
Atomic Force Microscopy Study of the Temperature and Storage Duration Dependencies of Horseradish Peroxidase Oligomeric State
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Protein
2.2. Experiment Design
2.3. Sample Preparation for AFM Experiments
- (1)
- The series for the determination of the effect of temperature
- (2)
- The series for the determination of the effect of storage duration
- (3)
- Blank experiments
2.4. AFM Scanning
2.5. AFM Data Processing
2.6. Spectrophotometric Measurements
3. Results
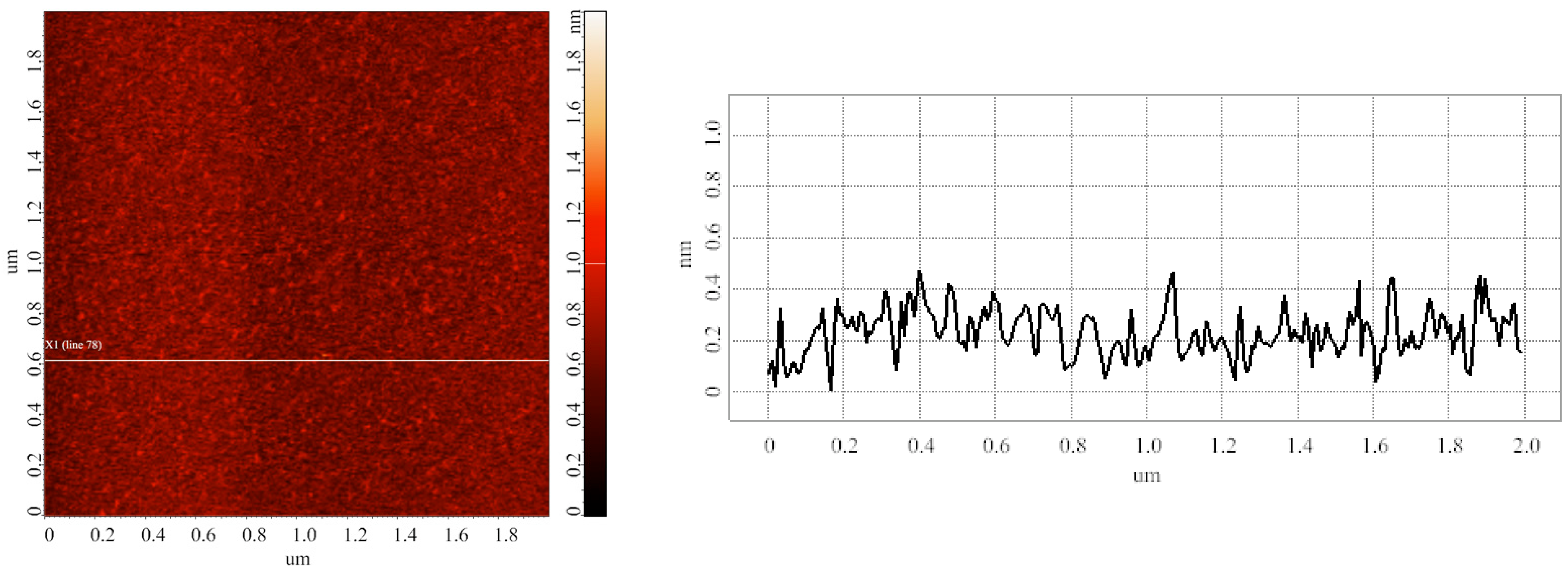
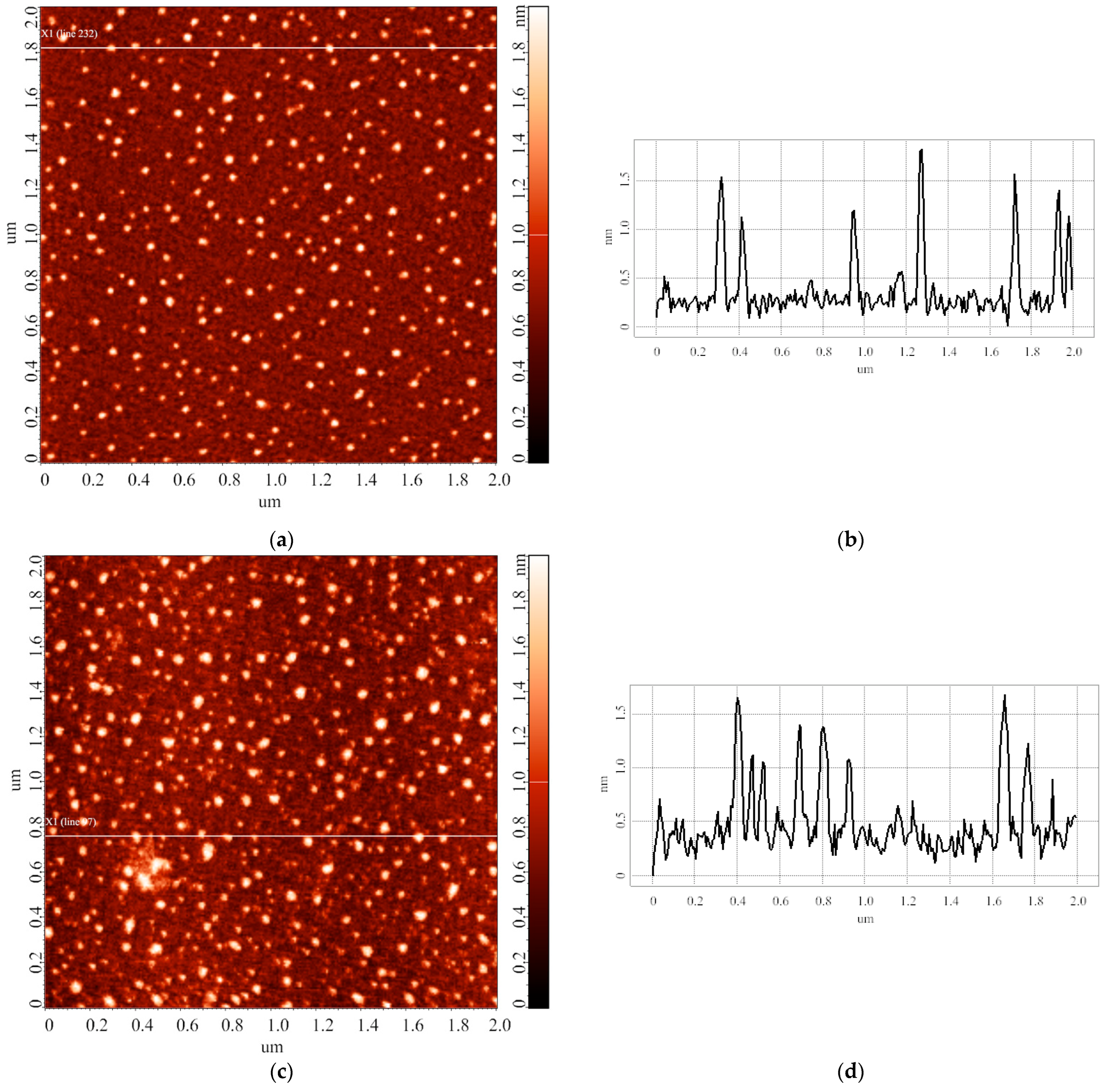
3.1. Blank Experiments
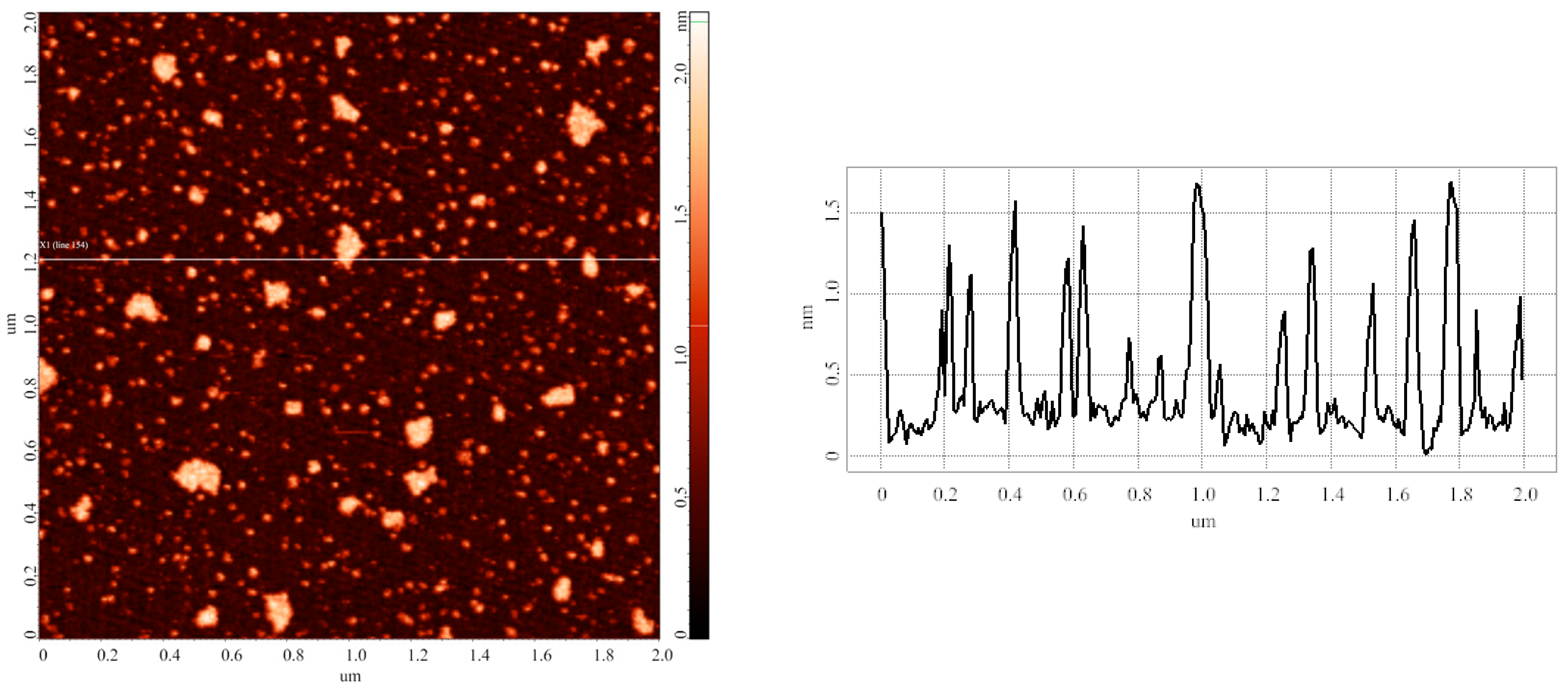
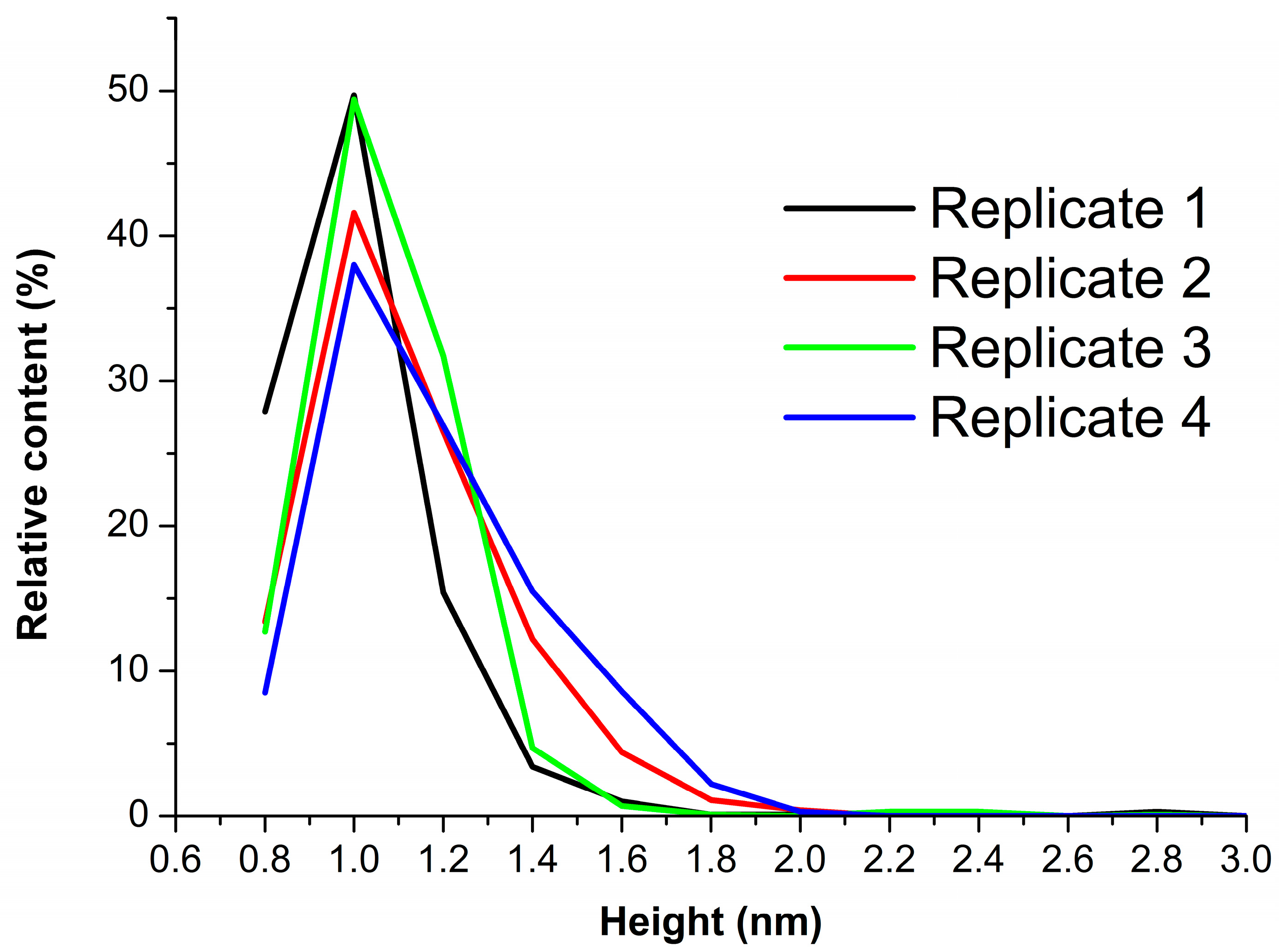
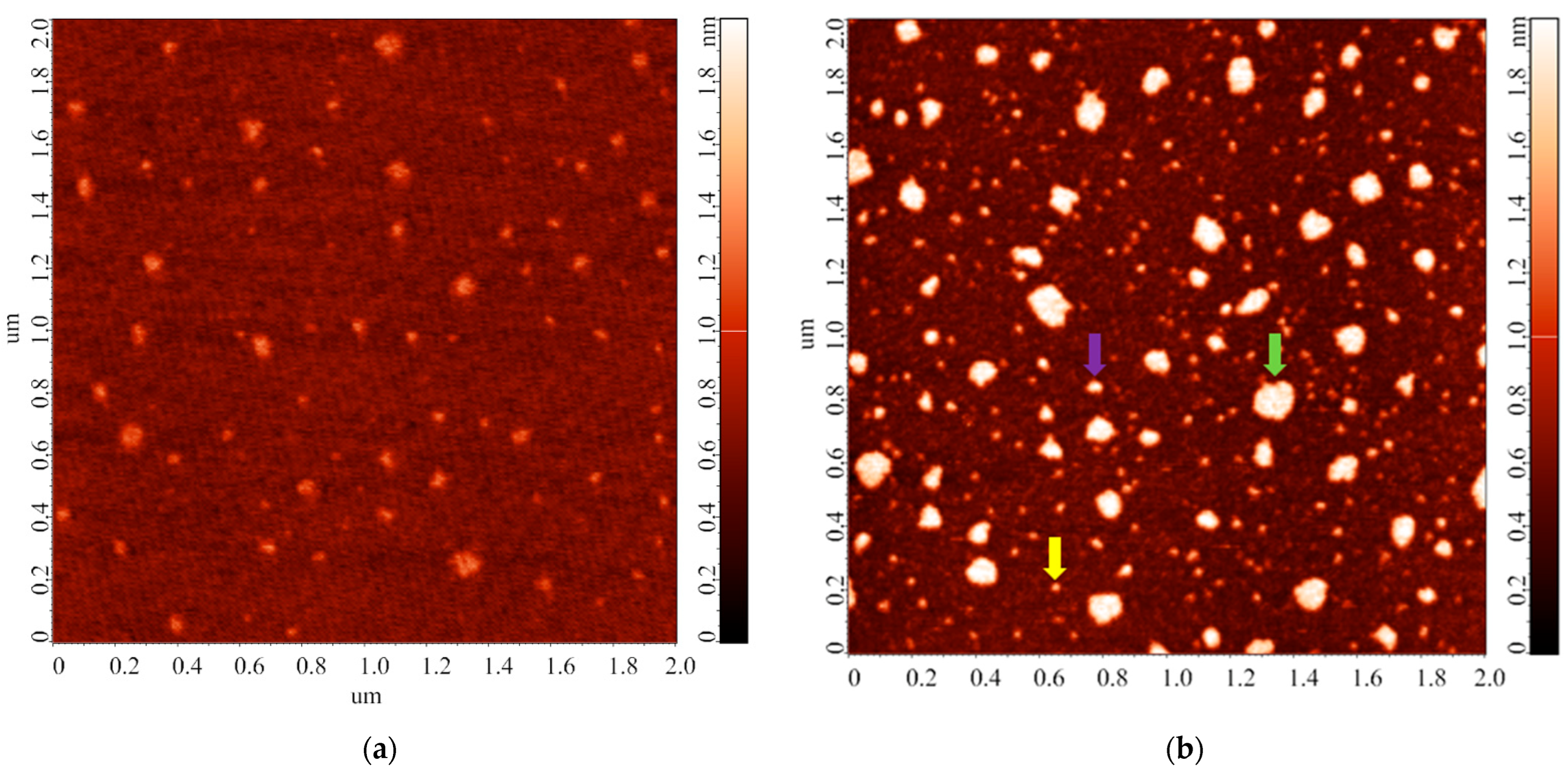
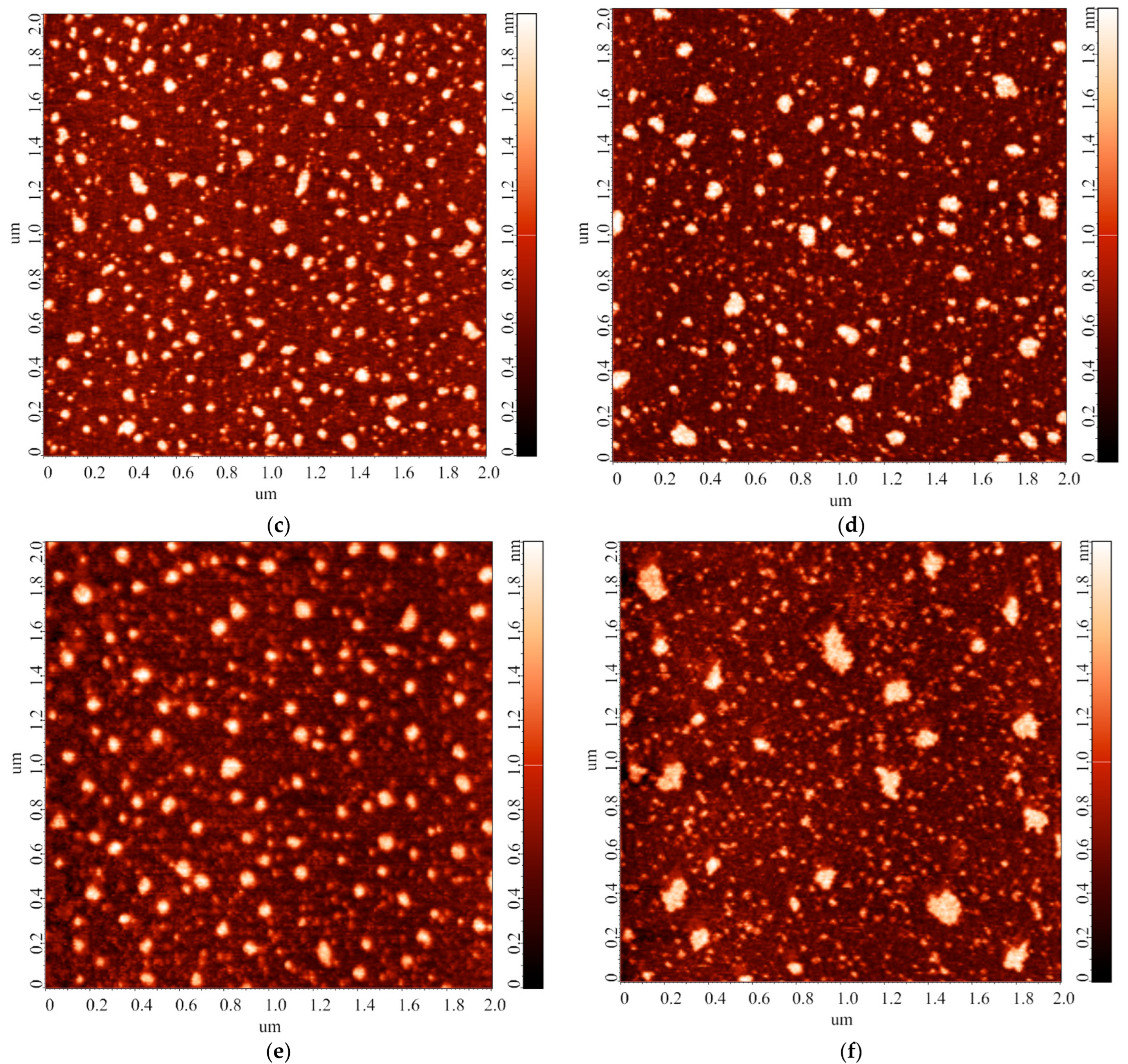
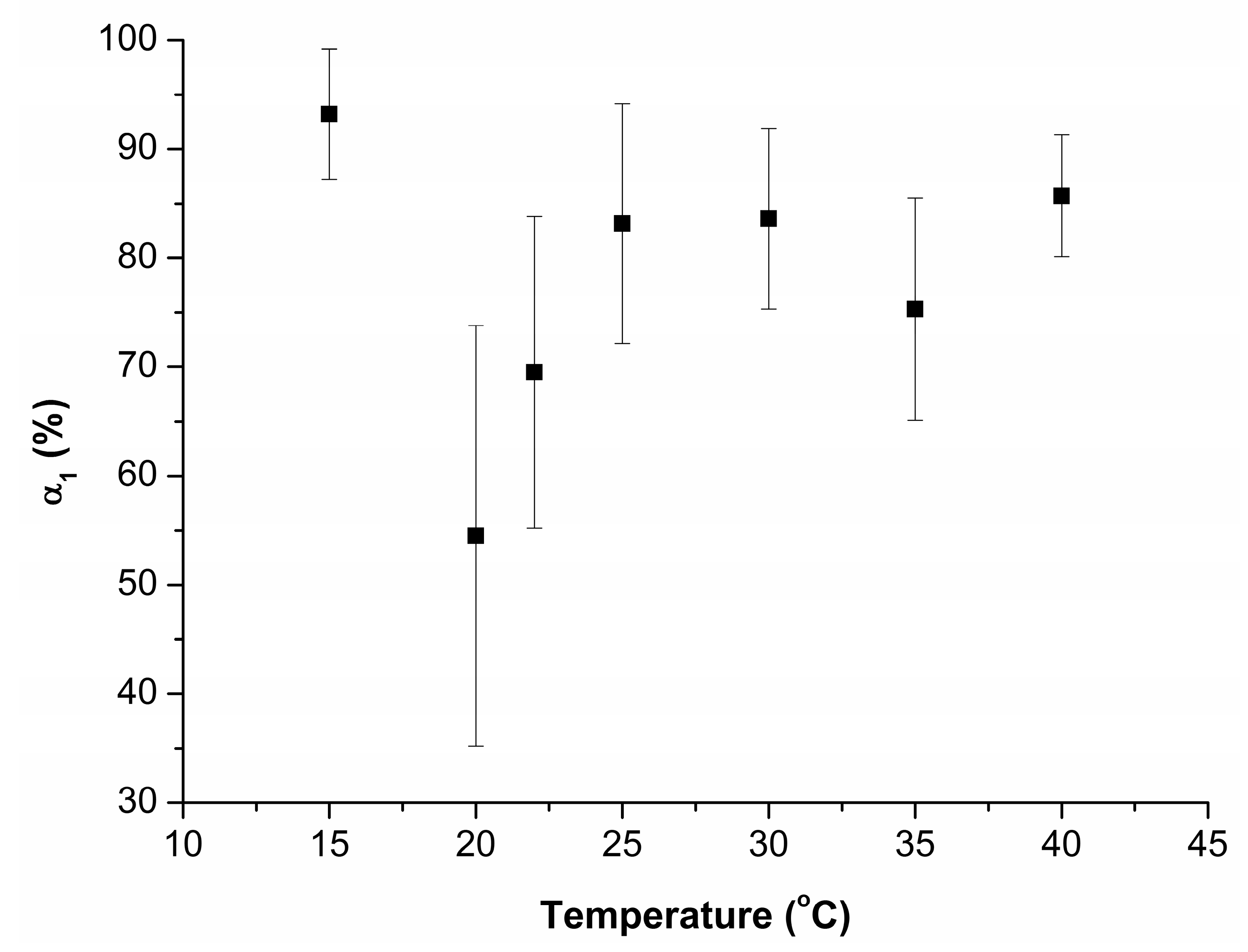
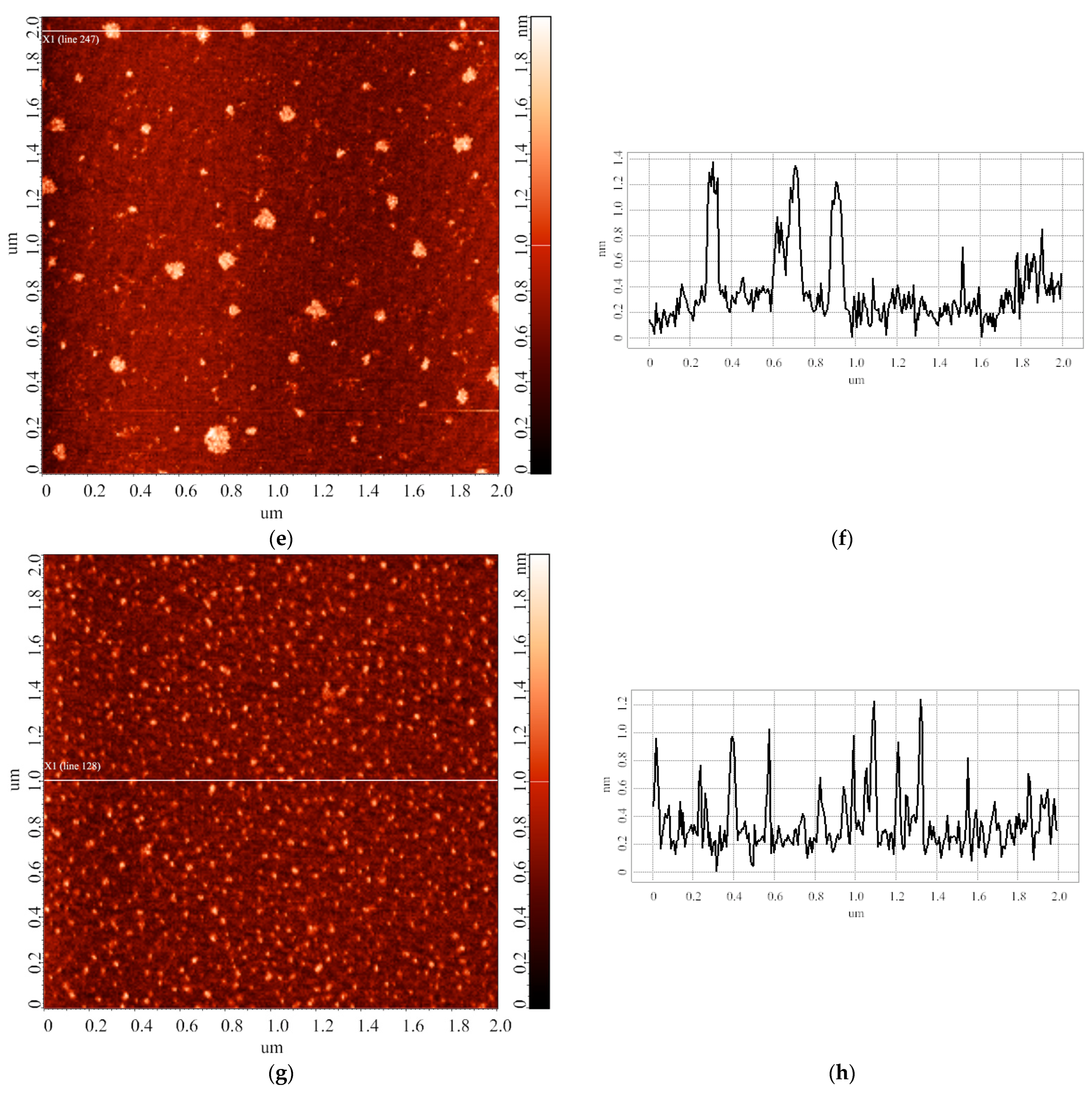
3.2. Effect of Incubation Temperature of HRP Solution on Enzyme Aggregation State on Mica
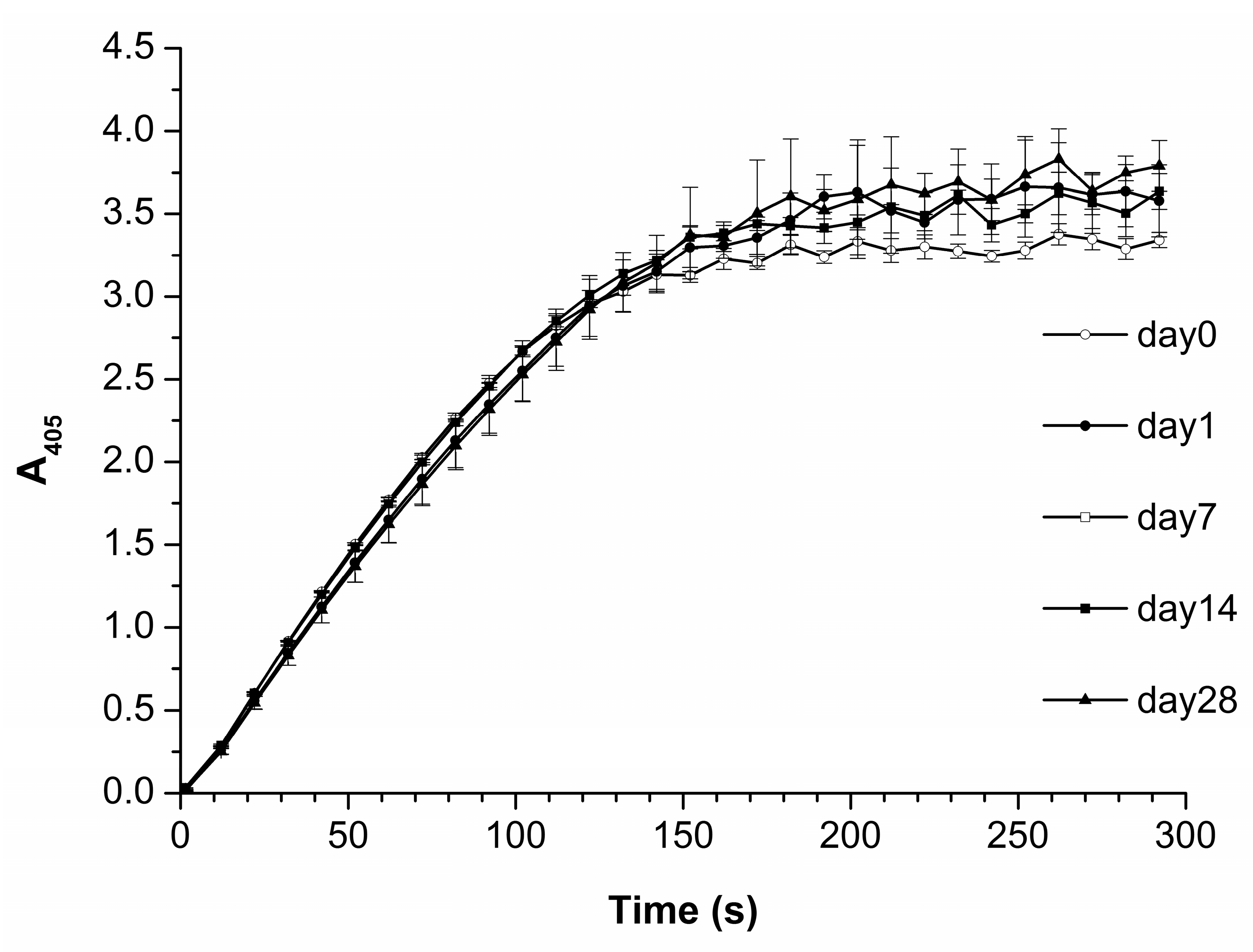
3.3. Effect of Storage Duration of HRP Solution on Enzyme Aggregation State on Mica
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kishore, D.; Kundu, S.; Kayastha, A.M. Thermal, chemical and pH induced denaturation of a multimeric β-galactosidase reveals multiple unfolding pathways. PLoS ONE 2012, 7, e50380. [Google Scholar] [CrossRef] [Green Version]
- Robinson, P.K. Enzymes: Principles and biotechnological applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef]
- Ivanov, Y.D.; Bukharina, N.S.; Frantsuzov, P.A.; Pleshakova, T.O.; Kanashenko, S.L.; Medvedeva, N.V.; Argentova, V.V.; Zgoda, V.G.; Munro, A.W.; Archakov, A.I. AFM study of cytochrome CYP102A1 oligomeric state. Soft Matter 2012, 8, 4602–4608. [Google Scholar] [CrossRef]
