
Influence of the Immune Microenvironment Provided by Implanted Biomaterials on the Biological Properties of Masquelet-Induced Membranes in Rats: Metakaolin as an Alternative Spacer

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Surgical Procedures
2.3. Spacers
2.4. Bone Turnover Assessment
2.5. Histology
2.6. BMP-2 Immunostaining
2.7. Real-Time PCR Analysis
2.8. Immunofluorescence Assays and Macrophage Quantification
2.9. Liquid Chromatography–Tandem Mass Spectrometry (LC-MS/MS)
2.10. MicroCT
2.11. Statistical Analysis
3. Results
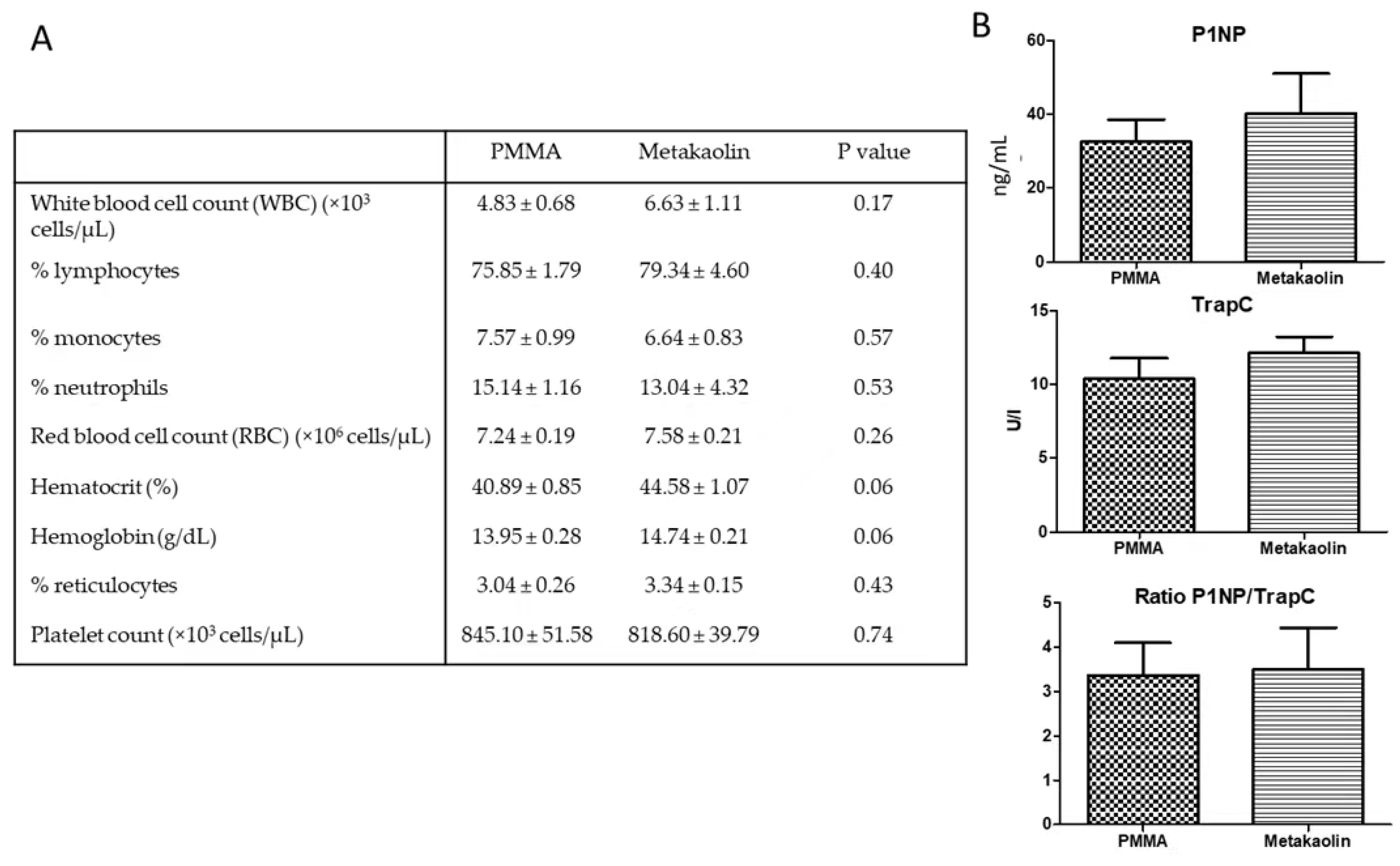
3.1. Animals and Blood Parameters at the End of IMT Stage 1
3.2. Comparison of Biological Properties between Metakaolin- and PMMA-Induced Membranes
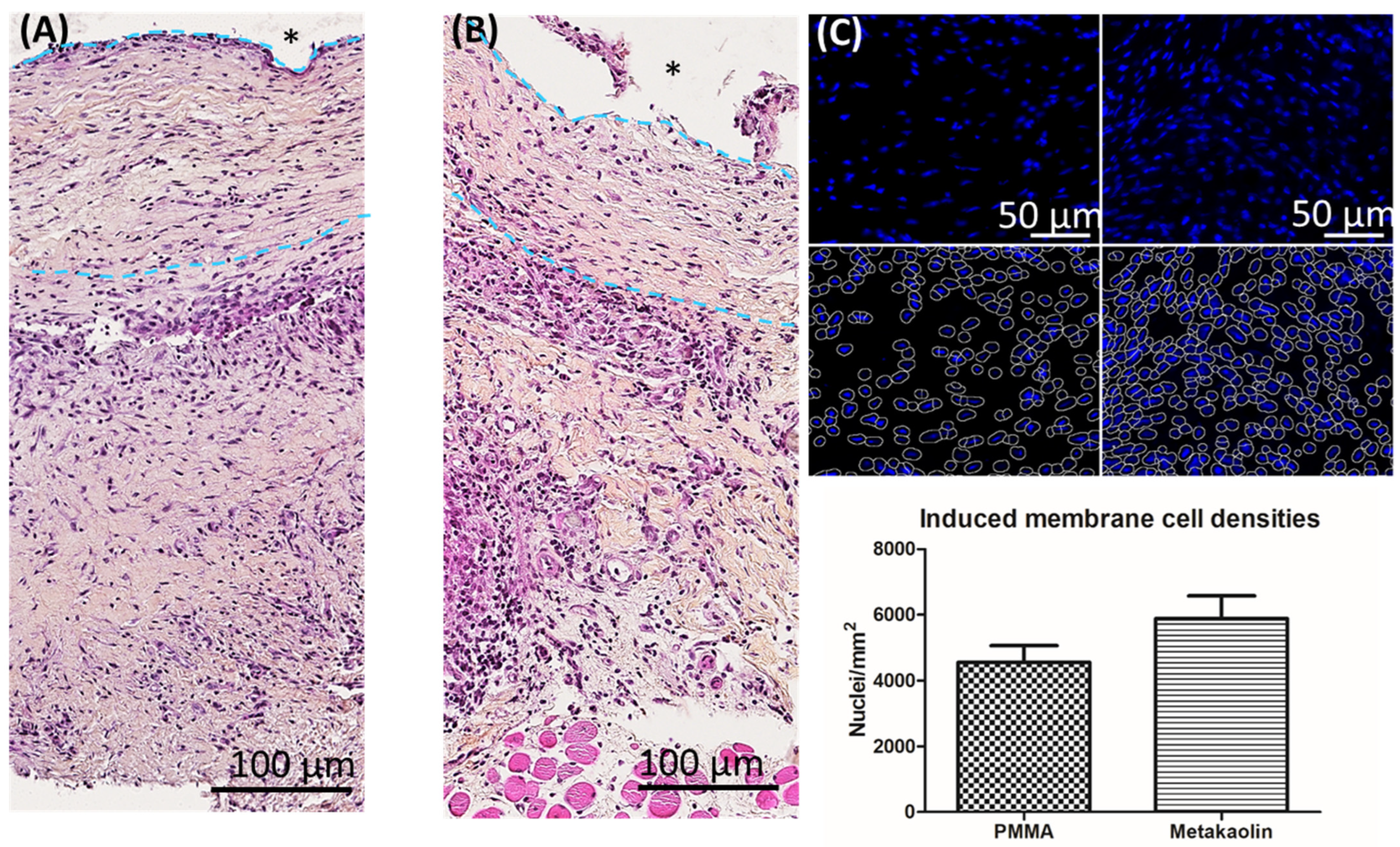
3.2.1. Membrane Architecture and Cellularity
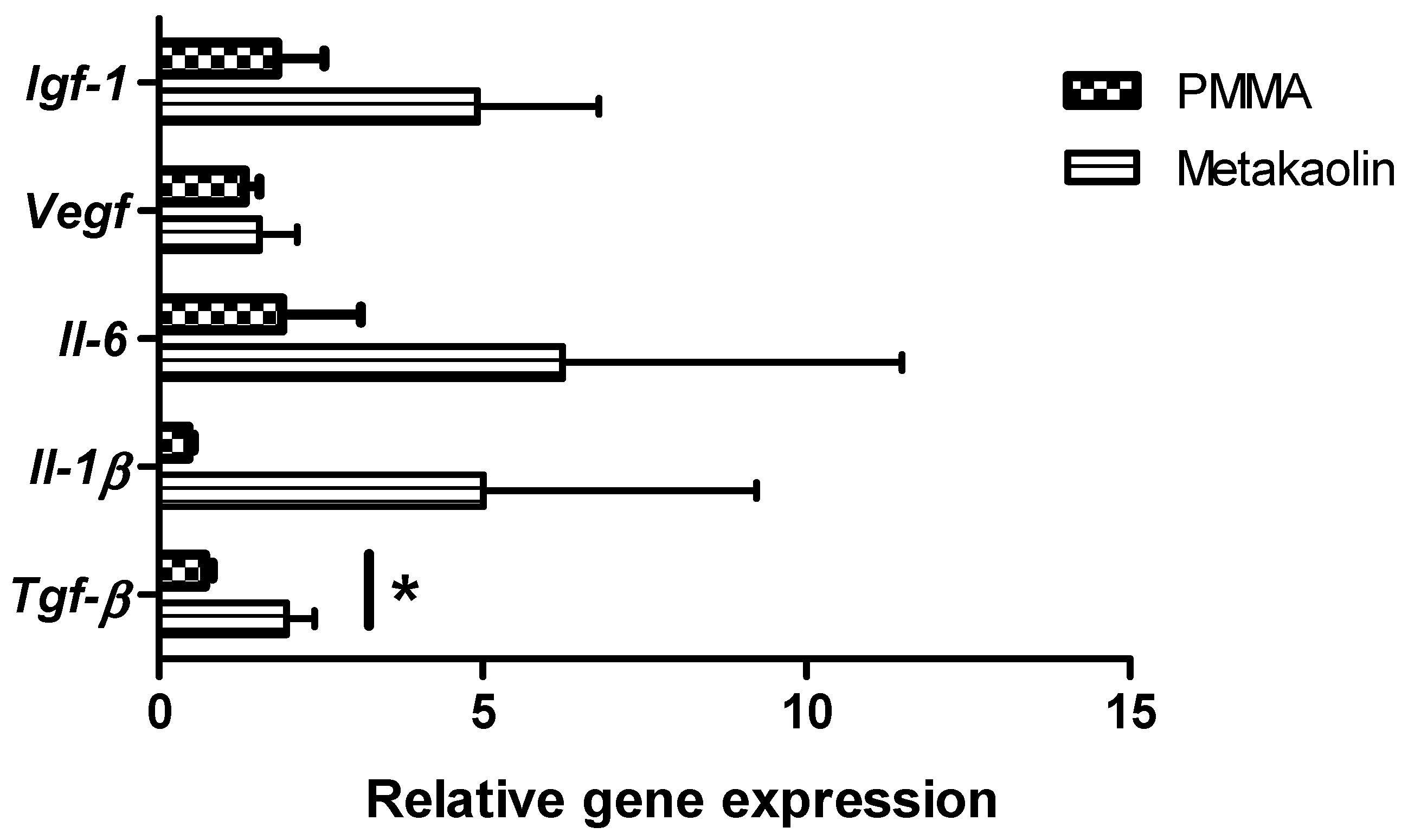
3.2.2. Gene Expression within Membranes
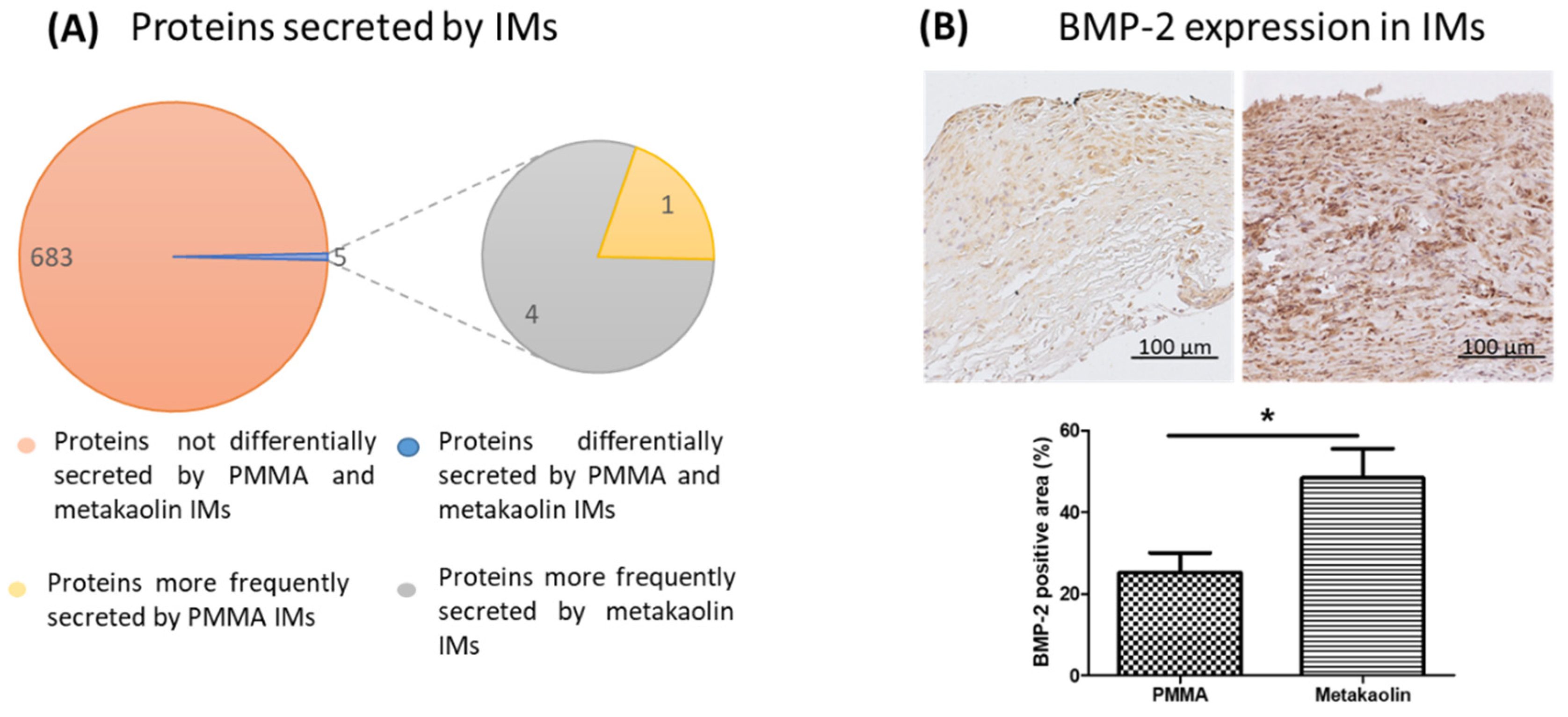
3.2.3. Secretion of Proteins by the IM and BMP-2 Expression within Membranes
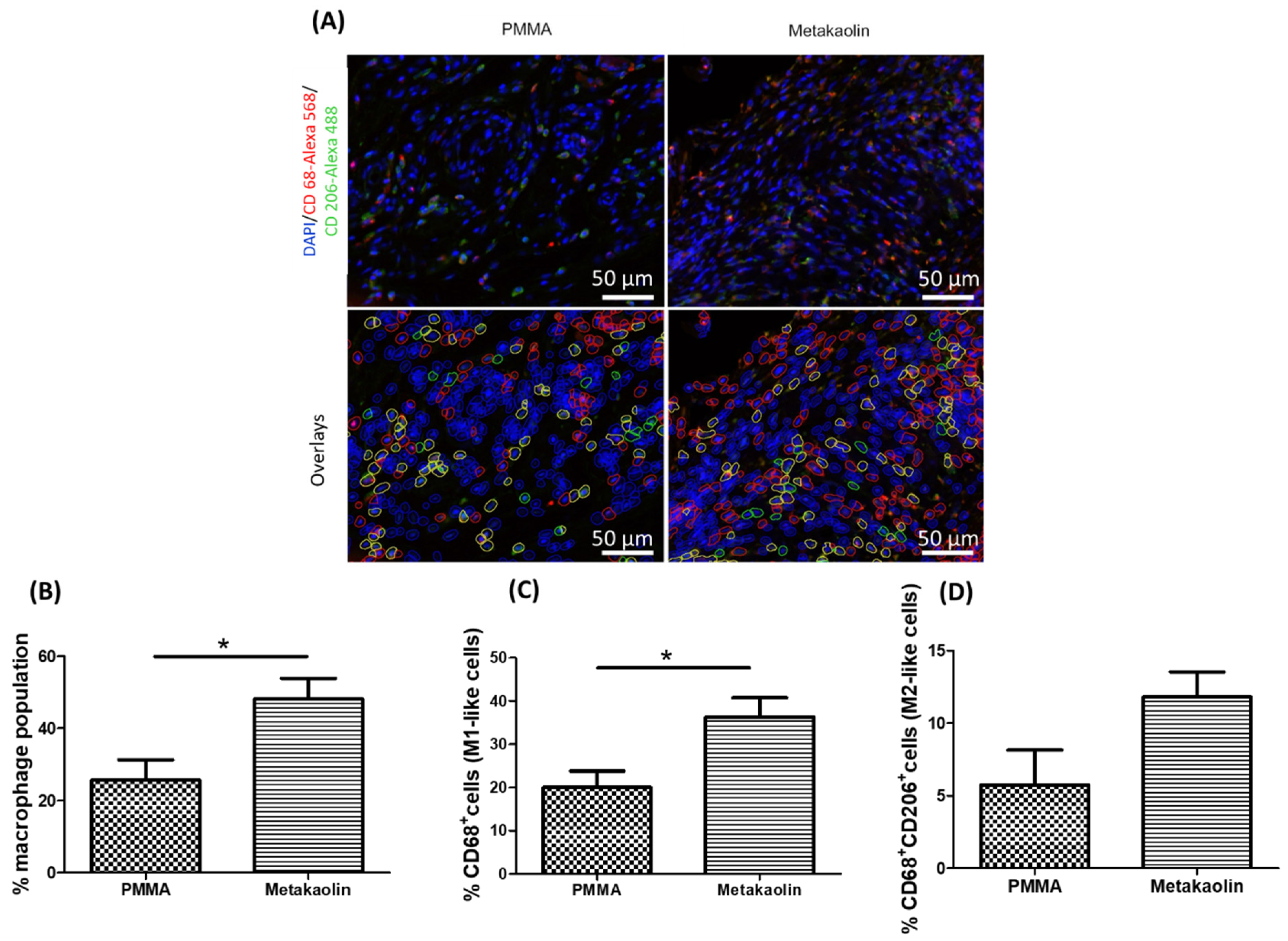
3.3. Macrophage Distribution in IMs
3.4. Bone Healing after IMT Stage 2 Surgery
4. Discussion
4.1. Metakaolin Modifies Several Osteogenic and Biological Parameters of IMs
4.2. Metakaolin Spacers Modulate the IM Immune Microenvironment
4.3. Metakaolin Slightly Improves Bone Repair Efficiency
5. Limitations, Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Masquelet, A.C.; Giannoudis, P.V. The Induced Membrane Technique for Treatment of Bone Defects: What Have I Learned? Trauma. Case Rep. 2021, 36, 100556. [Google Scholar] [CrossRef] [PubMed]
- Zwetyenga, N.; Catros, S.; Emparanza, A.; Deminiere, C.; Siberchicot, F.; Fricain, J.