
Association of Epicardial Adipose Tissue Adipocytes Hypertrophy with Biomarkers of Low-Grade Inflammation and Extracellular Matrix Remodeling in Patients with Coronary Artery Disease
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Echocardiography
2.3. Adipose Tissue Sampling and Processing
2.4. Blood Sampling and Processing
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Biochemical Assays
2.7. Statistical Analysis
3. Results
3.1. Morphometric and Biochemical Parameters
3.2. Biomarkers of Low-Grade Inflammation
3.3. Components of Extracellular Matrix Remodeling
3.4. Associations between EAT Adipocytes Size, Anthropometric, Biochemical Parameters and Low-Grade Inflammation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carbone, F.; Lattanzio, M.S.; Minetti, S.; Ansaldo, A.M.; Ferrara, D.; Molina-Molina, E.; Belfiore, A.; Elia, E.; Pugliese, S.; Palmieri, V.O.; et al. Circulating CRP Levels Are Associated with Epicardial and Visceral Fat Depots in Women with Metabolic Syndrome Criteria. Int. J. Mol. Sci. 2019, 20, 5981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, J.; Luzardo, E.; Mejías, J.C.; Rojas, J.; Ferreira, A.; Rivas-Ríos, J.R.; Bermúdez, V. Epicardial Fat: Physiological, Pathological, and Therapeutic Implications. Cardiol. Res. Pract. 2016, 2016, 1291537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacobellis, G. Epicardial adipose tissue in contemporary cardiology. Nat. Rev. Cardiol. 2022, 19, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Varga, I. Microenvironment of Immune Cells within the Visceral Adipose Tissue Sensu Lato vs. Epicardial Adipose Tissue: What Do We Know? Inflammation 2018, 41, 1142–1156. [Google Scholar] [CrossRef]
- Murdolo, G.; Smith, U. The Dysregulated Adipose Tissue: A Connecting Link between Insulin Resistance, Type 2 Diabetes Mellitus and Atherosclerosis. Nutr. Metab. Cardiovasc. Dis. 2006, 16, S35–S38. [Google Scholar] [CrossRef]
- Vianello, E.; Dozio, E.; Arnaboldi, F.; Marazzi, M.G.; Martinelli, C.; Lamont, J.; Tacchini, L.; Sigrüner, A.; Schmitz, G.; Corsi Romanelli, M.M. Epicardial Adipocyte Hypertrophy: Association with M1-Polarization and Toll-Like Receptor Pathways in Coronary Artery Disease Patients. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 246–253. [Google Scholar] [CrossRef]
- Wadey, R.M.; Connolly, K.D.; Mathew, D.; Walters, G.; Rees, D.A.; James, P.E. Inflammatory Adipocyte-Derived Extracellular Vesicles Promote Leukocyte Attachment to Vascular Endothelial Cells. Atherosclerosis 2019, 283, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Unamuno, X.; Gómez-Ambrosi, J.; Rodríguez, A.; Becerril, S.; Frühbeck, G.; Catalán, V. Adipokine Dysregulation and Adipose Tissue Inflammation in Human Obesity. Eur. J. Clin. Investig. 2018, 48, e12997. [Google Scholar] [CrossRef] [Green Version]
