
Macrophage Phenotype in Combination with Tumor Microbiome Composition Predicts RCC Patients’ Survival: A Pilot Study

 ,
,  , ,
, ,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Ethics Statement
2.2. Immunohistochemical Study
2.3. Quantitative PCR (qPCR)
2.4. 16S rRNA Gene Library Preparation and MiSeq Sequencing
2.5. Bioinformatics Treatment
2.6. Availability of Data
2.7. Statistical Analyses
3. Results
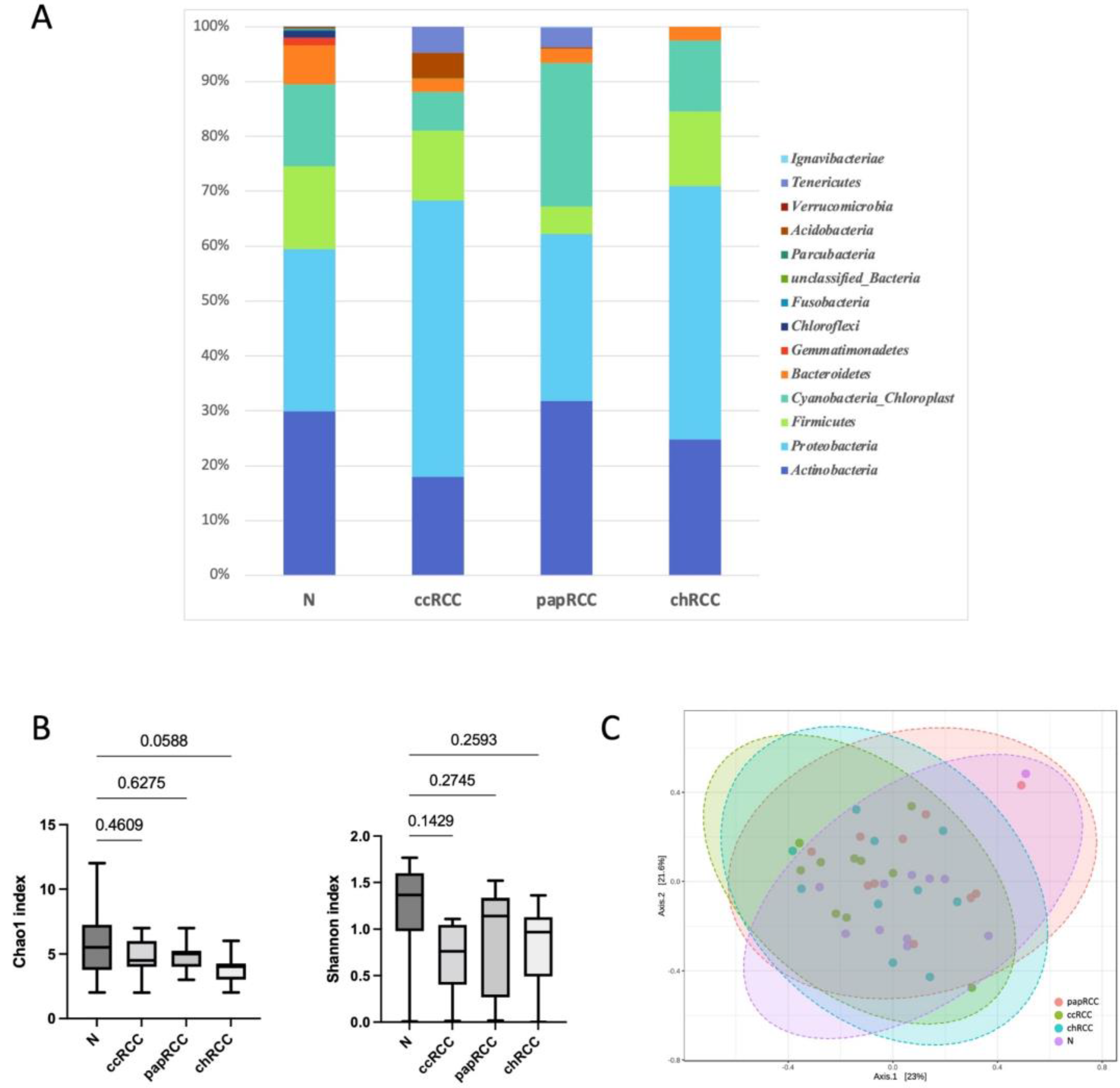
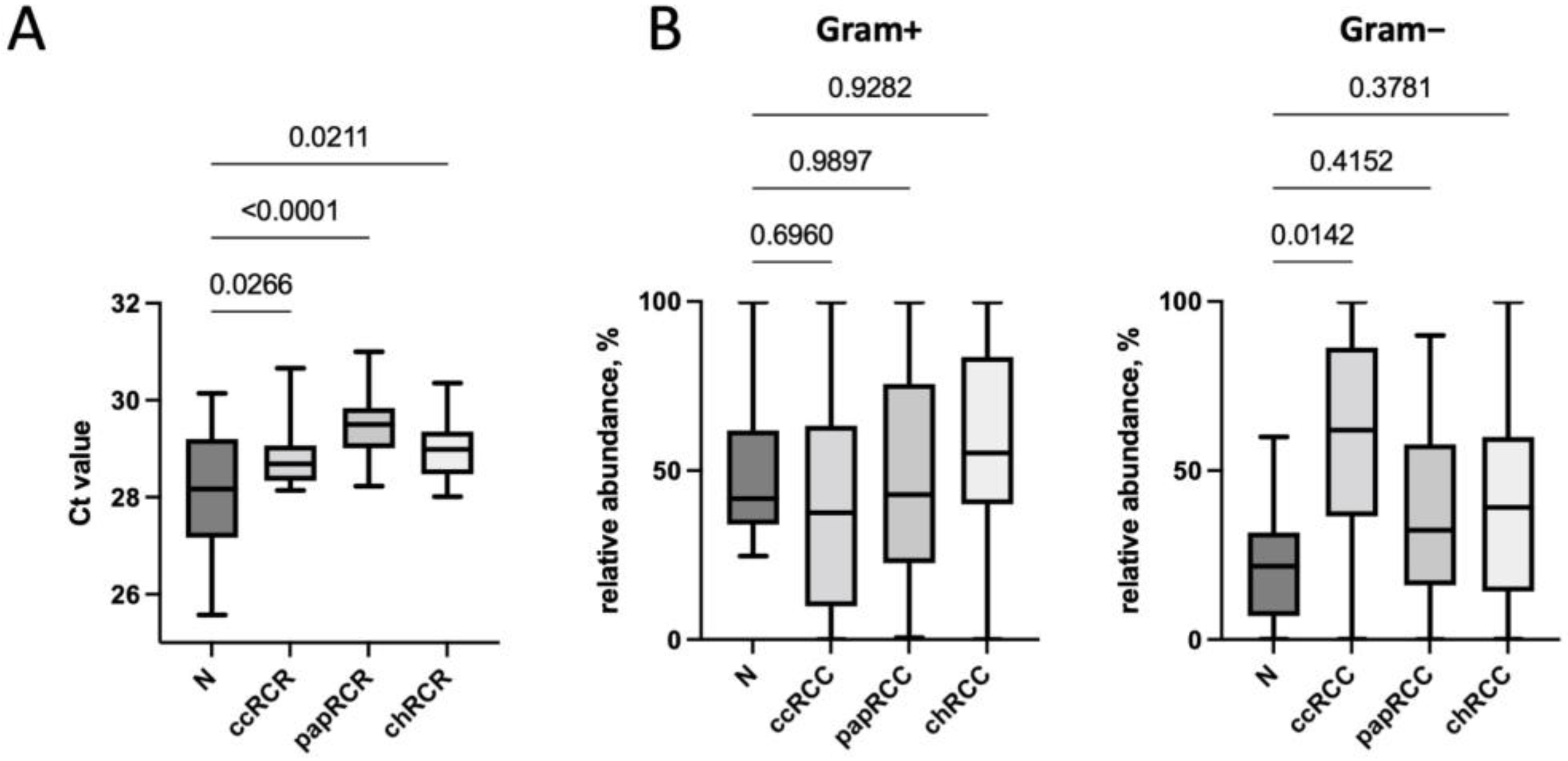
3.1. Renal Tissue Microbiome
3.2. Renal Tissue Stroma
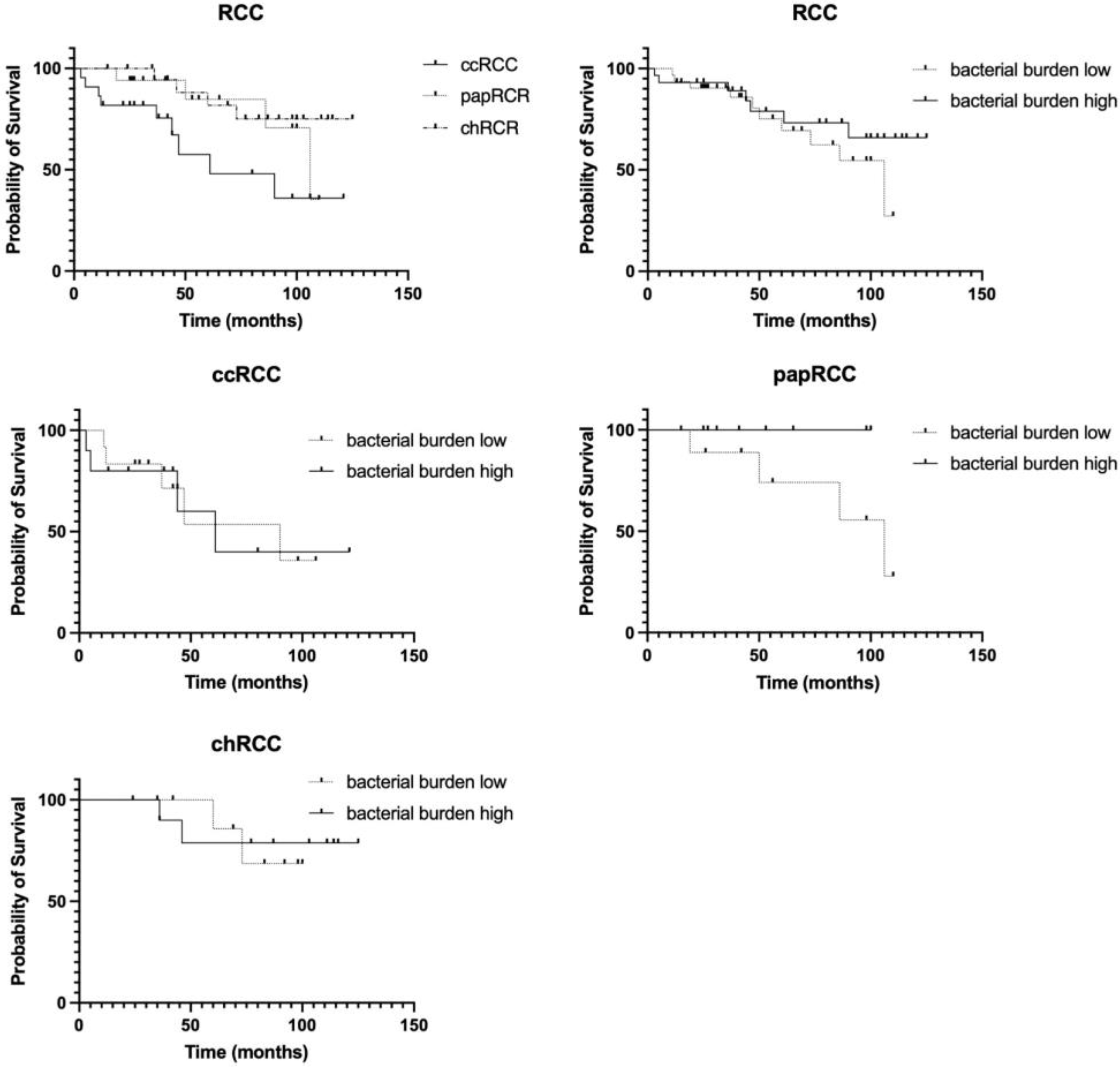
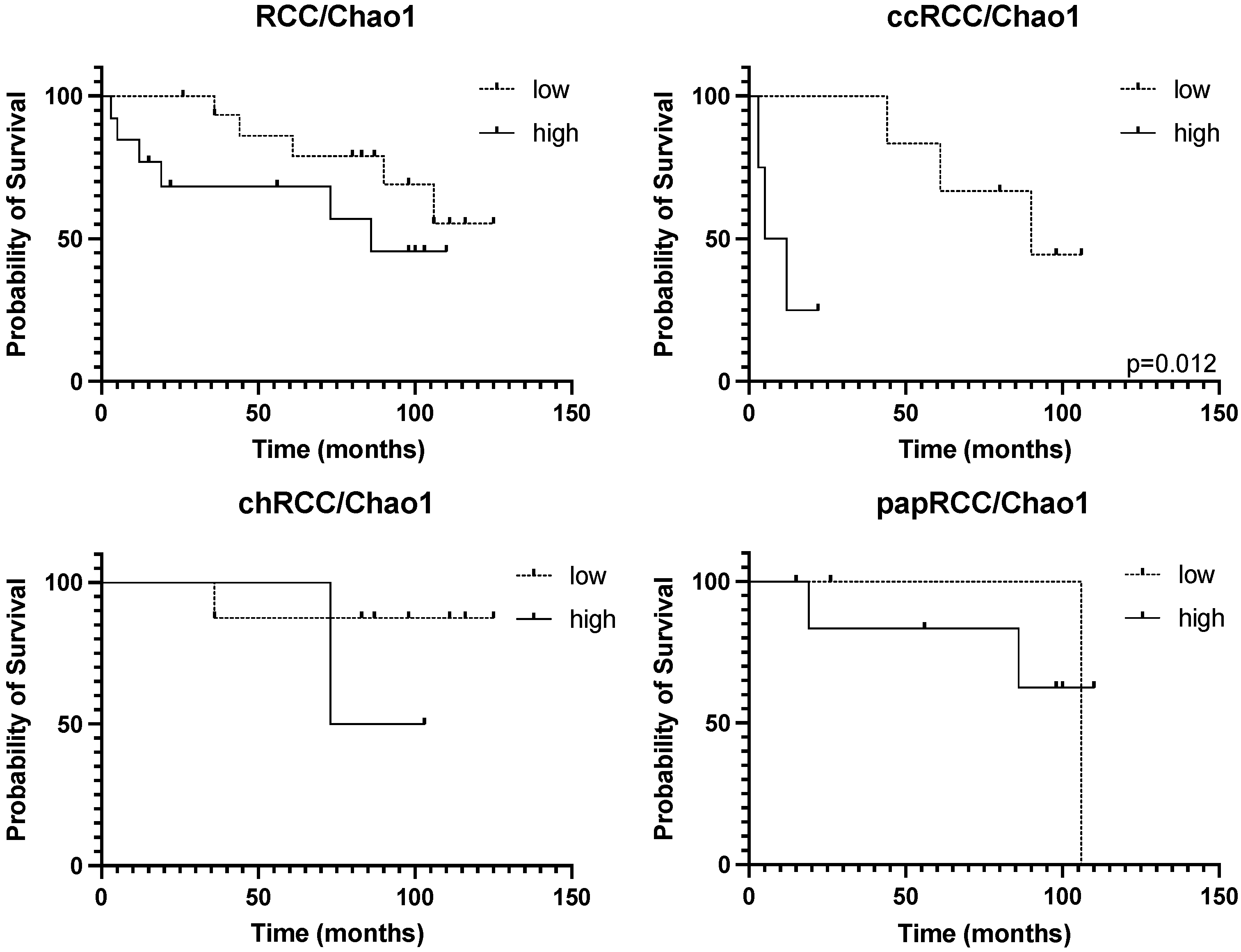
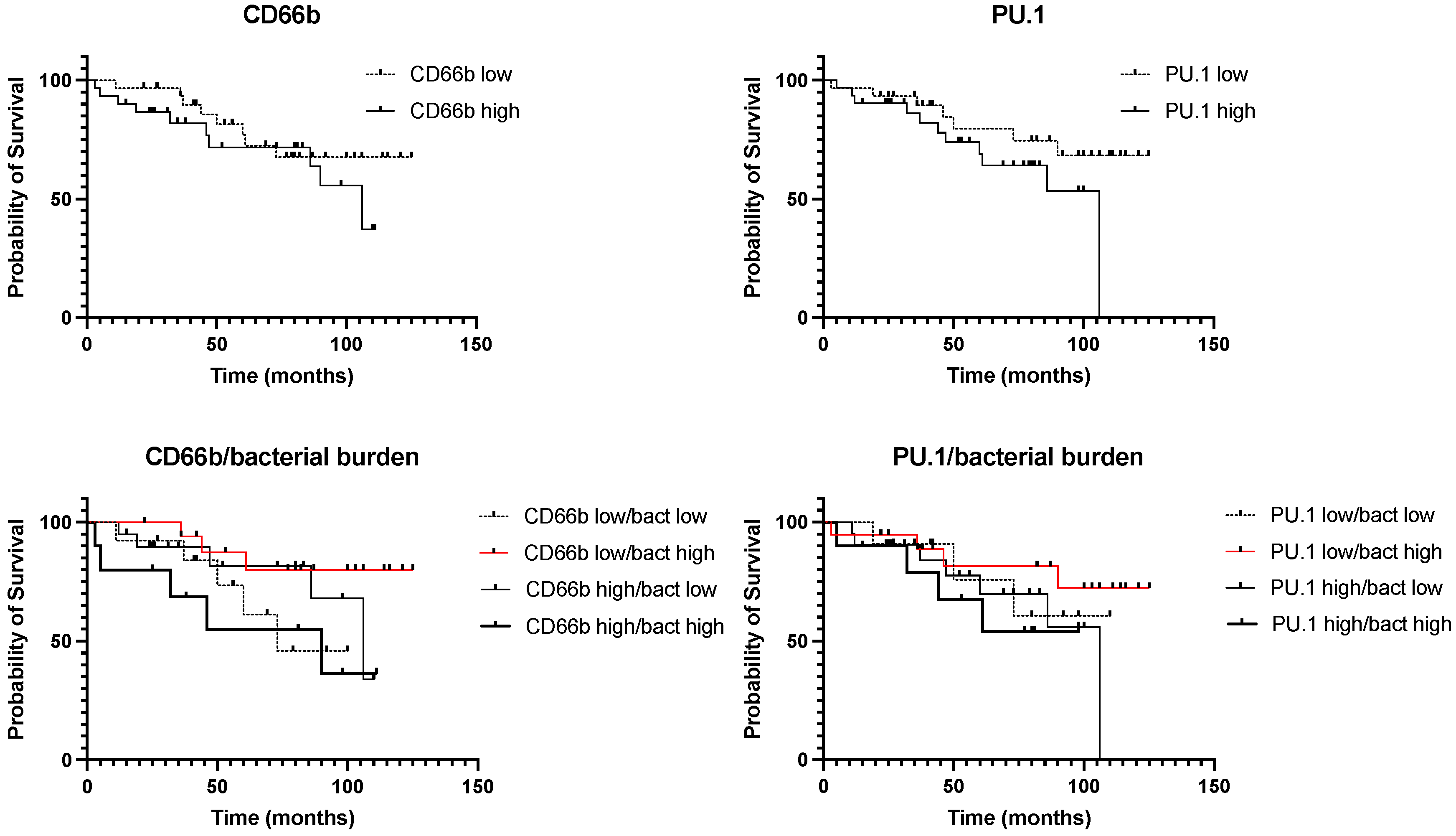
3.3. Survival Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Padala, S.A.; Barsouk, A.; Thandra, K.C.; Saginala, K.; Mohammed, A.; Vakiti, A.; Rawla, P.; Barsouk, A. Epidemiology of Renal Cell Carcinoma. World J. Oncol. 2020, 11, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, K.; Miller, J.D.; Li, J.Z.; Russell, M.W.; Charbonneau, C. Epidemiologic and socioeconomic burden of metastatic renal cell carcinoma (mRCC): A literature review. Cancer Treat. Rev. 2008, 34, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Tsimafeyeu, I.; Zolotareva, T.; Varlamov, S.; Zukov, R.; Petkau, V.; Mazhbich, M.; Statsenko, G.; Safina, S.; Zaitsev, I.; Sakaeva, D.; et al. Five-year Survival of Patients With Metastatic Renal Cell Carcinoma in the Russian Federation: Results From the RENSUR5 Registry. Clin. Genitourin. Cancer 2017, 15, e1069–e1072. [Google Scholar] [CrossRef]