-C. Mandibular Reconstruction Using Induced Membranes with Autologous Cancellous Bone Graft and HA-BetaTCP: Animal Model Study and Preliminary Results in Patients. Int. J. Oral Maxillofac. Surg. 2009, 38, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Christou, C.; Oliver, R.A.; Yu, Y.; Walsh, W.R. The Masquelet Technique for Membrane Induction and the Healing of Ovine Critical Sized Segmental Defects. PLoS ONE 2014, 9, e114122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, M.; Barbier, L.; Mathieu, L.; Poyot, T.; Demoures, T.; Souraud, J.-B.; Masquelet, A.-C.; Collombet, J.-M. Towards Understanding Therapeutic Failures in Masquelet Surgery: First Evidence That Defective Induced Membrane Properties Are Associated with Clinical Failures. J. Clin. Med. 2020, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Henrich, D.; Seebach, C.; Nau, C.; Basan, S.; Relja, B.; Wilhelm, K.; Schaible, A.; Frank, J.; Barker, J.; Marzi, I. Establishment and Characterization of the Masquelet Induced Membrane Technique in a Rat Femur Critical-Sized Defect Model. J. Tissue Eng. Regen. Med. 2016, 10, E382–E396. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, W.; Fan, X.; Li, H.; Wang, H.; Liao, R.; Hu, Y.; Zeng, M. Masquelet Technique: Effects of Vancomycin Concentration on Quality of the Induced Membrane. Injury 2022, 53, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign Body Reaction to Biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Klopfleisch, R. Macrophage Reaction against Biomaterials in the Mouse Model-Phenotypes, Functions and Markers. Acta Biomater. 2016, 43, 3–13. [Google Scholar] [CrossRef]
- Davidovits, J. Geopolymers. J. Therm. Anal. 1991, 37, 1633–1656. [Google Scholar] [CrossRef]
- Jung, M.; Shin, M.-K.; Jung, Y.-K.; Yoo, H.S. Modulation of Macrophage Activities in Proliferation, Lysosome, and Phagosome by the Nonspecific Immunostimulator, Mica. PLoS ONE 2015, 10, e0117838. [Google Scholar] [CrossRef]
