
Retinal Pigment Epithelium Cell Development: Extrapolating Basic Biology to Stem Cell Research

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
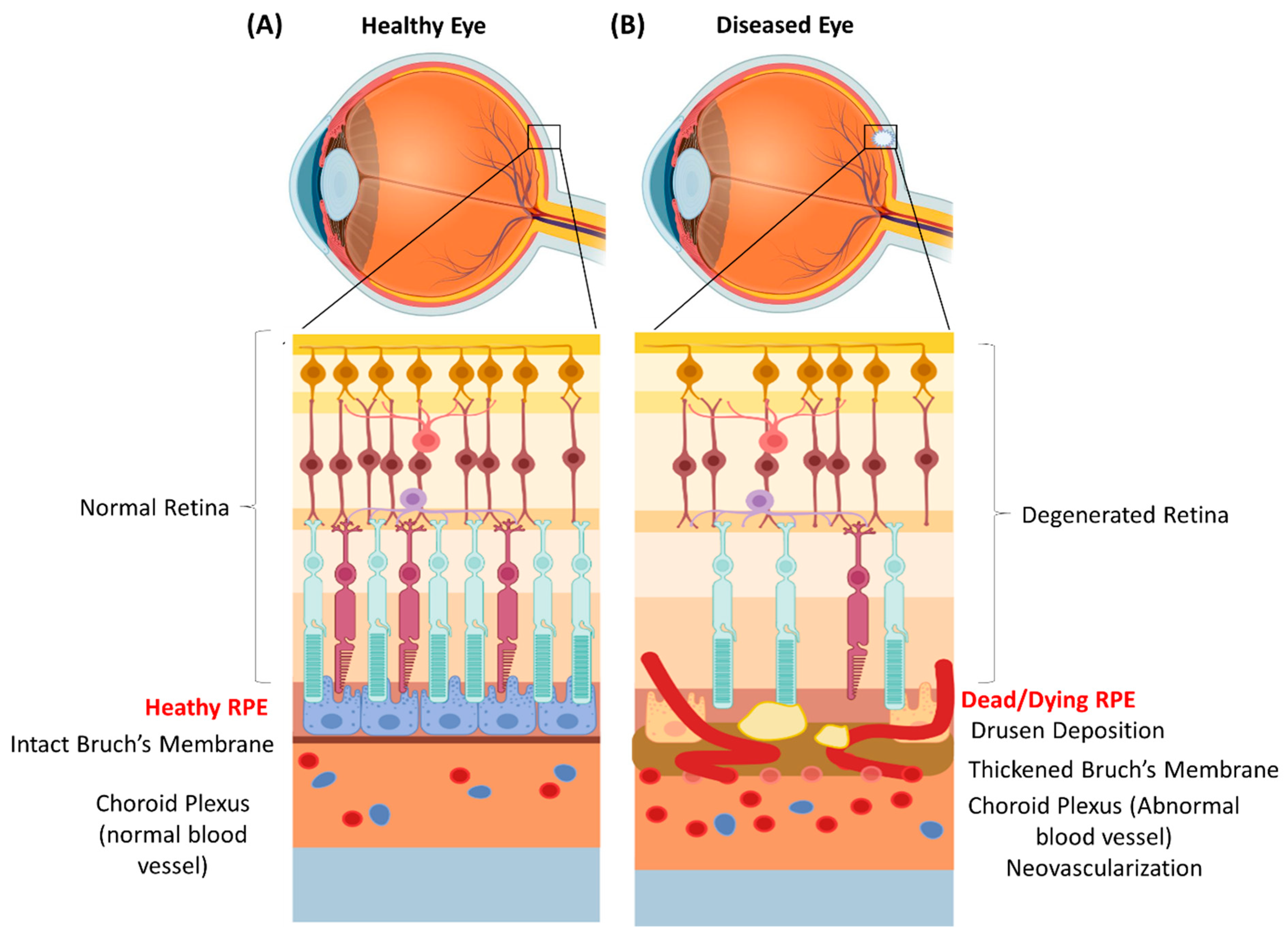
2. RPE Physiology and Function
3. RPE Origin and Developmental Progression
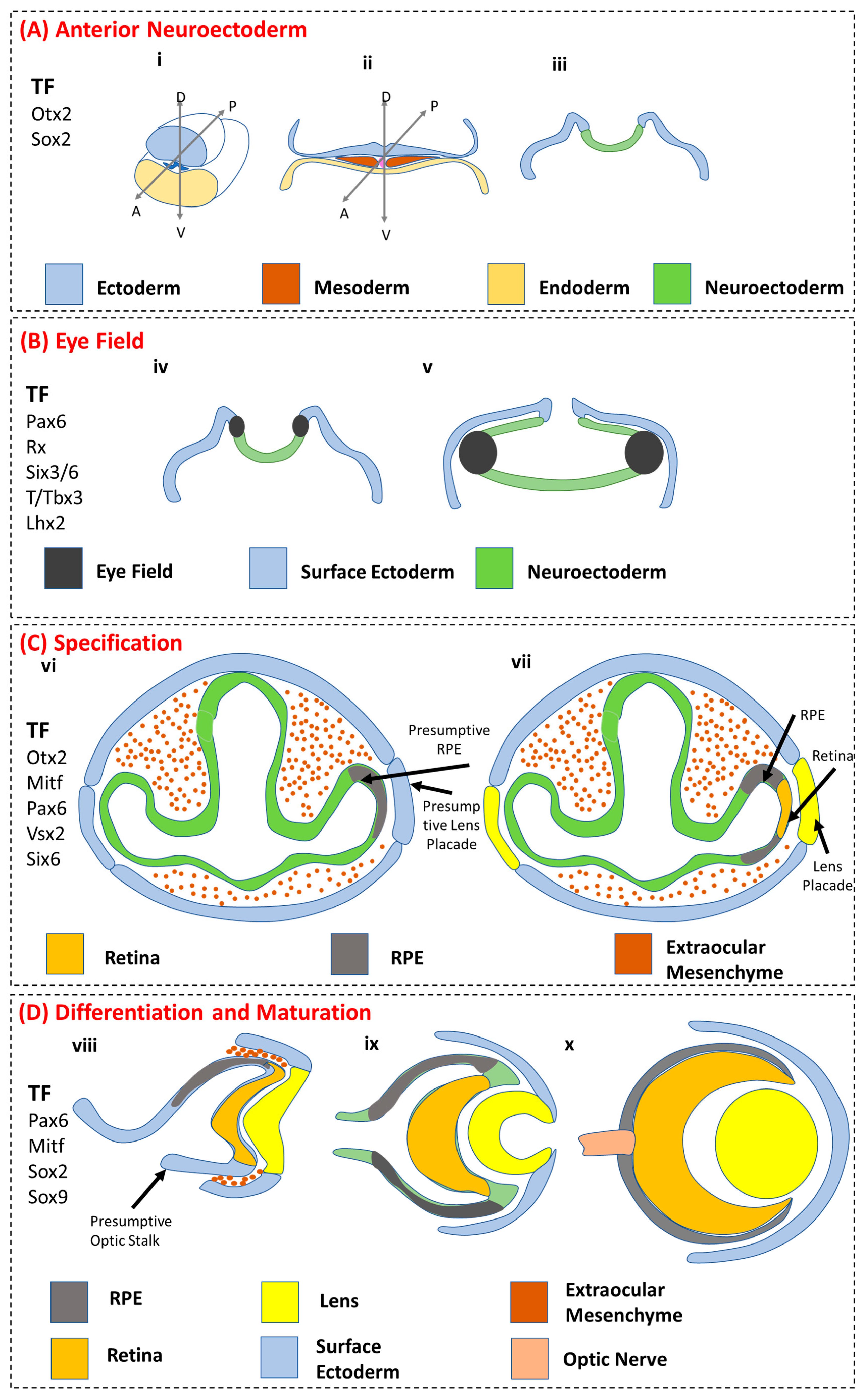
3.1. Anterior Neuroectoderm
3.2. Eye Field
3.3. RPE Specification
3.4. RPE Differentiation and Maturation
4. Genetic and Molecular Regulation of RPE Development and Differentiation
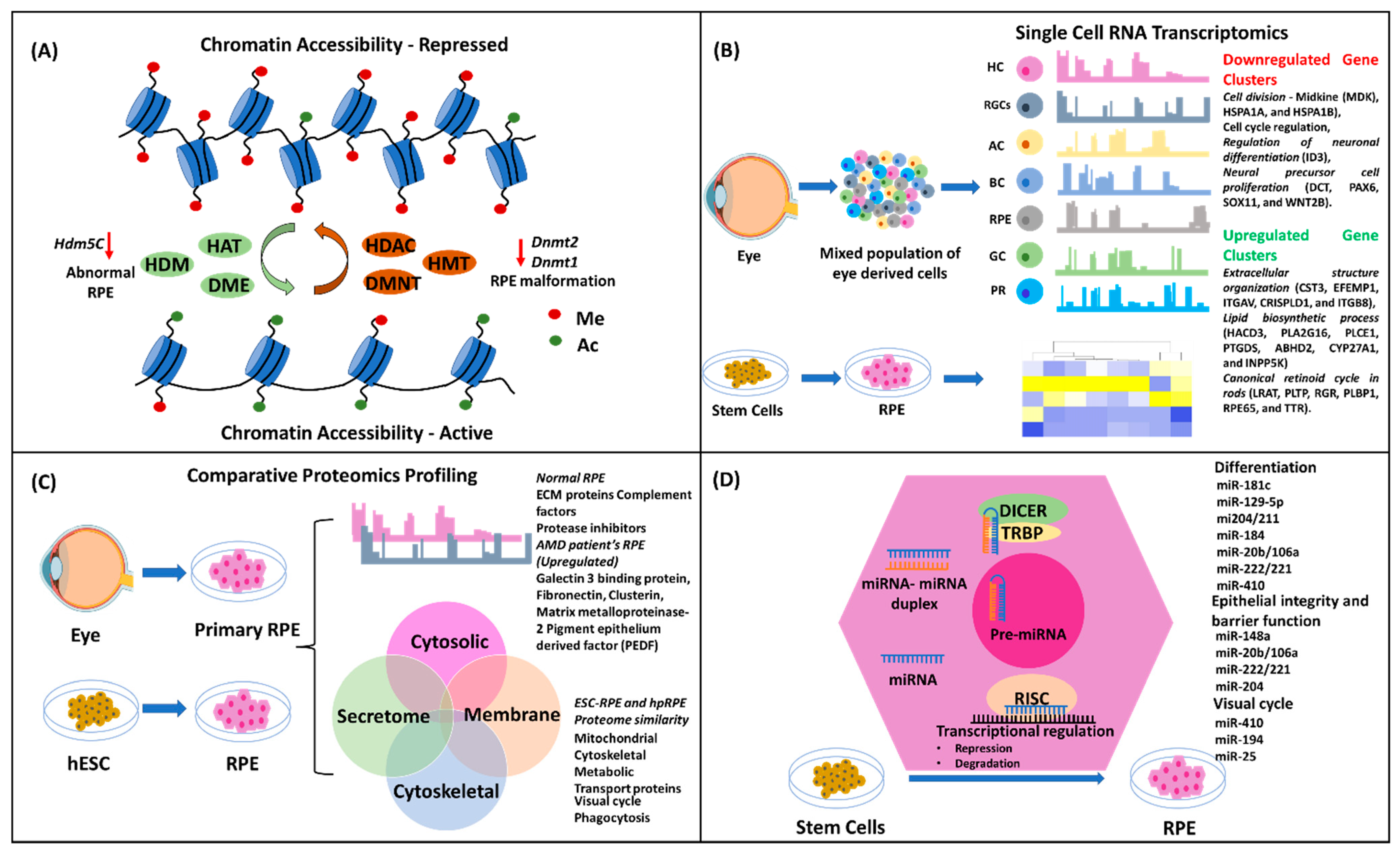
4.1. Epigenetics
4.2. Transcriptomics
4.3. Proteomics
4.4. MicroRNA
5. Significance of the Developmental Studies
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jager, R.D.; Mieler, W.F.; Miller, J.W. Age-Related Macular Degeneration. N. Engl. J. Med. 2008, 358, 2606–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heesterbeek, T.J.; Lorés-Motta, L.; Hoyng, C.B.; Lechanteur, Y.T.E.; den Hollander, A.I. Risk factors for progression of age-related macular degeneration. Ophthalmic Physiol. Opt. 2020, 40, 140–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cachafeiro, M.; Bemelmans, A.-P.; Samardzija, M.; Afanasieva, T.; Pournaras, J.-A.; Grimm, C.; Kostic, C.; Philippe, S.; Wenzel, A.; Arsenijevic, Y. Hyperactivation of retina by light in mice leads to photoreceptor cell death mediated by VEGF and retinal pigment epithelium permeability. Cell Death Dis. 2013, 4, e781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLeod, D.S.; Grebe, R.; Bhutto, I.; Merges, C.; Baba, T.; Lutty, G.A. Relationship between RPE and Choriocapillaris in Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4982–4991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugazhendhi, A.; Hubbell, M.; Jairam, P.; Ambati, B. Neovascular Macular Degeneration: A Review of Etiology, Risk Factors, and Recent Advances in Research and Therapy. Int. J. Mol. Sci. 2021, 22, 1170. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhou, J.; Li, D. Functions and Diseases of the Retinal Pigment Epithelium. Front. Pharmacol. 2021, 12, 1170. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kim, Y.; Oh, Y. Inflammatory pathways in pathological neovascularization in retina and choroid: A narrative review on the inflammatory drug target molecules in retinal and choroidal neovascularization. Ann. Eye Sci. 2021, 6, 1–17. [Google Scholar] [CrossRef]
- Zarbin, M. Cell-Based Therapy for Degenerative Retinal Disease. Trends Mol. Med. 2016, 22, 115–134. [Google Scholar] [CrossRef]
- Cai, J.; Nelson, K.C.; Wu, M.; Sternberg, P.; Jones, D.P. Oxidative damage and protection of the RPE. Prog. Retin. Eye Res. 2000, 19, 205–221. [Google Scholar] [CrossRef]
- Whitmore, S.S.; Sohn, E.H.; Chirco, K.R.; Drack, A.V.; Stone, E.M.; Tucker, B.A.; Mullins, R.F. Complement activation and choriocapillaris loss in early AMD: Implications for pathophysiology and therapy. Prog. Retin. Eye Res. 2015, 45, 1–29. [Google Scholar] [CrossRef]
- Feher, J.; Kovacs, I.; Artico, M.; Cavallotti, C.; Papale, A.; Balacco Gabrieli, C. Mitochondrial alterations of retinal pigment epithelium in age-related macular degeneration. Neurobiol. Aging 2006, 27, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.E.; DeWeerd, A.J.; Ildefonso, C.J.; Lewin, A.S.; Ash, J.D. Mitochondrial oxidative stress in the retinal pigment epithelium (RPE) led to metabolic dysfunction in both the RPE and retinal photoreceptors. Redox Biol. 2019, 24, 101201. [Google Scholar] [CrossRef]
