
Resveratrol and ω-3 PUFAs Promote Human Macrophage Differentiation and Function

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Media
2.2. Isolation and Culture of Monocytes from Human Peripheral Blood Mononuclear Cells (PBMC)
2.3. THP-1 Cells
2.4. Cytofluorometry
2.5. Analysis of Cytokines, Chemokines and PGE2
2.6. Statistical Analysis
2.7. Dosage Information
3. Results
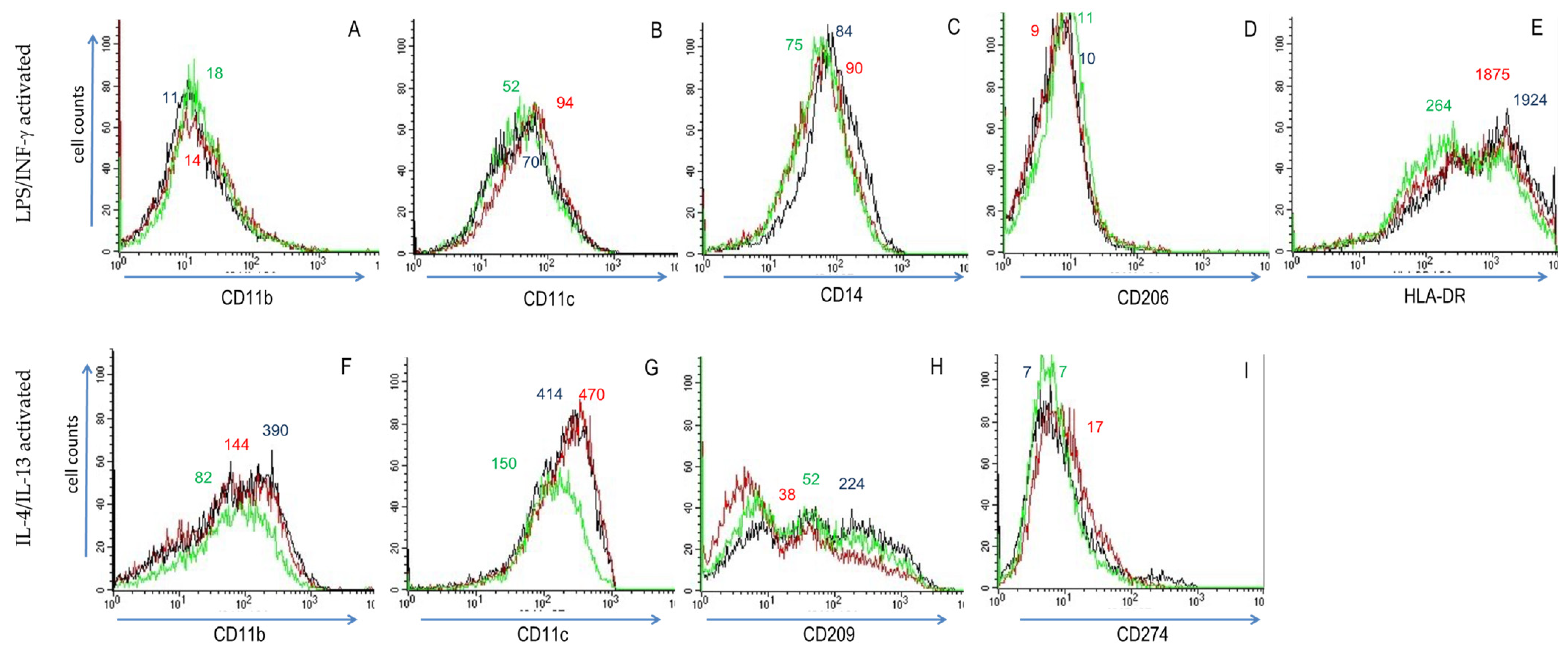
3.1. Activation of THP-1 Cells with LPS/IFN-γ or IL-4/IL-13 Induced Phenotypic Changes That Are Modulated by Res and ω-3 PUFAs
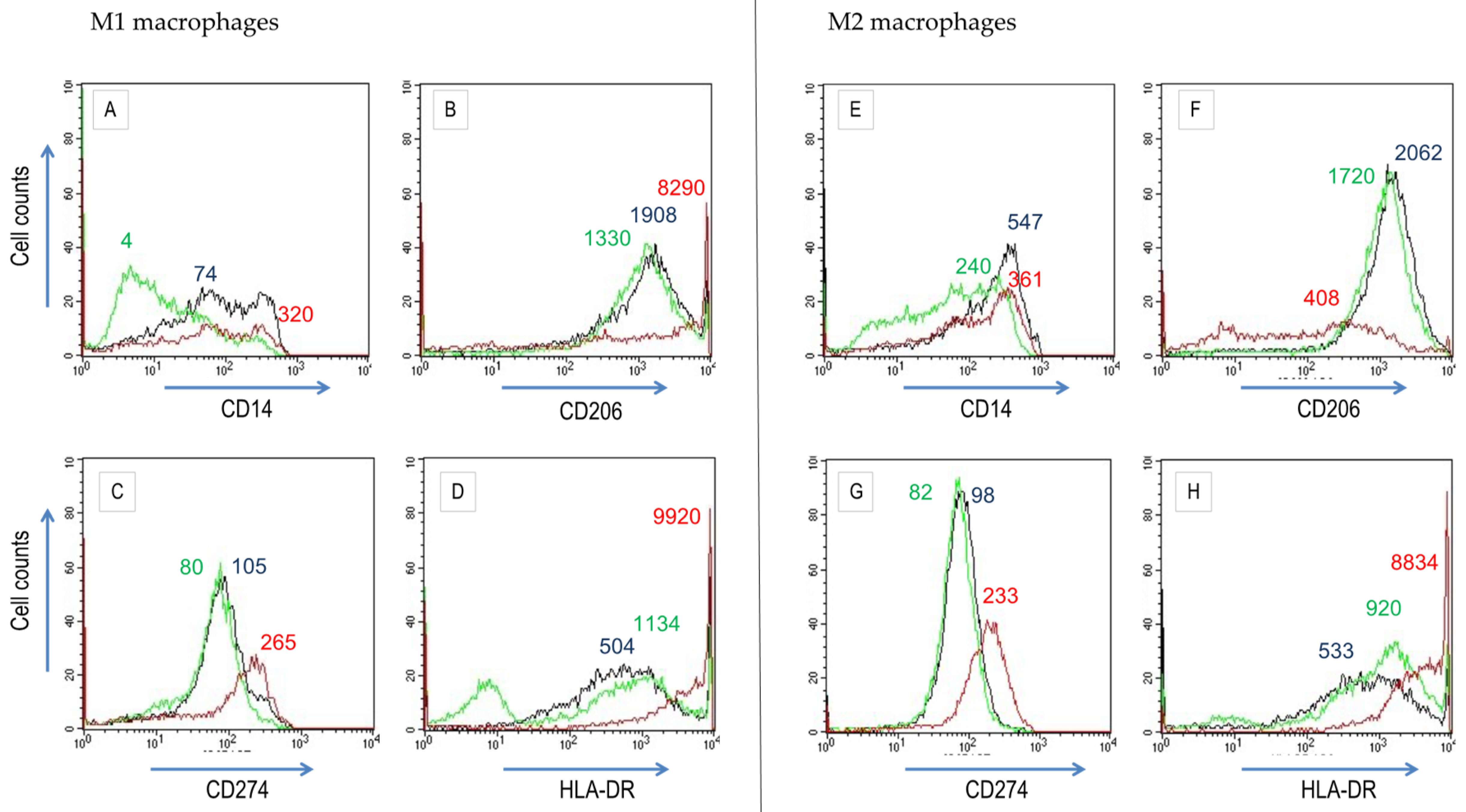
3.2. Res and Omega-3 PUFAs Modulate M1 and M2 Differentiation of Macrophages Derived from Peripheral Blood Mononuclear Cells (PBMCs)
3.3. Res and Omega-3 PUFAs Influence the Functions of Activated THP-1 Cells
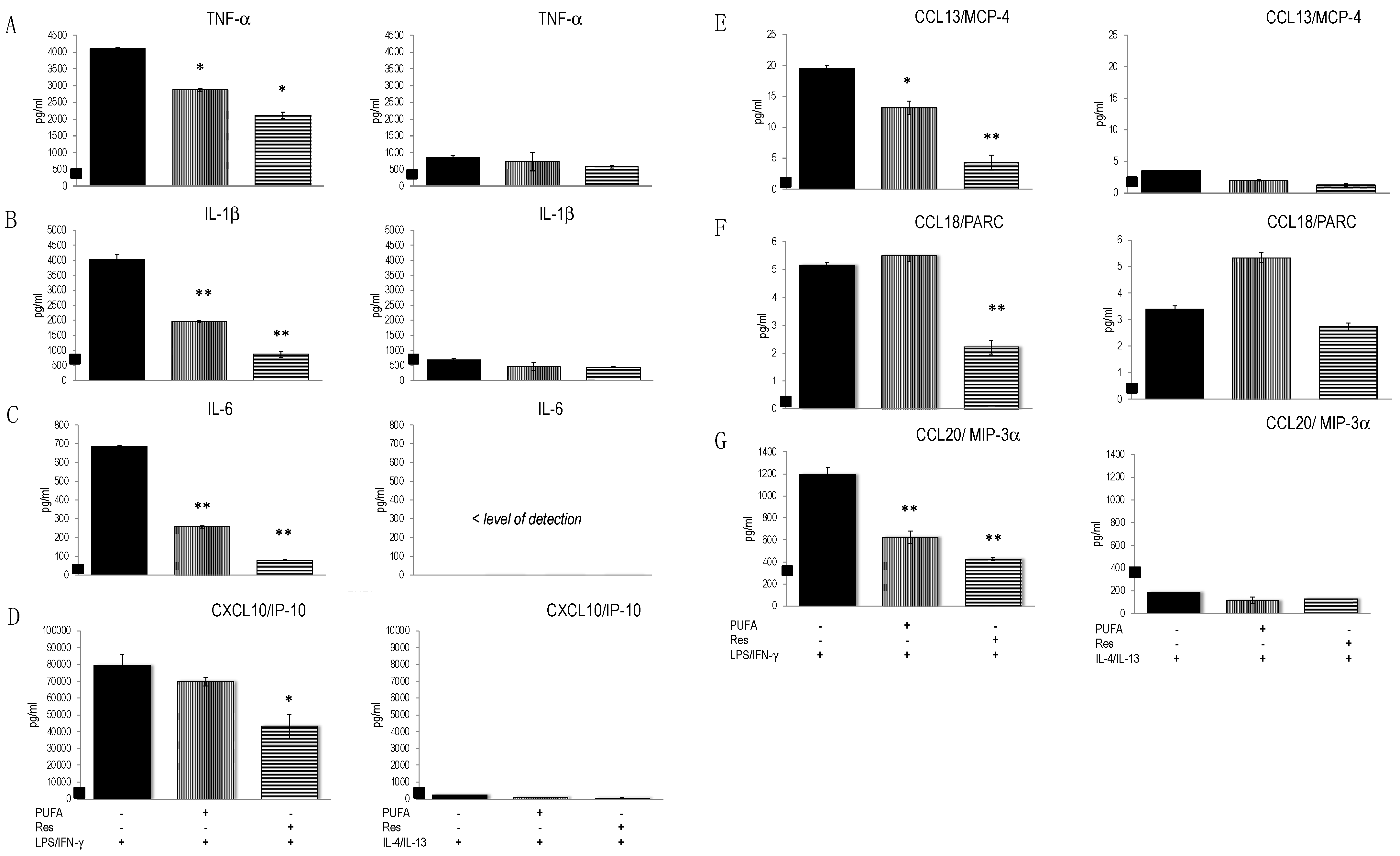
3.4. Res and ω-3 PUFAs Modulated the Secretion Pattern of PBMC-Derived M1 and M2 Macrophages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gordon, S.; Pluddemann, A.; Martinez Estrada, F. Macrophage heterogeneity in tissues: Phenotypic diversity and functions. Immunol. Rev. 2014, 262, 36–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.; Taylor, P.R. Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Fleetwood, A.J.; Dinh, H.; Cook, A.D.; Hertzog, P.J.; Hamilton, J.A. GM-CSF- and M-CSF-dependent macrophage phenotypes display differential dependence on type I interferon signaling. J. Leukoc. Biol. 2009, 86, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Fleetwood, A.J.; Lawrence, T.; Hamilton, J.A.; Cook, A.D. Granulocyte-macrophage colony-stimulating factor (CSF) and macrophage CSF-dependent macrophage phenotypes display differences in cytokine profiles and transcription factor activities: Implications for CSF blockade in inflammation. J. Immunol. 2007, 178, 5245–5252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra-Filardi, E.; Vega, M.A.; Sanchez-Mateos, P.; Corbi, A.L.; Puig-Kroger, A. Heme Oxygenase-1 expression in M-CSF-polarized M2 macrophages contributes to LPS-induced IL-10 release. Immunobiology 2010, 215, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Verreck, F.A.; de Boer, T.; Langenberg, D.M.; Hoeve, M.A.; Kramer, M.; Vaisberg, E.; Kastelein, R.; Kolk, A.; de Waal-Malefyt, R.; Ottenhoff, T.H. Human IL-23-producing type 1 macrophages promote but IL-10-producing type 2 macrophages subvert immunity to (myco)bacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 4560–4565. [Google Scholar] [CrossRef] [Green Version]