- Koshland, D.E. Correlation of Structure and Function in Enzyme Action. Science 1963, 142, 1533–1541. [Google Scholar] [CrossRef]
- Dalby, P.A. Optimising enzyme function by directed evolution. Curr. Opin. Struct. Biol. 2003, 13, 500–505. [Google Scholar] [CrossRef]
- Keleti, T. Mechanism of Action and Regulation of Enzymes; American Elsevier: New York, NY, USA, 1975. [Google Scholar]
- Gouda, M.D.; Singh, S.A.; Appu Rao, A.G.; Thakur, M.S.; Karanth, N.G. Thermal inactivation of glucose oxidase: Mechanism and stabilization using additives. J. Biol. Chem. 2003, 278, 24324–24333. [Google Scholar] [CrossRef] [Green Version]
- Gulla, K.C.; Gouda, M.D.; Thakur, M.S.; Karanth, N.G. Enhancement of stability of immobilized glucose oxidase by modification of free thiols generated by reducing disulfide bonds and using additives. Biosens. Bioelectron. 2004, 19, 621–625. [Google Scholar] [CrossRef]
- Altikatoglu, M.; Basaran, Y. Additive effect of dextrans on the stability of horseradish peroxidase. Protein J. 2011, 30, 84–90. [Google Scholar] [CrossRef]
- Mozhaev, V.V. Mechanism-based strategies for protein thermostabilization. Trends Biotechnol. 1993, 11, 88–95. [Google Scholar] [CrossRef]
- Goryanova, B.; Goldman, L.M.; Ming, S.; Amyes, T.L.; Gerlt, J.A.; Richard, J.P. Rate and Equilibrium Constants for an Enzyme Conformational Change during Catalysis by Orotidine 5′-Monophosphate Decarboxylase. Biochemistry 2015, 54, 4555–4564. [Google Scholar] [CrossRef]
