
Prevention and Therapy of Metastatic HER-2+ Mammary Carcinoma with a Human Candidate HER-2 Virus-like Particle Vaccine
 , , , and
, , , and 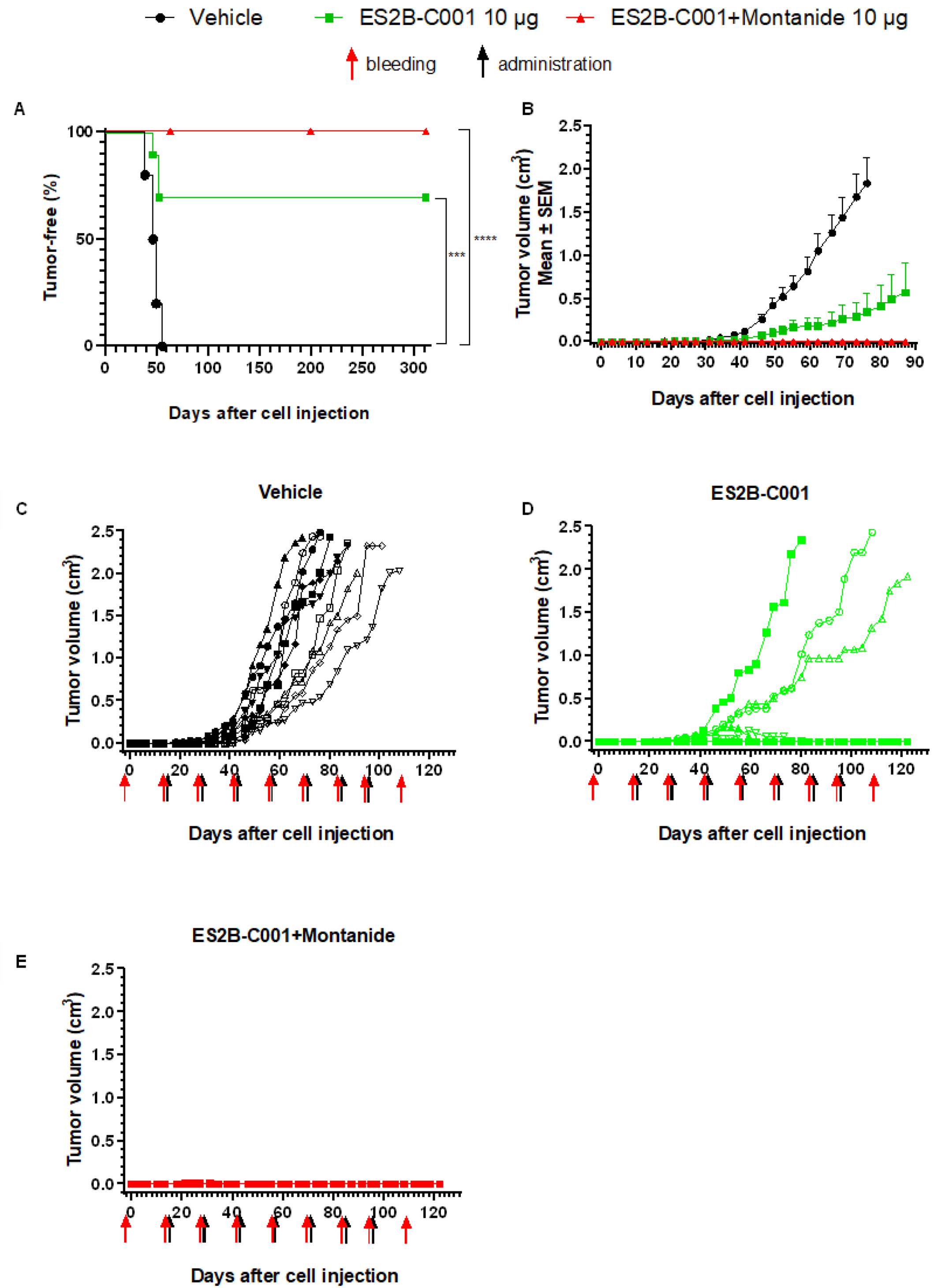{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Design, Expression and Purification of Catcher-HER-2 Antigen
2.3. Design, Expression and Purification of Tag-VLP
2.4. Formulation and Purification of ES2B-C001 Vaccine
2.5. Vaccinations
2.6. Cells
2.7. Local Tumor and Metastasis Therapy Models
2.8. Indirect Immunofluorescence and Flow Cytometry
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
2.10. Enzyme-Linked Immunospot (ELISpot)
2.11. Agar Colony Growth Inhibition
3. Results
3.1. Therapy of Mammary Carcinoma in FVB Mice
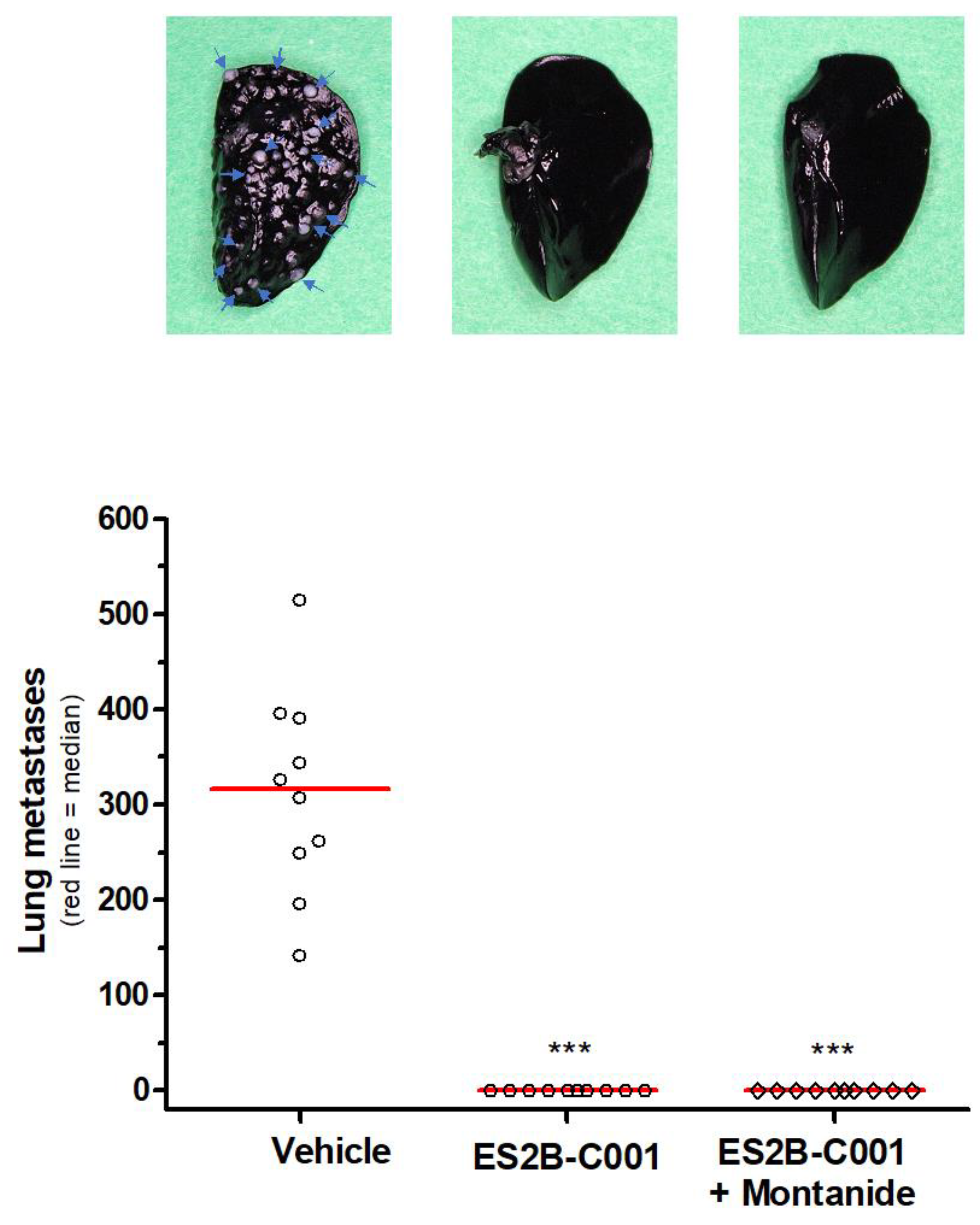
3.2. Therapy of Lung Metastases in FVB Mice
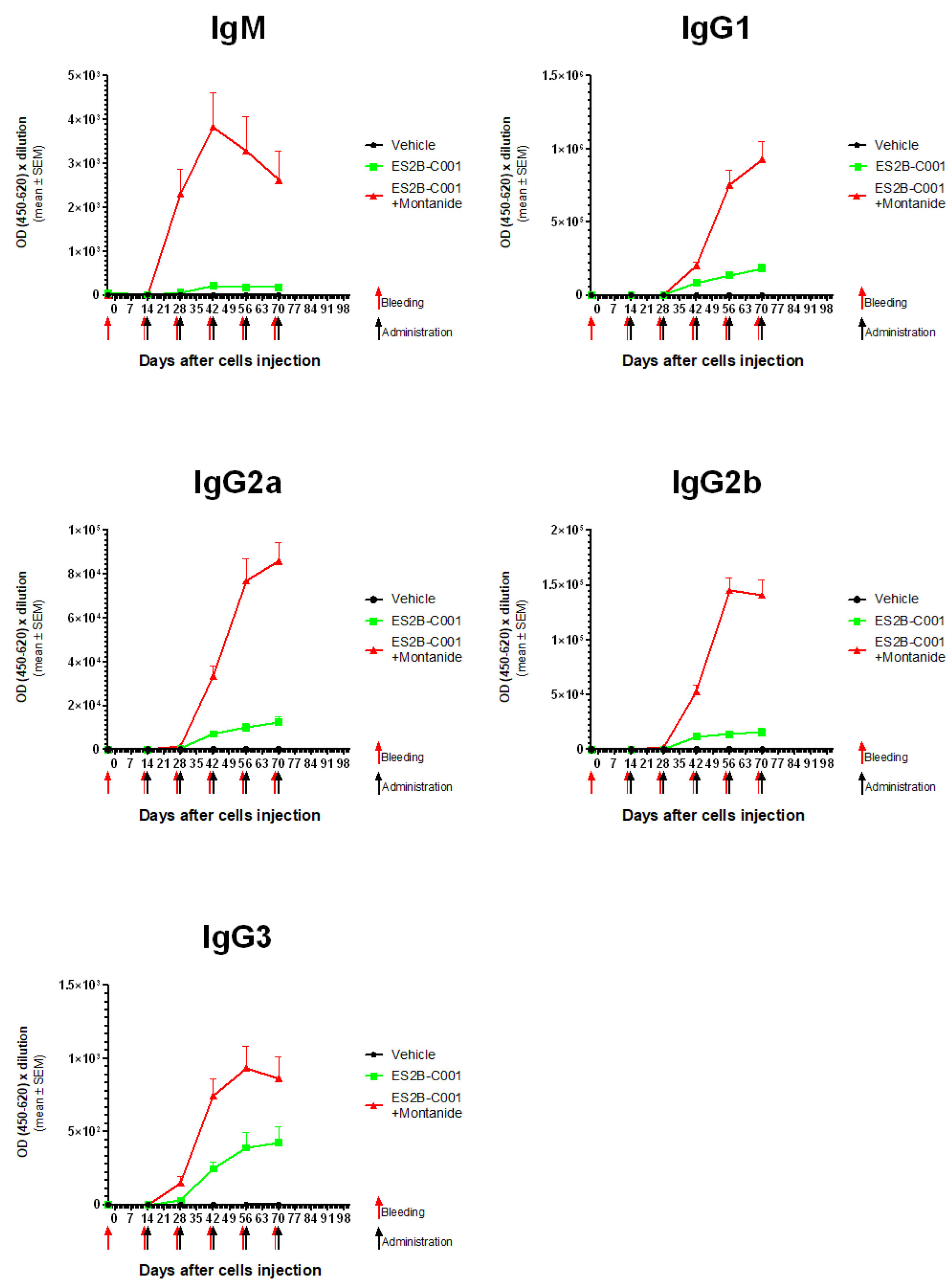
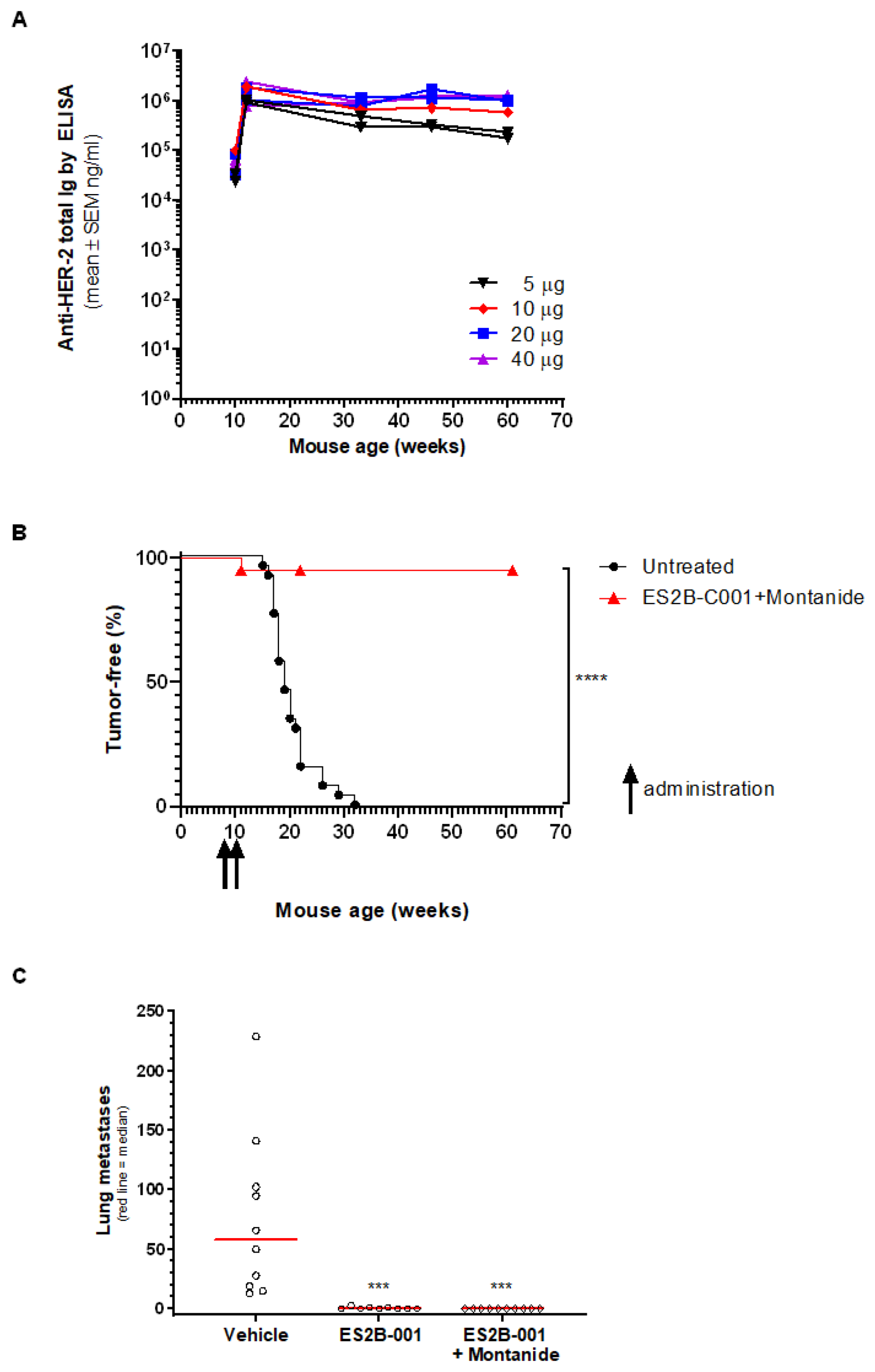
3.3. Anti-HER-2 Antibody Response
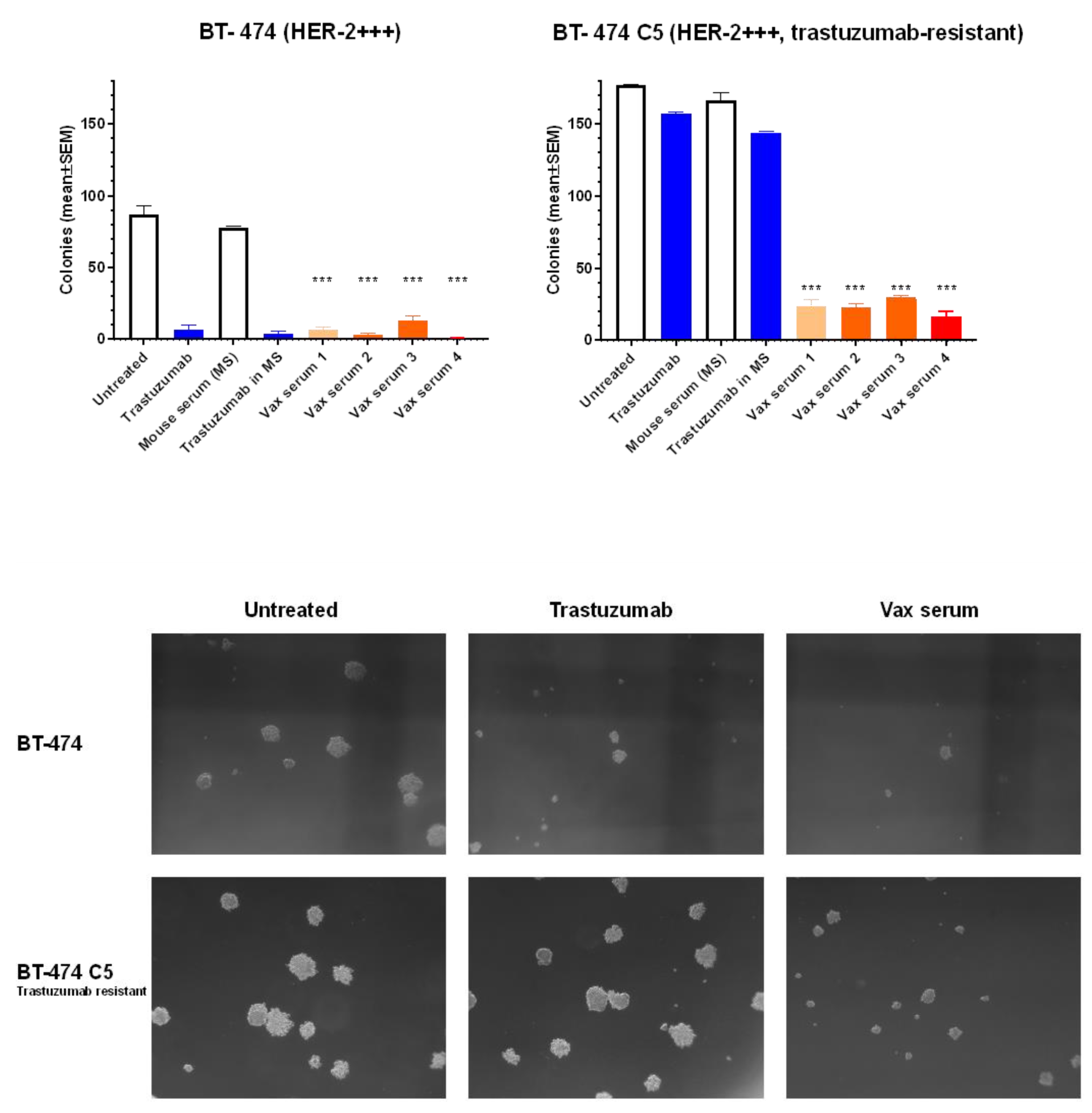
3.4. Inhibition of Human Breast Cancer Cells in 3D Culture
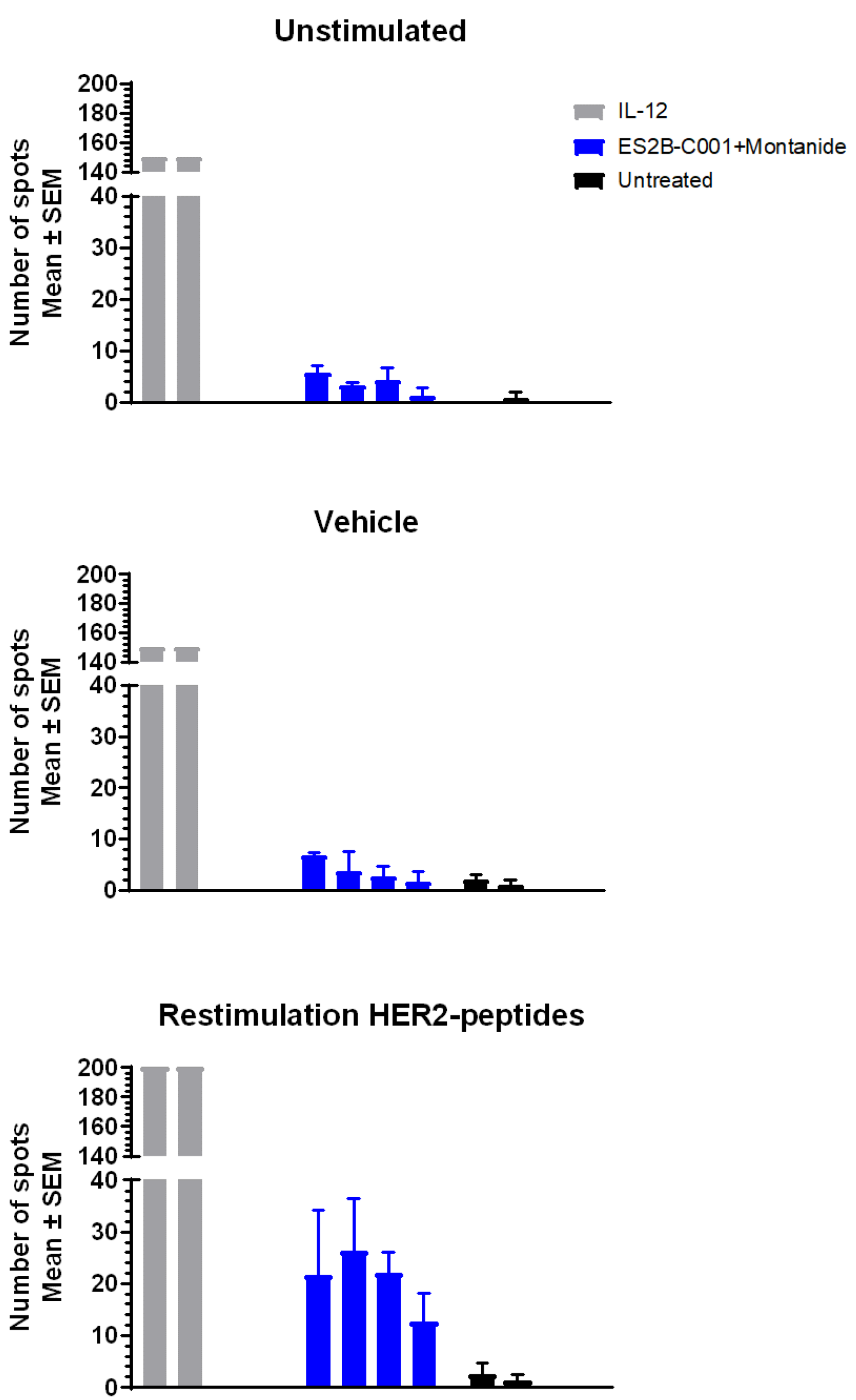
3.5. Cytokines and T Cell Responses
3.6. Prevention of Mammary Carcinoma Onset in Transgenic Mice
3.7. Therapy of Lung Metastases in Delta16 Mice Activity
3.8. Safety Profile of ES2B-C001
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slamon, D.J.; Godolphin, W.; Jones, L.A.; Holt, J.A.; Wong, S.G.; Keith, D.E.; Levin, W.J.; Stuart, S.G.; Udove, J.; Ullrich, A.; et al. Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science 1989, 244, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Iqbal, N. Human epidermal growth factor receptor 2 (HER2) in cancers: Overexpression and therapeutic implications. Mol. Biol. Int. 2014, 2014, 852748. [Google Scholar] [CrossRef] [PubMed]