- Strauss, O. The Retinal Pigment Epithelium in Visual Function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somasundaran, S.; Constable, I.J.; Mellough, C.B.; Carvalho, L.S. Retinal pigment epithelium and age-related macular degeneration: A review of major disease mechanisms. Clin. Experiment. Ophthalmol. 2020, 48, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Bose, D.; Maminishkis, A.; Bharti, K. Retinal Pigment Epithelium Replacement Therapy for Age-Related Macular Degeneration: Are We There Yet? Annu. Rev. Pharmacol. Toxicol. 2020, 60, 553–572. [Google Scholar] [CrossRef]
- Fuhrmann, S.; Zou, C.; Levine, E.M. Retinal pigment epithelium development, plasticity, and tissue homeostasis. Exp. Eye Res. 2014, 123, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Holz, F.G.; Schmitz-Valckenberg, S.; Fleckenstein, M. Recent developments in the treatment of age-related macular degeneration. J. Clin. Investig. 2014, 124, 1430–1438. [Google Scholar] [CrossRef] [Green Version]
- Ammar, M.J.; Hsu, J.; Chiang, A.; Ho, A.C.; Regillo, C.D. Age-related macular degeneration therapy: A review. Curr. Opin. Ophthalmol. 2020, 31, 215–221. [Google Scholar] [CrossRef]
- Zarbin, M.; Sugino, I.; Townes-Anderson, E. Concise Review: Update on Retinal Pigment Epithelium Transplantation for Age-Related Macular Degeneration. Stem Cells Transl. Med. 2019, 8, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Robredo, P.; Sancho, A.; Johnen, S.; Recalde, S.; Gama, N.; Thumann, G.; Groll, J.; García-Layana, A. Current Treatment Limitations in Age-Related Macular Degeneration and Future Approaches Based on Cell Therapy and Tissue Engineering. J. Ophthalmol. 2014, 2014, 510285. [Google Scholar] [CrossRef]
- Baradaran-Rafii, A.; Sarvari, M.; Alavi-Moghadam, S.; Payab, M.; Goodarzi, P.; Aghayan, H.R.; Larijani, B.; Rezaei-Tavirani, M.; Biglar, M.; Arjmand, B. Cell-based approaches towards treating age-related macular degeneration. Cell Tissue Bank. 2020, 21, 339–347. [Google Scholar] [CrossRef]
- Forest, D.L.; Johnson, L.V.; Clegg, D.O. Cellular models and therapies for age-related macular degeneration. Dis. Model. Mech. 2015, 8, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.D.; Hubschman, J.-P.; Heilwell, G.; Franco-Cardenas, V.; Pan, C.K.; Ostrick, R.M.; Mickunas, E.; Gay, R.; Klimanskaya, I.; Lanza, R. Embryonic stem cell trials for macular degeneration: A preliminary report. Lancet 2012, 379, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; et al. Autologous Induced Stem-Cell–Derived Retinal Cells for Macular Degeneration. N. Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef] [PubMed]
- van Meurs, J.C.; ter Averst, E.; Hofland, L.J.; van Hagen, P.M.; Mooy, C.M.; Baarsma, G.S.; Kuijpers, R.W.; Boks, T.; Stalmans, P. Autologous peripheral retinal pigment epithelium translocation in patients with subfoveal neovascular membranes. Br. J. Ophthalmol. 2004, 88, 110–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, S.; Krebs, I.; Hilgers, R.-D.; Abri, A.; Stolba, U.; Assadoulina, A.; Kellner, L.; Stanzel, B.V.; Jahn, C.; Feichtinger, H. Outcome of Transplantation of Autologous Retinal Pigment Epithelium in Age-Related Macular Degeneration: A Prospective Trial. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4151–4160. [Google Scholar] [CrossRef]
- Knoernschild, T.; Grasbon, T.; Wilsch, C.; Kampik, A.; Lütjen-Drecoll, E. RPE cell transplants to non-immune-privileged sites of the eye transform into fibroblast-like cells. Curr. Eye Res. 2003, 27, 25–34. [Google Scholar] [CrossRef]