- Athanazio, D.A.; Amorim, L.S.; da Cunha, I.W.; Leite, K.R.M.; da Paz, A.R.; de Paula Xavier Gomes, R.; Tavora, F.R.F.; Faraj, S.F.; Cavalcanti, M.S.; Bezerra, S.M. Classification of renal cell tumors—Current concepts and use of ancillary tests: Recommendations of the Brazilian Society of Pathology. Surg. Exp. Pathol. 2021, 4, 4. [Google Scholar] [CrossRef]
- Muglia, V.F.; Prando, A. Renal cell carcinoma: Histological classification and correlation with imaging findings. Radiol. Bras. 2015, 48, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.; Matrana, M.R.; Tsang, C.; Atkinson, B.; Choueiri, T.K.; Tannir, N.M. Systemic therapy for metastatic non-clear-cell renal cell carcinoma: Recent progress and future directions. Hematol. Oncol. Clin. N. Am. 2011, 25, 853–869. [Google Scholar] [CrossRef] [Green Version]
- Khan, Y.; Slattery, T.D.; Pickering, L.M. Individualizing Systemic Therapies in First Line Treatment and beyond for Advanced Renal Cell Carcinoma. Cancers 2020, 12, 3750. [Google Scholar] [CrossRef]
- Greef, B.; Eisen, T. Medical treatment of renal cancer: New horizons. Br. J. Cancer 2016, 115, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, A.; Mollica, V.; Santoni, M.; Ricci, A.D.; Rosellini, M.; Marchetti, A.; Montironi, R.; Ardizzoni, A.; Massari, F. Impact of Clinicopathological Features on Survival in Patients Treated with First-line Immune Checkpoint Inhibitors Plus Tyrosine Kinase Inhibitors for Renal Cell Carcinoma: A Meta-analysis of Randomized Clinical Trials. Eur. Urol. Focus 2021, 8, 514–521. [Google Scholar] [CrossRef]