- Verreck, F.A.; de Boer, T.; Langenberg, D.M.; van der Zanden, L.; Ottenhoff, T.H. Phenotypic and functional profiling of human proinflammatory type-1 and anti-inflammatory type-2 macrophages in response to microbial antigens and IFN-gamma- and CD40L-mediated costimulation. J. Leukoc. Biol. 2006, 79, 285–293. [Google Scholar] [CrossRef]
- Izquierdo, E.; Cuevas, V.D.; Fernandez-Arroyo, S.; Riera-Borrull, M.; Orta-Zavalza, E.; Joven, J.; Rial, E.; Corbi, A.L.; Escribese, M.M. Reshaping of Human Macrophage Polarization through Modulation of Glucose Catabolic Pathways. J. Immunol. 2015, 195, 2442–2451. [Google Scholar] [CrossRef] [Green Version]
- Mia, S.; Warnecke, A.; Zhang, X.M.; Malmstrom, V.; Harris, R.A. An optimized protocol for human M2 macrophages using M-CSF and IL-4/IL-10/TGF-beta yields a dominant immunosuppressive phenotype. Scand. J. Immunol. 2014, 79, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.K.; Mantovani, A. Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat. Immunol. 2010, 11, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A.; Nguyen, K.D.; Goh, Y.P. Macrophage-mediated inflammation in metabolic disease. Nat. Rev. Immunol. 2011, 11, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locati, M.; Mantovani, A.; Sica, A. Macrophage activation and polarization as an adaptive component of innate immunity. Adv. Immunol. 2013, 120, 163–184. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Keshav, S.; Harris, N.; Gordon, S. Interleukin 4 potently enhances murine macrophage mannose receptor activity: A marker of alternative immunologic macrophage activation. J. Exp. Med. 1992, 176, 287–292. [Google Scholar] [CrossRef]
- Akagawa, K.S.; Komuro, I.; Kanazawa, H.; Yamazaki, T.; Mochida, K.; Kishi, F. Functional heterogeneity of colony-stimulating factor-induced human monocyte-derived macrophages. Respirology 2006, 11, S32–S36. [Google Scholar] [CrossRef]
- Ambarus, C.A.; Krausz, S.; van Eijk, M.; Hamann, J.; Radstake, T.R.; Reedquist, K.A.; Tak, P.P.; Baeten, D.L. Systematic validation of specific phenotypic markers for in vitro polarized human macrophages. J. Immunol. Methods 2012, 375, 196–206. [Google Scholar] [CrossRef]
- Jaguin, M.; Houlbert, N.; Fardel, O.; Lecureur, V. Polarization profiles of human M-CSF-generated macrophages and comparison of M1-markers in classically activated macrophages from GM-CSF and M-CSF origin. Cell. Immunol. 2013, 281, 51–61. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S.; Locati, M.; Mantovani, A. Transcriptional profiling of the human monocyte-to-macrophage differentiation and polarization: New molecules and patterns of gene expression. J. Immunol. 2006, 177, 7303–7311. [Google Scholar] [CrossRef] [Green Version]
