
Effects of Monoamino-Oxidase-A (MAO-A) Inhibition on Skeletal Muscle Inflammation and Wasting through Pancreatic Ductal Adenocarcinoma in Triple Transgenic Mice

 ,
,
Abstract
:1. Introduction
2. Material and Methods
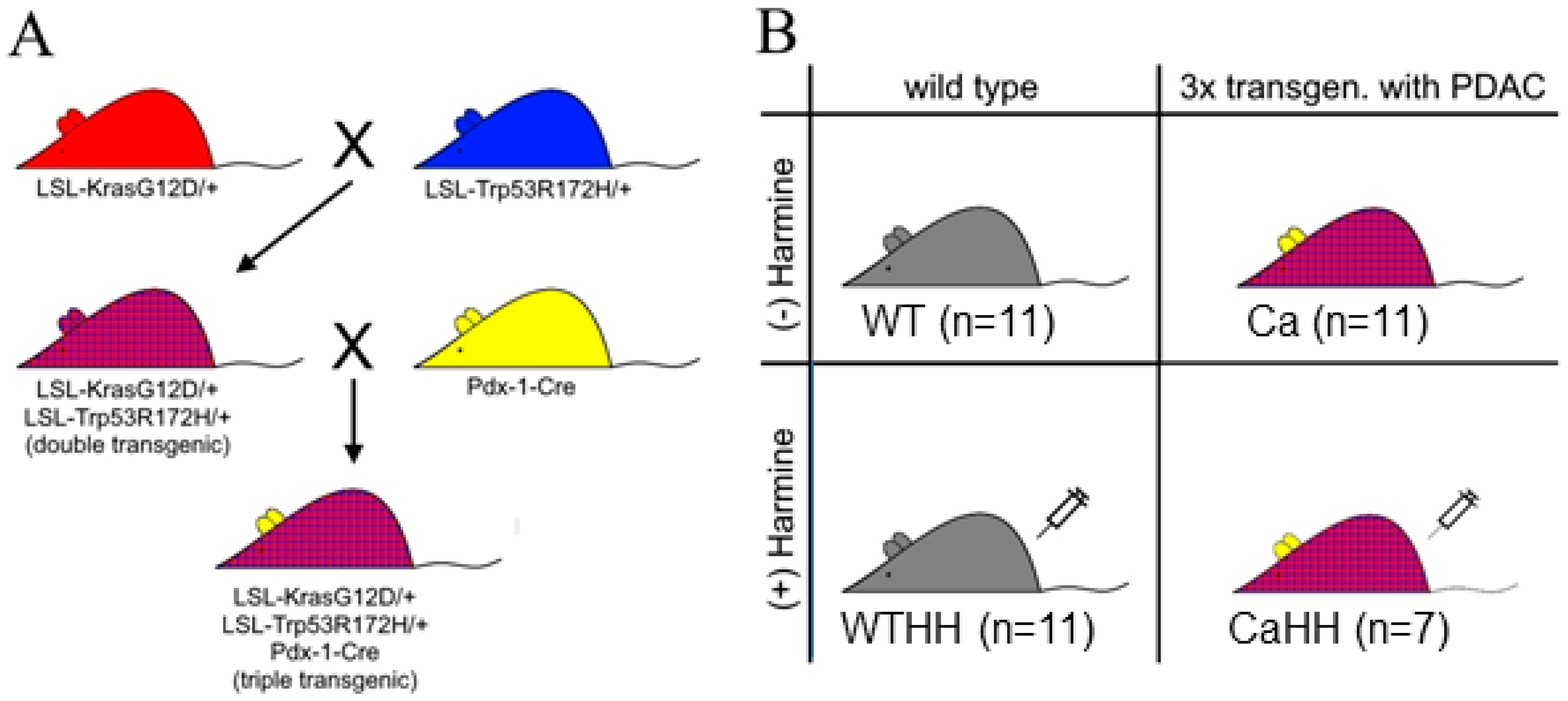
2.1. Tumor Mouse Model
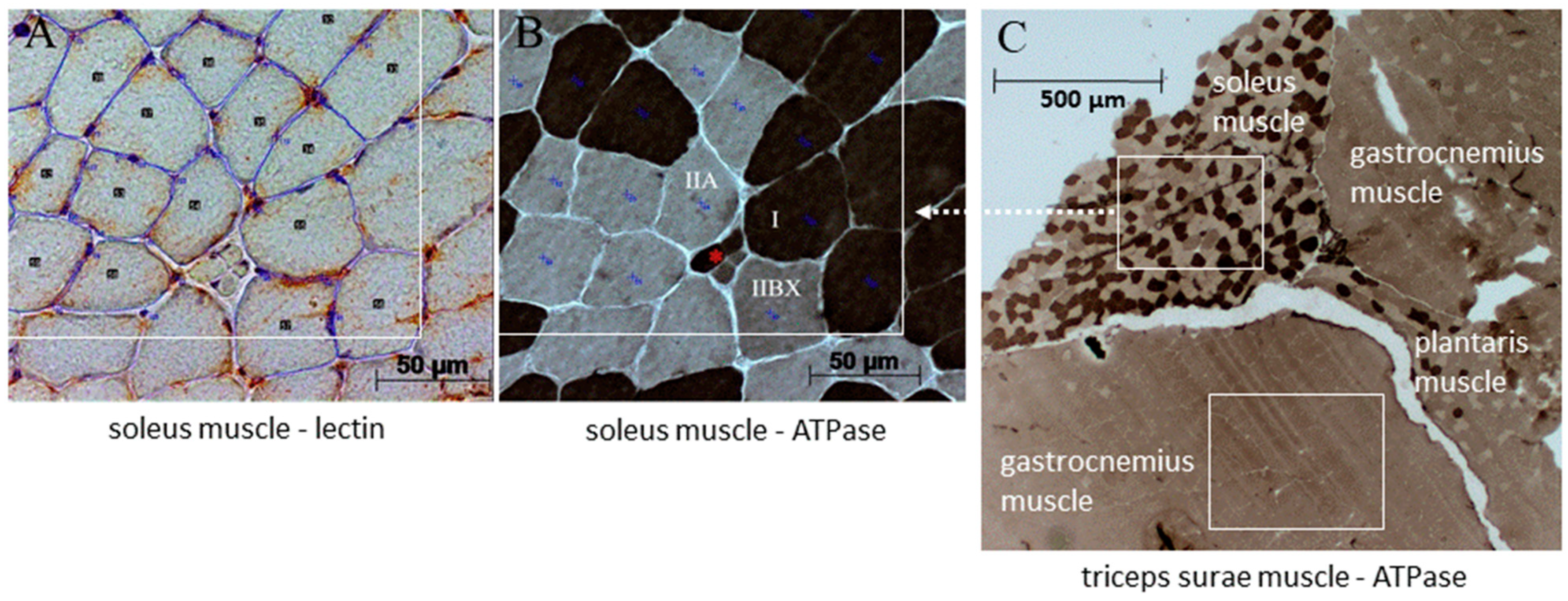
2.2. Histological Examination
2.3. Mitochondrial Ultrastructure
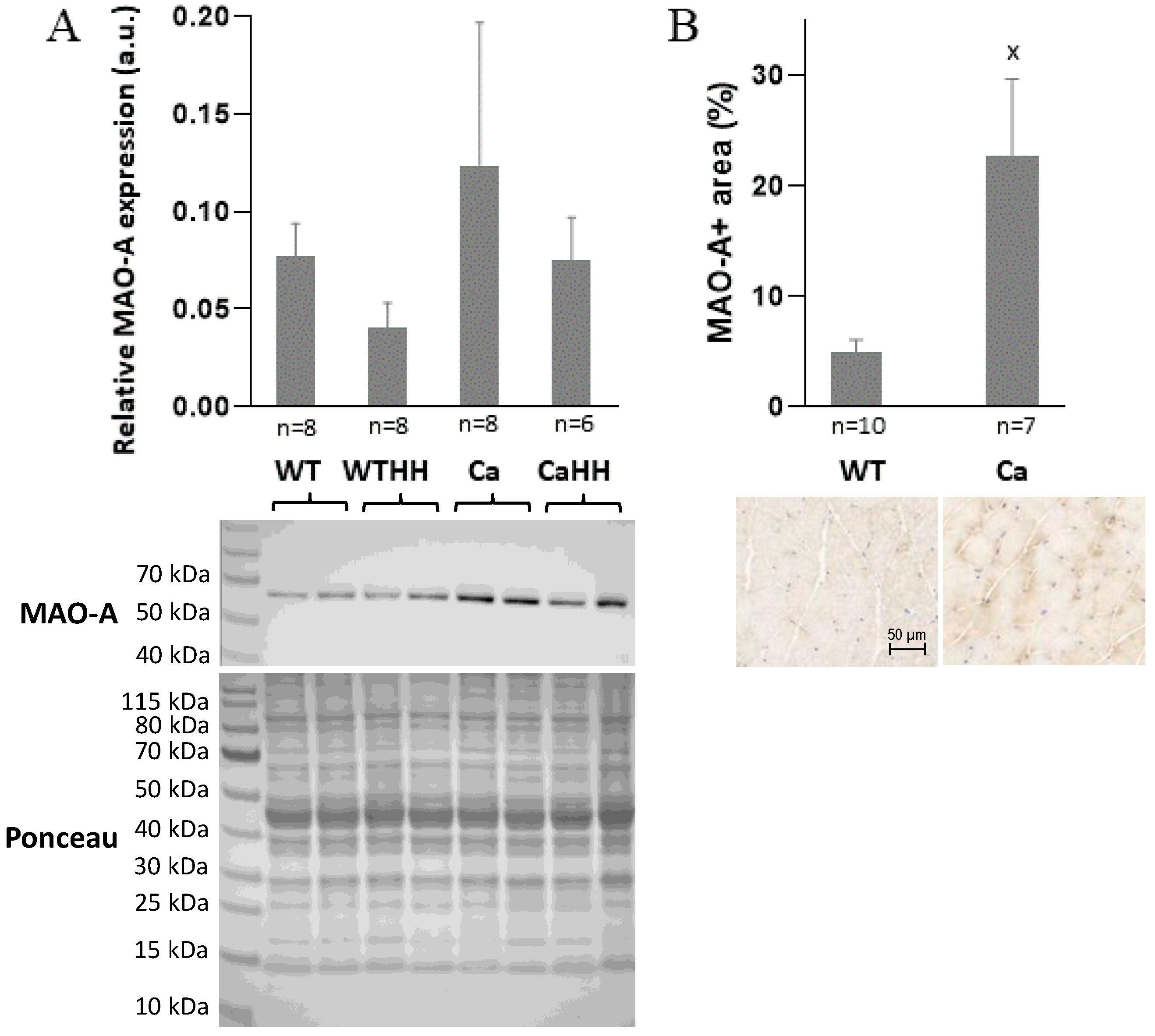
2.4. Western Blotting of MAO-A Expression
2.5. Intramyocellular Glutathione (GSH) Content and Glutathione Redox State
2.6. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.7. Statistical Analyses
3. Results
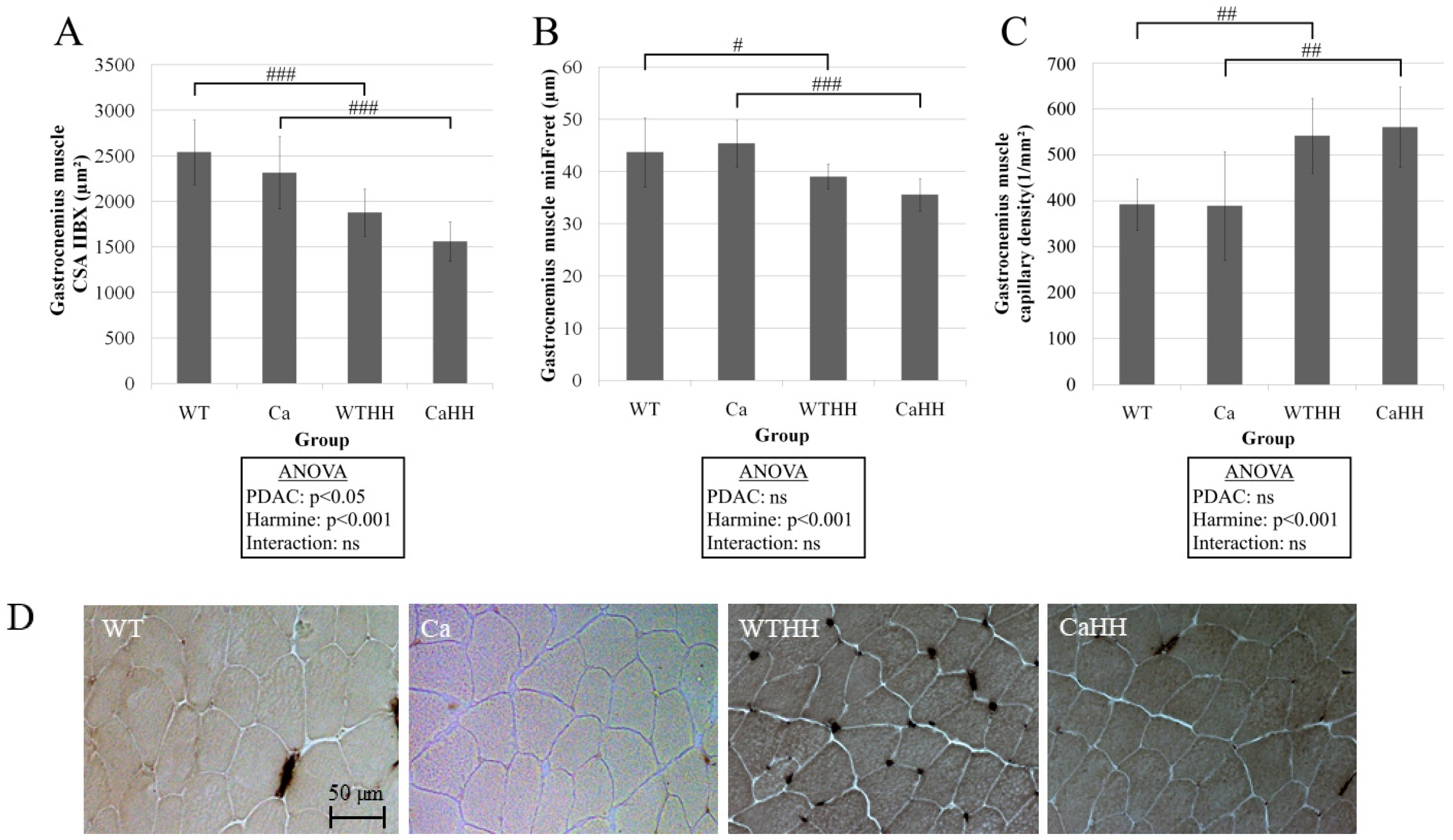
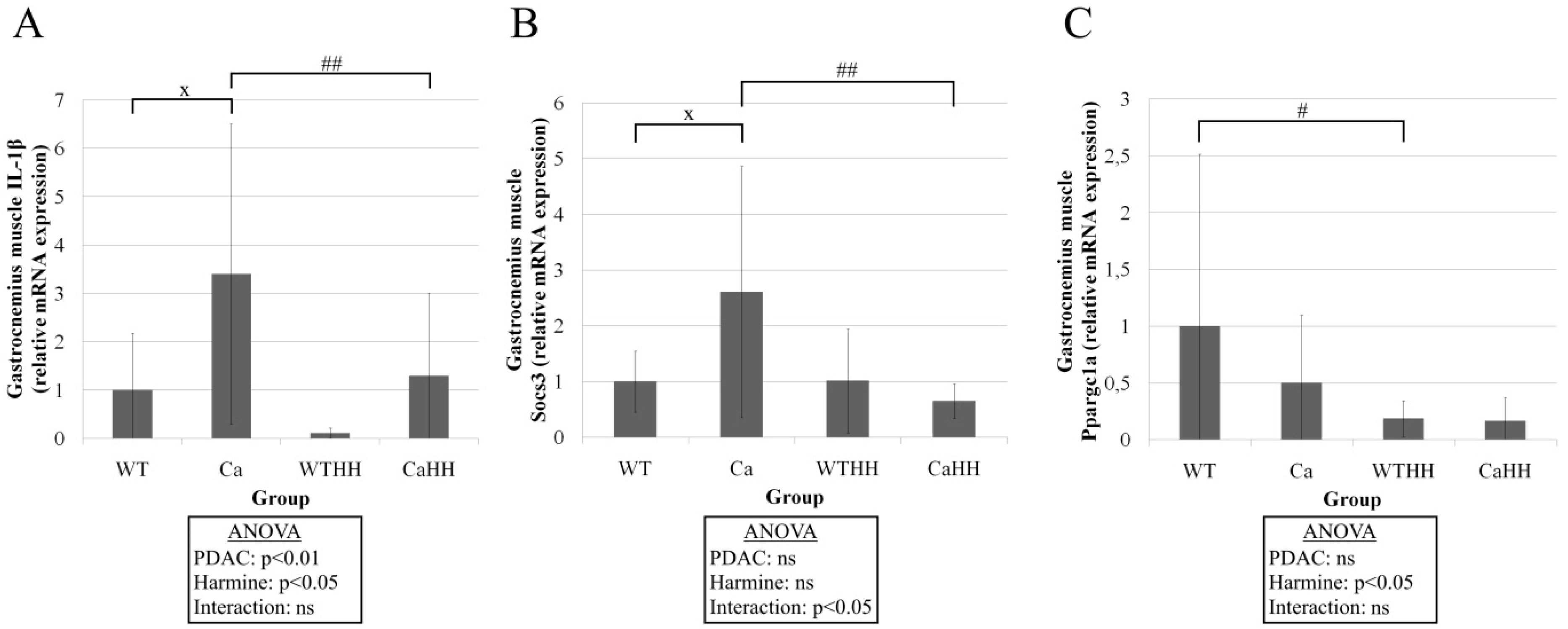
3.1. Gastrocnemius Muscle
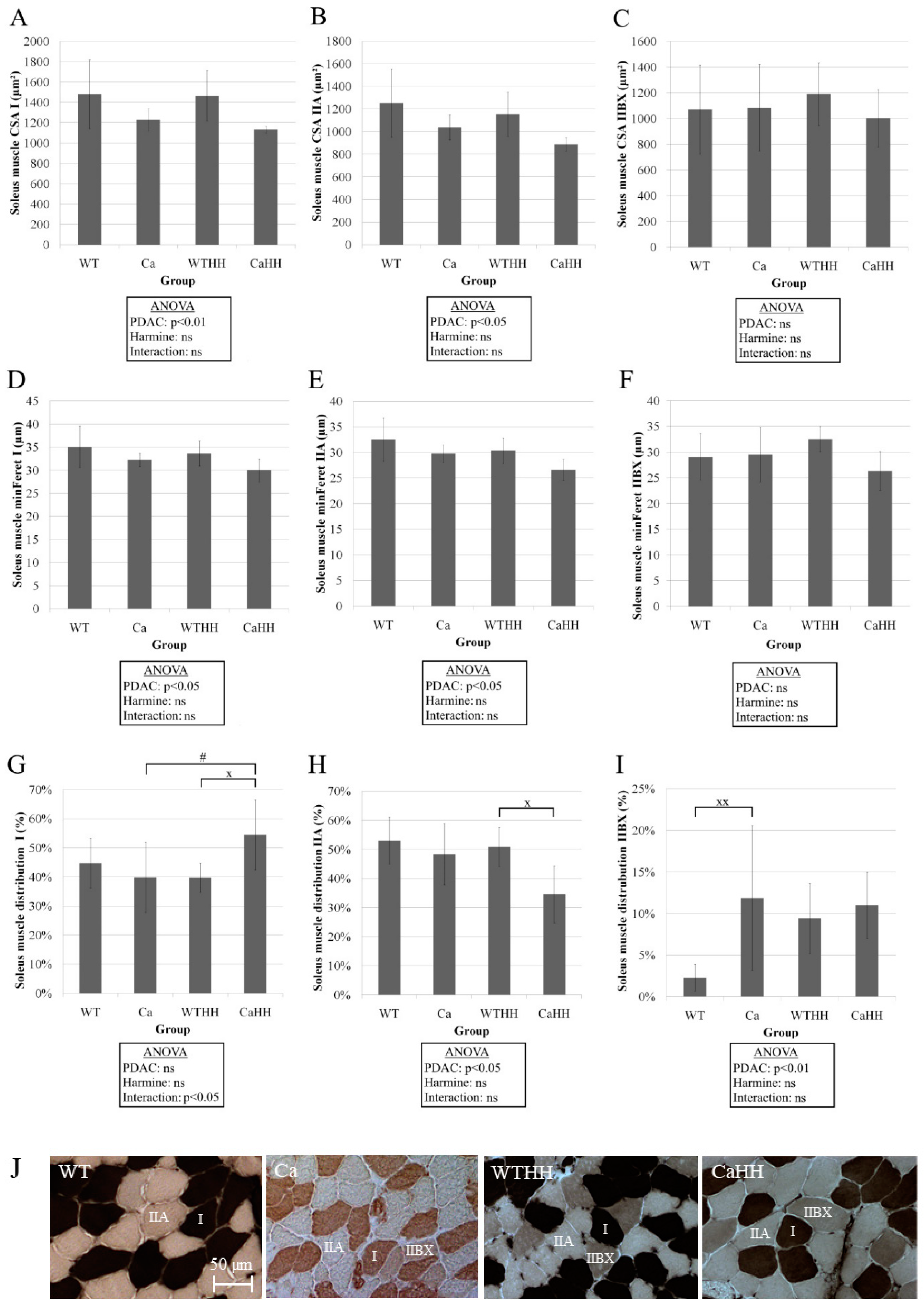
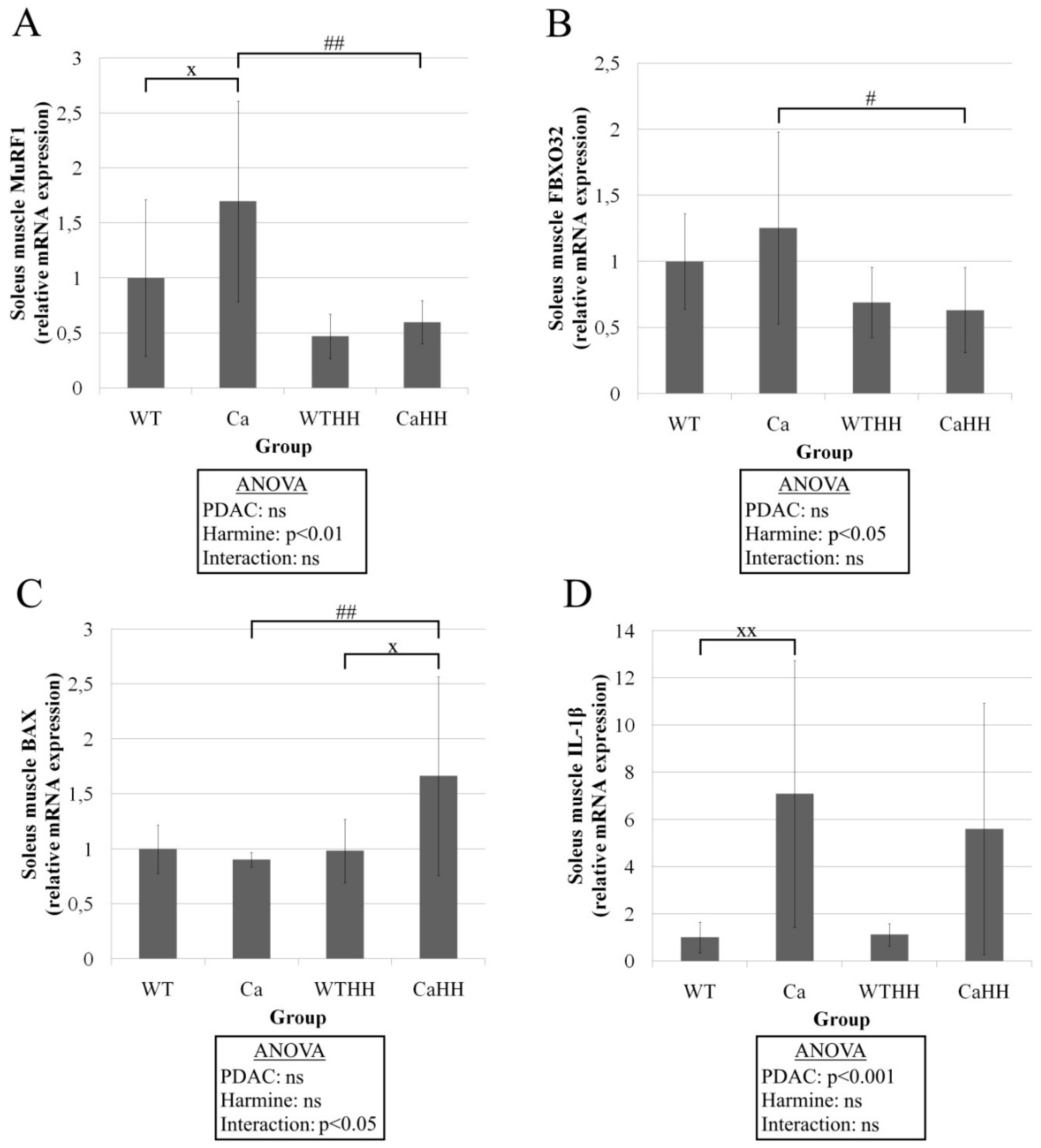
3.2. Soleus Muscle—Histomorphometry and Gene Expression
3.3. Gastrocnemius and Soleus Muscles—Mitochondrial Integrity and Centronucleation
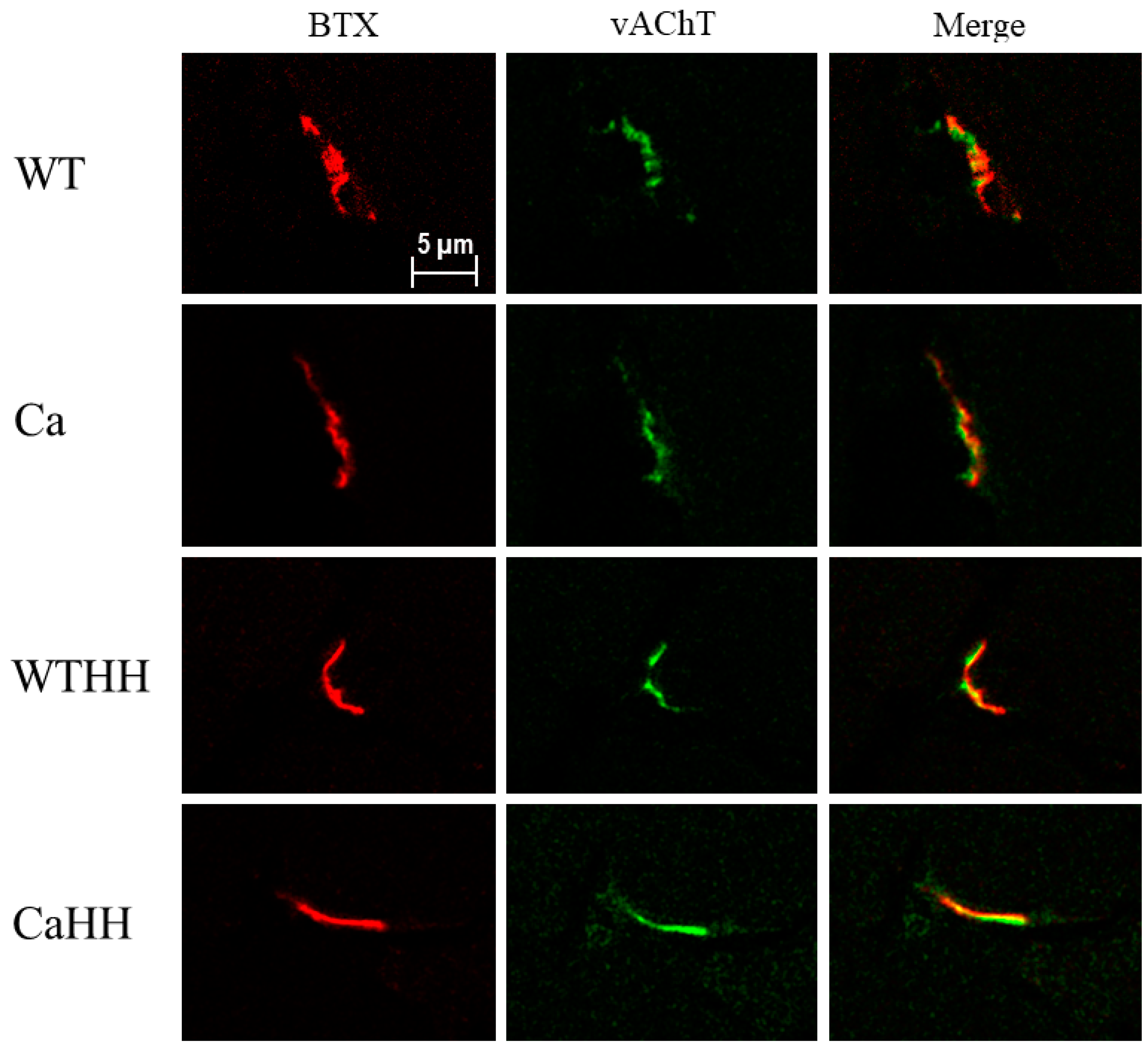
3.4. Quadriceps Muscle: Morphological NMJ Integrity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| BAX | Bcl2-associated X protein |
| Bcl2 | B-cell lymphoma 2 |
| BTX | α-bungarotoxin |
| Ca | untreated 3xtransgenic mice with PDAC |
| CaHH | HH-treated 3xtransgenic mice with PDAC |
| COX2 | cyclooxygenase 2 (syn: Prostaglandin endoperoxide synthase 2 (PTGS2)) |
| CSA | cross-sectional area (of muscle fibers in transversal cryosections) |
| DYRK1 A/B | dual-specificity tyrosine phosphorylation regulated kinases 1 A/B |
| FBXO32 | F-box only protein 32 (syn: MAFbx, Atrogin1) |
| GSSG | glutathione disulfide |
| rGSH | reduced glutathione |
| tGSH | total glutathione |
| HH | Harmine-Hydrochloride |
| IL | interleukin |
| KPC | denotes the transgenic mouse model of PDAC, that incorporates mutant |
| endogenous alleles of the Kras and P53 genes using Cre-Lox technology | |
| according to Hingorani et al. [44] | |
| MAO-A/B | monoamino oxidase A/B |
| MMP9 | matrix metallopeptidase 9 |
| MuRF1 | muscle ring finger protein 1 (syn: TRIM63) |
| NMJ | neuromuscular junction (motor endplate) |
| PDAC | pancreatic ductal adenocarcinoma |
| p62 | ubiquitin-binding protein p62 (syn: Sequestosome 1 (Sqstml1)) |
| PAX7 | paired box protein 7 |
| Ppargc1a | peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| (syn: PGC-1α) | |
| RER1 | retention of endoplasmatic reticulum 1 protein |
| ROS | reactive oxygen species |
| RT-qPCR | real-time quantitative polymerase chain reaction |
| Socs3 | suppressor of cytokine signaling 3 |
| TEM | transmission electron microscopy |
| TNFα | tumor necrosis factor α |
| WT | untreated wild type mice |
| WTHH | HH-treated wild type mice |
| vAChT | vesicular acetylcholine transporter (syn: SLC18A3) |
| VEGFA | vascular endothelial growth factor A |
References
- Porporato, P.E. Understanding cachexia as a cancer metabolism syndrome. Oncogenesis 2016, 5, e200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meza-Valderrama, D.; Marco, E.; Dávalos-Yerovi, V.; Muns, M.D.; Tejero-Sánchez, M.; Duarte, E.; Sánchez-Rodríguez, D. Sarcopenia, Malnutrition, and Cachexia: Adapting Definitions and Terminology of Nutritional Disorders in Older People with Cancer. Nutrients 2021, 13, 761. [Google Scholar] [CrossRef]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Johns, N.; Stephens, N.A.; Fearon, K.C. Muscle wasting in cancer. Int. J. Biochem. Cell Biol. 2013, 45, 2215–2229. [Google Scholar] [CrossRef]
- Al-Batran, S.E.; Hofheinz, R.D.; Reichart, A.; Pauligk, C.; Schönherr, C.; Schlag, R.; Siegler, G.; Dörfel, S.; Koenigsmann, M.; Zahn, M.O.; et al. Quality of life and outcome of patients with metastatic pancreatic cancer receiving first-line chemotherapy with nab-paclitaxel and gemcitabine: Real-life results from the prospective QOLIXANE trial of the Platform for Outcome, Quality of Life and Translational Research on Pancreatic Cancer registry. Int. J. Cancer 2021, 148, 1478–1488. [Google Scholar]
- Arthur, S.T.; Van Doren, B.A.; Roy, D.; Noone, J.M.; Zacherle, E.; Blanchette, C.M. Cachexia among US cancer patients. J. Med. Econ. 2016, 19, 874–880. [Google Scholar] [CrossRef]
- Arthur, S.T.; Noone, J.M.; Van Doren, B.A.; Roy, D.; Blanchette, C.M. One-year prevalence, comorbidities and cost of cachexia-related inpatient admissions in the USA. Drugs Context 2014, 3, 212265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, C.; Spiro, A.; Ahern, R.; Emery, P.W. Oral nutritional interventions in malnourished patients with cancer: A systematic review and meta-analysis. J. Natl. Cancer Inst. 2012, 104, 371–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande, A.J.; Silva, V.; Riera, R.; Medeiros, A.; Vitoriano, S.G.P.; Peccin, M.S.; Maddocks, M. Exercise for cancer cachexia in adults. Cochrane Database Syst. Rev. 2014, 11, CD010804. [Google Scholar] [CrossRef] [Green Version]
