta reveals developmental stage-dependent alterations in protein signature. Clin. Proteom. 2021, 18, 18. [Google Scholar] [CrossRef]
- Marshall, G.M.; Carter, D.R.; Cheung, B.B.; Liu, T.; Mateos, M.K.; Meyerowitz, J.G.; Weiss, W.A. The prenatal origins of cancer. Nat. Rev. Cancer 2014, 14, 277–289. [Google Scholar] [CrossRef] [Green Version]
- Suhail, Y.; Maziarz, D.J.; Dighe, A.; Wagner, G. Cis-Regulatory Differences Explaining Evolved Levels of Endometrial Invasibility in Eutherian Mammals. bioRxiv 2020. [Google Scholar] [CrossRef]
- D’Souza, A.W.; Wagner, G.P. Malignant cancer and invasive placentation: A case for positive pleiotropy between endometrial and malignancy phenotypes. Evol. Med. Public Health 2014, 2014, 136–145. [Google Scholar] [CrossRef]
- Ander, S.E.; Diamond, M.S. Immune responses at the maternal-fetal interface. Sci. Immunol. 2019, 4, eaat6114. [Google Scholar] [CrossRef]
- Cho-Chung, Y.S. Autoantibody biomarkers in the detection of cancer. Biochim. Biophys. Acta 2006, 1762, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Nesterova, M.; Johnson, N.; Cheadle, C.; Cho-Chung, Y.S. Autoantibody biomarker opens a new gateway for cancer diagnosis. Biochim. Biophys. Acta 2006, 1762, 398–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohayem, J.; Conrad, K.; Zimmermann, T.; Frank, K.H. Comparison of the diagnostic accuracy of three commercially available enzyme immunoassays for anti-p53 antibodies. Clin. Chem. 1999, 45, 2014–2016. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Goodell, V.; Disis, M.L. Humoral immunity directed against tumor-associated antigens as potential biomarkers for the early diagnosis of cancer. J. Proteome Res. 2008, 7, 1388–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, J.W.; Wandall, H.H. Autoantibodies as biomarkers in cancer. Lab. Med. 2011, 42, 623–628. [Google Scholar] [CrossRef] [Green Version]
- Sexauer, D.; Gray, E.; Zaenker, P. Tumour-associated autoantibodies as prognostic cancer biomarkers-a review. Autoimmun. Rev. 2022, 21, 103041. [Google Scholar] [CrossRef] [PubMed]
- Zaenker, P.; Gray, E.S.; Ziman, M.R. Autoantibody production in cancer—The humoral immune response toward autologous antigens in cancer patients. Autoimmun. Rev. 2016, 15, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Shi, J.; Wang, P.; Song, C.; Wang, K.; Zhang, J.; Ye, H. Tumour-associated autoantibodies as diagnostic biomarkers for breast cancer: A systematic review and meta-analysis. Scand. J. Immunol. 2016, 83, 393–408. [Google Scholar] [CrossRef]
- Hamrita, B.; Ben Nasr, H.; Chahed, K.; Kabbage, M.; Chouchane, L. Proteomic Analysis of Human Breast Cancer: New. Nat. Commun. 2010, 3, 896. [Google Scholar]
- Chen, H.; Werner, S.; Tao, S.; Zörnig, I.; Brenner, H. Blood autoantibodies against tumor-associated antigens as biomarkers in early detection of colorectal cancer. Cancer Lett. 2014, 346, 178–187. [Google Scholar] [CrossRef]
