
Thioredoxin Domain Containing 5 Suppression Elicits Serum Amyloid A-Containing High-Density Lipoproteins

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
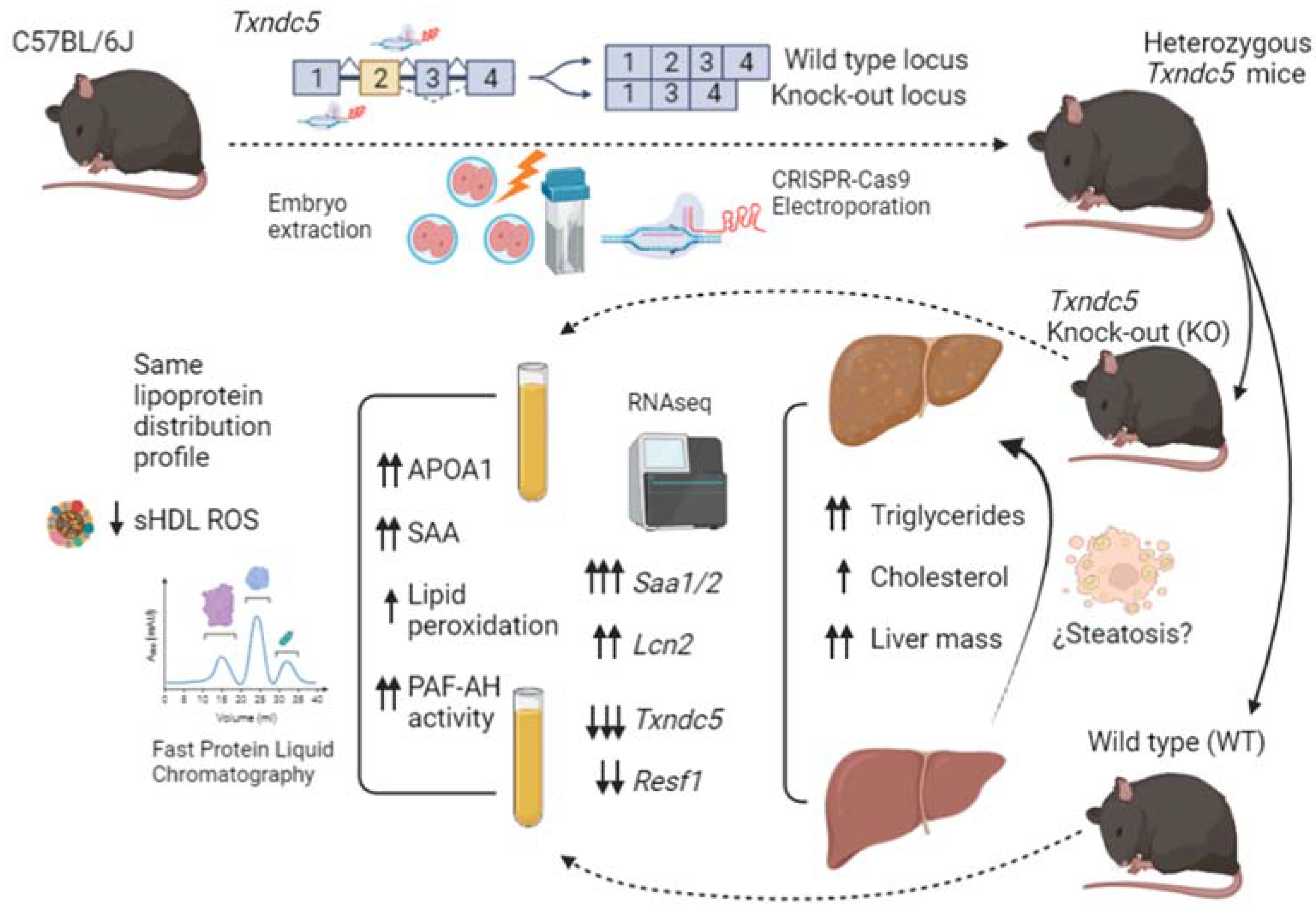
2.1. Generation of Txndc5-Deficient Mice Using CRISPR/hifiCas9 Genome-Editing
2.2. Animals and Diets
2.3. RNA Extraction
2.4. RNAseq Analysis
2.5. Quantification of mRNA and cDNA Synthesis
2.6. Western Blotting
2.7. Histological Analysis
2.8. Plasma Determinations and Liver Lipids
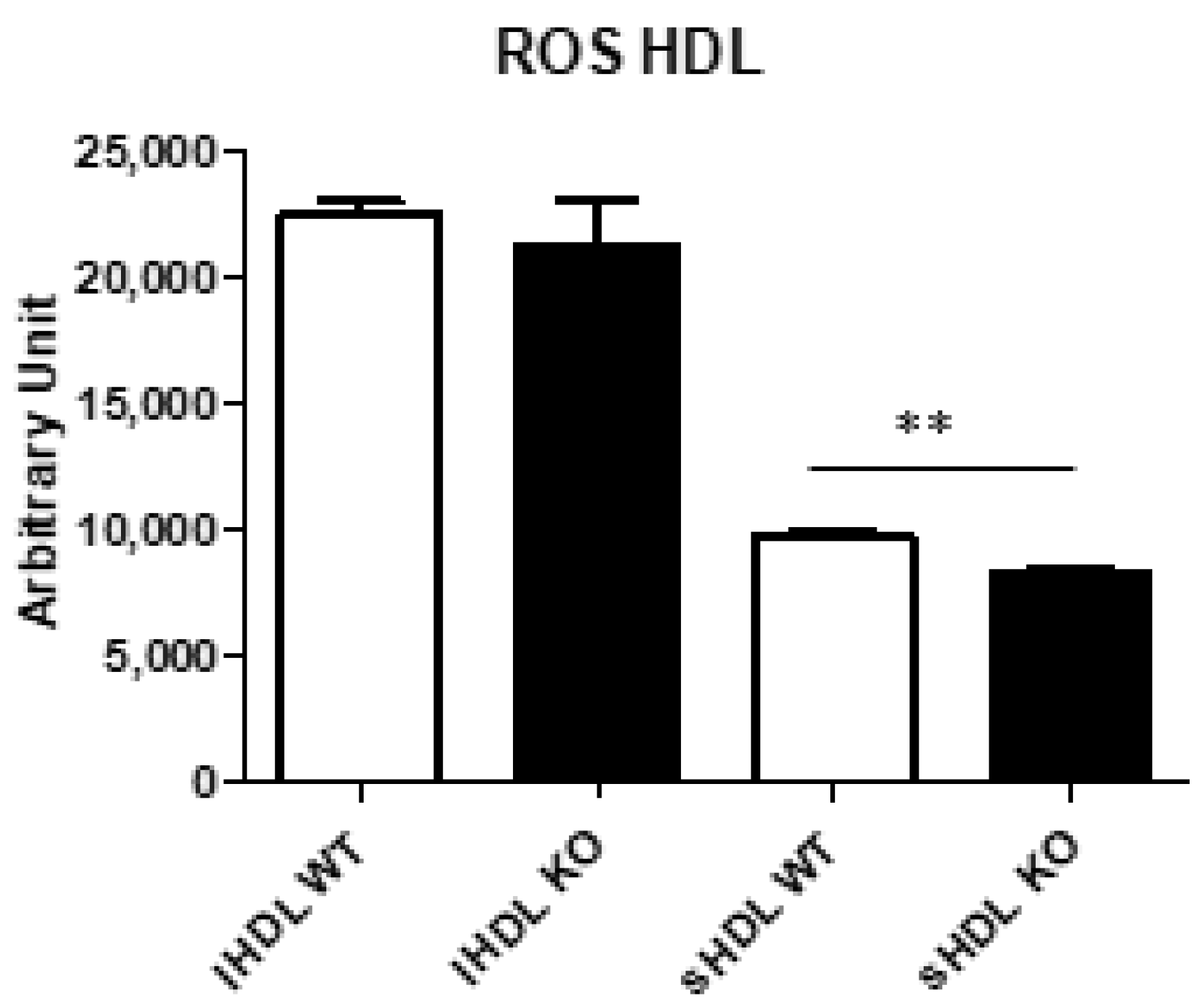
2.9. Reactive Oxygen Species (ROS) Content in Lipoproteins
2.10. Statistics
3. Results
3.1. Generation of Txndc5-Deficient Mice
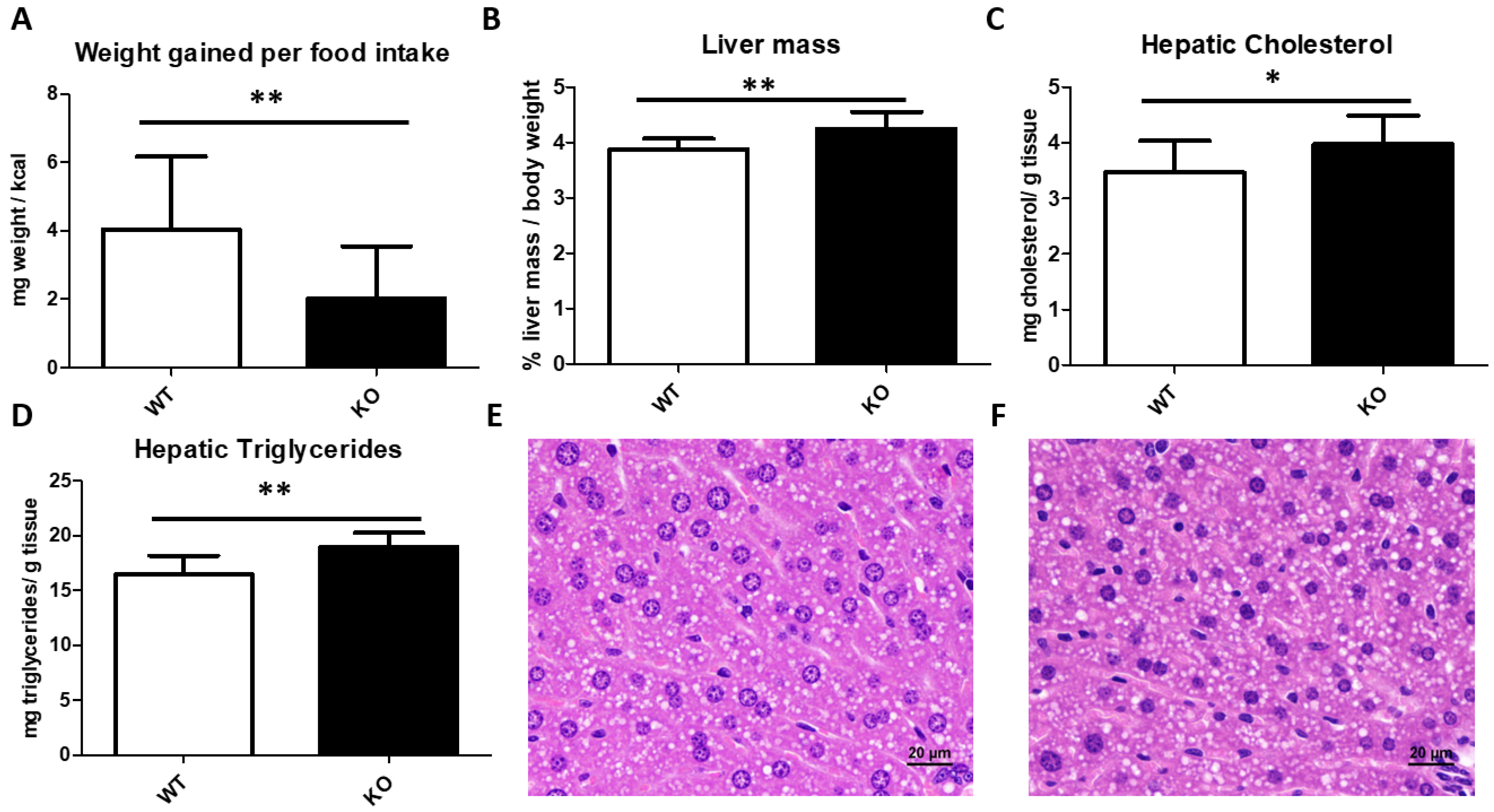
3.2. Somatometric Parameters
3.3. Liver Histological Analyses and Hepatic Lipid Content
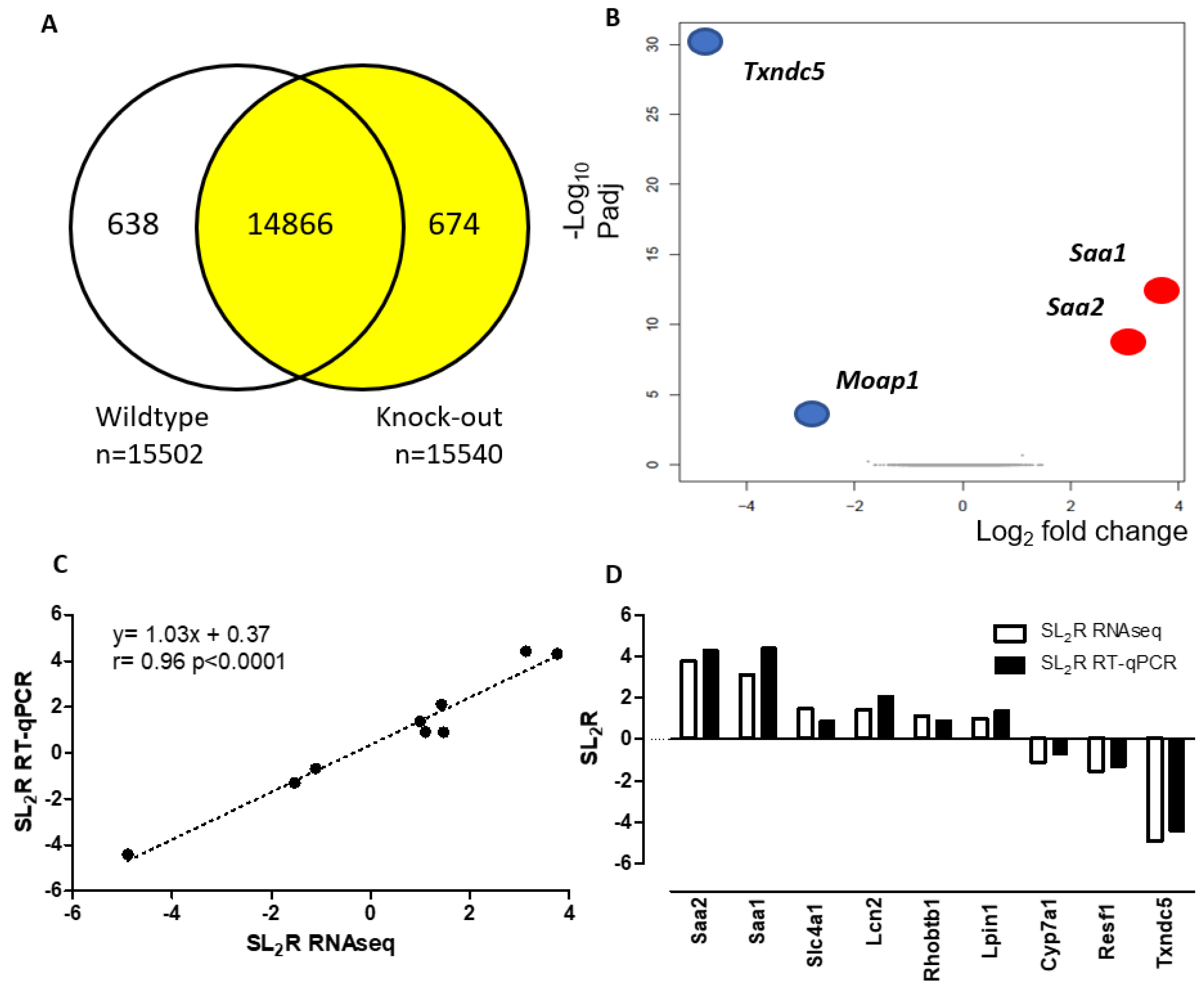
3.4. Hepatic Gene Expression of Txndc5-Deficient Male Mice Fed for 4 Weeks on a Chow Diet
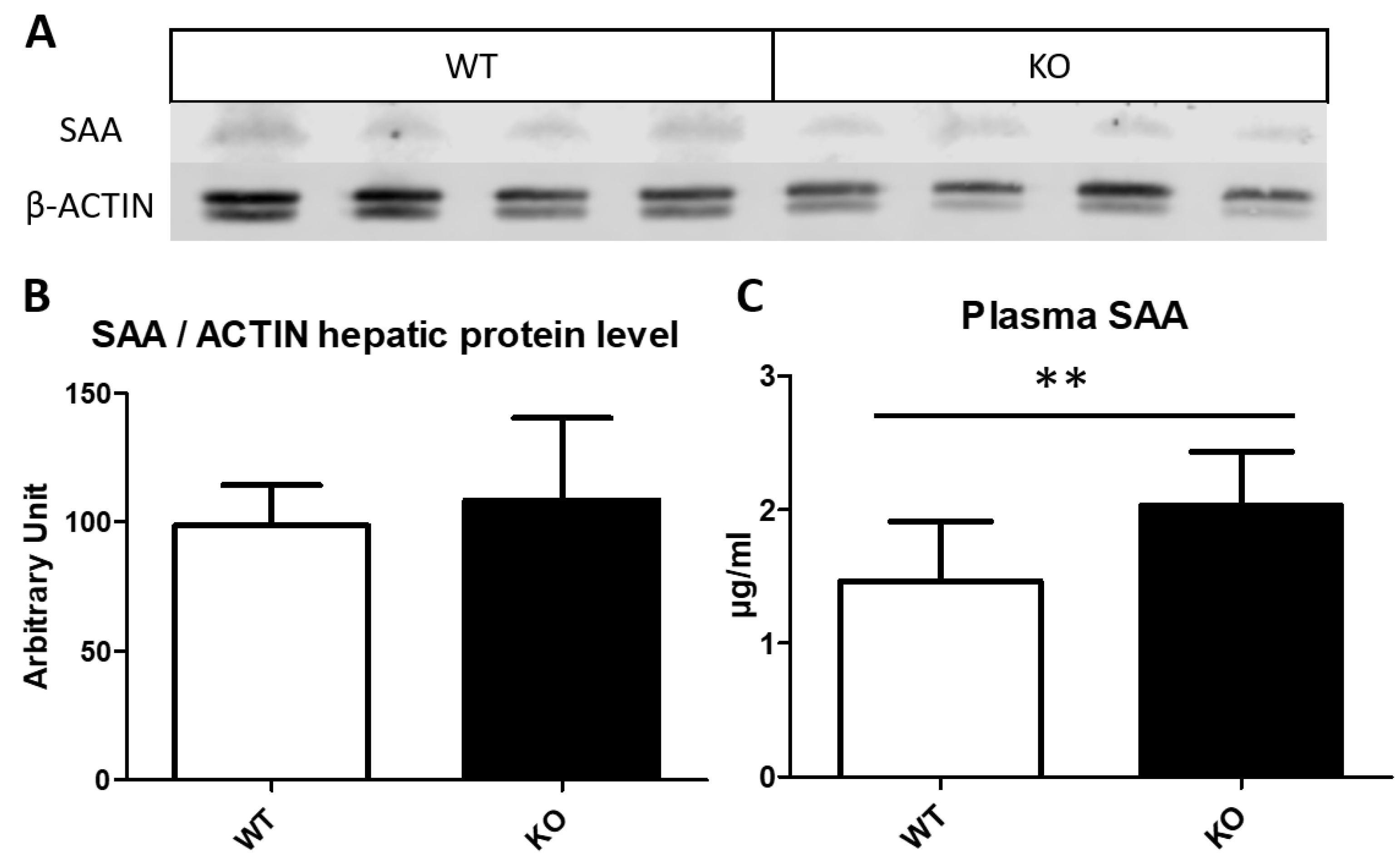
3.5. Serum Amyloid Content on Liver and Plasma
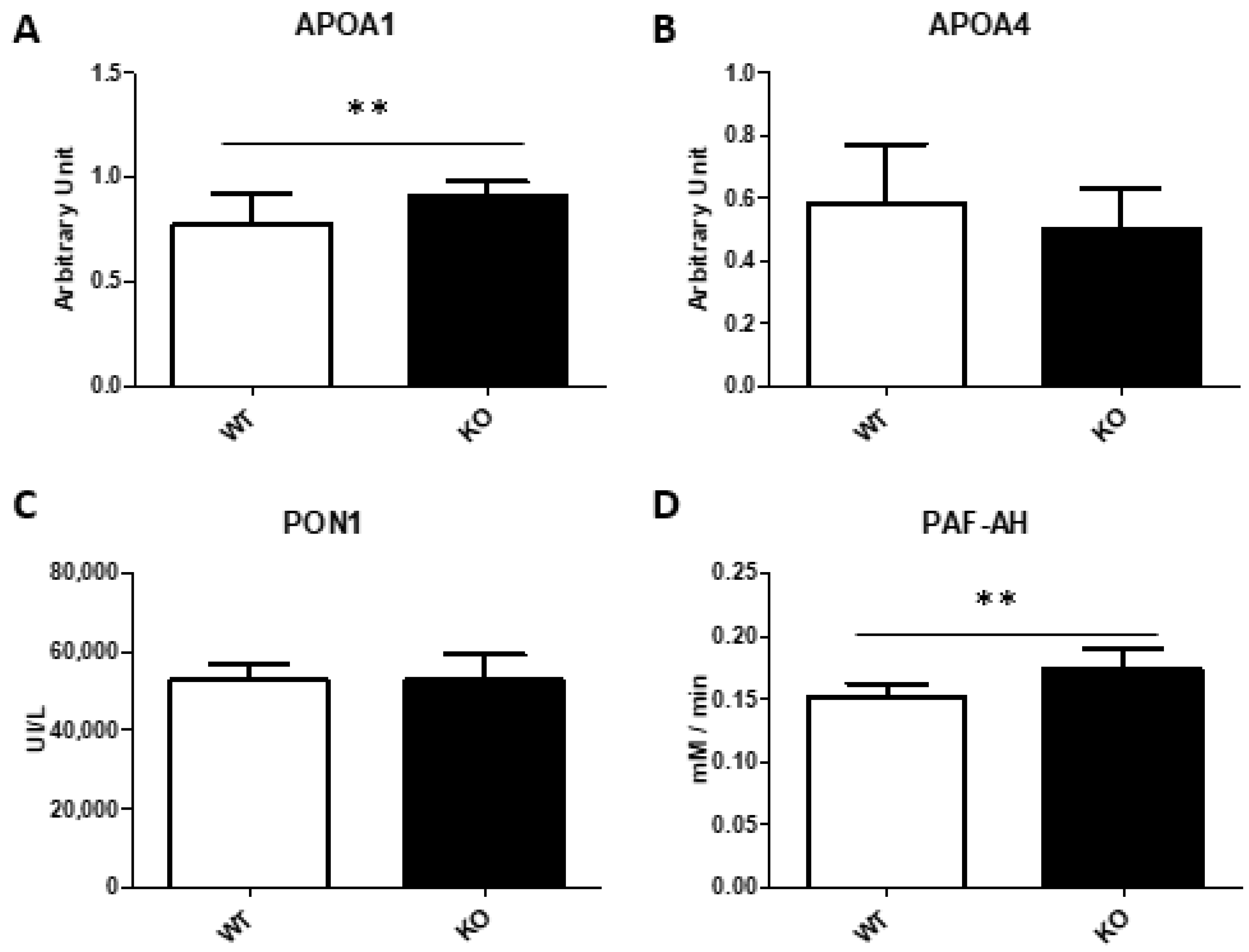
3.6. Plasma Parameters Determinations
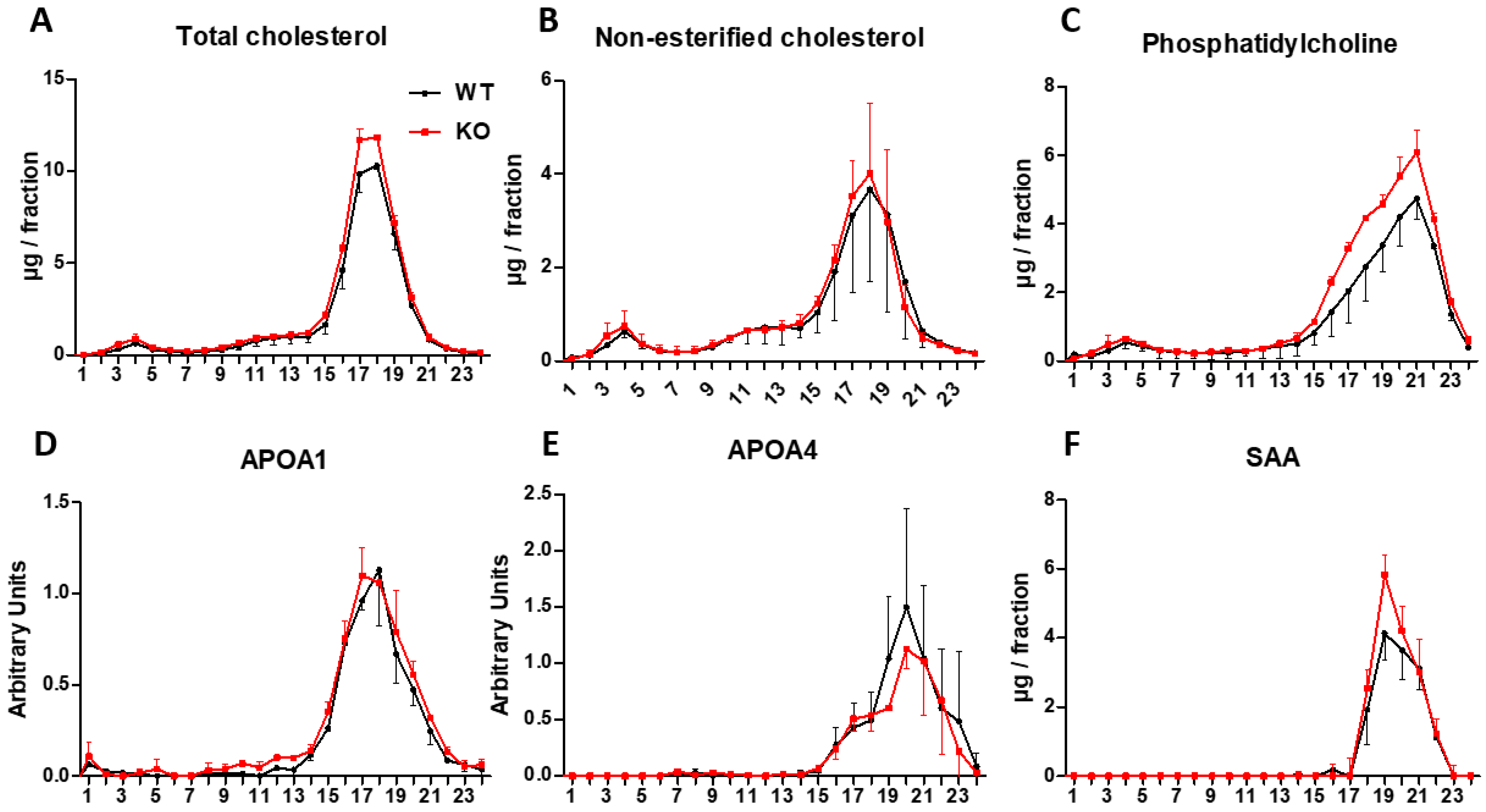
3.7. Analysis of Lipoproteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A Revolutionary Tool for Transcriptomics. Nat. Rev. Genet. 2009, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A Draft Map of the Human Proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flicek, P.; Amode, M.R.; Barrell, D.; Beal, K.; Brent, S.; Carvalho-Silva, D.; Clapham, P.; Coates, G.; Fairley, S.; Fitzgerald, S.; et al. Ensembl 2012. Nucleic Acids Res. 2012, 40, D84–D90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, C.; Sinskey, A.J.; Lodish, H.F. Oxidized Redox State of Glutathione in the Endoplasmic Reticulum. Science 1992, 257, 1496–1502. [Google Scholar] [CrossRef]
- Okumura, M.; Kadokura, H.; Inaba, K. Structures and Functions of Protein Disulfide Isomerase Family Members Involved in Proteostasis in the Endoplasmic Reticulum. Free Radic. Biol. Med. 2015, 83, 314–322. [Google Scholar] [CrossRef]
- Saibil, H. Chaperone Machines for Protein Folding, Unfolding and Disaggregation. Nat. Rev. Mol. Cell Biol. 2013, 14, 630–642. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Inaba, K. Disulfide Bond Formation Network in the Three Biological Kingdoms, Bacteria, Fungi and Mammals. FEBS J. 2012, 279, 2262–2271. [Google Scholar] [CrossRef]