- Balstad, T.R.; Solheim, T.S.; Strasser, F.; Kaasa, S.; Bye, A. Dietary treatment of weight loss in patients with advanced cancer and cachexia: A systematic literature review. Crit. Rev. Oncol. Hematol. 2014, 91, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Solheim, T.S.; Fearon, K.C.; Blum, D.; Kaasa, S. Non-steroidal anti-inflammatory treatment in cancer cachexia: A systematic literature review. Acta Oncol. 2013, 52, 6–17. [Google Scholar] [CrossRef] [Green Version]
- Ruiz Garcia, V.; López-Briz, E.; Carbonell Sanchis, R.; Gonzalvez Perales, J.L.; Bort-Marti, S. Megestrol acetate for treatment of anorexia-cachexia syndrome. Cochrane Database Syst. Rev. 2013, CD004310. [Google Scholar] [CrossRef] [PubMed]
- Strasser, F.; Luftner, D.; Possinger, K.; Ernst, G.; Ruhstaller, T.; Meissner, W.; Ko, Y.D.; Schnelle, M.; Reif, M.; Cerny, T.; et al. Comparison of orally administered cannabis extract and delta-9-tetrahydrocannabinol in treating patients with cancer-related anorexia-cachexia syndrome: A multicenter, phase III, randomized, double-blind, placebo-controlled clinical trial from the Cannabis-In-Cachexia-Study-Group. J. Clin. Oncol. 2006, 24, 3394–3400. [Google Scholar] [PubMed]
- Argilés, J.M.; Busquets, S.; Stemmler, B.; López-Soriano, F.J. Cancer cachexia: Understanding the molecular basis. Nat. Rev. Cancer 2014, 14, 754–762. [Google Scholar] [CrossRef]
- Martin, L.; Birdsell, L.; Macdonald, N.; Reiman, T.; Clandinin, M.T.; McCargar, L.J.; Murphy, R.; Ghosh, S.; Sawyer, M.B.; Baracos, V.E. Cancer cachexia in the age of obesity: Skeletal muscle depletion is a powerful prognostic factor, independent of body mass index. J. Clin. Oncol. 2013, 31, 1539–1547. [Google Scholar] [CrossRef]
- Prado, C.M.; Lieffers, J.R.; McCargar, L.J.; Reiman, T.; Sawyer, M.B.; Martin, L.; Baracos, V.E. Prevalence and clinical implications of sarcopenic obesity in patients with solid tumours of the respiratory and gastrointestinal tracts: A population-based study. Lancet Oncol. 2008, 9, 629–635. [Google Scholar] [CrossRef]
- Tan, B.H.; Birdsell, L.A.; Martin, L.; Baracos, V.E.; Fearon, K.C. Sarcopenia in an overweight or obese patient is an adverse prognostic factor in pancreatic cancer. Clin. Cancer Res. 2009, 15, 6973–6979. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.; Freyssenet, D. Phenotypic features of cancer cachexia-related loss of skeletal muscle mass and function: Lessons from human and animal studies. J. Cachexia Sarcopenia Muscle 2021, 12, 252–273. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.M.; Frye, G.S.; Ahn, B.; Ferreira, L.F.; Judge, A.R. Cancer cachexia decreases specific force and accelerates fatigue in limb muscle. Biochem. Biophys. Res. Commun. 2013, 435, 488–492. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Zhang, S.; Hu, W.; Lu, X.; Lou, N.; Yang, Z.; Chen, S.; Zhang, X.; Yang, H. Valproic acid attenuates skeletal muscle wasting by inhibiting C/EBPβ-regulated atrogin1 expression in cancer cachexia. Am. J. Physiol. Cell Physiol. 2016, 311, C101–C115. [Google Scholar] [CrossRef] [Green Version]
- Bonetto, A.; Rupert, J.E.; Barreto, R.; Zimmers, T.A. The Colon-26 Carcinoma Tumor-bearing Mouse as a Model for the Study of Cancer Cachexia. J. Vis. Exp. 2016, 30, e54893. [Google Scholar]
- Henderson, S.E.; Ding, L.Y.; Mo, X.; Bekaii-Saab, T.; Kulp, S.K.; Chen, C.S.; Huang, P.H. Suppression of Tumor Growth and Muscle Wasting in a Transgenic Mouse Model of Pancreatic Cancer by the Novel Histone Deacetylase Inhibitor AR-42. Neoplasia 2016, 18, 765–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delitto, D.; Judge, S.M.; Delitto, A.E.; Nosacka, R.L.; Rocha, F.G.; DiVita, B.B.; Gerber, M.H.; George, T.J.; Behrns, K.E.; Hughes, S.J.; et al. Human pancreatic cancer xenografts recapitulate key aspects of cancer cachexia. Oncotarget 2017, 8, 1177–1189. [Google Scholar] [CrossRef] [Green Version]
- Ciciliot, S.; Rossi, A.C.; Dyar, K.A.; Blaauw, B.; Schiaffino, S. Muscle type and fiber type specificity in muscle wasting. Int. J. Biochem. Cell Biol. 2013, 45, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Mehl, K.A.; Davis, J.M.; Berger, F.G.; Carson, J.A. Myofiber degeneration/regeneration is induced in the cachectic ApcMin/+ mouse. J. Appl. Physiol. 2005, 99, 2379–2387. [Google Scholar] [CrossRef]