- Liu, S.; Tan, Q.; Song, Y.; Shi, Y.; Han, X. Anti-p53 autoantibody in blood as a diagnostic biomarker for colorectal cancer: A meta-analysis. Scand. J. Immunol. 2020, 91, e12829. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Zhou, D.; Huang, J. Autoantibodies as biomarkers for colorectal cancer: A systematic review, meta-analysis, and bioinformatics analysis. Int. J. Biol. Markers 2019, 34, 334–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solassol, J.; Maudelonde, T.; Mange, A.; Pujol, J.L. Clinical relevance of autoantibody detection in lung cancer. J. Thorac. Oncol. 2011, 6, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Kazarian, M.; Laird-Offringa, I.A. Small-cell lung cancer-associated autoantibodies: Potential applications to cancer diagnosis, early detection, and therapy. Mol. Cancer 2011, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Li, X.; Ren, T.; Yin, Y. Autoantibodies as diagnostic biomarkers for lung cancer: A systematic review. Cell Death Discov. 2019, 5, 126. [Google Scholar] [CrossRef] [Green Version]
- Gadducci, A.; Cosio, S. Screening for Ovarian Cancer in the General Population: State of Art and Perspectives of Clinical Research. Anticancer. Res. 2022, 42, 4207–4216. [Google Scholar] [CrossRef] [PubMed]
- Bast, R.C.; Lu, Z.; Han, C.Y.; Lu, K.H.; Anderson, K.S.; Drescher, C.W.; Skates, S.J. Biomarkers and Strategies for Early Detection of Ovarian CancerEarly Detection of Ovarian Cancer. Cancer Epidemiol. Biomark. Prev. 2020, 29, 2504–2512. [Google Scholar] [CrossRef]
- Reddy, R.; Mintz, J.; Golan, R.; Firdaus, F.; Ponce, R.; Booven, D.V.; Manoharan, A.; Issa, I.; Blomberg, B.B.; Arora, H. Antibody Diversity in Cancer: Translational Implications and Beyond. Vaccines 2022, 10, 1165. [Google Scholar] [CrossRef]
- Saif, M.W.; Zalonis, A.; Syrigos, K. The clinical significance of autoantibodies in gastrointestinal malignancies: An overview. Expert. Opin. Biol. Ther. 2007, 7, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Chen, H.; Tao, S.; Brenner, H. Systematic review: Serum autoantibodies in the early detection of gastric cancer. Int. J. Cancer 2015, 136, 2243–2252. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Katayama, H.; Fahrmann, J.F.; Hanash, S.M. Development of autoantibody signatures for common cancers. Semin. Immunol. 2020, 47, 101388. [Google Scholar] [CrossRef] [PubMed]
- Jelski, W.; Mroczko, B. Molecular and Circulating Biomarkers of Gastric Cancer. Int. J. Mol. Sci. 2022, 23, 7588. [Google Scholar] [CrossRef]
- Costanzo, V.; Bardelli, A.; Siena, S.; Abrignani, S. Exploring the links between cancer and placenta development. Open Biol. 2018, 8, 180081. [Google Scholar] [CrossRef] [Green Version]
- Lala, P.K.; Nandi, P.; Hadi, A.; Halari, C. A crossroad between placental and tumor biology: What have we learnt? Placenta 2021, 116, 12–30. [Google Scholar] [CrossRef]
- Greaves, M.; Hughes, M. Cancer cell transmission via the placenta. Evol. Med. Public Health 2018, 2018, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Joseph, M.; Enting, D. Immune responses in bladder cancer-role of immune cell populations, prognostic factors and therapeutic implications. Front. Oncol. 2019, 9, 1270. [Google Scholar] [CrossRef] [Green Version]
- Okae, H.; Toh, H.; Sato, T.; Hiura, H.; Takahashi, S.; Shirane k Kabayama, Y.; Suyama, M.; Sasaki, H.; Arima, T. Derivation of human trophoblast stem cells. Cell Stem Cell 2018, 22, 50–63.e6. [Google Scholar] [CrossRef] [Green Version]