- Ivanov, Y.D.; Pleshakova, T.O.; Shumov, I.D.; Kozlov, A.F.; Ivanova, I.A.; Valueva, A.A.; Tatur, V.Y.; Smelov, M.V.; Ivanova, N.D.; Ziborov, V.S. AFM Imaging of Protein Aggregation in Studying the Impact of Knotted Electromagnetic Field on A Peroxidase. Sci. Rep. 2020, 10, 9022. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, Y.D.; Pleshakova, T.O.; Shumov, I.D.; Kozlov, A.F.; Ivanova, I.A.; Valueva, A.A.; Ershova, M.O.; Tatur, V.Y.; Stepanov, I.N.; Repnikov, V.V.; et al. AFM study of changes in properties of horseradish peroxidase after incubation of its solution near a pyramidal structure. Sci. Rep. 2021, 11, 9907. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, Y.D.; Tatur, V.Y.; Pleshakova, T.O.; Shumov, I.D.; Kozlov, A.F.; Valueva, A.A.; Ivanova, I.A.; Ershova, M.O.; Ivanova, N.D.; Repnikov, V.V.; et al. Effect of Spherical Elements of Biosensors and Bioreactors on the Physicochemical Properties of a Peroxidase Protein. Polymers 2021, 13, 1601. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; de Souza Santos, M.; Li, Y.; Tomchick, D.R.; Orth, K. High-resolution cryo-EM structures of the E. coli hemolysin ClyA oligomers. PLoS ONE 2019, 14, e0213423. [Google Scholar] [CrossRef] [Green Version]
- Vonck, J.; Mills, D.J. Advances in high-resolution cryo-EM of oligomeric enzymes. Curr. Opin. Struct. Biol. 2017, 46, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, Y.D.; Taldaev, A.; Lisitsa, A.V.; Ponomarenko, E.A.; Archakov, A.I. Prediction of Monomeric and Dimeric Structures of CYP102A1 Using AlphaFold2 and AlphaFold Multimer and Assessment of Point Mutation Effect on the Efficiency of Intra- and Interprotein Electron Transfer. Molecules 2022, 27, 1386. [Google Scholar] [CrossRef]