- Muscatello, L.V.; Di Oto, E.; Sarli, G.; Monti, V.; Pia Foschini, M.; Benazzi, C.; Brunetti, B. HER2 Amplification Status in Feline Mammary Carcinoma: A Tissue Microarray–Fluorescence In Situ Hydridization–Based Study. Vet. Pathol. 2019, 56, 230–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, M.; Ribeiro, R.; Najmudin, S.; Gameiro, A.; Rodrigues, R.V.; Cardoso, F.; Ferreira, F. Serum HER2 levels are increased in cats with mammary carcinomas and predict tissue HER2 status. Oncotarget 2016, 7, 17314–17326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, M.; Correia, J.; Peleteiro, M.C.; Ferreira, F. St Gallen molecular subtypes in feline mammary carcinoma and paired metastases—Disease progression and clinical implications from a 3-year follow-up study. Tumor Biol. 2015, 37, 4053–4064. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, B.; Bacci, B.; Sarli, G.; Pancioni, E.; Muscatello, L. Immunohistochemical Screening of HER2 in Canine Carcinomas: A Preliminary Study. Animals 2021, 11, 1006. [Google Scholar] [CrossRef]
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human breast cancer: Correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Ménard, S.; Fortis, S.; Castiglioni, F.; Agresti, R.; Balsari, A. HER2 as a prognostic factor in breast cancer. Oncology 2001, 61, 67–72. [Google Scholar] [CrossRef]
- Baselga, J.; Carbonell, X.; Castañeda-Soto, N.J.; Clemens, M.; Green, M.; Harvey, V.; Morales, S.; Barton, C.; Ghahramani, P. Phase II study of efficacy, safety, and pharmacokinetics of trastuzumab monotherapy administered on a 3-weekly schedule. J. Clin. Oncol. 2005, 23, 2162–2171. [Google Scholar] [CrossRef]
- Garg, A.; Quartino, A.; Li, J.; Jin, J.; Wada, D.R.; Li, H.; Cortés, J.; McNally, V.; Ross, G.; Visich, J.; et al. Population pharmacokinetic and covariate analysis of pertuzumab, a HER2-targeted monoclonal antibody, and evaluation of a fixed, non-weight-based dose in patients with a variety of solid tumors. Cancer Chemother. Pharmacol. 2014, 74, 819–829. [Google Scholar] [CrossRef]