- Roberts, B.M.; Ahn, B.; Smuder, A.J.; Al-Rajhi, M.; Gill, L.C.; Beharry, A.W.; Powers, S.K.; Fuller, D.D.; Ferreira, L.F.; Judge, A.R. Diaphragm and ventilatory dysfunction during cancer cachexia. FASEB J. 2013, 27, 2600–2610. [Google Scholar] [CrossRef] [Green Version]
- Bohnert, K.R.; Gallot, Y.S.; Sato, S.; Xiong, G.; Hindi, S.M.; Kumar, A. Inhibition of ER stress and unfolding protein response pathways causes skeletal muscle wasting during cancer cachexia. FASEB J. 2016, 30, 3053–3068. [Google Scholar] [CrossRef] [Green Version]
- Chacon-Cabrera, A.; Mateu-Jimenez, M.; Langohr, K.; Fermoselle, C.; García-Arumí, E.; Andreu, A.L.; Yelamos, J.; Barreiro, E. Role of PARP activity in lung cancer-induced cachexia: Effects on muscle oxidative stress, proteolysis, anabolic markers, and phenotype. J. Cell Physiol. 2017, 232, 3744–3761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar-Degracia, A.; Blanco, D.; Vilà-Ubach, M.; de Biurrun, G.; de Solórzano, C.O.; Montuenga, L.M.; Barreiro, E. Phenotypic and metabolic features of mouse diaphragm and gastrocnemius muscles in chronic lung carcinogenesis: Influence of underlying emphysema. J. Transl. Med. 2016, 14, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moylan, J.S.; Reid, M.B. Oxidative stress, chronic disease, and muscle wasting. Muscle Nerve 2007, 35, 411–429. [Google Scholar] [CrossRef]
- Tisdale, M.J. Mechanisms of cancer cachexia. Physiol. Rev. 2009, 89, 381–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoli, I.; Le, H.; Alesci, S.; McFann, K.K.; Su, Y.A.; Kino, T.; Chrousos, G.P.; Blackman, M.R. Monoamine oxidase-A is a major target gene for glucocorticoids in human skeletal muscle cells. FASEB J. 2005, 19, 1359–1361. [Google Scholar] [CrossRef]
- Bianchi, P.; Kunduzova, O.; Masini, E.; Cambon, C.; Bani, D.; Raimondi, L.; Seguelas, M.H.; Nistri, S.; Colucci, W.; Leducq, N.; et al. Oxidative stress by monoamine oxidase mediates receptor-independent cardiomyocyte apoptosis by serotonin and postischemic myocardial injury. Circulation 2005, 112, 3297–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pchejetski, D.; Kunduzova, O.; Dayon, A.; Calise, D.; Seguelas, M.H.; Leducq, N.; Seif, I.; Parini, A.; Cuvillier, O. Oxidative stress-dependent sphingosine kinase-1 inhibition mediates monoamine oxidase A-associated cardiac cell apoptosis. Circ. Res. 2007, 100, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umbarkar, P.; Singh, S.; Arkat, S.; Bodhankar, S.L.; Lohidasan, S.; Sitasawad, S.L. Monoamine oxidase-A is an important source of oxidative stress and promotes cardiac dysfunction, apoptosis, and fibrosis in diabetic cardiomyopathy. Free Radic. Biol. Med. 2015, 87, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Morin, N.; Visentin, V.; Calise, D.; Marti, L.; Zorzano, A.; Testar, X.; Valet, P.; Fischer, Y.; Carpéné, C. Tyramine stimulates glucose uptake in insulin-sensitive tissues in vitro and in vivo via its oxidation by amine oxidases. J. Pharmacol. Exp. Ther. 2002, 303, 1238–1247. [Google Scholar] [CrossRef] [Green Version]
- Duarte, J.A.; Carvalho, F.; Fernandes, E.; Remião, F.; Bastos, M.L.; Magalhães, J.; Appell, H.J. D-amphetamine-induced hydrogen peroxide production in skeletal muscle is modulated by monoamine oxidase inhibition. Int. J. Sports Med. 2004, 25, 446–449. [Google Scholar] [CrossRef]
- Centers for Disease Control, Prevention. An Update on Cancer Deaths in the United States; US Department of Health and Human Services, Centers for Disease Control and Prevention, Division of Cancer Prevention and Control: Atlanta, GA, USA, 2020.
- Dewys, W.D.; Begg, C.; Lavin, P.T.; Band, P.R.; Bennett, J.M.; Bertino, J.R.; Cohen, M.H.; Douglass, H.O.; Engstrom, P.F.; Ezdinli, E.Z.; et al. Prognostic effect of weight loss prior to chemotherapy in cancer patients. Eastern Cooperative Oncology Group. Am. J. Med. 1980, 69, 491–497. [Google Scholar] [CrossRef]
- Wigmore, S.J.; Plester, C.E.; Richardson, R.A.; Fearon, K.C. Changes in nutritional status associated with unresectable pancreatic cancer. Br. J. Cancer 1997, 75, 106–109. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, J.; Ketterer, K.; Marsch, C.; Fechtner, K.; Krakowski-Roosen, H.; Büchler, M.W.; Friess, H.; Martignoni, M.E. Pancreatic cancer related cachexia: Influence on metabolism and correlation to weight loss and pulmonary function. BMC Cancer 2009, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, J.; Büchler, M.W.; Friess, H.; Martignoni, M.E. Cachexia in patients with chronic pancreatitis and pancreatic cancer: Impact on survival and outcome. Nutr. Cancer 2013, 65, 827–833. [Google Scholar] [CrossRef]