- Braga, P.C.; Ricci, D. Methods in molecular biology. In Atomic Force Microscopy. Biomedical Methods and Applications; Humana Press: Totowa, NJ, USA, 2003; Volume 242. [Google Scholar]
- Kuznetsov, V.Y.; Ivanov, Y.D.; Archakov, A.I. Atomic force microscopy revelation of molecular complexes in the multiprotein cytochrome P450 2B4-containing system. Proteomics 2004, 4, 2390–2396. [Google Scholar] [CrossRef]
- Ierardi, V.; Ferrera, F.; Millo, E.; Damonte, G.; Filaci, G.; Valbusa, U. Bioactive surfaces for antibody-antigen complex detection by Atomic Force Microscopy. J. Phys. Conf. Ser. 2013, 439, 012001. [Google Scholar] [CrossRef]
- Ramanaviciene, A.; Snitka, V.; Mieliauskiene, R.; Kazlauskas, R.; Ramanavicius, A. AFM-study of complement system assembly initiated by antigen-antibody complex. Open Chem. 2006, 4, 194–206. [Google Scholar] [CrossRef]
- Kiselyova, O.I.; Yaminsky, I.V.; Ivanov, Y.D.; Kanaeva, I.P.; Kuznetsov, V.Y.; Archakov, A.I. AFM study of membrane proteins, cytochrome P450 2B4 and NADPH-cytochrome P450 reductase, and of their complex formation. Arch. Biochem. Biophys. 1999, 371, 1–7. [Google Scholar] [CrossRef]
- Kuznetsov, V.Y.; Ivanov, Y.D.; Bykov, V.A.; Saunin, S.A.; Fedorov, I.A.; Lemeshko, S.V.; Hoa, H.B.; Archakov, A.I. Atomic force microscopy detection of molecular complexes in multiprotein P450cam-containing monooxygenase system. Proteomics 2002, 2, 1699–1705. [Google Scholar] [CrossRef]
- Archakov, A.I.; Ivanov, Y.D. Application of AFM and optical biosensor for investigation of complexes formed in P450-containing monooxygenase systems. Biochim. Biophys. Acta—Proteins Proteom. 2011, 1814, 102–110. [Google Scholar] [CrossRef]