- Auwerx, J. The human leukemia cell line, THP-1: A multifacetted model for the study of monocyte-macrophage differentiation. Experientia 1991, 47, 22–31. [Google Scholar] [CrossRef]
- Daigneault, M.; Preston, J.A.; Marriott, H.M.; Whyte, M.K.; Dockrell, D.H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE 2010, 5, e8668. [Google Scholar] [CrossRef]
- Feng, Y.H.; Zhu, Y.N.; Liu, J.; Ren, Y.X.; Xu, J.Y.; Yang, Y.F.; Li, X.Y.; Zou, J.P. Differential regulation of resveratrol on lipopolysacchride-stimulated human macrophages with or without IFN-gamma pre-priming. Int. Immunopharmacol. 2004, 4, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.; Eklund, A.; Patarroyo, M. Regulated expression of integrins and other adhesion molecules during differentiation of monocytes into macrophages. Cell. Immunol. 1994, 156, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Hikiji, H.; Okinaga, T.; Hashidate-Yoshida, T.; Shindou, H.; Ariyoshi, W.; Shimizu, T.; Tominaga, K.; Nishihara, T. Essential Role of Lysophosphatidylcholine Acyltransferase 3 in the Induction of Macrophage Polarization in PMA-Treated U937 Cells. J. Cell. Biochem. 2015, 116, 2840–2848. [Google Scholar] [CrossRef]
- Berges, C.; Naujokat, C.; Tinapp, S.; Wieczorek, H.; Hoh, A.; Sadeghi, M.; Opelz, G.; Daniel, V. A cell line model for the differentiation of human dendritic cells. Biochem. Biophys. Res. Commun. 2005, 333, 896–907. [Google Scholar] [CrossRef]
- Buchacher, T.; Ohradanova-Repic, A.; Stockinger, H.; Fischer, M.B.; Weber, V. M2 Polarization of Human Macrophages Favors Survival of the Intracellular Pathogen Chlamydia pneumoniae. PLoS ONE 2015, 10, e0143593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNelis, J.C.; Olefsky, J.M. Macrophages, immunity, and metabolic disease. Immunity 2014, 41, 36–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, J.L.; Mohamadzadeh, M. Macrophages and chemokines as mediators of angiogenesis. Front. Physiol. 2013, 4, 159. [Google Scholar] [CrossRef] [Green Version]
- Pallares, V.; Calay, D.; Cedo, L.; Castell-Auvi, A.; Raes, M.; Pinent, M.; Ardevol, A.; Arola, L.; Blay, M. Enhanced anti-inflammatory effect of resveratrol and EPA in treated endotoxin-activated RAW 264.7 macrophages. Br. J. Nutr. 2012, 108, 1562–1573. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Bhattacharya, A.; Fernandes, G. Docosahexaenoic acid is more potent inhibitor of osteoclast differentiation in RAW 264.7 cells than eicosapentaenoic acid. J. Cell. Physiol. 2008, 214, 201–209. [Google Scholar] [CrossRef]