- Ramírez-Torres, A.; Barceló-Batllori, S.; Martínez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Arnal, C.; Guillén, N.; Acín, S.; Osada, J. Proteomics and Gene Expression Analyses of Squalene-Supplemented Mice Identify Microsomal Thioredoxin Domain-Containing Protein 5 Changes Associated with Hepatic Steatosis. J. Proteom. 2012, 77, 27–39. [Google Scholar] [CrossRef]
- Horna-Terrón, E.; Pradilla-Dieste, A.; Sánchez-de-Diego, C.; Osada, J. TXNDC5, a Newly Discovered Disulfide Isomerase with a Key Role in Cell Physiology and Pathology. Int. J. Mol. Sci. 2014, 15, 23501–23518. [Google Scholar] [CrossRef] [Green Version]
- Chawsheen, H.A.; Ying, Q.; Jiang, H.; Wei, Q. A Critical Role of the Thioredoxin Domain Containing Protein 5 (TXNDC5) in Redox Homeostasis and Cancer Development. Genes Dis. 2018, 5, 312–322. [Google Scholar] [CrossRef]
- Xu, B.; Li, J.; Liu, X.; Li, C.; Chang, X. TXNDC5 Is a Cervical Tumor Susceptibility Gene That Stimulates Cell Migration, Vasculogenic Mimicry and Angiogenesis by down-Regulating SERPINF1 and TRAF1 Expression. Oncotarget 2017, 8, 91009–91024. [Google Scholar] [CrossRef] [Green Version]
- Couasnay, G.; Bon, N.; Devignes, C.S.; Sourice, S.; Bianchi, A.; Véziers, J.; Weiss, P.; Elefteriou, F.; Provot, S.; Guicheux, J.; et al. PiT1/Slc20a1 Is Required for Endoplasmic Reticulum Homeostasis, Chondrocyte Survival, and Skeletal Development. J. Bone Miner. Res. 2019, 34, 387–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, H.; Li, Y.; Zhang, X.; Huang, G. Circ_0000517 Contributes to Hepatocellular Carcinoma Progression by Upregulating TXNDC5 via Sponging Mir-1296-5p. Cancer Manag. Res. 2020, 12, 3457–3468. [Google Scholar] [CrossRef]
- Wang, H.; Yang, X.; Guo, Y.; Shui, L.; Li, S.; Bai, Y.; Liu, Y.; Zeng, M.; Xia, J. HERG1 Promotes Esophageal Squamous Cell Carcinoma Growth and Metastasis through TXNDC5 by Activating the PI3K/AKT Pathway. J. Exp. Clin. Cancer Res. 2019, 38, 324. [Google Scholar] [CrossRef] [PubMed]
- Duivenvoorden, W.C.M.; Hopmans, S.N.; Austin, R.C.; Pinthus, J.H. Endoplasmic Reticulum Protein ERp46 in Prostate Adenocarcinoma. Oncol. Lett. 2017, 13, 3624–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedrick, E.; Mohankumar, K.; Lacey, A.; Safe, S. Inhibition of NR4A1 Promotes ROS Accumulation and IL24-Dependent Growth Arrest in Rhabdomyosarcoma. Mol. Cancer Res. 2019, 17, 2221–2232. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, E.; Lee, S.O.; Kim, G.; Abdelrahim, M.; Jin, U.H.; Safe, S.; Abudayyeh, A. Nuclear Receptor 4A1 (NR4A1) as a Drug Target for Renal Cell Adenocarcinoma. PLoS ONE 2015, 10, e0128308. [Google Scholar] [CrossRef]