- Ivanov, Y.D.; Frantsuzov, P.A.; Zöllner, A.; Medvedeva, N.V.; Archakov, A.I.; Reinle, W.; Bernhardt, R. Atomic Force Microscopy Study of Protein–Protein Interactions in the Cytochrome CYP11A1 (P450scc)-Containing Steroid Hydroxylase System. Nanoscale Res. Lett. 2011, 6, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, Y.D.; Pleshakova, T.O.; Shumov, I.D.; Kozlov, A.F.; Romanova, T.S.; Valueva, A.A.; Tatur, V.Y.; Stepanov, I.N.; Ziborov, V.S. Investigation of the Influence of Liquid Motion in a Flow-Based System on an Enzyme Aggregation State with an Atomic Force Microscopy Sensor: The Effect of Water Flow. Appl. Sci. 2020, 10, 4560. [Google Scholar] [CrossRef]
- Ziborov, V.S.; Pleshakova, T.O.; Shumov, I.D.; Kozlov, A.F.; Ivanova, I.A.; Valueva, A.A.; Tatur, V.Y.; Negodailov, A.N.; Lukyanitsa, A.A.; Ivanov, Y.D. Investigation of the Influence of Liquid Motion in a Flow-Based System on an Enzyme Aggregation State with an Atomic Force Microscopy Sensor: The Effect of Glycerol Flow. Appl. Sci. 2020, 10, 4825. [Google Scholar] [CrossRef]
- Ivanov, Y.D.; Pleshakova, T.O.; Shumov, I.D.; Kozlov, A.F.; Valueva, A.A.; Ivanova, I.A.; Ershova, M.O.; Larionov, D.I.; Repnikov, V.V.; Ivanova, N.D.; et al. AFM and FTIR Investigation of the Effect of Water Flow on Horseradish Peroxidase. Molecules 2021, 26, 306. [Google Scholar] [CrossRef]
- Ivanov, Y.D.; Pleshakova, T.O.; Shumov, I.D.; Kozlov, A.F.; Ivanova, I.A.; Ershova, M.O.; Tatur, V.Y.; Ziborov, V.S. AFM Study of the Influence of Glycerol Flow on Horseradish Peroxidase near the in/out Linear Sections of a Coil. Appl. Sci. 2021, 11, 1723. [Google Scholar] [CrossRef]