- Wang, S.; Wu, D.; Lamon-Fava, S.; Matthan, N.R.; Honda, K.L.; Lichtenstein, A.H. In vitro fatty acid enrichment of macrophages alters inflammatory response and net cholesterol accumulation. Br. J. Nutr. 2009, 102, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Weldon, S.M.; Mullen, A.C.; Loscher, C.E.; Hurley, L.A.; Roche, H.M. Docosahexaenoic acid induces an anti-inflammatory profile in lipopolysaccharide-stimulated human THP-1 macrophages more effectively than eicosapentaenoic acid. J. Nutr. Biochem. 2007, 18, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Im, D.S. Functions of omega-3 fatty acids and FFA4 (GPR120) in macrophages. Eur. J. Pharmacol. 2015, 785, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Raptis, D.A.; Limani, P.; Jang, J.H.; Ungethum, U.; Tschuor, C.; Graf, R.; Humar, B.; Clavien, P.A. GPR120 on Kupffer cells mediates hepatoprotective effects of omega3-fatty acids. J. Hepatol. 2014, 60, 625–632. [Google Scholar] [CrossRef] [PubMed]
- De Boer, A.A.; Monk, J.M.; Robinson, L.E. Docosahexaenoic acid decreases pro-inflammatory mediators in an in vitro murine adipocyte macrophage co-culture model. PLoS ONE 2014, 9, e85037. [Google Scholar] [CrossRef] [Green Version]
- Gladine, C.; Zmojdzian, M.; Joumard-Cubizolles, L.; Verny, M.A.; Comte, B.; Mazur, A. The omega-3 fatty acid docosahexaenoic acid favorably modulates the inflammatory pathways and macrophage polarization within aorta of LDLR(−/−) mice. Genes Nutr. 2014, 9, 424. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Kuan, Y.H.; Li, J.R.; Chen, W.Y.; Ou, Y.C.; Pan, H.C.; Liao, S.L.; Raung, S.L.; Chang, C.J.; Chen, C.J. Docosahexaenoic acid reduces cellular inflammatory response following permanent focal cerebral ischemia in rats. J. Nutr. Biochem. 2013, 24, 2127–2137. [Google Scholar] [CrossRef]
- Honda, K.L.; Lamon-Fava, S.; Matthan, N.R.; Wu, D.; Lichtenstein, A.H. Docosahexaenoic acid differentially affects TNFalpha and IL-6 expression in LPS-stimulated RAW 264.7 murine macrophages. Prostaglandins Leukot. Essent. Fat. Acids 2015, 97, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Ye, J.; Gao, Z.; Youn, H.S.; Lee, W.H.; Zhao, L.; Sizemore, N.; Hwang, D.H. Reciprocal modulation of Toll-like receptor-4 signaling pathways involving MyD88 and phosphatidylinositol 3-kinase/AKT by saturated and polyunsaturated fatty acids. J. Biol. Chem. 2003, 278, 37041–37051. [Google Scholar] [CrossRef] [Green Version]