- Vogel, C.L.; Cobleigh, M.A.; Tripathy, D.; Gutheil, J.C.; Harris, L.N.; Fehrenbacher, L.; Slamon, D.J.; Murphy, M.; Novotny, W.F.; Burchmore, M.; et al. Efficacy and safety of trastuzumab as a single agent in first-line treatment of HER2-overexpressing metastatic breast cancer. J. Clin. Oncol. 2002, 20, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Nami, B.; Maadi, H.; Wang, Z. Mechanisms Underlying the Action and Synergism of Trastuzumab and Pertuzumab in Targeting HER2-Positive Breast Cancer. Cancers 2018, 10, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhães, P.R.; Machuqueiro, M.; Almeida, J.G.; Melo, A.; Cordeiro, M.N.D.S.; Verde, S.C.; Gümüş, Z.H.; Moreira, I.S.; Correia, J.D.G. Dynamical Rearrangement of Human Epidermal Growth Factor Receptor 2 upon Antibody Binding: Effects on the Dimerization. Biomolecules 2019, 9, 706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, D.-Y.; Bang, Y.-J. HER2-targeted therapies—A role beyond breast cancer. Nat. Rev. Clin. Oncol. 2019, 17, 33–48. [Google Scholar] [CrossRef]
- Nahta, R.; Esteva, F.J. Herceptin: Mechanisms of action and resistance. Cancer Lett. 2006, 232, 123–138. [Google Scholar] [CrossRef]
- Clynes, R.A.; Towers, T.L.; Presta, L.G.; Ravetch, J.V. Inhibitory Fc receptors modulate in vivo cytotoxicity against tumor targets. Nat. Med. 2000, 6, 443–446. [Google Scholar] [CrossRef]
- Capelan, M.; Pugliano, L.; De Azambuja, E. Pertuzumab: New hope for patients with HER2–positive breast cancer. Annals. Oncol. 2013, 24, 273–282. [Google Scholar] [CrossRef]
- Lilienthal, G.-M.; Rahmöller, J.; Petry, J.; Bartsch, Y.C.; Leliavski, A.; Ehlers, M. Potential of Murine IgG1 and Human IgG4 to Inhibit the Classical Complement and Fcγ Receptor Activation Pathways. Front. Immunol. 2018, 9, 958. [Google Scholar] [CrossRef] [Green Version]