- Mo, R.; Peng, J.; Xiao, J.; Ma, J.; Li, W.; Wang, J.; Ruan, Y.; Ma, S.; Hong, Y.; Wang, C.; et al. High TXNDC5 Expression Predicts Poor Prognosis in Renal Cell Carcinoma. Tumor Biol. 2016, 37, 9797–9806. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, L.; Li, N.; Sha, L.; Zhang, K. An Immunohistochemical Study of Thioredoxin Domain-Containing 5 Expression in Gastric Adenocarcinoma. Oncol. Lett. 2015, 9, 1154–1158. [Google Scholar] [CrossRef] [Green Version]
- Tan, F.; Zhu, H.; He, X.; Yu, N.; Zhang, X.; Xu, H.; Pei, H. Role of TXNDC5 in Tumorigenesis of Colorectal Cancer Cells: In Vivo and in Vitro Evidence. Int. J. Mol. Med. 2018, 42, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Ao, N.; Yang, J.; Wang, X.; Du, J. Glucagon-like Peptide-1 Preserves Non-Alcoholic Fatty Liver Disease through Inhibition of the Endoplasmic Reticulum Stress-Associated Pathway. Hepatol. Res. 2016, 46, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, Y.; Saito, M.; Kadokura, H.; Miyazaki, J.-I.; Tashiro, F.; Imagawa, Y.; Iwawaki, T.; Kohno, K. IRE1-XBP1 Pathway Regulates Oxidative Proinsulin Folding in Pancreatic β Cells. J. Cell Biol. 2018, 217, 1287–1301. [Google Scholar] [CrossRef]
- Li, J.; Xu, B.; Wu, C.; Yan, X.; Zhang, L.; Chang, X. TXNDC5 Contributes to Rheumatoid Arthritis by Down-Regulating IGFBP1 Expression. Clin. Exp. Immunol. 2018, 192, 82–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Li, J.; Wu, C.; Liu, C.; Yan, X.; Chang, X. CXCL10 and TRAIL Are Upregulated by TXNDC5 in Rheumatoid Arthritis Fibroblast-like Synoviocytes. J. Rheumatol. 2018, 45, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.; Fairus, S.; Zulfarina, M.S.; Mohamed, I.N. The Efficacy of Squalene in Cardiovascular Disease Risk-A Systematic Review. Nutrients 2020, 12, 414. [Google Scholar] [CrossRef] [Green Version]
- Guillén, N.; Acín, S.; Navarro, M.A.; Perona, J.S.; Arbonés-Mainar, J.M.; Arnal, C.; Sarría, A.J.; Surra, J.C.; Carnicer, R.; Orman, I.; et al. Squalene in a Sex-Dependent Manner Modulates Atherosclerotic Lesion Which Correlates with Hepatic Fat Content in ApoE-Knockout Male Mice. Atherosclerosis 2008, 197, 72–83. [Google Scholar] [CrossRef]
- Lou-Bonafonte, J.M.; Beamonte, R.M.; Sanclemente, T.; Surra, J.C.; Herrera-Marcos, L.V.; Sanchez-Marco, J.; Arnal, C.; Osada, J. Current Insights into the Biological Action of Squalene. Mol. Nutr. Food Res. 2018, 62, 1800136. [Google Scholar] [CrossRef]
- Fabbrini, E.; Magkos, F. Hepatic Steatosis as a Marker of Metabolic Dysfunction. Nutrients 2015, 7, 4995–5019. [Google Scholar] [CrossRef] [Green Version]