- Teunissen, S.C.; Wesker, W.; Kruitwagen, C.; de Haes, H.C.; Voest, E.E.; de Graeff, A. Symptom prevalence in patients with incurable cancer: A systematic review. J. Pain Symptom Manag. 2007, 34, 94–104. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Wang, L.; Multani, A.S.; Combs, C.; Deramaudt, T.B.; Hruban, R.H.; Rustgi, A.K.; Chang, S.; Tuveson, D.A. Trp53R172H and KrasG12D cooperate to promote chromosomal instability and widely metastatic pancreatic ductal adenocarcinoma in mice. Cancer Cell 2005, 7, 469–483. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Komar, C.A.; Bengsch, F.; Graham, K.; Beatty, G.L. Genetically Engineered Mouse Models of Pancreatic Cancer: The KPC Model (LSL-Kras(G12D/+); LSL-Trp53(R172H/+); Pdx-1-Cre), Its Variants, and Their Application in Immuno-oncology Drug Discovery. Curr. Protoc. Pharmacol. 2016, 73, 14.39.11–14.39.20. [Google Scholar] [CrossRef] [Green Version]
- Hasselluhn, M.C.; Klein, L.; Patzak, M.S.; Buchholz, S.M.; Ströbel, P.; Ellenrieder, V.; Maisonneuve, P.; Neesse, A. Stromal Features of the Primary Tumor Are Not Prognostic in Genetically Engineered Mice of Pancreatic Cancer. Cells 2019, 9, 58. [Google Scholar] [CrossRef] [Green Version]
- Fendrich, V.; Chen, N.M.; Neef, M.; Waldmann, J.; Buchholz, M.; Feldmann, G.; Slater, E.P.; Maitra, A.; Bartsch, D.K. The angiotensin-I-converting enzyme inhibitor enalapril and aspirin delay progression of pancreatic intraepithelial neoplasia and cancer formation in a genetically engineered mouse model of pancreatic cancer. Gut 2010, 59, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Fendrich, V.; Schneider, R.; Maitra, A.; Jacobsen, I.D.; Opfermann, T.; Bartsch, D.K. Detection of precursor lesions of pancreatic adenocarcinoma in PET-CT in a genetically engineered mouse model of pancreatic cancer. Neoplasia 2011, 13, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrandt, W.; Keck, J.; Schmich, S.; Bonaterra, G.A.; Wilhelm, B.; Schwarzbach, H.; Eva, A.; Bertoune, M.; Slater, E.P.; Fendrich, V.; et al. Inflammation and Wasting of Skeletal Muscles in Kras-p53-Mutant Mice with Intraepithelial Neoplasia and Pancreatic Cancer-When Does Cachexia Start? Cells 2022, 11, 1607. [Google Scholar] [CrossRef] [PubMed]
- Fritzsche, J. Untersuchungen über die Samen von Peganum Harmala. In Bulletin de la Classe Physico-Mathématique de l’Académie Impériale des Sciences de Saint-Pétersbourg 1847; t.5-8 (1847–1850); Russian Academy of Science: St. Petersbourg, Russia, 1847; Volume 6, pp. 49–72. [Google Scholar]
- Neuner, A.; Tappeiner, H. Ueber die Wirkungen der Alkaloide von Peganum Harmala, insbesondere des Harmalins. Arch. Exp. Pathol. Pharmakol. 1894, 35, 69–76. [Google Scholar] [CrossRef]
- Herraiz, T.; González, D.; Ancín-Azpilicueta, C.; Arán, V.J.; Guillén, H. beta-Carboline alkaloids in Peganum harmala and inhibition of human monoamine oxidase (MAO). Food Chem. Toxicol. 2010, 48, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Alroy, J.; Goyal, V.; Skutelsky, E. Lectin histochemistry of mammalian endothelium. Histochemistry 1987, 86, 603–607. [Google Scholar] [CrossRef]
- Brooke, M.H.; Kaiser, K.K. Muscle fiber types: How many and what kind? Arch. Neurol. 1970, 23, 369–379. [Google Scholar] [CrossRef]
- Friedmann-Bette, B.; Bauer, T.; Kinscherf, R.; Vorwald, S.; Klute, K.; Bischoff, D.; Müller, H.; Weber, M.A.; Metz, J.; Kauczor, H.U.; et al. Effects of strength training with eccentric overload on muscle adaptation in male athletes. Eur. J. Appl. Physiol. 2010, 108, 821–836. [Google Scholar] [CrossRef]
- Bannow, L.I.; Bonaterra, G.A.; Bertoune, M.; Maus, S.; Schulz, R.; Weissmann, N.; Kraut, S.; Kinscherf, R.; Hildebrandt, W. Effect of chronic intermittent hypoxia (CIH) on neuromuscular junctions and mitochondria in slow- and fast-twitch skeletal muscles of mice-the role of iNOS. Skelet. Muscle 2022, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Sander, H.; Wallace, S.; Plouse, R.; Tiwari, S.; Gomes, A.V. Ponceau S waste: Ponceau S staining for total protein normalization. Anal. Biochem. 2019, 575, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: Applications to mammalian blood and other tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Hack, V.; Gross, A.; Kinscherf, R.; Bockstette, M.; Fiers, W.; Berke, G.; Dröge, W. Abnormal glutathione and sulfate levels after interleukin 6 treatment and in tumor-induced cachexia. FASEB J. 1996, 10, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aulino, P.; Berardi, E.; Cardillo, V.M.; Rizzuto, E.; Perniconi, B.; Ramina, C.; Padula, F.; Spugnini, E.P.; Baldi, A.; Faiola, F.; et al. Molecular, cellular and physiological characterization of the cancer cachexia-inducing C26 colon carcinoma in mouse. BMC Cancer 2010, 10, 363. [Google Scholar] [CrossRef]