- Dutour, A.; Achard, V.; Sell, H.; Naour, N.; Collart, F.; Gaborit, B.; Silaghi, A.; Eckel, J.; Alessi, M.C.; Henegar, C.; et al. Secretory Type II Phospholipase A2 is Produced and Secreted by Epicardial Adipose Tissue and Overexpressed in Patients with Coronary Artery Disease. J. Clin. Endocrinol. Metab. 2010, 95, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Mráz, M.; Cinkajzlová, A.; Kloučková, J.; Lacinová, Z.; Kratochvílová, H.; Lipš, M.; Pořízka, M.; Kopecký, P.; Lindner, J.; Kotulák, T.; et al. Dendritic Cells in Subcutaneous and Epicardial Adipose Tissue of Subjects with Type 2 Diabetes, Obesity, and Coronary Artery Disease. Mediat. Inflamm. 2019, 2019, 5481725. [Google Scholar] [CrossRef]
- Kitagawa, T.; Yamamoto, H.; Hattori, T.; Sentani, K.; Takahashi, S.; Senoo, A.; Kubo, Y.; Yasui, W.; Sueda, T.; Kihara, Y. Tumor Necrosis Factor-α Gene Expression in Epicardial Adipose Tissue is Related to Coronary Atherosclerosis Assessed by Computed Tomography. J. Atheroscler. Thromb. 2018, 25, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Leiguez, E.; Motta, P.; Maia Marques, R.; Lomonte, B.; Sampaio, S.V.; Teixeira, C. A Representative GIIA Phospholipase A2 Activates Preadipocytes to Produce Inflammatory Mediators Implicated in Obesity Development. Biomolecules 2020, 10, 1593. [Google Scholar] [CrossRef] [PubMed]
- Tandon, P.; Wafer, R.; Minchin, J.E. Adipose Morphology and Metabolic Disease. J. Exp. Biol. 2018, 221, jeb164970. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, T.; Craig, C.; Liu, L.F.; Perelman, D.; Allister, C.; Spielman, D.; Cushman, S.W. Adipose Cell Size and Regional Fat Deposition as Predictors of Metabolic Response to Overfeeding in Insulin-Resistant and Insulin-Sensitive Humans. Diabetes 2016, 65, 1245–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenkula, K.G.; Erlanson-Albertsson, C. Adipose Cell Size: Importance in Health and Disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R284–R295. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, T.; Sherman, A.; Tsao, P.; Gonzalez, O.; Yee, G.; Lamendola, C.; Reaven, G.M.; Cushman, S.W. Enhanced Proportion of Small Adipose Cells in Insulin-Resistant vs. Insulin-Sensitive Obese Individuals Implicates Impaired Adipogenesis. Diabetologia 2007, 50, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- Khan, T.; Muise, E.S.; Iyengar, P.; Wang, Z.V.; Chandalia, M.; Abate, N.; Zhang, B.B.; Bonaldo, P.; Chua, S.; Scherer, P.E. Metabolic Dysregulation and Adipose Tissue Fibrosis: Role of Collagen VI. Mol. Cell. Biol. 2009, 29, 1575–1591. [Google Scholar] [CrossRef] [Green Version]
- de Sousa Neto, I.V.; Durigan, J.L.Q.; da Silva, A.S.R.; de Cássia Marqueti, R. Adipose Tissue Extracellular Matrix Remodeling in Response to Dietary Patterns and Exercise: Molecular Landscape, Mechanistic Insights, and Therapeutic Approaches. Biology 2022, 11, 765. [Google Scholar] [CrossRef]
- Gensini, G.G. A More Meaningful Scoring System for Determining the Severity of Coronary Heart Disease. Am. J. Cardiol. 1983, 51, 606. [Google Scholar] [CrossRef]