- Castellucci, M.; Kosanke, G.; Verdenelli, F.; Huppertz, B.; Kaufmann, P. Villous sprouting: Fundamental mechanisms of human placental development. Hum. Reprod. Update 2000, 6, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Ben-Hur, H.; Cohen, O.; Schneider, D.; Gurevich, P.; Halperin, R.; Bala, U.; Mozes, M.; Zusman, I. The role of lymphocytes and macrophages in human breast tumorigenesis: An immunohistochemical and morphometric study. Anticancer. Res. 2002, 22, 1231–1238. [Google Scholar] [PubMed]
- Froehlich, K.; Schmidt, A.; Heger, J.I.; Al-Kawlani, B.; Aberl, C.A.; Jeschke, U.; Loibl, S.; Markert, U.R. Breast cancer, placenta and pregnancy. Eur. J. Cancer 2019, 115, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.K.; Rani, R.; Agrawal, S. An update in recurrent spontaneous abortion. Arch. Gynecol. Obstet. 2005, 272, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Chang-Claude, J.; Becher, H.; Eby, N.; Bastert, G.; Wahrendorf, J.; Hamann, U. Modifying effect of reproductive risk factors on the age at onset of breast cancer for GermanBRCA1 mutation carriers. J. Cancer Res. Clin. Oncol. 1997, 123, 272–279. [Google Scholar] [CrossRef]
- Rebbeck, T.R.; Wang, Y.; Kantoff, P.W.; Krithivas, K.; Neuhausen, S.L.; Godwin, A.K.; Daly, M.B.; Narod, S.A.; Brunet, J.S.; Vesprini, D.; et al. Modification of BRCA1- and BRCA2-associated breast cancer risk by AIB1 genotype and reproductive history. Cancer Res. 2001, 61, 5420–5424. [Google Scholar]
- Tryggvadottir, L.; Olafsdottir, E.J.; Gudlaugsdottir, S.; Thorlacius, S.; Jonasson, J.G.; Tulinius, H.; Eyfjord, J.E. BRCA2 mutation carriers, reproductive factors and breast cancer risk. Breast Cancer Res. 2003, 5, R121–R128. [Google Scholar] [CrossRef] [Green Version]
- MacMahon, B.; Cole, P.; MLin, T.; Lowe, C.R.; Mirra, A.P.; Ravnihar, B.; Salber, E.J.; Valaoras, V.J.; Yuasa, S. Age at first birth and breast cancer risk. Bull. World Health Organ. 1970, 43, 209. [Google Scholar]
- Albrektsen, G.; Heuch, I.; Hansen, I.; Kvåle, G. Breast cancer risk by age at birth, time since birth and time intervals between births: Exploring interaction effects. Br. J. Cancer 2005, 92, 167–175. [Google Scholar] [CrossRef]
- Slepicka, P.F.; Cyrill, S.L.; Dos Santos, C.O. Pregnancy and Breast Cancer: Pathways to Understand Risk and Prevention. Trends Mol. Med. 2019, 25, 866–881. [Google Scholar] [CrossRef] [Green Version]
- Husby, A.; Wohlfahrt, J.; Øyen, N.; Melbye, M. Pregnancy duration and breast cancer risk. Nat. Commun. 2018, 9, 4255. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, S.; Kamat, M.M.; Walusimbi, S.S.; Pate, J.L.; Ott, T.L. Effects of early pregnancy on uterine lymphocytes and endometrial expression of immune-regulatory molecules in dairy heifers. Biol. Reprod. 2017, 97, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Moffett-King, A. Natural killer cells and pregnancy. Nat. Rev. Immunol. 2002, 2, 656–663. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Sample Size | Mean Age | Gender | Stage | ||
|---|---|---|---|---|---|---|
| Female | Male | Early | Late | |||
| CRC | 34 | 51 ± 11.86 | 14 | 20 | 19 | 15 |
| BrC | 100 | 44 ± 8.41 | 100 | 0 | 70 | 30 |
| BC | 29 | 57 ± 14.08 | 10 | 19 | 13 | 16 |
| GC | 30 | 57 ± 10.44 | 14 | 16 | 10 | 20 |
| Non-CRC | 34 | 51 ± 11.84 | 14 | 20 | - | - |
| Non-BrC | 100 | 43 ± 8.97 | 100 | 0 | - | - |
| Non-BC | 29 | 56 ± 31.03 | 10 | 19 | - | - |
