
Modulation of Rxrα Expression in Mononuclear Phagocytes Impacts on Cardiac Remodeling after Ischemia-Reperfusion Injury
 ,
,  ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Ischemia/Reperfusion (I/R) Injury
2.3. Tissue and Organ Harvesting
2.4. Blood Analysis
2.5. FACS (Sorting)
2.5.1. Preparation of Blood and BM Samples
2.5.2. Preparation of Heart and Brain Samples
2.5.3. Phagocytosis Assay Using pHrodo™ Red BioParticles®
2.6. Immunofluorescence Microscopy (IF)
2.7. Quantification of Apoptotic Cells in the Heart on d1 after MI
2.8. (Real Time Quantitative) PCR
2.9. Gel Electrophoresis
2.10. Cytokine Array
2.11. Positron Emission Tomography (PET) Imaging
2.12. Statistical Analysis
3. Results
3.1. Rxrα Expression Is Reduced in Blood Monocytes of Cx3cr1Cre-ert2-YFPRxrαflox/flox Mice
3.2. Reduced Rxrα Expression in Myeloid Cells Has no Impact on Haematopoiesis
3.3. RXRα Influences Monocyte and Macrophage Subset Composition
3.4. Reduced Expression of Rxrα in Monocytes and Macrophages Increases Infarct Size after MI
3.5. Phagocytic Function of Myeloid Cells Is Altered by a Reduced Rxrα-Expression
3.6. Proliferation of Cardiac Macrophages Is Dependent on RXRα
3.7. RXRα Is Involved in Regulation of Angiogenesis after MI
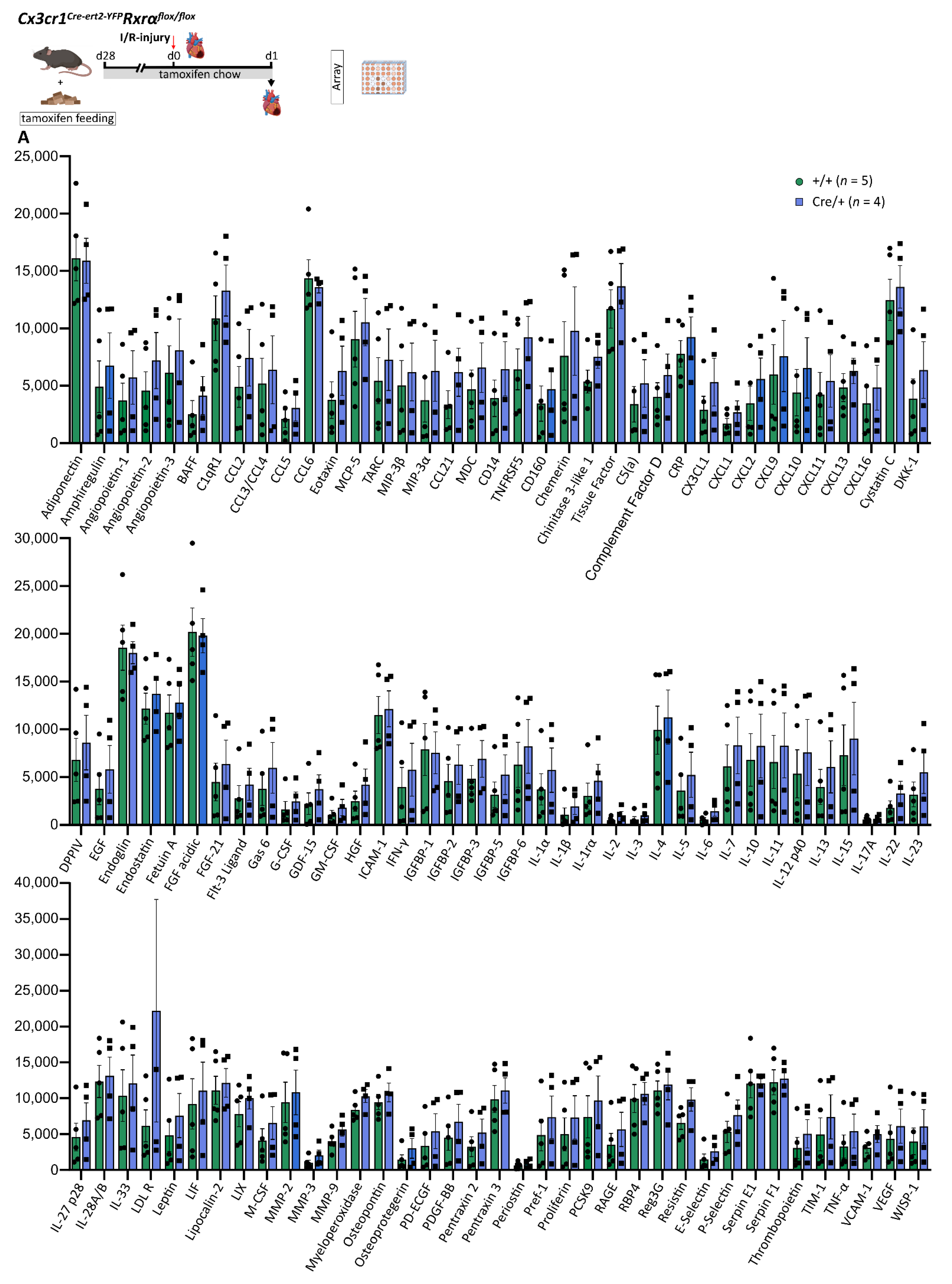
3.8. Reduced Rxrα Expression in Myeloid Cells Has No Impact on the Cytokine and Chemokine Profile in the Infarcted Heart on d1 after MI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yellon, D.M.; Hausenloy, D.J. Myocardial reperfusion injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Frantz, S.; Nahrendorf, M. Cardiac macrophages and their role in ischaemic heart disease. Cardiovasc. Res. 2014, 102, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Nahrendorf, M.; Swirski, F.K.; Aikawa, E.; Stangenberg, L.; Wurdinger, T.; Figueiredo, J.-L.; Libby, P.; Weissleder, R.; Pittet, M.J. The healing myocardium sequentially mobilizes two monocyte subsets with divergent and complementary functions. J. Exp. Med. 2007, 204, 3037–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panizzi, P.; Swirski, F.K.; Figueiredo, J.-L.; Waterman, P.; Sosnovik, D.E.; Aikawa, E.; Libby, P.; Pittet, M.; Weissleder, R.; Nahrendorf, M. Impaired infarct healing in atherosclerotic mice with Ly-6C(hi) monocytosis. J. Am. Coll. Cardiol. 2010, 55, 1629–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epelman, S.; Lavine, K.J.; Beaudin, A.E.; Sojka, D.K.; Carrero, J.A.; Calderon, B.; Brija, T.; Gautier, E.L.; Ivanov, S.; Satpathy, A.T.; et al. Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation. Immunity 2014, 40, 91–104. [Google Scholar] [CrossRef] [Green Version]
- van Amerongen, M.J.; Harmsen, M.C.; van Rooijen, N.; Petersen, A.H.; van Luyn, M.J.A. Macrophage depletion impairs wound healing and increases left ventricular remodeling after myocardial injury in mice. Am. J. Pathol. 2007, 170, 818–829. [Google Scholar] [CrossRef] [Green Version]
- Dawson, M.I.; Xia, Z. The retinoid X receptors and their ligands. Biochim. Biophys. Acta 2012, 1821, 21–56. [Google Scholar] [CrossRef] [Green Version]