- Jacobellis, G.; Assael, F.; Ribaudo, M.C. Epicardial Fat from Echocardiography: A New Method for Visceral Adipose Tissue Prediction. Obes. Res. 2003, 11, 304–310. [Google Scholar] [CrossRef]
- Naryzhnaya, N.V.; Koshelskaya, O.A.; Kologrivova, I.V.; Suslova, T.E.; Kharitonova, O.A.; Andreev, S.L.; Gorbunov, A.S.; Kurbatov, B.K.; Boshchenko, A.A. Production of Reactive Oxygen Species by Epicardial Adipocytes Is Associated with an Increase in Postprandial Glycemia, Postprandial Insulin, and a Decrease in Serum Adiponectin in Patients with Severe Coronary Atherosclerosis. Biomedicines 2022, 10, 2054. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, S.; Juge-Aubry, C.E.; Meier, C.A. Explant Cultures of White Adipose Tissue. Adipose Tissue Protocols. Methods in Molecular Biology. Methods Mol. Biol. 2008, 456, 195–199. [Google Scholar] [CrossRef]
- Suga, H.; Matsumoto, D.; Inoue, K.; Shigeura, T.; Eto, H.; Aoi, N.; Kato, H.; Abe, H.; Yoshimura, K. Numerical Measurement of Viable and Nonviable Adipocytes and Other Cellular Components in Aspirated Fat Tissue. Plast. Reconstr. Surg. 2008, 122, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tok, D.; Kadife, I.; Turak, O.; Ozcan, F.; Başar, N.; Cağlı, K.; Aras, D.; Topaloğlu, S.A.S. Increased Epicardial Fat Thickness is Associated with Low Grade Systemic Inflammation in Metabolic Syndrome. Turk. Kardiyol. Dern. Ars. 2012, 40, 690–695. [Google Scholar] [CrossRef] [Green Version]
- Malavazos, A.E.; Ermetici, F.; Coman, C.; Corsi, M.M.; Morricone, L.; Ambrosi, B. Influence of Epicardial Adipose Tissue and Adipocytokine Levels on Cardiac Abnormalities in Visceral Obesity. Int. J. Cardiol. 2007, 121, 132–134. [Google Scholar] [CrossRef]
- Aitken-Buck, H.M.; Moharram, M.; Babakr, A.A.; Reijers, R.; Van Hout, I.; Fomison-Nurse, I.C.; Sugunesegran, R.; Bhagwat, K.; Davis, P.J.; Bunton, R.W.; et al. Relationship between Epicardial Adipose Tissue Thickness and Epicardial Adipocyte Size with Increasing Body Mass Index. Adipocyte 2019, 8, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Guo, F.; Zhou, L.; Stahl, R.; Grams, J. The Cell Size and Distribution of Adipocytes from Subcutaneous and Visceral Fat is Associated with Type 2 Diabetes Mellitus in Humans. Adipocyte 2015, 4, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Alligier, M.; Meugnier, E.; Debard, C.; Lambert-Porcheron, S.; Chanseaume, E.; Sothier, M.; Loizon, E.; Hssain, A.A.; Brozek, J.; Scoazec, J.Y.; et al. Subcutaneous Adipose Tissue Remodeling during the Initial Phase of Weight Gain Induced by Overfeeding in Humans. J. Clin. Endocrinol. Metab. 2012, 97, E183–E192. [Google Scholar] [CrossRef] [Green Version]
- Hammarstedt, A.; Gogg, S.; Hedjazifar, S.; Nerstedt, A.; Smith, U. Impaired Adipogenesis and Dysfunctional Adipose Tissue in Human Hypertrophic Obesity. Physiol. Rev. 2018, 98, 1911–1941. [Google Scholar] [CrossRef] [Green Version]