- Bowman, P.D.; Wang, X.; Meledeo, M.A.; Dubick, M.A.; Kheirabadi, B.S. Toxicity of Aluminum Silicates Used in Hemostatic Dressings toward Human Umbilical Veins Endothelial Cells, HeLa Cells, and RAW267.4 Mouse Macrophages. J. Trauma. 2011, 71, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, M.; Vennemann, A.; Wohlleben, W. Lung Toxicity Analysis of Nano-Sized Kaolin and Bentonite: Missing Indications for a Common Grouping. Nanomaterials 2020, 10, 204. [Google Scholar] [CrossRef] [Green Version]
- Khurana, I.S.; Kaur, S.; Kaur, H.; Khurana, R.K. Multifaceted Role of Clay Minerals in Pharmaceuticals. Future Sci. OA 2015, 1, FSO6. [Google Scholar] [CrossRef] [PubMed]
- Awad, M.E.; López-Galindo, A.; Setti, M.; El-Rahmany, M.M.; Iborra, C.V. Kaolinite in Pharmaceutics and Biomedicine. Int. J. Pharm. 2017, 533, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, L.; Murison, J.C.; de Rousiers, A.; de l’Escalopier, N.; Lutomski, D.; Collombet, J.-M.; Durand, M. The Masquelet Technique: Can Disposable Polypropylene Syringes Be an Alternative to Standard PMMA Spacers? A Rat Bone Defect Model. Clin. Orthop. Relat. Res. 2021, 479, 2737–2751. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Schroeder, A.; Mueller, O.; Stocker, S.; Salowsky, R.; Leiber, M.; Gassmann, M.; Lightfoot, S.; Menzel, W.; Granzow, M.; Ragg, T. The RIN: An RNA Integrity Number for Assigning Integrity Values to RNA Measurements. BMC Mol. Biol. 2006, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Pugniere, P.; Banzet, S.; Chaillou, T.; Mouret, C.; Peinnequin, A. Pitfalls of Reverse Transcription Quantitative Polymerase Chain Reaction Standardization: Volume-Related Inhibitors of Reverse Transcription. Anal Biochem 2011, 415, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikovics, K.; Durand, M.; Castellarin, C.; Burger, J.; Sicherre, E.; Collombet, J.-M.; Oger, M.; Holy, X.; Favier, A.-L. Macrophages Characterization in an Injured Bone Tissue. Biomedicines 2022, 10, 1385. [Google Scholar] [CrossRef]