- Lytvynchuk, L.; Ebbert, A.; Studenovska, H.; Nagymihály, R.; Josifovska, N.; Rais, D.; Popelka, Š.; Tichotová, L.; Nemesh, Y.; Čížková, J.; et al. Subretinal Implantation of Human Primary RPE Cells Cultured on Nanofibrous Membranes in Minipigs. Biomedicines 2022, 10, 669. [Google Scholar] [CrossRef]
- Xiang, P.; Wu, K.-C.; Zhu, Y.; Xiang, L.; Li, C.; Chen, D.-L.; Chen, F.; Xu, G.; Wang, A.; Li, M.; et al. A novel Bruch’s membrane-mimetic electrospun substrate scaffold for human retinal pigment epithelium cells. Biomaterials 2014, 35, 9777–9788. [Google Scholar] [CrossRef] [Green Version]
- Kashani, A.H.; Lebkowski, J.S.; Rahhal, F.M.; Avery, R.L.; Salehi-Had, H.; Dang, W.; Lin, C.-M.; Mitra, D.; Zhu, D.; Thomas, B.B.; et al. A bioengineered retinal pigment epithelial monolayer for advanced, dry age-related macular degeneration. Sci. Transl. Med. 2018, 10, eaao4097. [Google Scholar] [CrossRef]
- Sharma, R.; Khristov, V.; Rising, A.; Jha, B.S.; Dejene, R.; Hotaling, N.; Li, Y.; Stoddard, J.; Stankewicz, C.; Wan, Q.; et al. Clinical-grade stem cell–derived retinal pigment epithelium patch rescues retinal degeneration in rodents and pigs. Sci. Transl. Med. 2019, 11, eaat5580. [Google Scholar] [CrossRef]
- Kuroda, T.; Ando, S.; Takeno, Y.; Kishino, A.; Kimura, T. Robust induction of retinal pigment epithelium cells from human induced pluripotent stem cells by inhibiting FGF/MAPK signaling. Stem Cell Res. 2019, 39, 101514. [Google Scholar] [CrossRef]
- Choudhary, P.; Booth, H.; Gutteridge, A.; Surmacz, B.; Louca, I.; Steer, J.; Kerby, J.; Whiting, P.J. Directing Differentiation of Pluripotent Stem Cells Toward Retinal Pigment Epithelium Lineage. Stem Cells Transl. Med. 2017, 6, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Boulton, M.; Dayhaw-Barker, P. The role of the retinal pigment epithelium: Topographical variation and ageing changes. Eye 2001, 15, 384–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, R.; Canto-Soler, M.V. Molecular mechanisms of optic vesicle development: Complexities, ambiguities and controversies. Dev. Biol. 2007, 305, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zaghloul, N.A.; Yan, B.; Moody, S.A. Step-wise specification of retinal stem cells during normal embryogenesis. Biol. Cell 2005, 97, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Zuber, M.E.; Gestri, G.; Viczian, A.S.; Barsacchi, G.; Harris, W.A. Specification of the vertebrate eye by a network of eye field transcription factors. Development 2003, 130, 5155–5167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, K.M.; Otsuna, H.; Kidokoro, H.; Carney, K.R.; Saijoh, Y.; Chien, C.-B. A complex choreography of cell movements shapes the vertebrate eye. Development 2012, 139, 359–372. [Google Scholar] [CrossRef] [Green Version]
- England, S.J.; Blanchard, G.B.; Mahadevan, L.; Adams, R.J. A dynamic fate map of the forebrain shows how vertebrate eyes form and explains two causes of cyclopia. Development 2006, 133, 4613–4617. [Google Scholar] [CrossRef] [Green Version]
- Muranishi, Y.; Terada, K.; Furukawa, T. An essential role for Rax in retina and neuroendocrine system development. Dev. Growth Differ. 2012, 54, 341–348. [Google Scholar] [CrossRef]
- Mathers, P.H.; Grinberg, A.; Mahon, K.A.; Jamrich, M. The Rx homeobox gene is essential for vertebrate eye development. Nature 1997, 387, 603–607. [Google Scholar] [CrossRef]