- Massari, F.; Rizzo, A.; Mollica, V.; Rosellini, M.; Marchetti, A.; Ardizzoni, A.; Santoni, M. Immune-based combinations for the treatment of metastatic renal cell carcinoma: A meta-analysis of randomised clinical trials. Eur. J. Cancer 2021, 154, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Mollica, V.; Santoni, M.; Matrana, M.R.; Basso, U.; De Giorgi, U.; Rizzo, A.; Maruzzo, M.; Marchetti, A.; Rosellini, M.; Bleve, S.; et al. Concomitant Proton Pump Inhibitors and Outcome of Patients Treated with Nivolumab Alone or Plus Ipilimumab for Advanced Renal Cell Carcinoma. Target. Oncol. 2022, 17, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Barata, P.C.; Rini, B.I. Treatment of renal cell carcinoma: Current status and future directions. CA A Cancer J. Clin. 2017, 67, 507–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, A.E.; Coughlin, L.A.; Kim, J.; Froehlich, T.W.; Xie, Y.; Frenkel, E.P.; Koh, A.Y. Metagenomic Shotgun Sequencing and Unbiased Metabolomic Profiling Identify Specific Human Gut Microbiota and Metabolites Associated with Immune Checkpoint Therapy Efficacy in Melanoma Patients. Neoplasia 2017, 19, 848–855. [Google Scholar] [CrossRef]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.L.; Luke, J.J.; Gajewski, T.F. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Li, S.; Ma, Z.; Liang, S.; Shan, T.; Zhang, M.; Zhu, X.; Zhang, P.; Liu, G.; Zhou, F.; et al. Presence of Porphyromonas gingivalis in esophagus and its association with the clinicopathological characteristics and survival in patients with esophageal cancer. Infect. Agents Cancer 2016, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; San Lucas, A.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell 2019, 178, 795–806.e12. [Google Scholar] [CrossRef]