- Temming, A.R.; Bentlage, A.E.; de Taeye, S.W.; Bosman, G.P.; Lissenberg-Thunnissen, S.N.; Derksen, N.I.; Brasser, G.; Mok, J.Y.; van Esch, W.J.; Howie, H.L.; et al. Cross-reactivity of mouse IgG subclasses to human Fc gamma receptors: Antibody deglycosylation only eliminates IgG2b binding. Mol. Immunol. 2020, 127, 79–86. [Google Scholar] [CrossRef]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG Subclasses and Allotypes: From Structure to Effector Functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef]
- Vernieri, C.; Milano, M.; Brambilla, M.; Mennitto, A.; Maggi, C.; Cona, M.S.; Prisciandaro, M.; Fabbroni, C.; Celio, L.; Mariani, G.; et al. Resistance mechanisms to anti-HER2 therapies in HER2-positive breast cancer: Current knowledge, new research directions and therapeutic perspectives. Crit. Rev. Oncol. Hematol. 2019, 139, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Shu, M.; Yan, H.; Xu, C.; Wu, Y.; Chi, Z.; Nian, W.; He, Z.; Xiao, J.; Wei, H.; Zhou, Q.; et al. A novel anti-HER2 antibody GB235 reverses Trastuzumab resistance in HER2-expressing tumor cells in vitro and in vivo. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cortés, J.; Fumoleau, P.; Bianchi, G.V.; Petrella, T.M.; Gelmon, K.; Pivot, X.; Verma, S.; Albanell, J.; Conte, P.; Lluch, A.; et al. Pertuzumab monotherapy after trastuzumab-based treatment and subsequent reintroduction of trastuzumab: Activity and tolerability in patients with advanced human epidermal growth factor receptor 2-positive breast cancer. J. Clin. Oncol. 2012, 30, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- Palladini, A.; Thrane, S.; Janitzek, C.M.; Pihl, J.; Clemmensen, S.B.; De Jongh, W.A.; Clausen, T.M.; Nicoletti, G.; Landuzzi, L.; Penichet, M.L.; et al. Virus-like particle display of HER2 induces potent anti-cancer responses. OncoImmunology 2017, 7, e1408749. [Google Scholar] [CrossRef] [Green Version]