- Schuster, S.; Cabrera, D.; Arrese, M.; Feldstein, A.E. Triggering and Resolution of Inflammation in NASH. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 349–364. [Google Scholar] [CrossRef]
- Yeh, C.F.; Cheng, S.H.; Lin, Y.S.; Shentu, T.P.; Huang, R.T.; Zhu, J.; Chen, Y.T.; Kumar, S.; Lin, M.S.; Kao, H.L.; et al. Targeting Mechanosensitive Endothelial TXNDC5 to Stabilize ENOS and Reduce Atherosclerosis in Vivo. Sci. Adv. 2022, 8, 8096. [Google Scholar] [CrossRef]
- Hung, C.-T.; Su, T.-H.; Chen, Y.-T.; Wu, Y.-F.; Chen, Y.-T.; Lin, S.-J.; Lin, S.-L.; Yang, K.-C. Targeting ER Protein TXNDC5 in Hepatic Stellate Cell Mitigates Liver Fibrosis by Repressing Non-Canonical TGFβ Signalling. Gut 2021, gutjnl-2021-325065. [Google Scholar] [CrossRef]
- Zeng, Y.; Ma, W.; Ma, C.; Ren, X.; Wang, Y. Inhibition of TXNDC5 Attenuates Lipopolysaccharide-Induced Septic Shock by Altering Inflammatory Responses. Lab. Investig. 2021, 1–10. [Google Scholar] [CrossRef]
- Tröder, S.E.; Ebert, L.K.; Butt, L.; Assenmacher, S.; Schermer, B.; Zevnik, B. An Optimized Electroporation Approach for Efficient CRISPR/Cas9 Genome Editing in Murine Zygotes. PLoS ONE 2018, 13, e0196891. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Calvo, R.; Guadall, A.; Calvayrac, O.; Navarro, M.A.; Alonso, J.; Ferrán, B.; de Diego, A.; Muniesa, P.; Osada, J.; Rodríguez, C.; et al. Over-Expression of Neuron-Derived Orphan Receptor-1 (NOR-1) Exacerbates Neointimal Hyperplasia after Vascular Injury. Hum. Mol. Genet. 2013, 22, 1949–1959. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Marcos, L.V.; Sancho-Knapik, S.; Gabás-Rivera, C.; Barranquero, C.; Gascón, S.; Romanos, E.; Martínez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Arnal, C.; et al. Pgc1a Is Responsible for the Sex Differences in Hepatic Cidec/Fsp27β MRNA Expression in Hepatic Steatosis of Mice Fed a Western Diet. Am. J. Physiol. Endocrinol Metab. 2020, 318, E249–E261. [Google Scholar] [CrossRef]
- Abuobeid, R.; Herrera-Marcos, L.; Navarro, M.A.; Arnal, C.; Martínez-Beamonte, R.; Surra, J.; Osada, J. Dietary Erythrodiol Modifies Hepatic Transcriptome in Mice in a Sex and Dose-Dependent Way. Int. J. Mol. Sci. 2020, 21, 7331. [Google Scholar] [CrossRef]
- Herrera-Marcos, L.V.; Martínez-Beamonte, R.; Macías-Herranz, M.; Arnal, C.; Barranquero, C.; Puente-Lanzarote, J.J.; Gascón, S.; Herrero-Continente, T.; Gonzalo-Romeo, G.; Alastrué-Vera, V.; et al. Hepatic Galectin-3 Is Associated with Lipid Droplet Area in Non-Alcoholic Steatohepatitis in a New Swine Model. Sci. Rep. 2022, 12, 1024. [Google Scholar] [CrossRef]