- Nosho, K.; Sukawa, Y.; Adachi, Y.; Ito, M.; Mitsuhashi, K.; Kurihara, H.; Kanno, S.; Yamamoto, I.; Ishigami, K.; Igarashi, H.; et al. Association of Fusobacterium nucleatum with immunity and molecular alterations in colorectal cancer. World J. Gastroenterol. 2016, 22, 557–566. [Google Scholar] [CrossRef]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Gnanasekar, A.; Lee, A.; Li, W.T.; Haas, M.; Wang-Rodriguez, J.; Chang, E.Y.; Rajasekaran, M.; Ongkeko, W.M. Influence of Intratumor Microbiome on Clinical Outcome and Immune Processes in Prostate Cancer. Cancers 2020, 12, 2524. [Google Scholar] [CrossRef]
- Daisley, B.A.; Chanyi, R.M.; Abdur-Rashid, K.; Al, K.F.; Gibbons, S.; Chmiel, J.A.; Wilcox, H.; Reid, G.; Anderson, A.; Dewar, M.; et al. Abiraterone acetate preferentially enriches for the gut commensal Akkermansia muciniphila in castrate-resistant prostate cancer patients. Nat. Commun. 2020, 11, 4822. [Google Scholar] [CrossRef] [PubMed]
- Maniecki, M.B.; Etzerodt, A.; Ulhoi, B.P.; Steiniche, T.; Borre, M.; Dyrskjot, L.; Orntoft, T.F.; Moestrup, S.K.; Moller, H.J. Tumor-promoting macrophages induce the expression of the macrophage-specific receptor CD163 in malignant cells. Int. J. Cancer J. Int. Du Cancer 2012, 131, 2320–2331. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, O.; Podlesnaya, P.; Rashidova, M.; Samoilova, D.; Petrenko, A.; Zborovskaya, I.; Mochalnikova, V.; Kataev, V.; Khlopko, Y.; Plotnikov, A.; et al. Lung Microbiome Differentially Impacts Survival of Patients with Non-Small Cell Lung Cancer Depending on Tumor Stroma Phenotype. Biomedicines 2020, 8, 349. [Google Scholar] [CrossRef]