- Rőszer, T.; Menéndez-Gutiérrez, M.P.; Cedenilla, M.; Ricote, M. Retinoid X receptors in macrophage biology. Trends Endocrinol. Metab. 2013, 24, 460–468. [Google Scholar] [CrossRef]
- Shao, M.; Lu, L.; Wang, Q.; Ma, L.; Tian, X.; Li, C.; Li, C.; Guo, D.; Wang, Q.; Wang, W.; et al. The multi-faceted role of retinoid X receptor in cardiovascular diseases. Biomed. Pharmacother. 2021, 137, 111264. [Google Scholar] [CrossRef]
- Núñez, V.; Alameda, D.; Rico, D.; Mota, R.; Gonzalo, P.; Cedenilla, M.; Fischer, T.; Boscá, L.; Glass, C.K.; Arroyo, A.G.; et al. Retinoid X receptor alpha controls innate inflammatory responses through the up-regulation of chemokine expression. Proc. Natl. Acad. Sci. USA 2010, 107, 10626–10631. [Google Scholar] [CrossRef] [Green Version]
- Sanz, M.-J.; Albertos, F.; Otero, E.; Juez, M.; Morcillo, E.J.; Piqueras, L. Retinoid X receptor agonists impair arterial mononuclear cell recruitment through peroxisome proliferator-activated receptor-γ activation. J. Immunol. 2012, 189, 411–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Storer, P.D.; Chavis, J.A.; Racke, M.K.; Drew, P.D. Agonists for the peroxisome proliferator-activated receptor-alpha and the retinoid X receptor inhibit inflammatory responses of microglia. J. Neurosci. Res. 2005, 81, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Allen, C.; Ballow, M. Retinoic acid enhances the production of IL-10 while reducing the synthesis of IL-12 and TNF-alpha from LPS-stimulated monocytes/macrophages. J. Clin. Immunol. 2007, 27, 193–200. [Google Scholar] [CrossRef]
- Daniel, B.; Nagy, G.; Hah, N.; Horvath, A.; Czimmerer, Z.; Poliska, S.; Gyuris, T.; Keirsse, J.; Gysemans, C.; Van Ginderachter, J.A.; et al. The active enhancer network operated by liganded RXR supports angiogenic activity in macrophages. Genes Dev. 2014, 28, 1562–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, A.; Sen, T.; Chatterjee, A. All-trans retinoic acid (ATRA) downregulates MMP-9 by modulating its regulatory molecules. Cell Adh. Migr. 2010, 4, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Montemurro, P.; Barbuti, G.; Conese, M.; Gabriele, S.; Petio, M.; Colucci, M.; Semeraro, N. Retinoic acid stimulates plasminogen activator inhibitor 2 production by blood mononuclear cells and inhibits urokinase-induced extracellular proteolysis. Br. J. Haematol. 1999, 107, 294–299. [Google Scholar] [CrossRef]
- Roszer, T.; Menéndez-Gutiérrez, M.P.; Lefterova, M.I.; Alameda, D.; Núñez, V.; Lazar, M.A.; Fischer, T.; Ricote, M. Autoimmune kidney disease and impaired engulfment of apoptotic cells in mice with macrophage peroxisome proliferator-activated receptor gamma or retinoid X receptor alpha deficiency. J. Immunol. 2011, 186, 621–631. [Google Scholar] [CrossRef]
- Ting, S.-M.; Zhao, X.; Sun, G.; Obertas, L.; Ricote, M.; Aronowski, J. Brain cleanup as a potential target for poststroke recovery: The role of RXR (retinoic X receptor) in phagocytes. Stroke 2020, 51, 958–966. [Google Scholar] [CrossRef]