- Zöller, N.; Schreiner, S.; Petry, L.; Hoffmann, S.; Steinhorst, K.; Kleemann, J.; Jäger, M.; Kaufmann, R.; Meissner, M.; Kippenberger, S. Collagen I Promotes Adipocytogenesis in Adipose-Derived Stem Cells in Vitro. Cells 2019, 8, 302. [Google Scholar] [CrossRef]
- Gao, Y.; Ma, K.; Kang, Y.; Liu, W.; Liu, X.; Long, X.; Hayashi, T.; Hattori, S.; Mizuno, K.; Fujisaki, H.; et al. Type I Collagen Reduces Lipid Accumulation During Adipogenesis of Preadipocytes 3T3-L1 via the YAP-mTOR-autophagy Axis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159181. [Google Scholar] [CrossRef] [PubMed]
- Datta, R.; Podolsky, M.J.; Atabai, K. Fat Fibrosis: Friend or Foe? JCI Insight 2018, 3, e122289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.K.; Anderson, P.H.; Morris, H.A.; Clifton, P.M. Visceral Fat is a Negative Determinant of Bone Health in Obese Postmenopausal Women. Int. J. Environ. Res. Public Health 2020, 17, 3996. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Kim, N.R.; Kang, J.W.; Eun, J.S.; Kang, Y.M. Radial BMD and Serum CTX-I Can Predict the Progression of Carotid Plaque in Rheumatoid Arthritis: A 3-year Prospective Cohort Study. Arthritis Res. Ther. 2021, 23, 258. [Google Scholar] [CrossRef]
- Small, A.; Kiss, D.; Giri, J.; Anwaruddin, S.; Siddiqi, H.; Guerraty, M.; Chirinos, J.A.; Ferrari, G.; Rader, D.J. Biomarkers of Calcific Aortic Valve Disease. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine Regulation of Energy Metabolism by the Skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.N.; Wei, P.; Yu, J. Higher Concentration of Serum C-Terminal Cross-Linking Telopeptide of Type I Collagen is Positively Related with Inflammatory Factors in Postmenopausal Women with H-Type Hypertension and Osteoporosis. Orthop. Surg. 2019, 11, 1135–1141. [Google Scholar] [CrossRef]
- Skjøt-Arkil, H.; Barascuk, N.; Larsen, L.; Dziegiel, M.; Henriksen, K.; Karsdal, M.A. Tumor Necrosis Factor-A and Receptor Activator of Nuclear Factor-κB Ligand Augment Human Macrophage Foam-Cell Destruction of Extracellular Matrix through Protease-Mediated Processes. Assay Drug Dev. Technol. 2012, 10, 69–77. [Google Scholar] [CrossRef]
- Djafari, F.; Eslahi, M.; Zandi, N.; Pazoki, B.; Reza Amini, M.; Shab-Bidar, S. The Inverse Association of Body Adiposity Index and Bone Health in the Older Adults: A Report from A Developing Country. Int. J. Clin. Pract. 2021, 75, e14718. [Google Scholar] [CrossRef]
- Iacobellis, G.; Barbaro, G. The Double Role of Epicardial Adipose Tissue as Pro- and Anti-inflammatory Organ. Horm. Metab. Res. 2008, 40, 442–445. [Google Scholar] [CrossRef]
- Conte, M.; Petraglia, L.; Poggio, P.; Valerio, V.; Cabaro, S.; Campana, P.; Comentale, G.; Attena, E.; Russo, V.; Pilato, E.; et al. Inflammation and Cardiovascular Diseases in the Elderly: The Role of Epicardial Adipose Tissue. Front. Med. 2022, 9, 844266. [Google Scholar] [CrossRef] [PubMed]