- Kovaleva, O.; Podlesnaya, P.; Rashidova, M.; Samoilova, D.; Petrenko, A.; Mochalnikova, V.; Kataev, V.; Khlopko, Y.; Plotnikov, A.; Gratchev, A. Prognostic Significance of the Microbiome and Stromal Cells Phenotype in Esophagus Squamous Cell Carcinoma. Biomedicines 2021, 9, 743. [Google Scholar] [CrossRef] [PubMed]
- Conway Morris, A.; Gadsby, N.; McKenna, J.P.; Hellyer, T.P.; Dark, P.; Singh, S.; Walsh, T.S.; McAuley, D.F.; Templeton, K.; Simpson, A.J.; et al. 16S pan-bacterial PCR can accurately identify patients with ventilator-associated pneumonia. Thorax 2017, 72, 1046–1048. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- USEARCH. Available online: http://drive5.com/usearch (accessed on 20 June 2022).
- Edgar, R.C. UCHIME2: Improved chimera prediction for amplicon sequencing. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- Trimmomatic. Available online: http://www.usadellab.org/cms/?page=trimmomatic (accessed on 20 June 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- RDP. Available online: http://rdp.cme.msu.edu/index.jsp (accessed on 20 June 2022).
- NCBI. Available online: https://blast.ncbi.nlm.nih.gov (accessed on 20 June 2022).
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- EnhancedVolcano. Available online: https://github.com/kevinblighe/EnhancedVolcano (accessed on 20 June 2022).