- Veitch, N.C.; Smith, A.T. Horseradish Peroxidase. Adv. Inorg. Chem. 2000, 51, 107–162. [Google Scholar] [CrossRef]
- Shannon, L.M.; Kay, E.; Lew, J.Y. Peroxidase Isozymes from Horseradish Roots. J. Biol. Chem. 1966, 241, 2166–2172. [Google Scholar] [CrossRef]
- Hassani, L.; Ranjbar, B.; Khajeh, K.; Naderi-Manesh, H.; Naderi-Manesh, M.; Sadeghi, M. Horseradish peroxidase thermostabilization: The combinatorial effects of the surface modification and the polyols. Enzym. Microb. Technol. 2006, 38, 118–125. [Google Scholar] [CrossRef]
- Gajhede, M.; Schuller, D.J.; Henriksen, A.; Smith, A.T.; Poulos, T.L. Crystal structure of horseradish peroxidase C at 2.15 Å resolution. Nat. Struct. Mol. Biol. 1997, 4, 1032–1038. [Google Scholar] [CrossRef]
- Bilal, M.; Barceló, D.; Iqbal, H.M. Nanostructured materials for harnessing the power of horseradish peroxidase for tailored environmental applications. Sci. Total Environ. 2020, 749, 142360. [Google Scholar] [CrossRef] [PubMed]
- Duan, N.; Li, C.; Song, M.; Wang, Z.; Zhu, C.; Wu, S. Signal amplification of SiO2 nanoparticle loaded horseradish peroxidase for colorimetric detection of lead ions in water. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2022, 265, 120342. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.; Kale, P.; Balasubramanian, P. Inhibition assays of horseradish peroxidase by hexavalent chromium and other heavy metals. Int. J. Environ. Anal. Chem. 2020, 1–13. [Google Scholar] [CrossRef]