- Clay, T.M.; Osada, T.; Hartman, Z.C.; Hobeika, A.; Devi, G.; Morse, M.A.; Lyerly, H.K. Polyclonal immune responses to antigens associated with cancer signaling pathways and new strategies to enhance cancer vaccines. Immunol. Res. 2010, 49, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Triulzi, C.; Vertuani, S.; Curcio, C.; Antognoli, A.; Seibt, J.; Akusjärvi, G.; Wei, W.Z.; Cavallo, F.; Kiessling, R. Antibody-dependent natural killer cell-mediated cytotoxicity engendered by a kinase-inactive human HER2 adenovirus-based vaccination mediates resistance to breast tumors. Cancer Res. 2010, 70, 7431–7441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, A.F.; Lollini, P.L. Virus-like antigen display for cancer vaccine development, what is the potential? Expert. Rev. Vaccines. 2018, 17, 285–288. [Google Scholar] [CrossRef] [Green Version]
- Castagnoli, L.; Ladomery, M.; Tagliabue, E.; Pupa, S.M. The d16HER2 Splice Variant: A Friend or Foe of HER2-Positive Cancers? Cancers 2019, 11, 902. [Google Scholar] [CrossRef] [Green Version]
- Marchini, C.; Gabrielli, F.; Iezzi, M.; Zenobi, S.; Montani, M.; Pietrella, L.; Kalogris, C.; Rossini, A.; Ciravolo, V.; Castagnoli, L.; et al. The Human Splice Variant Δ16HER2 Induces Rapid Tumor Onset in a Reporter Transgenic Mouse. PLoS ONE 2011, 6, e18727. [Google Scholar] [CrossRef]
- Landuzzi, L.; Palladini, A.; Ceccarelli, C.; Asioli, S.; Nicoletti, G.; Giusti, V.; Ruzzi, F.; Ianzano, M.L.; Scalambra, L.; Laranga, R.; et al. Early stability and late random tumor progression of a HER2-positive primary breast cancer patient-derived xenograft. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Nanni, P.; Nicoletti, G.; De Giovanni, C.; Landuzzi, L.; Di Carlo, E.; Cavallo, F.; Pupa, S.M.; Rossi, I.; Colombo, M.P.; Ricci, C.; et al. Combined allogeneic tumor cell vaccination and systemic interleukin 12 prevents mammary carcinogenesis in HER-2/neu transgenic mice. J. Exp. Med. 2001, 194, 1195–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, M.; Nechansky, A.; Kircheis, R. Cancer immunotherapy. Biotechnol. J. 2006, 1, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Milani, A.; Sangiolo, D.; Montemurro, F.; Aglietta, M.; Valabrega, G. Active immunotherapy in HER2 overexpressing breast cancer: Current status and future perspectives. Ann. Oncol. 2013, 24, 1740–1748. [Google Scholar] [CrossRef] [PubMed]