- Mullen, A.; Loscher, C.E.; Roche, H.M. Anti-inflammatory effects of EPA and DHA are dependent upon time and dose-response elements associated with LPS stimulation in THP-1-derived macrophages. J. Nutr. Biochem. 2010, 21, 444–450. [Google Scholar] [CrossRef]
- Schwager, J.; Richard, N.; Widmer, F.; Raederstorff, D. Resveratrol distinctively modulates the inflammatory profiles of immune and endothelial cells. BMC Complement. Altern. Med. 2017, 17, 309. [Google Scholar] [CrossRef] [Green Version]
- Wadsworth, T.L.; Koop, D.R. Effects of the wine polyphenolics quercetin and resveratrol on pro-inflammatory cytokine expression in RAW 264.7 macrophages. Biochem. Pharmacol. 1999, 57, 941–949. [Google Scholar] [CrossRef]
- Wan, J.; Benkdane, M.; Teixeira-Clerc, F.; Bonnafous, S.; Louvet, A.; Lafdil, F.; Pecker, F.; Tran, A.; Gual, P.; Mallat, A.; et al. M2 Kupffer cells promote M1 Kupffer cell apoptosis: A protective mechanism against alcoholic and nonalcoholic fatty liver disease. Hepatology 2014, 59, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Schwager, J.; Seifert, N.; Bompard, A.; Raederstorff, D.; Bendik, I. Resveratrol, EGCG and Vitamins Modulate Activated T Lymphocytes. Molecules 2021, 26, 5600. [Google Scholar] [CrossRef]
- Krausgruber, T.; Blazek, K.; Smallie, T.; Alzabin, S.; Lockstone, H.; Sahgal, N.; Hussell, T.; Feldmann, M.; Udalova, I.A. IRF5 promotes inflammatory macrophage polarization and TH1-TH17 responses. Nat. Immunol. 2011, 12, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Schwager, J.; Richard, N.; Mussler, B.; Raederstorff, D. Tomato Aqueous Extract Modulates the Inflammatory Profile of Immune Cells and Endothelial Cells. Molecules 2016, 21, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwager, J.; Bompard, A.; Weber, P.; Raederstorff, D. Ascorbic acid modulates cell migration in differentiated HL-60 cells and peripheral blood leukocytes. Mol. Nutr. Food Res. 2015, 59, 1513–1523. [Google Scholar] [CrossRef]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E., Jr.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [Green Version]
- Yurko-Mauro, K.; Kralovec, J.; Bailey-Hall, E.; Smeberg, V.; Stark, J.G.; Salem, N., Jr. Similar eicosapentaenoic acid and docosahexaenoic acid plasma levels achieved with fish oil or krill oil in a randomized double-blind four-week bioavailability study. Lipids Health Dis. 2015, 14, 99. [Google Scholar] [CrossRef] [Green Version]