- Fogelman, D.R.; Morris, J.; Xiao, L.; Hassan, M.; Vadhan, S.; Overman, M.; Javle, S.; Shroff, R.; Varadhachary, G.; Wolff, R.; et al. A predictive model of inflammatory markers and patient-reported symptoms for cachexia in newly diagnosed pancreatic cancer patients. Support. Care Cancer 2017, 25, 1809–1817. [Google Scholar] [CrossRef]
- Barber, M.D.; Powell, J.J.; Lynch, S.F.; Fearon, K.C.; Ross, J.A. A polymorphism of the interleukin-1 beta gene influences survival in pancreatic cancer. Br. J. Cancer 2000, 83, 1443–1447. [Google Scholar] [CrossRef] [Green Version]
- Song, K.S.; Kim, K.; Chung, K.C.; Seol, J.H.; Yoon, J.H. Interaction of SOCS3 with NonO attenuates IL-1beta-dependent MUC8 gene expression. Biochem. Biophys. Res. Commun. 2008, 377, 946–951. [Google Scholar] [CrossRef]
- Qin, H.; Holdbrooks, A.T.; Liu, Y.; Reynolds, S.L.; Yanagisawa, L.L.; Benveniste, E.N. SOCS3 deficiency promotes M1 macrophage polarization and inflammation. J. Immunol. 2012, 189, 3439–3448. [Google Scholar] [CrossRef] [Green Version]
- Sandri, M.; Lin, J.; Handschin, C.; Yang, W.; Arany, Z.P.; Lecker, S.H.; Goldberg, A.L.; Spiegelman, B.M. PGC-1alpha protects skeletal muscle from atrophy by suppressing FoxO3 action and atrophy-specific gene transcription. Proc. Natl. Acad. Sci. USA 2006, 103, 16260–16265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.; Li, J.L. Role of PGC-1α signaling in skeletal muscle health and disease. Ann. N. Y. Acad. Sci. 2012, 1271, 110–117. [Google Scholar] [CrossRef]
- Wang, X.; Pickrell, A.M.; Zimmers, T.A.; Moraes, C.T. Increase in muscle mitochondrial biogenesis does not prevent muscle loss but increased tumor size in a mouse model of acute cancer-induced cachexia. PLoS ONE 2012, 7, e33426. [Google Scholar] [CrossRef]
- Mastropasqua, F.; Girolimetti, G.; Shoshan, M. PGC1α: Friend or Foe in Cancer? Genes 2018, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Tao, Y.; Yang, H. Treatment with harmine ameliorates functional impairment and neuronal death following traumatic brain injury. Mol. Med. Rep. 2015, 12, 7985–7991. [Google Scholar] [CrossRef] [Green Version]
- Hamsa, T.P.; Kuttan, G. Harmine inhibits tumour specific neo-vessel formation by regulating VEGF, MMP, TIMP and pro-inflammatory mediators both in vivo and in vitro. Eur. J. Pharmacol. 2010, 649, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, M.; Tan, S.; Wang, C.; Fan, S.; Huang, C. Harmine is an inflammatory inhibitor through the suppression of NF-κB signaling. Biochem. Biophys. Res. Commun. 2017, 489, 332–338. [Google Scholar] [CrossRef]
- Mietto, B.S.; Mostacada, K.; Martinez, A.M. Neurotrauma and inflammation: CNS and PNS responses. Mediat. Inflamm. 2015, 2015, 251204. [Google Scholar] [CrossRef]
- Villarinho, J.G.; Pinheiro, K.e.V.; Pinheiro, F.e.V.; Oliveira, S.M.; Machado, P.; Martins, M.A.; Bonacorso, H.G.; Zanatta, N.; Fachinetto, R.; Ferreira, J. The antinociceptive effect of reversible monoamine oxidase-A inhibitors in a mouse neuropathic pain model. Prog. Neuro Sychopharmacol. Biol. Psychiatry 2013, 44, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.M.; Renno, W.M.; Sadek, H.L.; Kayali, N.M.; Al-Salem, A.; Rao, M.S.; Khan, K.M. Monoamine oxidase-B inhibitor protects degenerating spinal neurons, enhances nerve regeneration and functional recovery in sciatic nerve crush injury model. Neuropharmacology 2018, 128, 231–243. [Google Scholar] [CrossRef]
- Scalabrin, M.; Pollock, N.; Staunton, C.A.; Brooks, S.V.; McArdle, A.; Jackson, M.J.; Vasilaki, A. Redox responses in skeletal muscle following denervation. Redox Biol. 2019, 26, 101294. [Google Scholar] [CrossRef] [PubMed]
- Pollock, N.; Staunton, C.A.; Vasilaki, A.; McArdle, A.; Jackson, M.J. Denervated muscle fibers induce mitochondrial peroxide generation in neighboring innervated fibers: Role in muscle aging. Free Radic. Biol. Med. 2017, 112, 84–92. [Google Scholar] [CrossRef]