- Voronina, V.A.; Kozhemyakina, E.A.; O’Kernick, C.M.; Kahn, N.D.; Wenger, S.L.; Linberg, J.V.; Schneider, A.S.; Mathers, P.H. Mutations in the human RAX homeobox gene in a patient with anophthalmia and sclerocornea. Hum. Mol. Genet. 2004, 13, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lagutin, O.; Swindell, E.; Jamrich, M.; Oliver, G. Neuroretina specification in mouse embryos requires Six3-mediated suppression of Wnt8b in the anterior neural plate. J. Clin. Investig. 2010, 120, 3568–3577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagutin, O.V.; Zhu, C.C.; Kobayashi, D.; Topczewski, J.; Shimamura, K.; Puelles, L.; Russell, H.R.C.; McKinnon, P.J.; Solnica-Krezel, L.; Oliver, G. Six3 repression of Wnt signaling in the anterior neuroectoderm is essential for vertebrate forebrain development. Genes Dev. 2003, 17, 368–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loosli, F.; Winkler, S.; Wittbrodt, J. Six3 overexpression initiates the formation of ectopic retina. Genes Dev. 1999, 13, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, S. Chapter Three–Eye Morphogenesis and Patterning of the Optic Vesicle. In Current Topics in Developmental Biology; Cagan, R.L., Reh, T.A., Eds.; Academic Press: Cambridge, MA, USA, 2010; Volume 93, pp. 61–84. ISBN 0070-2153. [Google Scholar]
- Bharti, K.; Nguyen, M.-T.T.; Skuntz, S.; Bertuzzi, S.; Arnheiter, H. The other pigment cell: Specification and development of the pigmented epithelium of the vertebrate eye. Pigment Cell Res. 2006, 19, 380–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amram, B.; Cohen-Tayar, Y.; David, A.; Ashery-Padan, R. The retinal pigmented epithelium–from basic developmental biology research to translational approaches. Int. J. Dev. Biol. 2017, 61, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Tayar, Y.; Cohen, H.; Mitiagin, Y.; Abravanel, Z.; Levy, C.; Idelson, M.; Reubinoff, B.; Itzkovitz, S.; Raviv, S.; Kaestner, K.H.; et al. Pax6 regulation of Sox9 in the mouse retinal pigmented epithelium controls its timely differentiation and choroid vasculature development. Development 2018, 145, dev163691. [Google Scholar] [CrossRef] [Green Version]
- Bharti, K.; Liu, W.; Csermely, T.; Bertuzzi, S.; Arnheiter, H. Alternative promoter use in eye development: The complex role and regulation of the transcription factor MITF. Development 2008, 135, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Cavodeassi, F.; Bovolenta, P. New functions for old genes: Pax6 and Mitf in eye pigment biogenesis. Pigment Cell Melanoma Res. 2014, 27, 1005–1007. [Google Scholar] [CrossRef]
- Raviv, S.; Bharti, K.; Rencus-Lazar, S.; Cohen-Tayar, Y.; Schyr, R.; Evantal, N.; Meshorer, E.; Zilberberg, A.; Idelson, M.; Reubinoff, B.; et al. PAX6 Regulates Melanogenesis in the Retinal Pigmented Epithelium through Feed-Forward Regulatory Interactions with MITF. PLoS Genet. 2014, 10, e1004360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramón Martínez-Morales, J.; Rodrigo, I.; Bovolenta, P. Eye development: A view from the retina pigmented epithelium. BioEssays 2004, 26, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Nikitina, T.; Zhao, J.; Fleury, T.J.; Bhattacharyya, R.; Bouhassira, E.E.; Stein, A.; Woodcock, C.L.; Skoultchi, A.I. Histone H1 Depletion in Mammals Alters Global Chromatin Structure but Causes Specific Changes in Gene Regulation. Cell 2005, 123, 1199–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fyodorov, D.V.; Zhou, B.-R.; Skoultchi, A.I.; Bai, Y. Emerging roles of linker histones in regulating chromatin structure and function. Nat. Rev. Mol. Cell Biol. 2018, 19, 192–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luz-Madrigal, A.; Grajales-Esquivel, E.; Tangeman, J.; Kosse, S.; Liu, L.; Wang, K.; Fausey, A.; Liang, C.; Tsonis, P.A.; Del Rio-Tsonis, K. DNA demethylation is a driver for chick retina regeneration. Epigenetics 2020, 15, 998–1019. [Google Scholar] [CrossRef] [Green Version]