- Chanput, W.; Mes, J.J.; Savelkoul, H.F.; Wichers, H.J. Characterization of polarized THP-1 macrophages and polarizing ability of LPS and food compounds. Food Funct. 2013, 4, 266–276. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Oh, Y.C.; Kang, O.H.; Choi, J.G.; Chae, H.S.; Lee, Y.S.; Brice, O.O.; Jung, H.J.; Hong, S.H.; Lee, Y.M.; Kwon, D.Y. Anti-inflammatory effect of resveratrol by inhibition of IL-8 production in LPS-induced THP-1 cells. Am. J. Chin. Med. 2009, 37, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.H.; Lin-Shiau, S.Y.; Lin, J.K. Suppression of nitric oxide synthase and the down-regulation of the activation of NFkappaB in macrophages by resveratrol. Br. J. Pharmacol. 1999, 126, 673–680. [Google Scholar] [CrossRef] [Green Version]
- De la Lastra, C.A.; Villegas, I. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res. 2005, 49, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Richard, N.; Porath, D.; Radspieler, A.; Schwager, J. Effects of resveratrol, piceatannol, tri-acetoxystilbene, and genistein on the inflammatory response of human peripheral blood leukocytes. Mol. Nutr. Food Res. 2005, 49, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Shabani, M.; Sadeghi, A.; Hosseini, H.; Teimouri, M.; Babaei Khorzoughi, R.; Pasalar, P.; Meshkani, R. Resveratrol alleviates obesity-induced skeletal muscle inflammation via decreasing M1 macrophage polarization and increasing the regulatory T cell population. Sci. Rep. 2020, 10, 3791. [Google Scholar] [CrossRef] [PubMed]
- Buttari, B.; Profumo, E.; Segoni, L.; D’Arcangelo, D.; Rossi, S.; Facchiano, F.; Saso, L.; Businaro, R.; Iuliano, L.; Rigano, R. Resveratrol counteracts inflammation in human M1 and M2 macrophages upon challenge with 7-oxo-cholesterol: Potential therapeutic implications in atherosclerosis. Oxid. Med. Cell. Longev. 2014, 2014, 257543. [Google Scholar] [CrossRef]
- Calder, P.C. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505s–1519s. [Google Scholar] [CrossRef]
- Chang, H.Y.; Lee, H.N.; Kim, W.; Surh, Y.J. Docosahexaenoic acid induces M2 macrophage polarization through peroxisome proliferator-activated receptor gamma activation. Life Sci 2015, 120, 39–47. [Google Scholar] [CrossRef]
- Kawano, A.; Ariyoshi, W.; Yoshioka, Y.; Hikiji, H.; Nishihara, T.; Okinaga, T. Docosahexaenoic acid enhances M2 macrophage polarization via the p38 signaling pathway and autophagy. J. Cell. Biochem. 2019, 120, 12604–12617. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surface Determinant | Macrophage Subtype |
|---|---|