- Granér, M.; Seppälä-Lindroos, A.; Rissanen, A.; Hakkarainen, A.; Lundbom, N.; Kaprio, J.; Nieminen, M.S.; Pietiläinen, K.H. Epicardial Fat, Cardiac Dimensions, and Low-Grade Inflammation in Young Adult Monozygotic Twins Discordant for Obesity. Am. J. Cardiol. 2012, 109, 1295–1302. [Google Scholar] [CrossRef] [PubMed]
- Dixon, W.G.; Watson, K.D.; Lunt, M.; Hyrich, K.L.; Silman, A.J.; Symmons, D.P. Reduction in the Incidence of Myocardial Infarction in Patients with Rheumatoid Arthritis Who Respond to Anti-Tumor Necrosis Factor Alpha Therapy: Results from The British Society for Rheumatology Biologics Register. Arthritis Rheum. 2007, 56, 2905–2912. [Google Scholar] [CrossRef]
- Cawthorn, W.P.; Sethi, J.K. TNF-alpha and Adipocyte Biology. FEBS Lett. 2008, 582, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Aladhami, A.K.; Unger, C.A.; Ennis, S.L.; Altomare, D.; Ji, H.; Hope, M.C., 3rd; Velázquez, K.T.; Enos, R.T. Macrophage Tumor Necrosis Factor-Alpha Deletion Does Not Protect Against Obesity-Associated Metabolic Dysfunction. FASEB J. 2021, 35, e21665. [Google Scholar] [CrossRef] [PubMed]
- Cawthorn, W.P.; Heyd, F.; Hegyi, K.; Sethi, J.K. Tumour Necrosis Factor-Alpha Inhibits Adipogenesis via a Beta-Catenin/TCF4(TCF7L2)-Dependent Pathway. Cell Death Differ. 2007, 14, 1361–1373. [Google Scholar] [CrossRef] [PubMed]
- Crowl, R.M.; Stoller, T.J.; Conroy, R.R.; Stoner, C.R. Induction of Phospholipase A2 Gene Expression in Human Hepatoma Cells by Mediators of The Acute Phase Response. J. Biol. Chem. 1991, 266, 2647–2651. [Google Scholar] [CrossRef]
- Sato, H.; Taketomi, Y.; Ushida, A.; Isogai, Y.; Kojima, T.; Hirabayashi, T.; Miki, Y.; Yamamoto, K.; Nishito, Y.; Kobayashi, T.; et al. The Adipocyte-Inducible Secreted Phospholipases PLA2G5 and PLA2G2E Play Distinct Roles in Obesity. Cell Metab. 2014, 20, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Lasbleiz, A.; Gaborit, B.; Soghomonian, A.; Bartoli, A.; Ancel, P.; Jacquier, A.; Dutour, A. COVID-19 and Obesity: Role of Ectopic Visceral and Epicardial Adipose Tissues in Myocardial Injury. Front. Endocrinol. 2021, 12, 726967. [Google Scholar] [CrossRef]
- Sun, K.; Tordjman, J.; Clément, K.; Scherer, P.E. Fibrosis and Adipose Tissue Dysfunction. Cell Metab. 2013, 18, 470–477. [Google Scholar] [CrossRef]
- Ye, R.Z.; Richard, G.; Gévry, N.; Tchernof, A.; Carpentier, A.C. Fat Cell Size: Measurement Methods, Pathophysiological Origins, and Relationships with Metabolic Dysregulations. Endocr. Rev. 2022, 12, 35–60. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H.; Moors, C.C.; van der Zijl, N.J.; Venteclef, N.; Alili, R.; Jocken, J.W.; Essers, Y.; Cleutjens, J.P.; Clément, K.; Diamant, M.; et al. Valsartan Improves Adipose Tissue Function in Humans with Impaired Glucose Metabolism: A Randomized Placebo-Controlled Double-Blind Trial. PLoS ONE 2012, 7, e39930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.; Si, Y.; Xing, E.; Feng, Z.; Ding, Z.; Liu, Y.; Wei, C.; Tian, Y.; Zhang, Y.; Liu, J.; et al. Human Epicardial Adipose Tissue Inflammation Correlates with Coronary Artery Disease. Cytokine 2023, 162, 156119. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.C.; Wang, Z.Y.; Xu, Q.; Li, R.B.; Zhang, G.G.; Shi, R.Z. Exploring the Role of Epicardial Adipose Tissue in Coronary Artery Disease from the Difference of Gene Expression. Front. Physiol. 2021, 12, 605811. [Google Scholar] [CrossRef] [PubMed]