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Kovaleva, O.V.; Rashidova, M.A.; Samoilova, D.V.; Podlesnaya, P.A.; Tabiev, R.M.; Kuntsevich, N.V.; Efremov, G.D.; Alekseev, B.Y.; Gratchev, A. Immunosuppressive peculiarities of stromal cells of various kidney tumor types. Cancer Urol. 2020, 16, 29–35. [Google Scholar] [CrossRef]
- Xu, L.; Zhu, Y.; Chen, L.; An, H.; Zhang, W.; Wang, G.; Lin, Z.; Xu, J. Prognostic value of diametrically polarized tumor-associated macrophages in renal cell carcinoma. Ann. Surg. Oncol. 2014, 21, 3142–3150. [Google Scholar] [CrossRef] [PubMed]
- Jackute, J.; Zemaitis, M.; Pranys, D.; Sitkauskiene, B.; Miliauskas, S.; Sakalauskas, R. The prognostic influence of tumor infiltrating M1 and M2 phenotype macrophages in resected non-small cell lung cancer. Eur. Respir. J. 2016, 48, OA1775. [Google Scholar] [CrossRef]
- Masucci, M.T.; Minopoli, M.; Carriero, M.V. Tumor Associated Neutrophils. Their Role in Tumorigenesis, Metastasis, Prognosis and Therapy. Front. Oncol. 2019, 9, 1146. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.K.; Donskov, F.; Marcussen, N.; Nordsmark, M.; Lundbeck, F.; von der Maase, H. Presence of intratumoral neutrophils is an independent prognostic factor in localized renal cell carcinoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2009, 27, 4709–4717. [Google Scholar] [CrossRef]
- Donskov, F.; Hokland, M.; Marcussen, N.; Torp Madsen, H.H.; von der Maase, H. Monocytes and neutrophils as ‘bad guys’ for the outcome of interleukin-2 with and without histamine in metastatic renal cell carcinoma--results from a randomised phase II trial. Br. J. Cancer 2006, 94, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Yeh, C.R.; He, D.; Wang, Y.; Xie, H.; Pang, S.T.; Chang, L.S.; Li, L.; Yeh, S. Infiltrating neutrophils promote renal cell carcinoma progression via VEGFa/HIF2alpha and estrogen receptor beta signals. Oncotarget 2015, 6, 19290–19304. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, L.; Bai, Q.; Ou, C.; Xiong, Y.; Qu, Y.; Wang, Z.; Xia, Y.; Guo, J.; Xu, J. Tumor-infiltrating neutrophils predict therapeutic benefit of tyrosine kinase inhibitors in metastatic renal cell carcinoma. Oncoimmunology 2019, 8, e1515611. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, A.; Regolisti, G.; Brusasco, I.; Cabassi, A.; Morabito, S.; Fiaccadori, E. Alterations of intestinal barrier and microbiota in chronic kidney disease. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2015, 30, 924–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liss, M.A.; Chen, Y.; Rodriguez, R.; Pruthi, D.; Johnson-Pais, T.; Wang, H.; Mansour, A.; White, J.R.; Kaushik, D. Microbiome within Primary Tumor Tissue from Renal Cell Carcinoma May Be Associated with PD-L1 Expression of the Venous Tumor Thrombus. Adv. Urol. 2020, 2020, 9068068. [Google Scholar] [CrossRef] [PubMed]
- Heidler, S.; Lusuardi, L.; Madersbacher, S.; Freibauer, C. The Microbiome in Benign Renal Tissue and in Renal Cell Carcinoma. Urol. Int. 2020, 104, 247–252. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Wu, X.; Wang, Z.; Zhang, C.; Cao, G.; Liu, K.; Yan, T. Uncovering the microbiota in renal cell carcinoma tissue using 16S rRNA gene sequencing. J. Cancer Res. Clin. Oncol. 2021, 147, 481–491. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Number of Cases (%) |
|---|---|
| Age | |
| ≤58 | 34 (52%) |
| >58 | 32 (48%) |
| Gender | |
| Male | 34 (52%) |
| Female | 32 (48%) |
| Histology | |
| ccRCR | 23 (35%) |
| papRCC | 19 (29%) |
| chRCR | 24 (36%) |
| Stage | |
| I-II | 48 (73%) |
| III-IV | 18 (27%) |
| Tumor size | |
| T1-T2 | 50 (76%) |
| T3-T4 | 16 (24%) |
| Nodal status | |
| N0 | 63 (95%) |
| N+ | 3 (5%) |
| Phylum | N | ccRCC | papRCC | chRCC | p-Value |
|---|---|---|---|---|---|
| Actinobacteria | 29.88 | 18.00 | 31.77 | 24.79 | 0.7995 |