- Kosmac, K.; Peck, B.D.; Walton, R.G.; Mula, J.; Kern, P.A.; Bamman, M.M.; Dennis, R.A.; Jacobs, C.A.; Lattermann, C.; Johnson, D.L.; et al. Immunohistochemical Identification of Human Skeletal Muscle Macrophages. Bio. Protoc. 2018, 8, e2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gindraux, F.; Rondot, T.; de Billy, B.; Zwetyenga, N.; Fricain, J.-C.; Pagnon, A.; Obert, L. Similarities between Induced Membrane and Amniotic Membrane: Novelty for Bone Repair. Placenta 2017, 59, 116–123. [Google Scholar] [CrossRef]
- Gouron, R.; Petit, L.; Boudot, C.; Six, I.; Brazier, M.; Kamel, S.; Mentaverri, R. Osteoclasts and Their Precursors Are Present in the Induced-Membrane during Bone Reconstruction Using the Masquelet Technique. J. Tissue Eng Regen. Med. 2017, 11, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, R.J.; Churchman, S.M.; Tan, H.B.; McGonagle, D.; Jones, E.; Giannoudis, P.V. Induced Periosteum a Complex Cellular Scaffold for the Treatment of Large Bone Defects. Bone 2013, 57, 484–492. [Google Scholar] [CrossRef]
- Aho, O.-M.; Lehenkari, P.; Ristiniemi, J.; Lehtonen, S.; Risteli, J.; Leskelä, H.-V. The Mechanism of Action of Induced Membranes in Bone Repair. J. Bone Joint Surg. Am. 2013, 95, 597–604. [Google Scholar] [CrossRef]
- Wang, W.; Zuo, R.; Long, H.; Wang, Y.; Zhang, Y.; Sun, C.; Luo, G.; Zhang, Y.; Li, C.; Zhou, Y.; et al. Advances in the Masquelet Technique: Myeloid-Derived Suppressor Cells Promote Angiogenesis in PMMA-Induced Membranes. Acta Biomater. 2020, 108, 223–236. [Google Scholar] [CrossRef]
- Fischer, C.; Doll, J.; Tanner, M.; Bruckner, T.; Zimmermann, G.; Helbig, L.; Biglari, B.; Schmidmaier, G.; Moghaddam, A. Quantification of TGF-SS1, PDGF and IGF-1 Cytokine Expression after Fracture Treatment vs. Non-Union Therapy via Masquelet. Injury 2016, 47, 342–349. [Google Scholar] [CrossRef]
- Crane, J.L.; Xian, L.; Cao, X. Role of TGF-β Signaling in Coupling Bone Remodeling. Methods Mol. Biol. 2016, 1344, 287–300. [Google Scholar] [CrossRef]