- Naryzhnaya, N.V.; Koshelskaya, O.A.; Kologrivova, I.V.; Kharitonova, O.A.; Evtushenko, V.V.; Boshchenko, A.A. Hypertrophy and Insulin Resistance of Epicardial Adipose Tissue Ad-ipocytes: Association with the Coronary Artery Disease Severity. Biomedicines 2021, 9, 64. [Google Scholar] [CrossRef]
- Doukbi, E.; Soghomonian, A.; Sengenès, C.; Ahmed, S.; Ancel, P.; Dutour, A.; Gaborit, B. Browning Epicardial Adipose Tissue: Friend or Foe? Cells 2022, 11, 991. [Google Scholar] [CrossRef]
- Bielczyk-Maczynska, E. White Adipocyte Plasticity in Physiology and Disease. Cells 2019, 8, 1507. [Google Scholar] [CrossRef] [Green Version]
- Larson, C.J. Translational Pharmacology and Physiology of Brown Adipose Tissue in Human Disease and Treatment. Handb. Exp. Pharmacol. 2019, 251, 381–424. [Google Scholar] [CrossRef]
- Zhao, L.; Zhu, C.; Lu, M.; Chen, C.; Nie, X.; Abudukerimu, B.; Zhang, K.; Ning, Z.; Chen, Y.; Cheng, J.; et al. The Key Role of a Glucagon-Like Peptide-1 Receptor Agonist in Body Fat Redistribution. J. Endocrinol. 2019, 240, 271–286. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Wang, D.; Zhao, W.; Xu, L. Deciphering the Roles of PPARγ in Adipocytes via Dynamic Change of Transcription Complex. Front. Endocrinol. 2018, 21, 473. [Google Scholar] [CrossRef]
- Sidibeh, C.O.; Pereira, M.J.; Lau Börjesson, J.; Kamble, P.G.; Skrtic, S.; Katsogiannos, P.; Sundbom, M.; Svensson, M.K.; Eriksson, J.W. Role of Cannabinoid Receptor 1 in Human Adipose Tissue for Lipolysis Regulation and Insulin Resistance. Endocrine 2017, 55, 839–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Franquet Elía, E.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A.; et al. Activation of Human Brown Adipose Tissue by A β3-Adrenergic Receptor Agonist. Cell Metab. 2015, 21, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbanowicz, T.; Michalak, M.; Olasińska-Wiśniewska, A.; Rodzki, M.; Witkowska, A.; Gąsecka, A.; Buczkowski, P.; Perek, B.; Jemielity, M. Neutrophil Counts, Neutrophil-to-Lymphocyte Ratio, and Systemic Inflammatory Response Index (SIRI) Predict Mortality after Off-Pump Coronary Artery Bypass Surgery. Cells 2022, 11, 1124. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | |
|---|---|
| Gender (m/f) | 28/14 |
| Age, years | 59 (56; 66) |
| History of myocardial infarction, n (%) | 23 (54.8) |
| History of atrial fibrillation, n (%) | 3 (7.1%) |
| Arterial hypertension, n (%) | 42 (100) |
| Diabetes mellitus, n (%) | 12 (28.6) |
| Gensini Score, points | 78.0 (42.5; 121.0) |
| Multivessel CAD, n (%) | 25 (59.5) |