| CD11b (integrin αM) | M2 |
| CD11c (integrin αX) | M2 |
| CD14 (LPS receptor) | M1 |
| CD80 (B7-1) | M1 |
| CD206 (macrophage mannose receptor) | M2 |
| CD209 (DC-SIGN) | M2 |
| CD274 (B7-H1) | M1 |
| CCR2 (CC chemokine receptor 2) | M1 |
| CCR7 (CC chemokine receptor 7) | M1 |
| HLA-DR (MHC class II) | M2 |
| Cytokines/Chemokines | Macrophage Subtype |
| IL-1β | M1 |
| IL-6 | M1 |
| TNF-α | M1 |
| IL-10 | M2 |
| IL-12p70 | M1 |
| CCL13/MCP4 | M2 |
| CCL18/PARC | M2 |
| CCL20/MIP-3α | M2 |
| CXCL10/IP-10 | M1 |
| CXCL1/GRO-α | M2 |
| Polarization | Activation | TNF-α | IL-1β | IL-6 | IL-12p70 | CCL13/MCP-4 | CCL18/PARC | CCL20/MIP-3α | CXCL1/GROα | CXCL10/IP-10 |
|---|---|---|---|---|---|---|---|---|---|---|
| 6 days | 24 h | |||||||||
| Medium | None | 490 ± 20 | 91 ± 2 | 0.0 ± 0.0 | 0.0 ± 0.0 | 1.0 ± 0.0 | 4.0 ± 0.0 | 77,633 ± 7145 | 1.5 ± 0.3 | 0.0 ± 0.0 |
| GM-CSF | None | 1737 ± 203 | 374 ± 34 | 0.0 ± 0.0 | 0.0 ± 0.0 | 11 ± 1 | 5.0 ± 1.0 | 131,667 ± 10,017 | 10.7 ± 1.4 | 57 ± 7 |
| M-CSF | None | 628 ± 54 | 115±11 | 0.0 ± 0.0 | 0.0 ± 0.0 | 2.0 ± 0.0 | 3.0 ± 0.0 | 104,333 ± 4726 | 1.9 ± 0.2 | 0.0 ± 0.0 |
| Medium | IL-4/IL13 | 252 ± 36 | 68 ± 2 | 0.0 ± 0.0 | 0.0 ± 0.0 | 304 ± 56 | 11 ± 1 | 61,933 ± 11,754 | 304.3 ± 56.2 | 61 ± 0 |
| GM-CSF | IL-4/IL13 | 1111 ± 146 | 378 ± 66 | 0.0 ± 0.0 | 0.0 ± 0.0 | 331 ± 52 | 15 ± 3 | 105,000 ± 2000 | 331.3 ± 52.0 | 81 ± 10 |
| M-CSF | IL-4/IL13 | 371 ± 24 | 98 ± 8 | 0.0 ± 0.0 | 0.0 ± 0.0 | 430 ± 14 | 11 ± 1 | 85,233 ± 5661 | 430.0 ± 13.7 | 66 ± 4 |
| Medium | LPS/IFN-γ | 25,533 ± 404 | 8140 ± 433 | 11,133 ± 551 | 9.0 ± 2.7 | 18 ± 1 | 15 ± 0 | 194,667 ± 11,846 | 17.6 ± 1.4 | 90,300 ± 15,455 |
| GM-CSF | LPS/IFN-γ | 33,567 ± 1872 | 11,800 ± 520 | 5730 ± 429 | 8.3 ± 1.4 | 24 ± 1 | 30 ± 3 | 279,000 ± 13,856 | 23.7 ± 1.0 | 81,500 ± 7892 |
| M-CSF | LPS/IFN-γ | 33,400 ± 781 | 9420 ± 243 | 11,467 ± 208 | 16.0 ± 9.3 | 20 ± 1 | 15 ± 1 | 236,333 ± 4163 | 19.9 ± 1.4 | 98,800 ± 15,029 |
| M-CSF pol. | M1/M2 ratio | 101 | 119 | >5000 * | > 16 | 0.1 | 1.4 | 3.1 | 0.05 | 1480 |
| GM-CSF pol. | M1/M2 ratio | 30 | 31 | >10,000 * | > 8 | 0.7 | 0.2 | 2.8 | 0.01 | 1006 |
| Activation | Substances | TNF-α | IL-1β | IL-6 | IL-12p70 | CCL13/MCP-4 | CCL18/PARC | CCL20/MIP-3α | CXCL1/GRO-α | CXCL10/IP-10 |
|---|---|---|---|---|---|---|---|---|---|---|
| None | None | 1443 ± 76 | 1410 ± 62 | 19.1 ± 1.9 | 27.9 ± 3.9 | 4.5 ± 0.4 | 20.9 ± 0.6 | 24,333 ± 737 | 5827 ± 339 | 197.7 ± 13.3 |
| LPS/IFN-γ | None | 22,100 ± 954 | 22,433 ± 1274 | 8740 ± 467 | 81.4 ± 16.3 | 218.5 ± 0.7 | 104.5 ± 2.1 | 38,867 ± 4654 | 33,400 ± 9617 | >10,000 |