- Raeisossadati, R.; Ferrari, M.F.R.; Kihara, A.H.; AlDiri, I.; Gross, J.M. Epigenetic regulation of retinal development. Epigenetics Chromatin 2021, 14, 11. [Google Scholar] [CrossRef]
- Dvoriantchikova, G.; Seemungal, R.J.; Ivanov, D. The epigenetic basis for the impaired ability of adult murine retinal pigment epithelium cells to regenerate retinal tissue. Sci. Rep. 2019, 9, 3860. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zibetti, C.; Shang, P.; Sripathi, S.R.; Zhang, P.; Cano, M.; Hoang, T.; Xia, S.; Ji, H.; Merbs, S.L.; et al. ATAC-Seq analysis reveals a widespread decrease of chromatin accessibility in age-related macular degeneration. Nat. Commun. 2018, 9, 1364. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Jeong, Y.; Kwon, K.; Ismail, T.; Lee, H.-K.; Kim, C.; Park, J.-W.; Kwon, O.-S.; Kang, B.-S.; Lee, D.-S.; et al. Physiological effects of KDM5C on neural crest migration and eye formation during vertebrate development. Epigenetics Chromatin 2018, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Karg, M.; Lu, Y.; Hoffmann, E.; Philipose, H.; Sinclair, D.; Ksander, B.; Saint-Geniez, M. In vivo epigenetic reprogramming reverses the age-induced morphological decline of retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2021, 62, 3283. [Google Scholar]
- Rai, K.; Chidester, S.; Zavala, C.V.; Manos, E.J.; James, S.R.; Karpf, A.R.; Jones, D.A.; Cairns, B.R. Dnmt2 functions in the cytoplasm to promote liver, brain, and retina development in zebrafish. Genes Dev. 2007, 21, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Nasonkin, I.O.; Merbs, S.L.; Lazo, K.; Oliver, V.F.; Brooks, M.; Patel, K.; Enke, R.A.; Nellissery, J.; Jamrich, M.; Le, Y.Z.; et al. Conditional knockdown of DNA methyltransferase 1 reveals a key role of retinal pigment epithelium integrity in photoreceptor outer segment morphogenesis. Development 2013, 140, 1330–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adil, A.; Kumar, V.; Jan, A.T.; Asger, M. Single-Cell Transcriptomics: Current Methods and Challenges in Data Acquisition and Analysis. Front. Neurosci. 2021, 15, 591122. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, X.; Hu, B.; Mao, Y.; Chen, Y.; Yan, L.; Yong, J.; Dong, J.; Wei, Y.; Wang, W.; et al. Dissecting the transcriptome landscape of the human fetal neural retina and retinal pigment epithelium by single-cell RNA-seq analysis. PLoS Biol. 2019, 17, e3000365. [Google Scholar] [CrossRef] [Green Version]
- Booij, J.C.; van Soest, S.; Swagemakers, S.M.A.; Essing, A.H.W.; Verkerk, A.J.M.H.; van der Spek, P.J.; Gorgels, T.G.M.F.; Bergen, A.A.B. Functional annotation of the human retinal pigment epithelium transcriptome. BMC Genom. 2009, 10, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lidgerwood, G.E.; Senabouth, A.; Smith-Anttila, C.J.A.; Gnanasambandapillai, V.; Kaczorowski, D.C.; Amann-Zalcenstein, D.; Fletcher, E.L.; Naik, S.H.; Hewitt, A.W.; Powell, J.E.; et al. Transcriptomic Profiling of Human Pluripotent Stem Cell-derived Retinal Pigment Epithelium over Time. Genom. Proteom. Bioinform. 2021, 19, 223–242. [Google Scholar] [CrossRef]