- Lou-Bonafonte, J.M.; Gabás-Rivera, C.; Navarro, M.A.; Osada, J. PON1 and Mediterranean Diet. Nutrients 2015, 7, 4068–4092. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Beamonte, R.; Navarro, M.A.; Acin, S.; Guillén, N.; Barranquero, C.; Arnal, C.; Surra, J.; Osada, J. Postprandial Changes in High Density Lipoproteins in Rats Subjected to Gavage Administration of Virgin Olive Oil. PLoS ONE 2013, 8, e55231. [Google Scholar] [CrossRef] [Green Version]
- Navab, M.; Hama, S.Y.; Hough, G.P.; Subbanagounder, G.; Reddy, S.T.; Fogelman, A.M. A Cell-Free Assay for Detecting HDL That Is Dysfunctional in Preventing the Formation of or Inactivating Oxidized Phospholipids. J. Lipid Res. 2001, 42, 1308–1317. [Google Scholar] [CrossRef]
- Arbonés-Mainar, J.M.; Navarro, M.A.; Guzmán, M.A.; Arnal, C.; Surra, J.C.; Acín, S.; Carnicer, R.; Osada, J.; Roche, H.M. Selective Effect of Conjugated Linoleic Acid Isomers on Atherosclerotic Lesion Development in Apolipoprotein E Knockout Mice. Atherosclerosis 2006, 189, 318–327. [Google Scholar] [CrossRef]
- Asimakopoulou, A.; Weiskirchen, S.; Weiskirchen, R. Lipocalin 2 (LCN2) Expression in Hepatic Malfunction and Therapy. Front. Physiol. 2016, 7, 430. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, K.; Okuda, A.; Yusa, K.; Shinkai, Y. A CRISPR Knockout Screen Identifies SETDB1-Target Retroelement Silencing Factors in Embryonic Stem Cells. Genome Res. 2018, 28, 846–858. [Google Scholar] [CrossRef] [Green Version]
- Marhaug, G.; Dowton, S.B. Serum Amyloid A: An Acute Phase Apolipoprotein and Precursor of AA Amyloid. Baillière’s Clin. Rheumatol. 1994, 8, 553–573. [Google Scholar] [CrossRef]
- Stearman, R.S.; Lowell, C.A.; Peltzman, C.G.; Morrow, J.F. The Sequence and Structure of a New Serum Amyloid a Gene. Nucleic Acids Res. 1986, 14, 797–809. [Google Scholar] [CrossRef] [Green Version]
- Mori, M.; Tian, G.; Ishikawa, A.; Higuchi, K. Diversity and Complexity of the Mouse Saa1 and Saa2 Genes. Exp. Anim. 2014, 63, 99. [Google Scholar] [CrossRef] [Green Version]
- del Rio, D.; Stewart, A.J.; Pellegrini, N. A Review of Recent Studies on Malondialdehyde as Toxic Molecule and Biological Marker of Oxidative Stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Shunmoogam, N.; Naidoo, P.; Chilton, R. Paraoxonase (PON)-1: A Brief Overview on Genetics, Structure, Polymorphisms and Clinical Relevance. Vasc. Health Risk Manag. 2018, 14, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wu, Z.; Riwanto, M.; Gao, S.; Levison, B.S.; Gu, X.; Fu, X.; Wagner, M.A.; Besler, C.; Gerstenecker, G.; et al. Myeloperoxidase, Paraoxonase-1, and HDL Form a Functional Ternary Complex. J. Clin. Investig. 2013, 123, 3815. [Google Scholar] [CrossRef] [Green Version]