- Piechocki, M.P.; Ho, Y.-S.; Pilon, S.; Wei, W.-Z. Human ErbB-2 (Her-2) Transgenic Mice: A Model System for Testing Her-2 Based Vaccines. J. Immunol. 2003, 171, 5787–5794. [Google Scholar] [CrossRef] [Green Version]
- Palladini, A.; Nicoletti, G.; Lamolinara, A.; Dall’Ora, M.; Balboni, T.; Ianzano, M.L.; Laranga, R.; Landuzzi, L.; Giusti, V.; Ceccarelli, C.; et al. HER2 isoforms co-expression differently tunes mammary tumor phenotypes affecting onset, vasculature and therapeutic response. Oncotarget. 2017, 8, 54444–54458. [Google Scholar] [CrossRef]
- Palladini, A.; Nicoletti, G.; Pappalardo, F.; Murgo, A.; Grosso, V.; Stivani, V.; Ianzano, M.L.; Antognoli, A.; Croci, S.; Landuzzi, L.; et al. In silico modeling and in vivo efficacy of cancer-preventive vaccinations. Cancer Res. 2010, 70, 7755–7763. [Google Scholar] [CrossRef] [Green Version]
- Veronesi, U.; Cascinelli, N.; Mariani, L.; Greco, M.; Saccozzi, R.; Luini, A.; Aguilar, M.; Marubini, E. Twenty-Year Follow-up of a Randomized Study Comparing Breast-Conserving Surgery with Radical Mastectomy for Early Breast Cancer. New Engl. J. Med. 2002, 347, 1227–1232. [Google Scholar] [CrossRef]
- Negoita, S.; Ramirez-Pena, E. Prevention of Late Recurrence: An Increasingly Important Target for Breast Cancer Research and Control. JNCI: J. Natl. Cancer Inst. 2021, 114, 340–341. [Google Scholar] [CrossRef]
- Pedersen, R.N.; Esen, B.Ö.; Mellemkjær, L.; Christiansen, P.; Ejlertsen, B.; Lash, T.L.; Nørgaard, M.; Cronin-Fenton, D. The Incidence of Breast Cancer Recurrence 10-32 Years After Primary Diagnosis. J. Natl. Cancer. Inst. 2022, 114, 391–399. [Google Scholar] [CrossRef]
- Yersal, O.; Barutca, S. Biological subtypes of breast cancer: Prognostic and therapeutic implications. World J. Clin. Oncol. 2014, 5, 412–424. [Google Scholar] [CrossRef]