- Tsiridis, E.; Upadhyay, N.; Giannoudis, P. Molecular Aspects of Fracture Healing: Which Are the Important Molecules? Injury 2007, 38 (Suppl. S1), S11–S25. [Google Scholar] [CrossRef]
- Tang, Q.; Tong, M.; Zheng, G.; Shen, L.; Shang, P.; Liu, H. Masquelet’s Induced Membrane Promotes the Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells by Activating the Smad and MAPK Pathways. Am. J. Transl. Res. 2018, 10, 1211–1219. [Google Scholar] [PubMed]
- Arango Duque, G.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.V.; Puleo, D.A. Infection, Inflammation, and Bone Regeneration. J. Dent. Res. 2011, 90, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Loi, F.; Córdova, L.A.; Pajarinen, J.; Lin, T.; Yao, Z.; Goodman, S.B. Inflammation, Fracture and Bone Repair. Bone 2016, 86, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Wang, Z.; Shi, Y.; Dong, L.; Wang, C. Modulating Macrophage Activities to Promote Endogenous Bone Regeneration: Biological Mechanisms and Engineering Approaches. Bioact. Mater. 2021, 6, 244–261. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Rhee, C.; Utsunomiya, T.; Zhang, N.; Ueno, M.; Yao, Z.; Goodman, S.B. Modulation of the Inflammatory Response and Bone Healing. Front. Endocrinol. 2020, 11, 386. [Google Scholar] [CrossRef]
- Chung, L.; Maestas, D.R.; Housseau, F.; Elisseeff, J.H. Key Players in the Immune Response to Biomaterial Scaffolds for Regenerative Medicine. Adv. Drug Deliv. Rev. 2017, 114, 184–192. [Google Scholar] [CrossRef]
- Martin, K.E.; García, A.J. Macrophage Phenotypes in Tissue Repair and the Foreign Body Response: Implications for Biomaterial-Based Regenerative Medicine Strategies. Acta Biomater. 2021, 133, 4–16. [Google Scholar] [CrossRef]
- Kyriakides, T.R.; Kim, H.-J.; Zheng, C.; Harkins, L.; Tao, W.; Deschenes, E. Foreign Body Response to Synthetic Polymer Biomaterials and the Role of Adaptive Immunity. Biomed. Mater. 2022, 17, 022007. [Google Scholar] [CrossRef]