| Ejection fraction, % | 62.0 (47.0; 66.0) |
| Duration of arterial hypertension, years | 20 (15; 23) |
| Duration of coronary artery disease, years | 2 (2; 10) |
| Systolic blood pressure, mm Hg | 130 (123; 140) |
| Diastolic blood pressure, mm Hg | 80 (70; 85) |
| Smoking, n (%) | 17 (40,5) |
| Body Mass Index, kg/m2 | 30.7 (28.1; 33.3) |
| Obesity, n (%) | 25 (59.5) |
| Waist circumference, cm | 106.4 (100; 116) |
| EAT thickness, mm | 5.23 (4.37; 6.30) |
| Metformin, n (%) | 10 (23.8) |
| RAAS inhibitors, n (%) | 36 (85.7) |
| Calcium channels antagonists, n (%) | 26 (61.9) |
| Beta-blockers, n (%) | 38 (90.5) |
| Diuretics, n (%) | 16 (38.1) |
| Statins, n (%) | 41 (97.6) |
| Parameters | Group 1 Diameter of EAT Adipocytes ≤ 87.32 μm (n = 21) | Group 2 Diameter of EAT Adipocytes > 87.32 μm (n = 21) | p |
|---|---|---|---|
| Men/women, n% | 13 (61.9%)/8 (38.1%) | 15 (71.4%)/6 (28.6%) | 0.530 |
| Body Mass Index, kg/m2 | 29.2 (27.0; 31.2) | 32.8 (29.9; 35.4) | 0.036 |
| Waist circumference, cm | 102 (76; 128) | 112 (99; 122) | 0.010 |
| Hip circumference, cm | 104 (101; 108) | 110 (105; 117) | 0.037 |
| EAT thickness, mm | 5.05 (4.37; 6.15) | 5.45 (4.35; 6.60) | 0.690 |
| EAT adipocyte size, µm | 83.79 (79.27; 86.03) | 90.95 (89.09; 95.13) | <0.001 |
| %EAT adipocytes > 100 µm | 8.88 (5.91; 10.82) | 28.22 (18.24; 34.75) | <0.001 |
| %EAT adipocytes < 50 µm | 1.58 (1.04; 2.89) | 1.2 (0.27; 3.85) | 0.630 |
| History of myocardial infarction, n (%) | 11 (52.4) | 12 (57.1) | 0.999 |
| History of atrial fibrillation, n (%) | 1 (4.8) | 2 (9.5) | 0.999 |
| Gensini Score, points | 70.0 (28.0; 100.0) | 107.0 (71.0; 144.0) | 0.008 |
| Multivessel CAD, n (%) | 9 (42.9) | 16 (76.2) | 0.058 |
| Ejection fraction, % | 64.0 (47.0; 69.5) | 61.0 (48.0; 64.0) | 0.346 |
| Parameters | Total (n = 42) | Group 1 Diameter of EAT Adipocytes ≤ 87.32 μm (n = 21) | Group 2 Diameter of EAT Adipocytes > 87.32 μm (n = 21) | p |
|---|---|---|---|---|
| Fasting glucose, mM | 5.89 (5.30; 6.60) | 5.6 (5.1; 6.04) | 6.21 (5.70; 6.79) | 0.078 |
| Fasting insulin, µIU/mL | 5.1 (2.64; 7.84) | 6.02 (2.64; 8.65) | 4.65 (2.66; 5.4) | 0.300 |
| Triglycerides, mM | 1.41 (1.08; 1.92) | 1.2 (0.9; 1.37) | 1.7 (1.41; 2.16) | 0.007 |
| Fasting C-peptide, ng/mL | 2.3 (1.97; 2.98) | 2.15 (1.73; 2.75) | 2.78 (2.18; 3.1) | 0.030 |
| HDL cholesterol, mM | 1.03 (0.88; 1.18) | 1.04 (0.88; 1.18) | 0.99 (0.86; 1.16) | 0.400 |
| LDL cholesterol, mM | 2.04 (1.72; 2.42) | 2.0 (1.75; 2.4) | 2.21 (1.62; 2.56) | 0.082 |
| Apolipoprotein B, mg/dL | 105.28 (8.5; 131.29) | 90.16 (67.28; 112.34) | 112.34 (91.82; 136.82) | 0.080 |
| Apolipoprotein A1, mg/dL | 146.20 (124.57; 168.42) | 140.12 (121.07; 167.56) | 148.06 (134.13; 168.42) | 0.490 |
| apoB/apoA1 | 0.69 (0.49; 0.83) | 0.61 (0.45; 0.76) | 0.73 (0.55; 0.92) | 0.230 |
| EAT Adipocyte Size | Percentage of EAT Adipocytes > 100 μm | Percentage of EAT Adipocytes < 50 μm | ||||
|---|---|---|---|---|---|---|