- Wang, L.T.; Pereira, L.S.; Kiyuka, P.K.; Schön, A.; Kisalu, N.K.; Vistein, R.; Dillon, M.; Bonilla, B.G.; Molina-Cruz, A.; Barillas-Mury, C.; et al. Protective effects of combining monoclonal antibodies and vaccines against the Plasmodium falciparum circumsporozoite protein. PLOS Pathog. 2021, 17, e1010133. [Google Scholar] [CrossRef] [PubMed]
- Rovero, S.; Amici, A.; Di Carlo, E.; Bei, R.; Nanni, P.; Quaglino, E.; Porcedda, P.; Boggio, K.; Smorlesi, A.; Lollini, P.L.; et al. DNA vaccination against rat her-2/Neu p185 more effectively inhibits carcinogenesis than transplantable carcinomas in transgenic BALB/c mice. J. Immunol. 2000, 165, 5133–5142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randy, C.Q.; Behrens, E.M. Cytokine Storm Syndrome; Springer Nature: Cham, Switzerland, 2019. [Google Scholar]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr. Approaching the Asymptote? Evolution and Revolution in Immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 54, 1–13. [Google Scholar] [CrossRef]
- Ascarateil, S.; Puget, A.; Koziol, M.-E. Safety data of Montanide ISA 51 VG and Montanide ISA 720 VG, two adjuvants dedicated to human therapeutic vaccines. J. Immunother. Cancer 2015, 3, P428. [Google Scholar] [CrossRef] [Green Version]
- Jegerlehner, A.; Wiesel, M.; Dietmeier, K.; Zabel, F.; Gatto, D.; Saudan, P.; Bachmann, M.F. Carrier induced epitopic suppression of antibody responses induced by virus-like particles is a dynamic phenomenon caused by carrier-specific antibodies. Vaccine 2010, 28, 5503–5512. [Google Scholar] [CrossRef] [PubMed]
- Fougeroux, C.; Goksøyr, L.; Idorn, M.; Soroka, V.; Myeni, S.K.; Dagil, R.; Janitzek, C.M.; Søgaard, M.; Aves, K.L.; Horsted, E.W. Capsid-like particles decorated with the SARS-CoV-2 receptor-binding domain elicit strong virus neutralization activity. Nat. Commun. 2021, 12, 324. [Google Scholar] [CrossRef]