- Daou, N.; Hassani, M.; Matos, E.; De Castro, G.S.; Costa, R.G.F.; Seelaender, M.; Moresi, V.; Rocchi, M.; Adamo, S.; Li, Z.; et al. Displaced Myonuclei in Cancer Cachexia Suggest Altered Innervation. Int. J. Mol. Sci. 2020, 21, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anagnostou, M.E.; Hepple, R.T. Mitochondrial Mechanisms of Neuromuscular Junction Degeneration with Aging. Cells 2020, 9, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa-Caldwell, M.E.; Benson, C.A.; Lee, D.E.; Brown, J.L.; Washington, T.A.; Greene, N.P.; Wiggs, M.P. Mitochondrial Function and Protein Turnover in the Diaphragm are Altered in LLC Tumor Model of Cancer Cachexia. Int. J. Mol. Sci. 2020, 21, 7841. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, J.; Riverón-Negrete, L.; Abdullaev, F.; Espinosa-Aguirre, J.; Rodríguez-Arnaiz, R. Cytotoxicity of the beta-carboline alkaloids harmine and harmaline in human cell assays in vitro. Exp. Toxicol. Pathol. 2008, 60, 381–389. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Suzuki, T.; Ishii, H.; Ogata, A.; Nakae, D. Mitochondrial dysfunction and biotransformation of β-carboline alkaloids, harmine and harmaline, on isolated rat hepatocytes. Chem. Biol. Interact. 2010, 188, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Nafisi, S.; Bonsaii, M.; Maali, P.; Khalilzadeh, M.A.; Manouchehri, F. Beta-carboline alkaloids bind DNA. J. Photochem. Photobiol. B 2010, 100, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Pagano, B.; Caterino, M.; Filosa, R.; Giancola, C. Binding of Harmine Derivatives to DNA: A Spectroscopic Investigation. Molecules 2017, 22, 1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.R.; Li, Q.; Liu, Z.L.; Liu, H.H.; Wang, W.; Liao, X.L.; Pan, Y.L.; Jiang, J.W. Harmine induces apoptosis in HepG2 cells via mitochondrial signaling pathway. Hepatobiliary Pancreat. Dis. Int. 2011, 10, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Huang, C.R.; Wang, W.; Zhang, X.K.; Chen, J.J.; Wang, J.J.; Lin, C.; Jiang, J.W. Harmine Hydrochloride Triggers G2 Phase Arrest and Apoptosis in MGC-803 Cells and SMMC-7721 Cells by Upregulating p21, Activating Caspase-8/Bid, and Downregulating ERK/Bad Pathway. Phytother. Res. 2016, 30, 31–40. [Google Scholar] [CrossRef]
- Sobhani, A.M.; Ebrahimi, S.A.; Mahmoudian, M. An in vitro evaluation of human DNA topoisomerase I inhibition by Peganum harmala L. seeds extract and its beta-carboline alkaloids. J. Pharm. Pharm. Sci. 2002, 5, 19–23. [Google Scholar]
- Song, Y.; Kesuma, D.; Wang, J.; Deng, Y.; Duan, J.; Wang, J.H.; Qi, R.Z. Specific inhibition of cyclin-dependent kinases and cell proliferation by harmine. Biochem. Biophys. Res. Commun. 2004, 317, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Giacobbo, B.L.; Doorduin, J.; Moraga-Amaro, R.; Nazario, L.R.; Schildt, A.; Bromberg, E.; Dierckx, R.A.J.O.; de Vries, E.F.J. Chronic harmine treatment has a delayed effect on mobility in control and socially defeated rats. Psychopharmacology 2020, 237, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- Arque, G.; Casanovas, A.; Dierssen, M. Dyrk1A is dynamically expressed on subsets of motor neurons and in the neuromuscular junction: Possible role in Down syndrome. PLoS ONE 2013, 8, e54285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Lee, K.S.; Kim, A.K.; Choi, M.; Choi, K.; Kang, M.; Chi, S.W.; Lee, M.S.; Lee, J.S.; Lee, S.Y.; et al. A chemical with proven clinical safety rescues Down-syndrome-related phenotypes in through DYRK1A inhibition. Dis. Model. Mech. 2016, 9, 839–948. [Google Scholar]
- Habib, M.Z.; Tadros MGAbd-Alkhalek, H.A.; Mohamad, M.I.; Eid, D.M.; Hassan, F.E.; Elhelaly, H.; Faramawy, Y.E.; Aboul-Fotouh, S. Harmine prevents 3-nitropropionic acid-induced neurotoxicity in rats via enhancing NRF2-mediated signaling: Involvement of p21 and AMPK. Eur. J. Pharmacol. 2022, 927, 175046. [Google Scholar] [CrossRef]
- Deng, X.; Mercer, S.E.; Sun, C.Y.; Friedman, E. The normal function of the cancer kinase Mirk/dyrk1B is to reduce reactive oxygen species. Genes Cancer 2014, 5, 22–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, E. Mirk/Dyrk1B in Cancer. J. Cell. Biochem. 2007, 102, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Bhat, N.; Narayanan, A.; Fathzadeh, M.; Shah, K.; Dianatpour, M.; Abou Ziki, M.D.; Mani, A. Dyrk1b promotes autophagy during skeletal muscle differentiation by upregulating 4e-bp1. Cell Signal. 2022, 90, 110186. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Assay | Symbol | Amplicon Length (bp) | Cat. No. |
|---|---|---|---|
| Actin Beta | ACTB | 77 | QT01136772 |
| BCL2 Associated X, Apoptosis Regulator | BAX | 76 | QT00102536 |
| BCL2 Apoptosis Regulator | BCL-2 | 104 | QT02392292 |
| Caspase 3 | Caspase3 | 150 | QT01164779 |
| CD68 Molecule | CD68 | 67 | QT00254051 |