| Parameters | rs | p | rs | p | rs | p |
| BMI | 0.46 | 0.002 | 0.49 | 0.001 | 0.07 | 0.650 |
| Waist circumference | 0.55 | <0.001 | 0.54 | <0.001 | 0.10 | 0.530 |
| Hip circumference | 0.39 | 0.010 | 0.43 | 0.004 | 0.13 | 0.410 |
| WHR | 0.37 | 0.017 | 0.33 | 0.030 | −0.13 | 0.400 |
| EAT thickness | 0.18 | 0.280 | 0.13 | 0.430 | −0.32 | 0.048 |
| EAT Adipocyte Size | Percentage of EAT Adipocytes > 100 μm | |||
|---|---|---|---|---|
| Parameters | rs | p | rs | p |
| Fasting glycemia | 0.40 | 0.009 | 0.44 | 0.003 |
| Fasting C-peptide | −0.16 | 0.290 | −0.03 | 0.850 |
| Triglycerides | 0.49 | 0.001 | 0.48 | 0.001 |
| Triglycerides/HDL cholesterol | 0.37 | 0.017 | 0.33 | 0.030 |
| LDL cholesterol | 0.09 | 0.590 | 0.15 | 0.330 |
| ApoB | 0.36 | 0.034 | 0.34 | 0.047 |
| ApoA1 | 0.18 | 0.302 | 0.05 | 0.793 |
| EAT Adipocyte Size | Percentage of EAT Adipocytes > 100 μm | Percentage of EAT Adipocytes < 50 μm | ||||
|---|---|---|---|---|---|---|
| Parameters | rs | p | rs | p | rs | p |
| IL-1β | 0.34 | 0.026 | 0.31 | 0.048 | −0.29 | 0.060 |
| IL-6 | −0.12 | 0.450 | −0.001 | 0.990 | 0.40 | 0.009 |
| IL-10 | 0.14 | 0.370 | 0.22 | 0.170 | 0.03 | 0.870 |
| TNF-α | 0.49 | 0.001 | 0.41 | 0.006 | −0.33 | 0.030 |
| LpPLA2 | 0.34 | 0.040 | 0.44 | 0.008 | 0.08 | 0.630 |
| sPLA2 | 0.23 | 0.150 | 0.25 | 0.110 | −0.17 | 0.300 |
| hsCRP | 0.15 | 0.340 | 0.22 | 0.170 | 0.06 | 0.730 |
| CTX-I | −0.43 | 0.005 | −0.41 | 0.007 | −0.13 | 0.42 |
| Parameters | Estimate | p |
|---|---|---|
| Intercept | 0.438 | 0.751 |
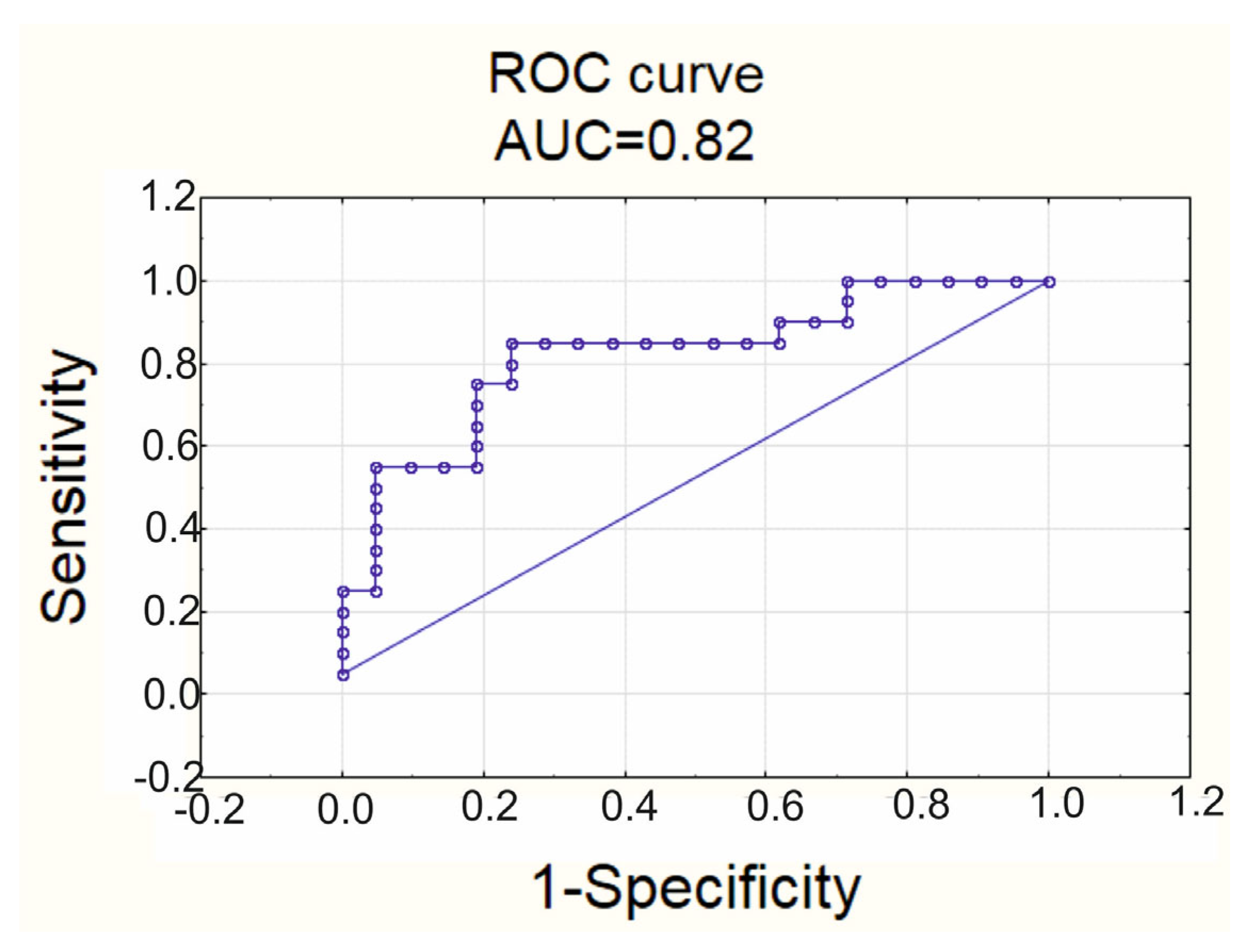
| TNF-α | 0.961 | 0.044 |
| sPLA2 | 0.198 | 0.116 |
| CTX-I | −4.405 | 0.009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kologrivova, I.V.; Naryzhnaya, N.V.; Koshelskaya, O.A.; Suslova, T.E.; Kravchenko, E.S.; Kharitonova, O.A.; Evtushenko, V.V.; Boshchenko, A.A. Association of Epicardial Adipose Tissue Adipocytes Hypertrophy with Biomarkers of Low-Grade Inflammation and Extracellular Matrix Remodeling in Patients with Coronary Artery Disease. Biomedicines 2023, 11, 241. https://doi.org/10.3390/biomedicines11020241
Kologrivova IV, Naryzhnaya NV, Koshelskaya OA, Suslova TE, Kravchenko ES, Kharitonova OA, Evtushenko VV, Boshchenko AA. Association of Epicardial Adipose Tissue Adipocytes Hypertrophy with Biomarkers of Low-Grade Inflammation and Extracellular Matrix Remodeling in Patients with Coronary Artery Disease. Biomedicines. 2023; 11(2):241. https://doi.org/10.3390/biomedicines11020241
Chicago/Turabian StyleKologrivova, Irina V., Natalia V. Naryzhnaya, Olga A. Koshelskaya, Tatiana E. Suslova, Elena S. Kravchenko, Olga A. Kharitonova, Vladimir V. Evtushenko, and Alla A. Boshchenko. 2023. "Association of Epicardial Adipose Tissue Adipocytes Hypertrophy with Biomarkers of Low-Grade Inflammation and Extracellular Matrix Remodeling in Patients with Coronary Artery Disease" Biomedicines 11, no. 2: 241. https://doi.org/10.3390/biomedicines11020241