| Proteobacteria | 29.59 | 50.31 | 30.49 | 46.11 | 0.3673 |
| Firmicutes | 15.04 | 12.75 | 4.99 | 13.60 | 0.7174 |
| Cyanobacteria_Chloroplast | 15.00 | 7.11 | 26.08 | 12.96 | 0.4798 |
| Bacteroidetes | 7.11 | 2.28 | 2.68 | 2.52 | 0.6439 |
| Gemmatimonadetes | 1.32 | 0.00 | 0.00 | 0.00 | 0.0700 |
| Chloroflexi | 1.24 | 0.00 | 0.00 | 0.00 | 0.1629 |
| Fusobacteria | 0.20 | 0.00 | 0.00 | 0.00 | 0.1073 |
| unclassified_Bacteria | 0.18 | 0.14 | 0.00 | 0.00 | 0.5198 |
| Parcubacteria | 0.15 | 0.00 | 0.00 | 0.00 | 0.4040 |
| Acidobacteria | 0.12 | 4.59 | 0.17 | 0.00 | 0.5429 |
| Verrucomicrobia | 0.12 | 0.00 | 0.00 | 0.00 | 0.4040 |
| Tenericutes | 0.02 | 4.82 | 3.62 | 0.00 | 0.5654 |
| Ignavibacteriae | 0.00 | 0.00 | 0.22 | 0.00 | 0.4040 |
| Genus | N | ccRCC | papRCC | chRCC | p-Value |
|---|---|---|---|---|---|
| Kocuria | 8.93 | 0.00 | 0.00 | 0.95 | 0.3949 |
| Phyllobacterium | 5.92 | 2.25 | 3.97 | 0.00 | 0.3311 |
| Micrococcus | 3.67 | 0.00 | 0.00 | 3.15 | 0.4231 |
| Cutibacterium | 3.55 | 10.90 | 13.52 | 9.22 | 0.5331 |
| Corynebacterium | 3.07 | 0.00 | 9.16 | 4.19 | 0.2990 |
| Rothia | 2.93 | 1.46 | 0.00 | 1.47 | 0.2475 |
| Streptococcus | 2.67 | 3.86 | 0.73 | 0.00 | 0.7523 |
| Acinetobacter | 2.13 | 2.64 | 0.62 | 4.75 | 0.7671 |
| Nocardioides | 1.95 | 0.00 | 0.51 | 0.00 | 0.3232 |
| Massilia | 1.95 | 4.54 | 0.00 | 0.87 | 0.6030 |
| Enhydrobacter | 1.80 | 0.00 | 4.50 | 0.00 | 0.3910 |
| Sphingomonas | 1.80 | 7.57 | 1.00 | 2.24 | 0.3585 |
| Staphylococcus | 1.60 | 5.91 | 0.00 | 0.00 | 0.4409 |
| Lactobacillus | 1.60 | 0.00 | 0.00 | 1.17 | 0.4023 |
| Fibrella | 1.58 | 0.00 | 0.29 | 0.00 | 0.4727 |
| Clavibacter | 1.53 | 0.00 | 5.41 | 0.00 | 0.4462 |
| Psychrobacter | 1.30 | 0.00 | 0.38 | 7.14 | 0.5743 |
| Flavobacterium | 1.26 | 1.45 | 0.00 | 0.00 | 0.5244 |
| Lactococcus | 1.19 | 0.00 | 1.53 | 5.44 | 0.3018 |
| Croceibacterium | 1.14 | 0.00 | 2.69 | 0.00 | 0.4798 |
| Phocaeicola | 1.07 | 0.00 | 0.12 | 0.29 | 0.6109 |
| Methylobacterium | 1.04 | 0.00 | 0.00 | 0.00 | 0.4040 |
| Brevundimonas | 0.94 | 0.66 | 1.01 | 0.69 | 0.8674 |
| Escherichia_Shigella | 0.65 | 4.34 | 5.95 | 14.28 | 0.7391 |
| Pseudomonas | 0.63 | 2.42 | 2.51 | 0.00 | 0.6729 |
| Brachybacterium | 0.42 | 0.00 | 0.00 | 2.77 | 0.4619 |
| Aureimonas | 0.41 | 0.00 | 0.00 | 1.28 | 0.4733 |
| Burkholderia | 0.38 | 1.14 | 0.06 | 0.00 | 0.4501 |
| Paracoccus | 0.36 | 3.24 | 1.02 | 0.00 | 0.3761 |
| Lawsonella | 0.35 | 4.15 | 0.00 | 0.66 | 0.5178 |
| Haemophilus | 0.35 | 1.37 | 0.00 | 0.00 | 0.4667 |
| Photobacterium | 0.10 | 4.29 | 0.00 | 0.00 | 0.4203 |
| Mesomycoplasma | 0.02 | 4.82 | 3.62 | 0.00 | 0.5654 |
| Novosphingobium | 0.02 | 0.00 | 0.00 | 12.35 | 0.1438 |
| Anaerobutyricum | 0.00 | 2.97 | 0.00 | 0.00 | 0.4040 |
| Aquicella | 0.00 | 2.75 | 0.00 | 0.00 | 0.4094 |
| Blastocatella | 0.00 | 4.15 | 0.00 | 0.00 | 0.4034 |
| Blastococcus | 0.00 | 1.38 | 0.00 | 0.00 | 0.4040 |
| Catonella | 0.00 | 0.00 | 1.55 | 0.00 | 0.4042 |
| Caulobacter | 0.00 | 0.00 | 1.00 | 0.00 | 0.4040 |
| Cloacibacterium | 0.00 | 0.00 | 0.00 | 1.84 | 0.4041 |
| Dermabacter | 0.00 | 0.00 | 0.00 | 2.18 | 0.4042 |
| Jeotgalicoccus | 0.00 | 0.00 | 0.00 | 4.50 | 0.4041 |
| Lautropia | 0.00 | 1.10 | 0.00 | 0.00 | 0.4043 |
| Neisseria | 0.00 | 1.86 | 0.00 | 0.00 | 0.4042 |
| Pedobacter | 0.00 | 0.00 | 1.32 | 0.00 | 0.4040 |
| Roseomonas | 0.00 | 6.33 | 0.00 | 0.07 | 0.4100 |
| Simplicispira | 0.00 | 2.40 | 2.75 | 0.00 | 0.5740 |
| Tetragenococcus | 0.00 | 0.00 | 0.00 | 2.20 | 0.4044 |
| Histology | Stage | ||||||
|---|---|---|---|---|---|---|---|
| ccRCC | papRCC | chRCC | p | I–II | III–IV | p | |
| CD68 | 18.0 (8.6–22.4) | 11.4 (5.8–15.4) | 9.5 (4.5–13.4) | 0.041 * | 10.8 (6.1–16.8) | 18.5 (4.9–22.5) | 0.230 |
| PU.1 | 17.4 (7.2–25.8) | 20.2 (7.8–29.8) | 7.0 (3.6–14.6) | 0.054 | 10.8 (5.3–24.7) | 17.3 (6.7–27.7) | 0.268 |