- Zimmerman, T.L.; Thevananther, S.; Ghose, R.; Burns, A.R.; Karpen, S.J. Nuclear export of retinoid X receptor alpha in response to interleukin-1beta-mediated cell signaling: Roles for JNK and SER260. J. Biol. Chem. 2006, 281, 15434–15440. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Tabeta, K.; Mann, N.; Crozat, K.; Mudd, S.; Beutler, B. An essential role for Rxr alpha in the development of Th2 responses. Eur. J. Immunol. 2005, 35, 3414–3423. [Google Scholar] [CrossRef]
- Chen, J.; Kubalak, S.W.; Chien, K.R. Ventricular muscle-restricted targeting of the RXRalpha gene reveals a non-cell-autonomous requirement in cardiac chamber morphogenesis. Development 1998, 125, 1943–1949. [Google Scholar] [CrossRef] [PubMed]
- Kiermayer, C.; Conrad, M.; Schneider, M.; Schmidt, J.; Brielmeier, M. Optimization of spatiotemporal gene inactivation in mouse heart by oral application of tamoxifen citrate. Genesis 2007, 45, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Zacherl, M.J.; Weckbach, L.; Paintmayer, L.; Weinberger, T.; Stark, K.; Massberg, S.; Bartenstein, P.; Lehner, S.; Schulz, C.; et al. Cardiac 18F-FDG Positron Emission Tomography: An Accurate Tool to Monitor In vivo Metabolic and Functional Alterations in Murine Myocardial Infarction. Front. Cardiovasc. Med. 2021, 8, 656742. [Google Scholar] [CrossRef]
- Taschner, S.; Koesters, C.; Platzer, B.; Jörgl, A.; Ellmeier, W.; Benesch, T.; Strobl, H. Down-regulation of RXRalpha expression is essential for neutrophil development from granulocyte/monocyte progenitors. Blood 2007, 109, 971–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Shen, L.-H.; Hu, L.-H.; Ge, H.; Pu, J.; Chai, D.-J.; Shao, Q.; Wang, L.; Zeng, J.-Z.; He, B. Retinoid X receptor agonists inhibit phorbol-12-myristate-13-acetate (PMA)-induced differentiation of monocytic THP-1 cells into macrophages. Mol. Cell. Biochem. 2010, 335, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Ricote, M.; Snyder, C.S.; Leung, H.-Y.; Chen, J.; Chien, K.R.; Glass, C.K. Normal hematopoiesis after conditional targeting of RXRalpha in murine hematopoietic stem/progenitor cells. J. Leukoc. Biol. 2006, 80, 850–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahrendorf, M.; Pittet, M.J.; Swirski, F.K. Monocytes: Protagonists of infarct inflammation and repair after myocardial infarction. Circulation 2010, 121, 2437–2445. [Google Scholar] [CrossRef]
- Lavine, K.J.; Epelman, S.; Uchida, K.; Weber, K.J.; Nichols, C.G.; Schilling, J.D.; Ornitz, D.M.; Randolph, G.J.; Mann, D.L. Distinct macrophage lineages contribute to disparate patterns of cardiac recovery and remodeling in the neonatal and adult heart. Proc. Natl. Acad. Sci. USA 2014, 111, 16029–16034. [Google Scholar] [CrossRef] [Green Version]
- Certo, M.; Endo, Y.; Ohta, K.; Sakurada, S.; Bagetta, G.; Amantea, D. Activation of RXR/PPARγ underlies neuroprotection by bexarotene in ischemic stroke. Pharmacol. Res. 2015, 102, 298–307. [Google Scholar] [CrossRef]