- Rahemi, V.; Trashin, S.; Hafideddine, Z.; van Doorslaer, S.; Meynen, V.; Gorton, L.; de Wael, K. Amperometric Flow-Injection Analysis of Phenols Induced by Reactive Oxygen Species Generated under Daylight Irradiation of Titania Impregnated with Horseradish Peroxidase. Anal. Chem. 2020, 92, 3643–3649. [Google Scholar] [CrossRef]
- Huddleston, S.; Robertson, S.; Dobson, C.; Kwong, F.Y.P.; Charalambous, B.M. Structural and functional stability of horseradish peroxidase. Biochem. Soc. Trans. 1995, 23, 108S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://www.sigmaaldrich.com/RU/en/product/sigma/p6782 (accessed on 22 August 2022).
- Costa, S.A.; Tzanov, T.; Carneiro, A.F.; Paar, A.; Gübitz, G.M.; Cavaco-Paulo, A. Studies of stabilization of native catalase using additives. Enzym. Microb. Technol. 2002, 30, 387–391. [Google Scholar] [CrossRef] [Green Version]
- Pleshakova, T.O.; Kaysheva, A.L.; Shumov, I.D.; Ziborov, V.S.; Bayzyanova, J.M.; Konev, V.A.; Uchaikin, V.F.; Archakov, A.I.; Ivanov, Y.D. Detection of hepatitis C virus core protein in serum using aptamer-functionalized AFM chips. Micromachines 2019, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Sanders, S.A.; Bray, R.C.; Smith, A.T. pH-Dependent properties of a mutant horseradish peroxidase isoenzyme C in which Arg38 has been replaced with lysine. Eur. J. Biochem. 1994, 224, 1029–1037. [Google Scholar] [CrossRef]
- Enzymatic Assay of Peroxidase (EC 1.11.1.7) 2,2′-Azino-Bis(3-Ethylbenzthiazoline-6-Sulfonic Acid) as a Substrate Sigma Prod. No. P-6782. Available online: https://www.sigmaaldrich.com/RU/en/technical-documents/protocol/protein-biology/enzyme-activity-assays/enzymatic-assay-of-peroxidase-abts-as-substrate (accessed on 18 February 2022).