| Non-GC | 30 | 57 ± 10.28 | 14 | 16 | - | - |
| Cancer Type | Sample Size | Mean Age (Year) | Gestational Age (Weeks) |
|---|---|---|---|
| First-trimester pregnant women | 100 | 33 ± 3.77 | 11–13 |
| Second trimester pregnant women | 100 | 32 ± 4.07 | 15–18 |
| Third trimester pregnant women | 110 | 29 ± 6.67 | 37–41 |
| Normal males | 100 | 38 ± 9.57 | - |
| Groups | Inclusion Criteria | Exclusion Criteria |
|---|---|---|
| Case | Pathologically confirmed cancer | Current or past history of autoimmune diseases |
| Available clinical pathological data | Previous history of cancer | |
| No history of any treatment (surgery, radiotherapy and/or chemotherapy, hormone therapy) before sampling | Received immunosuppressive drugs during the three months before sample collection | |
| Newly diagnosed cases | History of chemo, adjuvant, or radiotherapy and cancer surgery | |
| Control | Age- and sex-matched subjects | Not age- and sex-matched |
| Subjects with no history of cancer | History of chronic diseases or cancer | |
| Non-BrC females with confirmed non-malignant report on breast Fine Needle Aspiration (FNA) | Last pregnancy ≤ 3 years before cancer onset (female patients) | |
| Non-GC patients as confirmed by endoscopic gastric sampling and pathology confirmation | More than two abortions | |
| Non-CRC and non-BC as confirmed by colonoscopy and sonography | Current or past history of autoimmune diseases | |
| Normal males referred to medical laboratories for a routine checkup with normal biochemical and hematological test results | History of immunosuppressive drug intake three months before sampling | |
| Pregnant females in first or second trimesters of pregnancy with normal screening tests | Having no clinical pathological data | |
| Pregnant female in third trimester of pregnancy referred for cesarean section | More than 10 pregnancies |
| Parameter | B | 95% Wald Confidence Interval | Sig. | |
|---|---|---|---|---|
| Lower | Upper | |||
| BrC | 0.051 | 0.016 | 0.086 | 0.005 |
| Non-BrC | Referent | |||
| Gravidity ≥ 1 | 0.060 | 0.104 | 0.016 | 0.007 |
| Gravidity = 0 | Referent | |||
| Abortion ≥ 1 | 0.001 | −0.040 | 0.043 | 0.957 |
| Abortion = 0 | Referent | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khorami Sarvestani, S.; Shojaeian, S.; Sarrami-Forooshani, R.; Yekaninejad, M.S.; Gilany, K.; Ghaderi, A.; Hashemnejad, M.; Olfatbakhsh, A.; Notash Haghighat, F.; Montazeri, S.; et al. Cancer Is Associated with the Emergence of Placenta-Reactive Autoantibodies. Biomedicines 2023, 11, 316. https://doi.org/10.3390/biomedicines11020316
Khorami Sarvestani S, Shojaeian S, Sarrami-Forooshani R, Yekaninejad MS, Gilany K, Ghaderi A, Hashemnejad M, Olfatbakhsh A, Notash Haghighat F, Montazeri S, et al. Cancer Is Associated with the Emergence of Placenta-Reactive Autoantibodies. Biomedicines. 2023; 11(2):316. https://doi.org/10.3390/biomedicines11020316
Chicago/Turabian StyleKhorami Sarvestani, Sara, Sorour Shojaeian, Ramin Sarrami-Forooshani, Mir Saeed Yekaninejad, Kambiz Gilany, Abbas Ghaderi, Maryam Hashemnejad, Asiie Olfatbakhsh, Farzane Notash Haghighat, Samaneh Montazeri, and et al. 2023. "Cancer Is Associated with the Emergence of Placenta-Reactive Autoantibodies" Biomedicines 11, no. 2: 316. https://doi.org/10.3390/biomedicines11020316