| IL-4/IL-13 | None | 964 ± 150 | 5363 ± 3058 | 13.1 ± 12.6 | 125.5 ± 23.3 | 143.5 ± 0.7 | 27.6 ± 2.4 | 25,750 ± 6576 | 2510 ± 1994 | 230.0 ± 50.9 |
| M1/M2 Ratio | None | 23 | 4.2 | 667 | 0.6 | 1.5 | 3.7 | 1.5 | 13 | >43 * |
| None | Res | 718 ± 87 | 1247 ± 125 | 12.8 ± 1.1 | 115.5 ± 9.2 | 3.6 ± 0.6 | 24.1 ± 3.4 | 21,900 ± 2088 | 3297 ± 197 | 203.3 ± 9.5 |
| None | PUFAs | 268 ± 33 | 388 ± 67 | 2.9 ± 0.4 | 15.1 ± 2.4 | 2.6 ± 0.4 | 17.6 ± 1.2 | 6823 ± 1508 | 2697 ± 390 | 88.2 ± 6.5 |
| Ratio untreat./Res | 2.0 | 1.1 | 1.5 | 0.2 | 1.3 | 0.9 | 1.1 | 1.8 | 1.0 | |
| Ratio untreat./PUFA | 5.4 | 3.6 | 6.3 | 1.9 | 1.7 | 1.2 | 3.6 | 2.2 | 2.2 |
| PBMC | TNF-α | IL-1β | IL-6 | IL-10 | IL-12p70 | CXCL10/IP-10 | CCL13/MCP-4 | CCL18/PARC | CCL20/MIP-3α | CXCL1/GRO-α |
|---|---|---|---|---|---|---|---|---|---|---|
| M0 | 908 ± 38 | 2573 ± 337 | 28,133 ± 3066 | 5.6 ± 0.4 | 21.3 ± 3.4 | 126.3 ± 15.5 | 424 ± 158 | 16,966 ± 1464 | 490 ± 69 | 93,733 ± 16,815 |
| M1-polarized | 920 ± 13 | 1370 ± 140 | 25,433 ± 1357 | 4.6 ± 0.5 | 22.5 ± 5.2 | 65.2 ± 6.3 | 2023 ± 538 | 7563 ± 792 | 315 ± 39 | 90,000 ± 13,928 |
| M2-polarized | 886 ± 83 | 1550 ± 87 | 24,000 ± 1833 | 6.1 ± 0.6 | 27.1 ± 10.1 | 129.0 ± 4.0 | 1044 ± 115 | 12,000 ± 1135 | 455 ± 36 | 78,633 ± 8467 |
| M1/M2 ratio | 1.04 | 0.88 | 1.06 | 0.76 | 0.83 | 0.51 | 1.94 | 0.63 | 0.69 | 1.14 |
| M1 polarized and activated | 2065 ± 275 | 5045 ± 77 | 49,000 ± 9475 | 6.0 ± 1.4 | 19 ± 5 | 63 ± 32 | 272 ± 8 | 1915 ± 233 | 545 ± 172 | 30,450 ± 9828 |
| M2 polarized and activated | 1790 ± 141 | 5100 ± 735 | 50,600 ± 11,313 | 10.0 ± 2.2 | 6 ± 2 | 130 ± 30 | 908 ± 54 | 217 ± 4 | 525± 118 | 60,700 ± 14,283 |
| M1/M2 ratio | 1.15 | 1.0 | 1.0 | 0.6 | 3.0 | 0.5 | 0.3 | 8.8 | 1.0 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwager, J.; Bompard, A.; Raederstorff, D.; Hug, H.; Bendik, I. Resveratrol and ω-3 PUFAs Promote Human Macrophage Differentiation and Function. Biomedicines 2022, 10, 1524. https://doi.org/10.3390/biomedicines10071524
Schwager J, Bompard A, Raederstorff D, Hug H, Bendik I. Resveratrol and ω-3 PUFAs Promote Human Macrophage Differentiation and Function. Biomedicines. 2022; 10(7):1524. https://doi.org/10.3390/biomedicines10071524
Chicago/Turabian StyleSchwager, Joseph, Albine Bompard, Daniel Raederstorff, Hubert Hug, and Igor Bendik. 2022. "Resveratrol and ω-3 PUFAs Promote Human Macrophage Differentiation and Function" Biomedicines 10, no. 7: 1524. https://doi.org/10.3390/biomedicines10071524