- Voigt, A.P.; Mulfaul, K.; Mullin, N.K.; Flamme-Wiese, M.J.; Giacalone, J.C.; Stone, E.M.; Tucker, B.A.; Scheetz, T.E.; Mullins, R.F. Single-cell transcriptomics of the human retinal pigment epithelium and choroid in health and macular degeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 24100–24107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitmore, S.S.; Wagner, A.H.; DeLuca, A.P.; Drack, A.V.; Stone, E.M.; Tucker, B.A.; Zeng, S.; Braun, T.A.; Mullins, R.F.; Scheetz, T.E. Transcriptomic analysis across nasal, temporal, and macular regions of human neural retina and RPE/choroid by RNA-Seq. Exp. Eye Res. 2014, 129, 93–106. [Google Scholar] [CrossRef] [Green Version]
- West, K.A.; Yan, L.; Shadrach, K.; Sun, J.; Hasan, A.; Miyagi, M.; Crabb, J.S.; Hollyfield, J.G.; Marmorstein, A.D.; Crabb, J.W. Protein Database, Human Retinal Pigment Epithelium. Mol. Cell. Proteom. 2003, 2, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Hongisto, H.; Jylhä, A.; Nättinen, J.; Rieck, J.; Ilmarinen, T.; Veréb, Z.; Aapola, U.; Beuerman, R.; Petrovski, G.; Uusitalo, H.; et al. Comparative proteomic analysis of human embryonic stem cell-derived and primary human retinal pigment epithelium. Sci. Rep. 2017, 7, 6016. [Google Scholar] [CrossRef]
- Pelkonen, L.; Sato, K.; Reinisalo, M.; Kidron, H.; Tachikawa, M.; Watanabe, M.; Uchida, Y.; Urtti, A.; Terasaki, T. LC–MS/MS Based Quantitation of ABC and SLC Transporter Proteins in Plasma Membranes of Cultured Primary Human Retinal Pigment Epithelium Cells and Immortalized ARPE19 Cell Line. Mol. Pharm. 2017, 14, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, B.A.; Fliesler, S.J. Reassessing the suitability of ARPE-19 cells as a valid model of native RPE biology. Exp. Eye Res. 2022, 219, 109046. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Geathers, J.S.; Grillo, S.L.; Weber, S.R.; Wang, W.; Zhao, Y.; Sundstrom, J.M. Role of Epithelial-Mesenchymal Transition in Retinal Pigment Epithelium Dysfunction. Front. Cell Dev. Biol. 2020, 8, 501. [Google Scholar] [CrossRef] [PubMed]
- Alge, C.S.; Suppmann, S.; Priglinger, S.G.; Neubauer, A.S.; May, C.A.; Hauck, S.; Welge-Lussen, U.; Ueffing, M.; Kampik, A. Comparative Proteome Analysis of Native Differentiated and Cultured Dedifferentiated Human RPE Cells. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3629–3641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, E.; Lu, X.; Flippin, J.; Devaney, J.M.; Halligan, B.; Hoffman, E.; Csaky, K.; Hathout, Y. Secreted Proteome Profiling in Human RPE Cell Cultures Derived from Donors with Age Related Macular Degeneration and Age Matched Healthy Donors. J. Proteome Res. 2006, 5, 2599–2610. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-4. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-4 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Wang, H.-C.; Greene, W.A.; Kaini, R.R.; Shen-Gunther, J.; Chen, H.-I.H.; Car, H.; Wang, Y. Profiling the microRNA Expression in Human iPS and iPS-derived Retinal Pigment Epithelium. Cancer Inform. 2014, 13, CIN-S14074. [Google Scholar] [CrossRef]
- Hu, G.; Huang, K.; Yu, J.; Gopalakrishna-Pillai, S.; Kong, J.; Xu, H.; Liu, Z.; Zhang, K.; Xu, J.; Luo, Y.; et al. Identification of miRNA Signatures during the Differentiation of hESCs into Retinal Pigment Epithelial Cells. PLoS ONE 2012, 7, e37224. [Google Scholar] [CrossRef]
- Davis, N.; Mor, E.; Ashery-Padan, R. Roles for Dicer1 in the patterning and differentiation of the optic cup neuroepithelium. Development 2011, 138, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Sundermeier, T.R.; Sakami, S.; Sahu, B.; Howell, S.J.; Gao, S.; Dong, Z.; Golczak, M.; Maeda, A.; Palczewski, K. MicroRNA-processing Enzymes Are Essential for Survival and Function of Mature Retinal Pigmented Epithelial Cells in Mice. J. Biol. Chem. 2017, 292, 3366–3378. [Google Scholar] [CrossRef] [Green Version]