- Stahl, A.; Hao, D.; Barrera, J.; Henn, D.; Lin, S.; Moeinzadeh, S.; Kim, S.; Maloney, W.; Gurtner, G.; Wang, A.; et al. A Bioactive Compliant Vascular Graft Modulates Macrophage Polarization and Maintains Patency with Robust Vascular Remodeling. Bioact. Mater. 2023, 19, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.N.; Valentin, J.E.; Stewart-Akers, A.M.; McCabe, G.P.; Badylak, S.F. Macrophage Phenotype and Remodeling Outcomes in Response to Biologic Scaffolds with and without a Cellular Component. Biomaterials 2009, 30, 1482–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Wang, W.; Nassiri, S.; Kwan, T.; Dang, C.; Liu, W.; Spiller, K.L. Temporal and Spatial Distribution of Macrophage Phenotype Markers in the Foreign Body Response to Glutaraldehyde-Crosslinked Gelatin Hydrogels. J. Biomater. Sci. Polym. Ed. 2016, 27, 721–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Liang, H.; Liu, X.; Wu, J.; Yang, C.; Wong, T.M.; Kwan, K.Y.H.; Cheung, K.M.C.; Wu, S.; Yeung, K.W.K. Regulation of Macrophage Polarization through Surface Topography Design to Facilitate Implant-to-Bone Osteointegration. Sci. Adv. 2021, 7, eabf6654. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, P.; Jones, E.; Panagiotopoulou, V.; Jha, A.; Blanchy, M.; Antimisiaris, S.; Anton, M.; Dhuiège, B.; Marotta, M.; Marjanovic, N.; et al. Electrospun and 3D Printed Polymeric Materials for One-Stage Critical-Size Long Bone Defect Regeneration Inspired by the Masquelet Technique: Recent Advances. Injury 2022, 53, S2–S12. [Google Scholar] [CrossRef]
- Liodakis, E.; Giannoudis, V.P.; Sehmisch, S.; Jha, A.; Giannoudis, P.V. Bone Defect Treatment: Does the Type and Properties of the Spacer Affect the Induction of Masquelet Membrane? Evidence Today. Eur. J. Trauma. Emerg. Surg. 2022, 1–22. [Google Scholar] [CrossRef]