- Stafforini, D.M. Plasma PAF-AH (PLA2G7): Biochemical Properties, Association with LDLs and HDLs, and Regulation of Expression. Enzymes 2015, 38, 71–93. [Google Scholar] [CrossRef]
- Lee, T.H.; Yeh, C.F.; Lee, Y.T.; Shih, Y.C.; Chen, Y.T.; Hung, C.T.; You, M.Y.; Wu, P.C.; Shentu, T.P.; Huang, R.T.; et al. Fibroblast-Enriched Endoplasmic Reticulum Protein TXNDC5 Promotes Pulmonary Fibrosis by Augmenting TGFβ Signaling through TGFBR1 Stabilization. Nat. Commun. 2020, 11, 4254. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.-C.; Shih, Y.-C.; Chen, C.-L.; Zhang, Y.; Mellor, R.L.; Kanter, E.M.; Fang, Y.; Wang, H.-C.; Hung, C.-T.; Nong, J.-Y.; et al. Molecular Medicine Endoplasmic Reticulum Protein TXNDC5 Augments Myocardial Fibrosis by Facilitating Extracellular Matrix Protein Folding and Redox-Sensitive Cardiac Fibroblast Activation. Circ. Res. 2018, 122, 1052–1068. [Google Scholar] [CrossRef]
- Buck, M.; Gouwy, M.; Wang, J.; Snick, J.; Opdenakker, G.; Struyf, S.; Damme, J. Structure and Expression of Different Serum Amyloid A (SAA) Variants and Their Concentration-Dependent Functions during Host Insults. Curr. Med. Chem. 2016, 23, 1725–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chait, A.; den Hartigh, L.J.; Wang, S.; Goodspeed, L.; Babenko, I.; Altemeier, W.A.; Vaisar, T. Presence of Serum Amyloid A3 in Mouse Plasma Is Dependent on the Nature and Extent of the Inflammatory Stimulus. Sci. Rep. 2020, 10, 10397. [Google Scholar] [CrossRef]
- Price, A.J.; Manjegowda, M.C.; Kain, J.; Anandh, S.; Bochkis, I.M. Hdac3, Setdb1, and Kap1 Mark H3K9me3/H3K14ac Bivalent Regions in Young and Aged Liver. Aging Cell 2020, 19, e13092. [Google Scholar] [CrossRef] [Green Version]
- Yagnik, D.; Hills, F. Urate Crystals Induce Macrophage PAF-AH Secretion Which Is Differentially Regulated by TGFβ1 and Hydrocortisone. Mol. Med. Rep. 2018, 18, 3506–3512. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.T.; Yudell, B.E.; Loor, J.J. Regulation of Energy Metabolism by Long-Chain Fatty Acids. Prog. Lipid Res. 2014, 53, 124–144. [Google Scholar] [CrossRef]