- Iacovelli, J.; Zhao, C.; Wolkow, N.; Veldman, P.; Gollomp, K.; Ojha, P.; Lukinova, N.; King, A.; Feiner, L.; Esumi, N.; et al. Generation of Cre Transgenic Mice with Postnatal RPE-Specific Ocular Expression. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1378–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohana, R.; Weiman-Kelman, B.; Raviv, S.; Tamm, E.R.; Pasmanik-Chor, M.; Rinon, A.; Netanely, D.; Shamir, R.; Solomon, A.S.; Ashery-Padan, R. MicroRNAs are essential for differentiation of the retinal pigmented epithelium and maturation of adjacent photoreceptors. Development 2015, 142, 2487–2498. [Google Scholar] [CrossRef] [PubMed]
- Du, S.W.; Palczewski, K. MicroRNA regulation of critical retinal pigment epithelial functions. Trends Neurosci. 2022, 45, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Adijanto, J.; Castorino, J.J.; Wang, Z.-X.; Maminishkis, A.; Grunwald, G.B.; Philp, N.J. Microphthalmia-associated Transcription Factor (MITF) Promotes Differentiation of Human Retinal Pigment Epithelium (RPE) by Regulating microRNAs-204/211 Expression. J. Biol. Chem. 2012, 287, 20491–20503. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Qin, B.; Liu, G.; Sun, X.; Shi, H.; Ding, S.; Liu, Y.; Zhu, M.; Chen, X.; Zhao, C. MicroRNA-184 promotes differentiation of the retinal pigment epithelium by targeting the AKT2/mTOR signaling pathway. Oncotarget 2016, 7, 52340–52353. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.W.; Kim, J.-J.; Seo, M.-S.; Park, S.-B.; Kang, T.-W.; Lee, J.Y.; Lee, B.-C.; Kang, I.; Shin, T.-H.; Kim, H.-S.; et al. miR-410 Inhibition Induces RPE Differentiation of Amniotic Epithelial Stem Cells via Overexpression of OTX2 and RPE65. Stem Cell Rev. Rep. 2015, 11, 376–386. [Google Scholar] [CrossRef]
- Harhaj, N.S.; Antonetti, D.A. Regulation of tight junctions and loss of barrier function in pathophysiology. Int. J. Biochem. Cell Biol. 2004, 36, 1206–1237. [Google Scholar] [CrossRef]
- Wang, F.E.; Zhang, C.; Maminishkis, A.; Dong, L.; Zhi, C.; Li, R.; Zhao, J.; Majerciak, V.; Gaur, A.B.; Chen, S.; et al. MicroRNA-204/211 alters epithelial physiology. FASEB J. 2010, 24, 1552–1571. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Kaneko, H.; Hwang, S.-J.; Ye, F.; Higuchi, A.; Tsunekawa, T.; Matsuura, T.; Iwase, T.; Asami, T.; Ito, Y.; et al. Increased Ocular Levels of MicroRNA-148a in Cases of Retinal Detachment Promote Epithelial–Mesenchymal Transition. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2699–2705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Miyagishima, K.J.; Dong, L.; Rising, A.; Nimmagadda, M.; Liang, G.; Sharma, R.; Dejene, R.; Wang, Y.; Abu-Asab, M.; et al. Regulation of phagolysosomal activity by miR-204 critically influences structure and function of retinal pigment epithelium/retina. Hum. Mol. Genet. 2019, 28, 3355–3368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murad, N.; Kokkinaki, M.; Gunawardena, N.; Gunawan, M.S.; Hathout, Y.; Janczura, K.J.; Theos, A.C.; Golestaneh, N. miR-184 regulates ezrin, LAMP-1 expression, affects phagocytosis in human retinal pigment epithelium and is downregulated in age-related macular degeneration. FEBS J. 2014, 281, 5251–5264. [Google Scholar] [CrossRef]
- Choi, S.W.; Kim, J.-J.; Seo, M.-S.; Park, S.-B.; Shin, T.-H.; Shin, J.-H.; Seo, Y.; Kim, H.-S.; Kang, K.-S. Inhibition by miR-410 facilitates direct retinal pigment epithelium differentiation of umbilical cord blood-derived mesenchymal stem cells. J. Vet. Sci. 2017, 18, 59–65. [Google Scholar] [CrossRef]