- Toth, Z.; Roi, M.; Evans, E.; Watson, J.T.; Nicolaou, D.; McBride-Gagyi, S. Masquelet Technique: Effects of Spacer Material and Micro-Topography on Factor Expression and Bone Regeneration. Ann. Biomed. Eng. 2019, 47, 174–189. [Google Scholar] [CrossRef]
- Murison, J.-C.; Pfister, G.; Amar, S.; Rigal, S.; Mathieu, L. Metacarpal Bone Reconstruction by a Cementless Induced Membrane Technique. Hand Surg. Rehabil. 2019, 38, 83–86. [Google Scholar] [CrossRef]
- Ma, Y.-F.; Jiang, N.; Zhang, X.; Qin, C.-H.; Wang, L.; Hu, Y.-J.; Lin, Q.-R.; Yu, B.; Wang, B.-W. Calcium Sulfate Induced versus PMMA-Induced Membrane in a Critical-Sized Femoral Defect in a Rat Model. Sci. Rep. 2018, 8, 637. [Google Scholar] [CrossRef] [Green Version]
- Haffner-Luntzer, M.; Fischer, V.; Ignatius, A. Differences in Fracture Healing Between Female and Male C57BL/6J Mice. Front. Physiol. 2021, 12, 712494. [Google Scholar] [CrossRef]
- Strube, P.; Mehta, M.; Baerenwaldt, A.; Trippens, J.; Wilson, C.J.; Ode, A.; Perka, C.; Duda, G.N.; Kasper, G. Sex-Specific Compromised Bone Healing in Female Rats Might Be Associated with a Decrease in Mesenchymal Stem Cell Quantity. Bone 2009, 45, 1065–1072. [Google Scholar] [CrossRef]
- Betjes, M.G.; Haks, M.C.; Tuk, C.W.; Beelen, R.H. Monoclonal Antibody EBM11 (Anti-CD68) Discriminates between Dendritic Cells and Macrophages after Short-Term Culture. Immunobiology 1991, 183, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Chistiakov, D.A.; Killingsworth, M.C.; Myasoedova, V.A.; Orekhov, A.N.; Bobryshev, Y.V. CD68/Macrosialin: Not Just a Histochemical Marker. Lab. Investig. 2017, 97, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, M.H.M.; Hauck, F.; Dreyer, J.H.; Kempkes, B.; Niedobitek, G. Macrophage Polarisation: An Immunohistochemical Approach for Identifying M1 and M2 Macrophages. PLoS ONE 2013, 8, e80908. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, A.; Karim, M.R.; Kuramochi, M.; Izawa, T.; Kuwamura, M.; Yamate, J. Characterization of Macrophages and Myofibroblasts Appearing in Dibutyltin Dichloride-Induced Rat Pancreatic Fibrosis. Toxicol. Pathol. 2020, 48, 509–523. [Google Scholar] [CrossRef]