- Degos, L.; Wang, Z.Y. All trans retinoic acid in acute promyelocytic leukemia. Oncogene 2001, 20, 7140–7145. [Google Scholar] [CrossRef] [Green Version]
- Shiohara, M.; Dawson, M.I.; Hobbs, P.D.; Sawai, N.; Higuchi, T.; Koike, K.; Komiyama, A.; Koeffler, H.P. Effects of novel RAR- and RXR-selective retinoids on myeloid leukemic proliferation and differentiation in vitro. Blood 1999, 93, 2057–2066. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 13 April 2021).
- Germain, P.; Chambon, P.; Eichele, G.; Evans, R.M.; Lazar, M.A.; Leid, M.; De Lera, A.R.; Lotan, R.; Mangelsdorf, D.J.; Gronemeyer, H. International Union of Pharmacology. LXIII. Retinoid X receptors. Pharmacol. Rev. 2006, 58, 760–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochain, C.; Channon, K.M.; Silvestre, J.-S. Angiogenesis in the infarcted myocardium. Antioxid. Redox Signal. 2013, 18, 1100–1113. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Maeda, K.; Takefuji, M.; Kikuchi, R.; Morishita, Y.; Hirashima, M.; Murohara, T. Dynamics of angiogenesis in ischemic areas of the infarcted heart. Sci. Rep. 2017, 7, 7156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Zhao, W.; Chen, Y.; Ahokas, R.A.; Sun, Y. Vascular endothelial growth factor (VEGF)-A: Role on cardiac angiogenesis following myocardial infarction. Microvasc. Res. 2010, 80, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Henson, P.M. Dampening inflammation. Nat. Immunol. 2005, 6, 1179–1181. [Google Scholar] [CrossRef]
- Ravichandran, K.S.; Lorenz, U. Engulfment of apoptotic cells: Signals for a good meal. Nat. Rev. Immunol. 2007, 7, 964–974. [Google Scholar] [CrossRef]
- Hilgendorf, I.; Gerhardt, L.M.S.; Tan, T.C.; Winter, C.; Holderried, T.A.W.; Chousterman, B.G.; Iwamoto, Y.; Liao, R.; Zirlik, A.; Scherer-Crosbie, M.; et al. Ly-6Chigh monocytes depend on Nr4a1 to balance both inflammatory and reparative phases in the infarcted myocardium. Circ. Res. 2014, 114, 1611–1622. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Räuber, S.; Fischer, M.; Messerer, D.; Wimmler, V.; Konda, K.; Todica, A.; Lorenz, M.; Titova, A.; Schulz, C.; Weinberger, T. Modulation of Rxrα Expression in Mononuclear Phagocytes Impacts on Cardiac Remodeling after Ischemia-Reperfusion Injury. Biomedicines 2022, 10, 1274. https://doi.org/10.3390/biomedicines10061274
Räuber S, Fischer M, Messerer D, Wimmler V, Konda K, Todica A, Lorenz M, Titova A, Schulz C, Weinberger T. Modulation of Rxrα Expression in Mononuclear Phagocytes Impacts on Cardiac Remodeling after Ischemia-Reperfusion Injury. Biomedicines. 2022; 10(6):1274. https://doi.org/10.3390/biomedicines10061274
Chicago/Turabian StyleRäuber, Saskia, Maximilian Fischer, Denise Messerer, Vanessa Wimmler, Kumaraswami Konda, Andrei Todica, Michael Lorenz, Anna Titova, Christian Schulz, and Tobias Weinberger. 2022. "Modulation of Rxrα Expression in Mononuclear Phagocytes Impacts on Cardiac Remodeling after Ischemia-Reperfusion Injury" Biomedicines 10, no. 6: 1274. https://doi.org/10.3390/biomedicines10061274