- Cardoso, F.; Piccart, M.J.; Durbecq, V.; Di Leo, A. Resistance to Trastuzumab: A Necessary Evil or a Temporary Challenge? Clin. Breast Cancer 2002, 3, 247–257. [Google Scholar] [CrossRef]
- Wang, R.-X.; Chen, S.; Jin, X.; Chen, C.-M.; Shao, Z.-M. Weekly paclitaxel plus carboplatin with or without trastuzumab as neoadjuvant chemotherapy for HER2-positive breast cancer: Loss of HER2 amplification and its impact on response and prognosis. Breast Cancer Res. Treat. 2016, 161, 259–267. [Google Scholar] [CrossRef]
- Giusti, V.; Ruzzi, F.; Landuzzi, L.; Ianzano, M.L.; Laranga, R.; Nironi, E.; Scalambra, L.; Nicoletti, G.; De Giovanni, C.; Olivero, M.; et al. Evolution of HER2-positive mammary carcinoma: HER2 loss reveals claudin-low traits in cancer progression. Oncogenesis 2021, 10, 1–12. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruzzi, F.; Palladini, A.; Clemmensen, S.; Strøbæk, A.; Buijs, N.; Domeyer, T.; Dorosz, J.; Soroka, V.; Grzadziela, D.; Rasmussen, C.J.; et al. Prevention and Therapy of Metastatic HER-2+ Mammary Carcinoma with a Human Candidate HER-2 Virus-like Particle Vaccine. Biomedicines 2022, 10, 2654. https://doi.org/10.3390/biomedicines10102654
Ruzzi F, Palladini A, Clemmensen S, Strøbæk A, Buijs N, Domeyer T, Dorosz J, Soroka V, Grzadziela D, Rasmussen CJ, et al. Prevention and Therapy of Metastatic HER-2+ Mammary Carcinoma with a Human Candidate HER-2 Virus-like Particle Vaccine. Biomedicines. 2022; 10(10):2654. https://doi.org/10.3390/biomedicines10102654
Chicago/Turabian StyleRuzzi, Francesca, Arianna Palladini, Stine Clemmensen, Anette Strøbæk, Nicolaas Buijs, Tanja Domeyer, Jerzy Dorosz, Vladislav Soroka, Dagmara Grzadziela, Christina Jo Rasmussen, and et al. 2022. "Prevention and Therapy of Metastatic HER-2+ Mammary Carcinoma with a Human Candidate HER-2 Virus-like Particle Vaccine" Biomedicines 10, no. 10: 2654. https://doi.org/10.3390/biomedicines10102654