- Davies, P.; Rennke, H.; Cotran, R. Influence of molecular charge upon the endocytosis and intracellular fate of peroxidase activity in cultured arterial endothelium. J. Cell Sci. 1981, 49, 69–86. [Google Scholar] [CrossRef]
- Welinder, K.G. Amino Acid Sequence Studies of Horseradish Peroxidase. Amino and Carboxyl Termini, Cyanogen Bromide and Tryptic Fragments, the Complete Sequence, and Some Structural Characteristics of Horseradish Peroxidase C. JBIC J. Biol. Inorg. Chem. 1979, 96, 483–502. [Google Scholar] [CrossRef] [PubMed]
- Unger, B.P.; Gunsalus, I.C.; Sligar, S.G. Nucleotide sequence of the Pseudomonas putida cytochrome P-450cam gene and its expression in Escherichia coli. J. Biol. Chem. 1986, 261, 1158–1163. [Google Scholar] [CrossRef]
- Chu, J.W.; Kimura, T. Studies on adrenal steroid hydroxylases. Molecular and catalytic properties of adrenodoxin reductase (a flavoprotein). J. Biol. Chem. 1973, 248, 2089–2094. [Google Scholar] [CrossRef]
- Ivanov, Y.D.; Danichev, V.V.; Pleshakova, T.O.; Shumov, I.D.; Ziborov, V.S.; Krokhin, N.V.; Zagumenniy, M.N.; Ustinov, V.S.; Smirnov, L.P.; Shironin, A.V.; et al. Irreversible chemical AFM-based fishing for detection of low-copied proteins. Biochem. (Moscow) Suppl. Ser. B Biomed. Chem. 2013, 7, 46–61. [Google Scholar] [CrossRef]
- Kiselyova, O.I.; Yaminsky, I.V. Atomic Force Microscopy of Protein Complexes. In Atomic Force Microscopy. Methods in Molecular Biology; Braga, P.C., Ricci, D., Eds.; Humana Press: Totowa, NJ, USA, 2004; Volume 242, pp. 217–230. [Google Scholar]
- Bukharina, N.S.; Ivanov, Y.D.; Pleshakova, T.O.; Frantsuzov, P.A.; Ivanova, N.D.; Krohin, N.V.; Petushkova, N.A.; Archakov, A.I. Atomic force microscopy monitoring of the temperature dependence of the cytochrome BM3 oligomeric state. Biophysics 2015, 60, 66–72. [Google Scholar] [CrossRef]
- Maurer, S.C.; Schulze, H.; Schmid, R.D.; Urlacher, V. Immobilisation of P450 BM-3 and an NADP+ cofactor recycling system: Towards a technical appication of heme-containing monooxygenases in fine chemical synthesis. Adv. Synth. Catal. 2003, 345, 802–810. [Google Scholar] [CrossRef] [Green Version]
- Jamakhandi, A.P.; Jeffus, B.C.; Dass, V.R.; Miller, G.P. Thermal inactivation of the reductase domain of cytochrome P450 BM3. Arch. Biochem. Biophys. 2005, 439, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Day, R.; Bennion, B.J.; Ham, S.; Daggett, V. Increasing temperature accelerates protein unfolding without changing the pathway of unfolding. J. Mol. Biol. 2002, 322, 189–203. [Google Scholar] [CrossRef]
- Winzer, A.T.; Kraft, C.; Bhushan, S.; Stepanenko, V.; Tessmer, I. Correcting for AFM tip induced topography convolutions in protein–DNA samples. Ultramicrosc 2012, 121, 8–15. [Google Scholar] [CrossRef]
- Alev, B.; Tunali, S.; Yanardag, R.; Yarat, A. Influence of storage time and temperature on the activity of urease. Bulg. Chem. Commun. 2019, 51, 159–163. [Google Scholar] [CrossRef]
- Laboria, N.; Fragoso, A.; O’Sullivan, C.K. Storage properties of peroxidase labeled antibodies for the development of multiplexed packaged immunosensors for cancer markers. Anal. Lett. 2011, 44, 2019–2030. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technical Replicate | Group 1 | Group 2 | ||
|---|---|---|---|---|
| hmax1, nm | α1, % | hmax2, nm | α2, % | |
| 1 | 1 ± 0.2 | 93.7 | - | 6.3 |
| 2 | 79.8 | 1.4 ± 0.2 | 20.2 | |
| 3 | 96.4 | - | 3.6 | |
| 4 | 73.2 | 1.5 ± 0.2 | 26.8 | |
| Mean value | 1 ± 0.2 | 85.8 ± 9.6 | 1.45 ± 0.2 | 14.2 ± 9.6 |
| Storage Duration | “Group 1”, α1 % | “Group 2”, α2 % |
|---|---|---|
| 1 day | 88.5 ± 9.3 | 11.5 ± 9.3 |
| 7 days | 92.6 ± 5.5 | 7.4 ± 5.5 |
| 14 days | 88.3 ± 9.1 | 11.7 ± 9.1 |
| 28 days | 89.3 ± 6.7 | 10.7 ± 6.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanova, I.A.; Ershova, M.O.; Shumov, I.D.; Valueva, A.A.; Ivanov, Y.D.; Pleshakova, T.O. Atomic Force Microscopy Study of the Temperature and Storage Duration Dependencies of Horseradish Peroxidase Oligomeric State. Biomedicines 2022, 10, 2645. https://doi.org/10.3390/biomedicines10102645
Ivanova IA, Ershova MO, Shumov ID, Valueva AA, Ivanov YD, Pleshakova TO. Atomic Force Microscopy Study of the Temperature and Storage Duration Dependencies of Horseradish Peroxidase Oligomeric State. Biomedicines. 2022; 10(10):2645. https://doi.org/10.3390/biomedicines10102645
Chicago/Turabian StyleIvanova, Irina A., Maria O. Ershova, Ivan D. Shumov, Anastasia A. Valueva, Yuri D. Ivanov, and Tatyana O. Pleshakova. 2022. "Atomic Force Microscopy Study of the Temperature and Storage Duration Dependencies of Horseradish Peroxidase Oligomeric State" Biomedicines 10, no. 10: 2645. https://doi.org/10.3390/biomedicines10102645