| F-Box Protein 32 | FBXO32 | 103 | QT00158543 |
| Interleukin 1 Beta | IL-1β | 150 | QT01048355 |
| Interleukin 6 | IL-6 | 128 | QT00098875 |
| Monoamine Oxidase A | MAO-A | 81 | QT00109326 |
| Monoamine Oxidase B | MAO-B | 93 | QT00145124 |
| Matrix Metallopeptidase 9 | MMP9 | 84 | QT00108815 |
| Myogenin | Myog | 115 | QT00112378 |
| Paired Box Protein 7 | PAX7 | 135 | QT00147728 |
| PPARG Coactivator 1 Alpha | Ppargc1a | 63 | QT00156303 |
| Prostaglandin-Endoperoxide Synthase 2 | PTGS2 (COX2) | 95 | QT00165347 |
| Retention of Endoplasmic Reticulum protein 1 (S. cerevisiae) | RER1 | 86 | QT00146580 |
| Suppressor of Cytokine Signaling 3 | SOCS3 | 90 | QT00156303 |
| Sequestosome 1 | Sqstm1 (P62) | 91 | QT00127855 |
| TATA Box Binding Protein | TBP | 114 | QT00198443 |
| Tumor Necrosis Factor | TNFα | 112 | QT00104006 |
| Tripartite Motif Containing 63 | TRIM63(MuRF1) | 116 | QT00291991 |
| Vascular Endothelial Growth Factor A | VEGFA | 117 | QT00160769 |
| WT | Ca | WTHH | CaHH | ANOVA | |||
|---|---|---|---|---|---|---|---|
| PDAC | HH | Interaction | |||||
| n (m/f) | 11(5/6) | 11 (7/4) | 11 (8/3) | 7 (4/3) | - | - | - |
| weight (g) | 27.88 ± 6.16 | 27.65 ± 3.47 | 27.17 ± 4.52 | 24.03 ± 3.16 | p = 0.314 | p = 0.186 | p = 0.332 |
| age(months) | 3.91 ± 0.54 | 5.42 ± 0.77 | 4.61 ± 0.11 | 4.50 ± 0.36 | p < 0.001 | p = 0.552 | p < 0.001 |
| WT | Ca | WTHH | CaHH | ANOVA | ||||
|---|---|---|---|---|---|---|---|---|
| PDAC | HH | Interaction | ||||||
| n (m/f) | 9 (4/5) | 11 (7/4) | 11 (8/3) | 7 (4/3) | - | - | - | |
| Apoptosis/Atrophy | BAX | 1 ± 1.57 | 0.65 ± 0.67 | 0.42 ± 0.58 | 0.73 ± 0.69 | p = 0.902 | p = 0.406 | p = 0.327 |
| BCL-2 | 1 ± 0.57 | 1.06 ± 0.58 | 1.62 ± 2.19 | 1.11 ± 0.85 | p = 0.662 | p = 0.441 | p = 0.535 | |
| Caspase 3 | 1 ± 1.09 | 0.79 ± 0.44 | 0.69 ± 0.36 | 0.92 ± 0.82 | p = 0.977 | p = 0.661 | p = 0.388 | |
| MuRF1 | 1 ± 2.45 | 0.14 ± 0.16 | 0.19 ± 0.53 | 0.11 ± 0.11 | p = 0.252 | p = 0.298 | p = 0.371 | |
| Fbxo32 | 1 ± 0.82 | 2.36 ± 2.84 | 0.57 ± 0.31 | 0.87 ± 0.43 | p = 0.128 | p = 0.107 | p = 0.349 | |
| MAO-A | 1 ± 1.44 | 0.55 ± 0.49 | 0.20 ± 0.10 | 0.52 ± 0.54 | p = 0.743 | p = 0.113 | p = 0.168 | |
| MAO-B | 1 ± 1.52 | 0.45 ± 0.54 | 0.21 ± 0.31 | 0.33 ± 0.31 | p = 0.407 | p = 0.104 | p = 0.253 | |
| MMP9 | 1 ± 2.43 | 0.25 ± 0.23 | 0.17 ± 0.37 | 0.15 ± 0.18 | p = 0.337 | p = 0.252 | p = 0.386 | |
| Inflammation | CD 68 | 1 ± 2.01 | 0.68 ± 0.69 | 0.64 ± 1.53 | 1.99 ± 2.63 | p = 0.457 | p = 0.499 | p = 0.180 |
| TNFα | 1 ± 0.99 | 1.65 ± 2.15 | 0.53 ± 0.51 | 0.51 ± 0.45 | p = 0.461 | p = 0.093 | p = 0.466 | |
| IL-6 | 1 ± 1.52 | 0.83 ± 0.77 | 0.30 ± 0.58 | 1.24 ± 2.29 | p = 0.461 | p = 0.689 | p = 0.239 | |
| COX2 | 1 ± 1.06 | 1.45 ± 2.27 | 0.60 ± 0.62 | 0.20 ± 0.36 | p = 0.903 | p = 0.105 | p = 0.381 |
| WT | Ca | WTHH | CaHH | ANOVA | |||
|---|---|---|---|---|---|---|---|
| PDAC | HH | Interaction | |||||
| n (m/f) | 11 (6/5) | 11 (7/4) | 10 (7/3) | 4 (2/2) | - | - | - |
| tGSH (pmol/g) | 10.40 ± 0.41 | 8.41 ± 0.81 | 5.32 ± 0.98 | 5.45 ± 1.97 | p = 0.342 | p < 0.001 | p = 0.281 |
| rGSH (pmol/g) | 7.89 ± 0.44 | 7.07 ± 0.83 | 4.03 ± 0.94 | 3.66 ± 1.81 | p = 0.536 | p = 0.001 | p = 0.811 |
| GSSG (pmol/g) | 2.51 ± 0.18 | 1.35 ± 0.19 | 1.29 ± 0.14 | 1.79 ± 0.69 | p = 0.189 | p = 0.129 | p = 0.002 |
| rGSH/GSSG (ratio) | 3.31 ± 0.34 | 6.25 ± 0.92 | 3.18 ± 0.84 | 3.88 ± 1.94 | p = 0.062 | p = 0.192 | p = 0.241 |
| WT | Ca | WTHH | CaHH | ANOVA | |||
|---|---|---|---|---|---|---|---|
| PDAC | HH | Interaction | |||||
| n (m/f) | 11 (5/6) | 8 (5/3) | 6 (6/0) | 4 (2/2) | - | - | - |
| CSA (µm2) | 1351 ± 299 | 1154 ± 86 | 1286 ± 199 | 1037 ± 31 | p = 0.019 | p = 0.338 | p = 0.777 |
| minFeret (µm) | 33.66 ± 4.11 | 31.15 ± 1.48 | 31.75 ± 2.33 | 28.54 ± 2.33 | p = 0.032 | p = 0.092 | p = 0.789 |
| capillary contacts/all fiber types (n/fiber) | 4.35 ± 0.81 | 4.49 ± 0.73 | 4.39 ± 0.49 | 4.02 ± 0.38 | p = 0.896 | p = 0.573 | p = 0.383 |
| capillary contacts/fiber type I (n/fiber) | 4.50 ± 0.90 | 4.57 ± 0.81 | 4.56 ± 0.54 | 4.17 ± 0.45 | p = 0.791 | p = 0.683 | p = 0.483 |
| capillary contacts/fiber type IIA (n/fiber) | 4.23 ± 0.79 | 4.37 ± 0.67 | 4.25 ± 0.52 | 3.83 ± 0.27 | p = 0.861 | p = 0.449 | p = 0.323 |
| capillary contacts/fiber type IIBX (n/fiber) | 4.08 ± 1.07 | 4.26 ± 0.98 | 4.47 ± 0.62 | 3.70 ± 0.55 | p = 0.705 | p = 0.999 | p = 0.233 |
| capillary density (n/mm2) | 1144 ± 176 | 1265 ± 246 | 1186 ± 308 | 1248 ± 141 | p = 0.282 | p = 0.851 | p = 0.762 |
| WT | Ca | WTHH | CaHH | ANOVA | ||||
|---|---|---|---|---|---|---|---|---|
| PDAC | HH | Interaction | ||||||