- Tsuji, Y.; Kuramochi, M.; Golbar, H.M.; Izawa, T.; Kuwamura, M.; Yamate, J. Acetaminophen-Induced Rat Hepatotoxicity Based on M1/M2-Macrophage Polarization, in Possible Relation to Damage-Associated Molecular Patterns and Autophagy. Int. J. Mol. Sci. 2020, 21, 8998. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Karim, M.R.; Izawa, T.; Kuwamura, M.; Yamate, J. Immunophenotypical Characterization of M1/M2 Macrophages and Lymphocytes in Cisplatin-Induced Rat Progressive Renal Fibrosis. Cells 2021, 10, 257. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durand, M.; Oger, M.; Nikovics, K.; Venant, J.; Guillope, A.-C.; Jouve, E.; Barbier, L.; Bégot, L.; Poirier, F.; Rousseau, C.; et al. Influence of the Immune Microenvironment Provided by Implanted Biomaterials on the Biological Properties of Masquelet-Induced Membranes in Rats: Metakaolin as an Alternative Spacer. Biomedicines 2022, 10, 3017. https://doi.org/10.3390/biomedicines10123017
Durand M, Oger M, Nikovics K, Venant J, Guillope A-C, Jouve E, Barbier L, Bégot L, Poirier F, Rousseau C, et al. Influence of the Immune Microenvironment Provided by Implanted Biomaterials on the Biological Properties of Masquelet-Induced Membranes in Rats: Metakaolin as an Alternative Spacer. Biomedicines. 2022; 10(12):3017. https://doi.org/10.3390/biomedicines10123017
Chicago/Turabian StyleDurand, Marjorie, Myriam Oger, Krisztina Nikovics, Julien Venant, Anne-Cecile Guillope, Eugénie Jouve, Laure Barbier, Laurent Bégot, Florence Poirier, Catherine Rousseau, and et al. 2022. "Influence of the Immune Microenvironment Provided by Implanted Biomaterials on the Biological Properties of Masquelet-Induced Membranes in Rats: Metakaolin as an Alternative Spacer" Biomedicines 10, no. 12: 3017. https://doi.org/10.3390/biomedicines10123017