| CD163 | 11.6 (6.2–15.8) | 8.4 (5.6–11.0) | 3.6 (2.7–6.8) | 0.043 * | 6.7 (3.3–11.5) | 9.9 (3.8–15.7) | 0.203 |
| CD66b | 2.6 (1.4–5.6) | 4.8 (2.0–8.0) | 1.7 (1.2–3.0) | 0.019 * | 2.5 (1.2–5.2) | 2.3 (1.6–5.0) | 0.980 |
| CD56 | 1.8 (0.6–11.8) | 1.2 (0.8–3.6) | 1.7 (0.7–9.5) | 0.611 | 1.6 (0.7–7.3) | 1.5 (0.9–9.85) | 0.788 |
| CD20 | 2.6 (1.0–5.8) | 3.2 (1.4–5.4) | 2.7 (0.6–6.5) | 0.603 | 3.0 (0.9–6.0) | 3.6 (0.9–4.9) | 0.957 |
| CD3 | 16.8 (4.2–32.8) | 7.2 (5.4–12.4) | 1.8 (1.1–6.1) | 0.001 * | 5.5 (1.9–12.6) | 9.7 (1.8–34.3) | 0.152 |
| CD8 | 7.4 (1.8–20.0) | 7.0 (2.0–10.6) | 2.4 (0.6–3.6) | 0.002 * | 3.5 (1.5–7.7) | 8.1 (1.2–22.75) | 0.141 |
| FoxP3 | 0.2 (0.0–0.6) | 0.4 (0.0–0.8) | 0.4 (0.0–0.8) | 0.383 | 0.4 (0.0–0.8) | 0.2 (0.0–0.6) | 0.336 |
| Bacterial Burden vs. CD68 | Bacterial Burden vs. PU.1 | Bacterial Burden vs. CD163 | Bacterial Burden vs. CD66b | Bacterial Burden vs. CD56 | |
|---|---|---|---|---|---|
| Spearman r | |||||
| r | 0.040 | 0.301 | 0.130 | 0.326 | 0.056 |
| 95% confidence interval | −0.2107 to 0.2864 | 0.05736 to 0.5123 | −0.1226 to 0.3672 | 0.08423 to 0.5320 | −0.1949 to 0.3013 |
| p-value | |||||
| p (two-tailed) | 0.747 | 0.013 * | 0.297 | 0.007 * | 0.651 |
| Bacterial Burden vs. iNOS | Bacterial Burden vs. CD20 | Bacterial Burden vs. CD3 | Bacterial Burden vs. CD8 | Bacterial Burden vs. FoxP3 | |
| Spearman r | |||||
| r | 0.124 | 0.161 | 0.172 | 0.100 | 0.186 |
| 95% confidence interval | −0.1289 to 0.3617 | −0.09073 to 0.3948 | −0.07977 to 0.4041 | −0.1524 to 0.3407 | −0.06501 to 0.4164 |
| p-value | |||||
| p (two-tailed) | 0.321 | 0.194 | 0.165 | 0.423 | 0.132 |
| Markers | Univariate Analysis | Multivariate Analysis | ||||
|---|---|---|---|---|---|---|
| HR | 95% CI | p | HR | 95% CI | p | |
| CD68 (high/low) | 4.798 | 1.883 to 12.23 | 0.002 * | 0.995 | 0.889 to 1.102 | 0.934 |
| PU.1 (high/low) | 1.906 | 0.755 to 4.822 | 0.165 | 0.995 | 0.881 to 1.086 | 0.937 |
| CD163 (high/low) | 7.734 | 3.003 to 19.92 | <0.0001 * | 1.181 | 1.019 to 1.435 | 0.046 * |
| CD66b (high/low) | 1.605 | 0.632 to 4.073 | 0.312 | 1.172 | 0.993 to 1.377 | 0.051 |
| CD56 (high/low) | 1.103 | 0.437 to 2.785 | 0.835 | 1.029 | 0.979 to 1.077 | 0.189 |
| CD20 (high/low) | 0.650 | 0.258 to 1.638 | 0.367 | 0.671 | 0.399 to 0.872 | 0.024 * |
| CD3 (high/low) | 1.650 | 0.649 to 4.195 | 0.271 | 1.045 | 0.922 to 1.206 | 0.501 |
| CD8 (high/low) | 4.670 | 1.836 to 11.88 | 0.002 * | 0.938 | 0.772 to 1.116 | 0.453 |
| FoxP3 (high/low) | 1.256 | 0.498 to 3.165 | 0.6263 | 0.414 | 0.055 to 1.980 | 0.3133 |
| iNOS (high/low) | 0.319 | 0.089 to 1.143 | 0.2404 | 0.005 | 0.00001 to 0.395 | 0.0402 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovaleva, O.V.; Podlesnaya, P.; Sorokin, M.; Mochalnikova, V.; Kataev, V.; Khlopko, Y.A.; Plotnikov, A.O.; Stilidi, I.S.; Kushlinskii, N.E.; Gratchev, A. Macrophage Phenotype in Combination with Tumor Microbiome Composition Predicts RCC Patients’ Survival: A Pilot Study. Biomedicines 2022, 10, 1516. https://doi.org/10.3390/biomedicines10071516
Kovaleva OV, Podlesnaya P, Sorokin M, Mochalnikova V, Kataev V, Khlopko YA, Plotnikov AO, Stilidi IS, Kushlinskii NE, Gratchev A. Macrophage Phenotype in Combination with Tumor Microbiome Composition Predicts RCC Patients’ Survival: A Pilot Study. Biomedicines. 2022; 10(7):1516. https://doi.org/10.3390/biomedicines10071516
Chicago/Turabian StyleKovaleva, Olga V., Polina Podlesnaya, Maxim Sorokin, Valeria Mochalnikova, Vladimir Kataev, Yuriy A. Khlopko, Andrey O. Plotnikov, Ivan S. Stilidi, Nikolay E. Kushlinskii, and Alexei Gratchev. 2022. "Macrophage Phenotype in Combination with Tumor Microbiome Composition Predicts RCC Patients’ Survival: A Pilot Study" Biomedicines 10, no. 7: 1516. https://doi.org/10.3390/biomedicines10071516