| n (m/f) | 11 (6/5) | 7 (5/2) | 6 (4/2) | 6 (3/3) | - | - | - | |
| Apoptosis/Atrophy | BCL-2 | 1 ± 0.52 | 0.94 ± 0.53 | 0.50 ± 0.36 | 0.70 ± 0.71 | p = 0.836 | p = 0.088 | p = 0.564 |
| Caspase 3 | 1 ± 0.47 | 0.92 ± 0.37 | 1.22 ± 0.58 | 0.49 ± 0.42 | p = 0.073 | p = 0.678 | p = 0.094 | |
| p62 | 1 ± 0.16 | 1.19 ± 0.34 | 1.05 ± 0.09 | 1.67 ± 1.10 | p = 0.090 | p = 0.262 | p = 0.321 | |
| MAO-A | 1 ± 0.21 | 0.94 ± 0.25 | 0.69 ± 0.10 | 1.00 ± 0.36 | p = 0.332 | p = 0.162 | p = 0.070 | |
| MAO-B | 1 ± 0.23 | 0.89 ± 0.13 | 0.79 ± 0.15 | 0.94 ± 0.48 | p = 0.949 | p = 0.422 | p = 0.245 | |
| MMP9 | 1 ± 0.61 | 0.63 ± 0.36 | 0.37 ± 0.22 | 0.50 ± 0.49 | p = 0.405 | p = 0.043 | p = 0.205 | |
| Inflammation | CD 68 | 1 ± 0.24 | 1.40 ± 0.49 | 1.59 ± 0.93 | 4.61 ± 5.41 | p = 0.145 | p = 0.086 | p = 0.203 |
| TNFα | 1 ± 0.81 | 1.16 ± 1.09 | 1.24 ± 0.40 | 0.65 ± 0.80 | p = 0.661 | p = 0.758 | p = 0.270 | |
| IL-6 | 1 ± 0.34 | 1.18 ± 0.63 | 0.95 ± 0.34 | 1.43 ± 1.52 | p = 0.336 | p = 0.788 | p = 0.639 | |
| COX 2 | 1 ± 1.09 | 2.55 ± 2.08 | 1.77 ± 1.21 | 1.22 ± 2.72 | p = 0.346 | p = 0.797 | p = 0.160 | |
| Socs3 | 1 ± 1.12 | 2.32 ± 1.89 | 0.71 ± 0.28 | 0.84 ± 0.59 | p = 0.088 | p = 0.091 | p = 0.219 | |
| Angiogenesis | VEGFA | 1 ± 0.24 | 1.14 ± 0.21 | 1.11 ± 0.52 | 1.13 ± 0.94 | p = 0.657 | p = 0.787 | p = 0.797 |
| Myogenesis/ | Ppargc1a | 1 ± 1.02 | 1.74 ± 1.05 | 1.99 ± 1.42 | 1.54 ± 1.21 | p = 0.602 | p = 0.340 | p = 0.213 |
| MyoG | 1 ± 0.41 | 0.92 ± 0.22 | 0.55 ± 0.16 | 1.83 ± 1.71 | p = 0.152 | p = 0.616 | p = 0.048 | |
| PAX7 | 1 ± 0.62 | 1.03 ± 0.59 | 1.04 ± 0.71 | 0.72 ± 0.78 | p = 0.662 | p = 0.660 | p = 0.526 | |
| WT | Ca | WTHH | CaHH | ANOVA | |||
|---|---|---|---|---|---|---|---|
| PDAC | HH | Interaction | |||||
| Gastrocnemius muscle | |||||||
| n (m/f) | 9 (3/6) | 7 (4/3) | 11 (8/3) | 6 (3/3) | - | - | - |
| % all fibers | 0.52 ± 0.23 | 0.71 ± 0.29 | 0.83 ± 0.27 | 2.17 ± 0.10 | p = 0.240 | p < 0.133 | p = 0.532 |
| Soleus muscle | |||||||
| n (m/f) | 11 (5/6) | 8 (5/3) | 6 (6/0) | 4 (2/2) | - | - | - |
| % all fibers | 0.08 ± 0.08 | 0.31 ± 0.21 | 0.63 ± 0.31 | 0.75 ± 0.55 | p = 0.475 | p = 0.059 | p = 0.815 |
| % I | 0.00 ± 0.00 | 0.51 ± 0.34 | 0.58 ± 0.36 | 0.71 ± 0.71 | p = 0.325 | p = 0.243 | p = 0.569 |
| % IIA | 0.18 ± 0.18 | 0.16 ± 0.16 | 0.00 ± 0.00 | 0.33 ± 0.33 | p = 0.443 | p = 0.975 | p = 0.395 |
| % IIBX | 00.0 ± 0.00 | 0.00 ± 0.00 | 2.50 ± 1.60 | 1.79 ± 1.79 | p = 0.679 | p = 0.019 | p = 0.679 |
| Gastrocnemius and soleus muscle IIBX fibers | |||||||
| n (m/f) | 20 (8/12) | 15 (9/6) | 17 (14/3) | 10 (5/5) | - | - | - |
| % IIBX | 0.23 ± 0.12 | 0.33 ± 0.16 | 1.41 ± 0.59 | 1.58 ± 0.74 | p = 0.760 | p = 0.006 | p = 0.938 |
| WT | Ca | WTHH | CaHH | ANOVA | |||
|---|---|---|---|---|---|---|---|
| PDAC | HH | Interaction | |||||
| n (m/f) | 5 (3/2) | 4 (4/0) | 5 (3/2) | 5 (3/2) | - | - | - |
| BTX + area (µm2) | 2.85 ± 0.18 | 3.06 ± 0.40 | 2.55 ± 0.14 | 2.17 ± 0.10 | p = 0.680 | p = 0.013 | p = 0.182 |
| vAChT + area (µm2) | 3.12 ± 0.28 | 4.31 ± 0.58 | 3.65 ± 0.27 | 2.73 ± 0.09 | p = 0.683 | p = 0.124 | p = 0.005 |
| vAChT/BTX overlap (µm2) | 0.53 ± 0.03 | 0.98 ± 0.25 | 0.68 ± 0.09 | 0.48 ± 0.03 | p = 0.279 | p = 0.127 | p = 0.010 |
| vAChT/BTX overlap (%) | 18.61 ± 0.57 | 30.54 ± 4.20 | 26.40 ± 2.34 | 21.99 ± 1.60 | p = 0.125 | p = 0.872 | p = 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmich, S.K.P.; Keck, J.; Bonaterra, G.A.; Bertoune, M.; Adam, A.; Wilhelm, B.; Slater, E.P.; Schwarzbach, H.; Fendrich, V.; Kinscherf, R.; et al. Effects of Monoamino-Oxidase-A (MAO-A) Inhibition on Skeletal Muscle Inflammation and Wasting through Pancreatic Ductal Adenocarcinoma in Triple Transgenic Mice. Biomedicines 2023, 11, 912. https://doi.org/10.3390/biomedicines11030912
Schmich SKP, Keck J, Bonaterra GA, Bertoune M, Adam A, Wilhelm B, Slater EP, Schwarzbach H, Fendrich V, Kinscherf R, et al. Effects of Monoamino-Oxidase-A (MAO-A) Inhibition on Skeletal Muscle Inflammation and Wasting through Pancreatic Ductal Adenocarcinoma in Triple Transgenic Mice. Biomedicines. 2023; 11(3):912. https://doi.org/10.3390/biomedicines11030912
Chicago/Turabian StyleSchmich, Simon K. P., Jan Keck, Gabriel A. Bonaterra, Mirjam Bertoune, Anna Adam, Beate Wilhelm, Emily P. Slater, Hans Schwarzbach, Volker Fendrich, Ralf Kinscherf, and et al. 2023. "Effects of Monoamino-Oxidase-A (MAO-A) Inhibition on Skeletal Muscle Inflammation and Wasting through Pancreatic Ductal Adenocarcinoma in Triple Transgenic Mice" Biomedicines 11, no. 3: 912. https://doi.org/10.3390/biomedicines11030912