- Cui, L.; Lyu, Y.; Jin, X.; Wang, Y.; Li, X.; Wang, J.; Zhang, J.; Deng, Z.; Yang, N.; Zheng, Z.; et al. miR-194 suppresses epithelial-mesenchymal transition of retinal pigment epithelial cells by directly targeting ZEB1. Ann. Transl. Med. 2019, 7, 751. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Zheng, L.; Wang, M.; Lu, Y.; Li, Z.; Lian, C.; Mao, S.; Hou, X.; Li, S.; et al. miR-25 Mediates Retinal Degeneration Via Inhibiting ITGAV and PEDF in Rat. Curr. Mol. Med. 2017, 17, 359–374. [Google Scholar] [CrossRef]
- Zhang, K.; Ding, S. Stem Cells and Eye Development. N. Engl. J. Med. 2011, 365, 370–372. [Google Scholar] [CrossRef] [Green Version]
- Sonoda, S.; Spee, C.; Barron, E.; Ryan, S.J.; Kannan, R.; Hinton, D.R. A protocol for the culture and differentiation of highly polarized human retinal pigment epithelial cells. Nat. Protoc. 2009, 4, 662–673. [Google Scholar] [CrossRef] [Green Version]
- Limnios, I.J.; Chau, Y.-Q.; Skabo, S.J.; Surrao, D.C.; O’Neill, H.C. Efficient differentiation of human embryonic stem cells to retinal pigment epithelium under defined conditions. Stem Cell Res. Ther. 2021, 12, 248. [Google Scholar] [CrossRef]
- Zahabi, A.; Shahbazi, E.; Ahmadieh, H.; Hassani, S.-N.; Totonchi, M.; Taei, A.; Masoudi, N.; Ebrahimi, M.; Aghdami, N.; Seifinejad, A.; et al. A New Efficient Protocol for Directed Differentiation of Retinal Pigmented Epithelial Cells from Normal and Retinal Disease Induced Pluripotent Stem Cells. Stem Cells Dev. 2011, 21, 2262–2272. [Google Scholar] [CrossRef] [PubMed]
- Greber, B.; Coulon, P.; Zhang, M.; Moritz, S.; Frank, S.; Müller-Molina, A.J.; Araúzo-Bravo, M.J.; Han, D.W.; Pape, H.-C.; Schöler, H.R. FGF signalling inhibits neural induction in human embryonic stem cells. EMBO J. 2011, 30, 4874–4884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Zhou, J.; Meng, X. Primary cilia in retinal pigment epithelium development and diseases. J. Cell. Mol. Med. 2021, 25, 9084–9088. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, S.; Levine, E.M.; Reh, T.A. Extraocular mesenchyme patterns the optic vesicle during early eye development in the embryonic chick. Development 2000, 127, 4599–4609. [Google Scholar] [CrossRef] [PubMed]
- Buse, E.; de Groot, H. Generation of developmental patterns in the neuroepithelium of the developing mammalian eye: The pigment epithelium of the eye. Neurosci. Lett. 1991, 126, 63–66. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, S.; Lytvynchuk, L.; Ardan, T.; Studenovska, H.; Faura, G.; Eide, L.; Znaor, L.; Erceg, S.; Stieger, K.; Motlik, J.; et al. Retinal Pigment Epithelium Cell Development: Extrapolating Basic Biology to Stem Cell Research. Biomedicines 2023, 11, 310. https://doi.org/10.3390/biomedicines11020310
Gupta S, Lytvynchuk L, Ardan T, Studenovska H, Faura G, Eide L, Znaor L, Erceg S, Stieger K, Motlik J, et al. Retinal Pigment Epithelium Cell Development: Extrapolating Basic Biology to Stem Cell Research. Biomedicines. 2023; 11(2):310. https://doi.org/10.3390/biomedicines11020310
Chicago/Turabian StyleGupta, Santosh, Lyubomyr Lytvynchuk, Taras Ardan, Hana Studenovska, Georgina Faura, Lars Eide, Ljubo Znaor, Slaven Erceg, Knut Stieger, Jan Motlik, and et al. 2023. "Retinal Pigment Epithelium Cell Development: Extrapolating Basic Biology to Stem Cell Research" Biomedicines 11, no. 2: 310. https://doi.org/10.3390/biomedicines11020310