- Wang, L.; Dong, H.; Song, G.; Zhang, R.; Pan, J.; Han, J. TXNDC5 Synergizes with HSC70 to Exacerbate the Inflammatory Phenotype of Synovial Fibroblasts in Rheumatoid Arthritis through NF-ΚB Signaling. Cell. Mol. Immunol. 2018, 15, 685–696. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, J.; Zhang, X.; Tian, R.; Qiao, L.; Ge, L.; Pan, J.; Wang, L. TXNDC5 Protects Synovial Fibroblasts of Rheumatoid Arthritis from the Detrimental Effects of Endoplasmic Reticulum Stress. Intractable Rare Dis. Res. 2020, 9, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Yang, M.; Zhou, B.; Luo, J.; Zhang, Z.; Zhang, W.; Yan, Z. CircRNA-104718 Acts as Competing Endogenous RNA and Promotes Hepatocellular Carcinoma Progression through MicroRNA-218-5p/TXNDC5 Signaling Pathway. Clin. Sci. 2019, 133, 1487–1503. [Google Scholar] [CrossRef]
- Seo, E.; Kang, H.; Choi, H.; Choi, W.; Jun, H.S. Reactive Oxygen Species-Induced Changes in Glucose and Lipid Metabolism Contribute to the Accumulation of Cholesterol in the Liver during Aging. Aging Cell 2019, 18, e12895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stonik, J.A.; Remaley, A.T.; Demosky, S.J.; Neufeld, E.B.; Bocharov, A.; Brewer, H.B. Serum Amyloid a Promotes ABCA1-Dependent and ABCA1-Independent Lipid Efflux from Cells. Biochem. Biophys. Res. Commun. 2004, 321, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.J.; Binder, C.J.; Krychtiuk, K.A.; Schillinger, M.; Minar, E.; Hoke, M. The Prognostic Value of Serum Amyloid A for Long-Term Mortality among Patients with Subclinical Carotid Atherosclerosis. Eur. J. Clin. Investig. 2019, 49, e13095. [Google Scholar] [CrossRef] [PubMed]
- Kosuge, M.; Ebina, T.; Ishikawa, T.; Hibi, K.; Tsukahara, K.; Okuda, J.; Iwahashi, N.; Ozaki, H.; Yano, H.; Kusama, I.; et al. Serum Amyloid A Is a Better Predictor of Clinical Outcomes than C-Reactive Protein in Non-ST-Segment Elevation Acute Coronary Syndromes. Circ. J. 2007, 71, 186–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Beer, M.C.; Wroblewski, J.M.; Noffsinger, V.P.; Rateri, D.L.; Howatt, D.A.; Balakrishnan, A.; Ji, A.; Shridas, P.; Thompson, J.C.; van der Westhuyzen, D.R.; et al. Deficiency of Endogenous Acute Phase Serum Amyloid A Does Not Affect Atherosclerotic Lesions in Apolipoprotein E-Deficient Mice. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 255–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | GenBank | Biological Process | Gene Symbol | SL2R RNA seq | p Value RNA seq | WT | KO | SL2R qPCR |
|---|---|---|---|---|---|---|---|---|
| Serum Amyloid A2 | NM_011314.3 | Acute phase protein | Saa2 | 3.76 | 5.91 × 10−17 | 1.3 ± 1 | 26 ± 27 *** | 4.31 |
| Serum Amyloid A1 | NM_009117.4 | Acute phase protein | Saa1 | 3.13 | 6.41 × 10−13 | 1.1 ± 0.5 | 23 ± 24 *** | 4.41 |
| Solute Carrier Family 4 Member 1 | NM_011403.2 | Anion Exchange Protein | Slc4a1 | 1.47 | 1.80 × 10−3 | 1.6 ± 1.1 | 2.9 ± 1.8 * | 0.90 |
| Lipocalin 2 | NM_008491.1 | Iron-trafficking protein | Lcn2 | 1.43 | 1.08 × 10−3 | 1.5 ± 1.2 | 6.3 ± 7 ** | 2.11 |
| Rho Related BTB Domain Containing 1 | NM_001081347.1 | Small GTPase of Rho family | Rhobtb1 | 1.11 | 7.12 × 10−3 | 1.1 ± 0.7 | 2.5 ± 2 * | 0.91 |
| Lipin 1 | NM_001355598.1 | Triglyceride synthesis | Lpin1 | 1.00 | 3.74 × 10−3 | 1.1 ± 0.6 | 2.9 ± 2 * | 1.37 |
| Cytochrome P450, family 7, subfamily a, polypeptide 1 | NM_007824.3 | Bile acid synthesis | Cyp7a1 | −1.11 | 2.79 × 10−3 | 1.4 ± 0.8 | 0.9 ± 0.6 | −0.68 |
| Retroelement Silencing Factor 1 | NM_001289662.1 | Regulation of imprinted gene expression | Resf1 | −1.53 | 1.24 × 10−3 | 1.2 ± 0.7 | 0.5 ± 0.6 ** | −1.30 |
| Thioredoxin Domain Containing 5 | NM_145367.4 | Disulfide isomerase | Txndc5 | −4.89 | 2.77 × 10−35 | 1.1 ± 0.4 | 0.1 ± 0 *** | −4.41 |
| Modulator of Apoptosis 1 | NM_001142937.2 | Receptor-dependent apoptosis | Moap1 | −2.85 | 6.35 × 10−8 | NA | NA | NA |
| Wild Type (n = 13) | Knock-Out (n = 10) | |
|---|---|---|
| Triglycerides (mM) | 1.8 ± 0.3 | 2 ± 0.5 |
| Cholesterol (mM) | 2.7 ± 0.4 | 3 ± 0.2 |
| Phosphatidylcholine (mM) | 4.2 ± 1.4 | 4.7 ± 1.8 |
| Ketone bodies (mM) | 1.7 ± 0.5 | 1.7 ± 0.4 |
| Malondialdehyde (mM) | 0.59 ± 0.17 | 0.67 ± 0.12 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Marco, J.; Martínez-Beamonte, R.; Diego, A.D.; Herrero-Continente, T.; Barranquero, C.; Arnal, C.; Surra, J.; Navarro, M.A.; Osada, J. Thioredoxin Domain Containing 5 Suppression Elicits Serum Amyloid A-Containing High-Density Lipoproteins. Biomedicines 2022, 10, 709. https://doi.org/10.3390/biomedicines10030709
Sánchez-Marco J, Martínez-Beamonte R, Diego AD, Herrero-Continente T, Barranquero C, Arnal C, Surra J, Navarro MA, Osada J. Thioredoxin Domain Containing 5 Suppression Elicits Serum Amyloid A-Containing High-Density Lipoproteins. Biomedicines. 2022; 10(3):709. https://doi.org/10.3390/biomedicines10030709
Chicago/Turabian StyleSánchez-Marco, Javier, Roberto Martínez-Beamonte, Alicia De Diego, Tania Herrero-Continente, Cristina Barranquero, Carmen Arnal, Joaquín Surra, María A. Navarro, and Jesús Osada. 2022. "Thioredoxin Domain Containing 5 Suppression Elicits Serum Amyloid A-Containing High-Density Lipoproteins" Biomedicines 10, no. 3: 709. https://doi.org/10.3390/biomedicines10030709