
VraSR Regulatory System Contributes to the Virulence of Community-Associated Methicillin-Resistant Staphylococcus aureus (CA-MRSA) in a 3D-Skin Model and Skin Infection of Humanized Mouse Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture and Reagents
2.2. Construction of Isogenic Knockout Mutant Strain
2.3. Generation of a 3D-Skin Model
2.4. Evaluation of MRSA Infection in 3D-Skin Model
2.5. Determination of Cytokine Levels
2.6. Generation of Humanized Mice
2.7. Cytotoxicity Assay
2.8. Humanized Mouse MRSA-SSTI Model
2.9. Confocal Laser Scanning Microscopy (CLSM) Analysis of Biofilms
3. Results
3.1. Construction of Isogenic Knockout Mutant Strain ST30ΔvraR
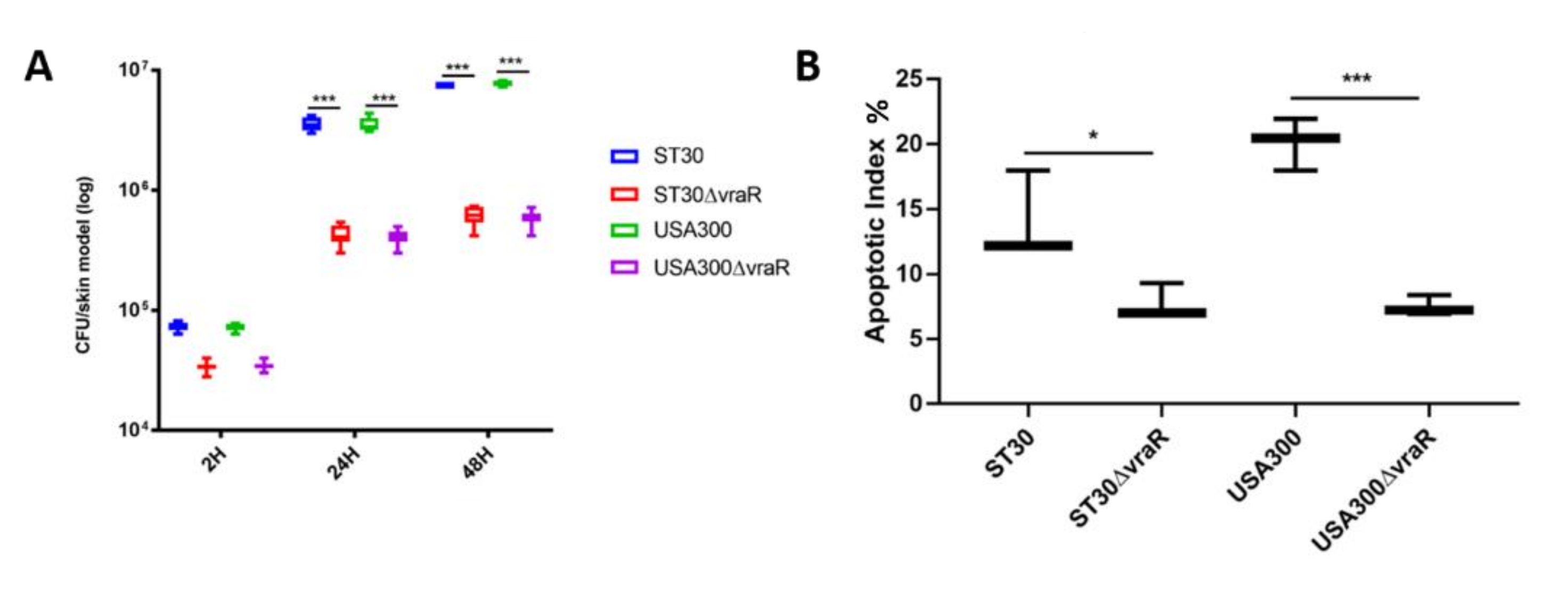
3.2. Infection of the Skin Model
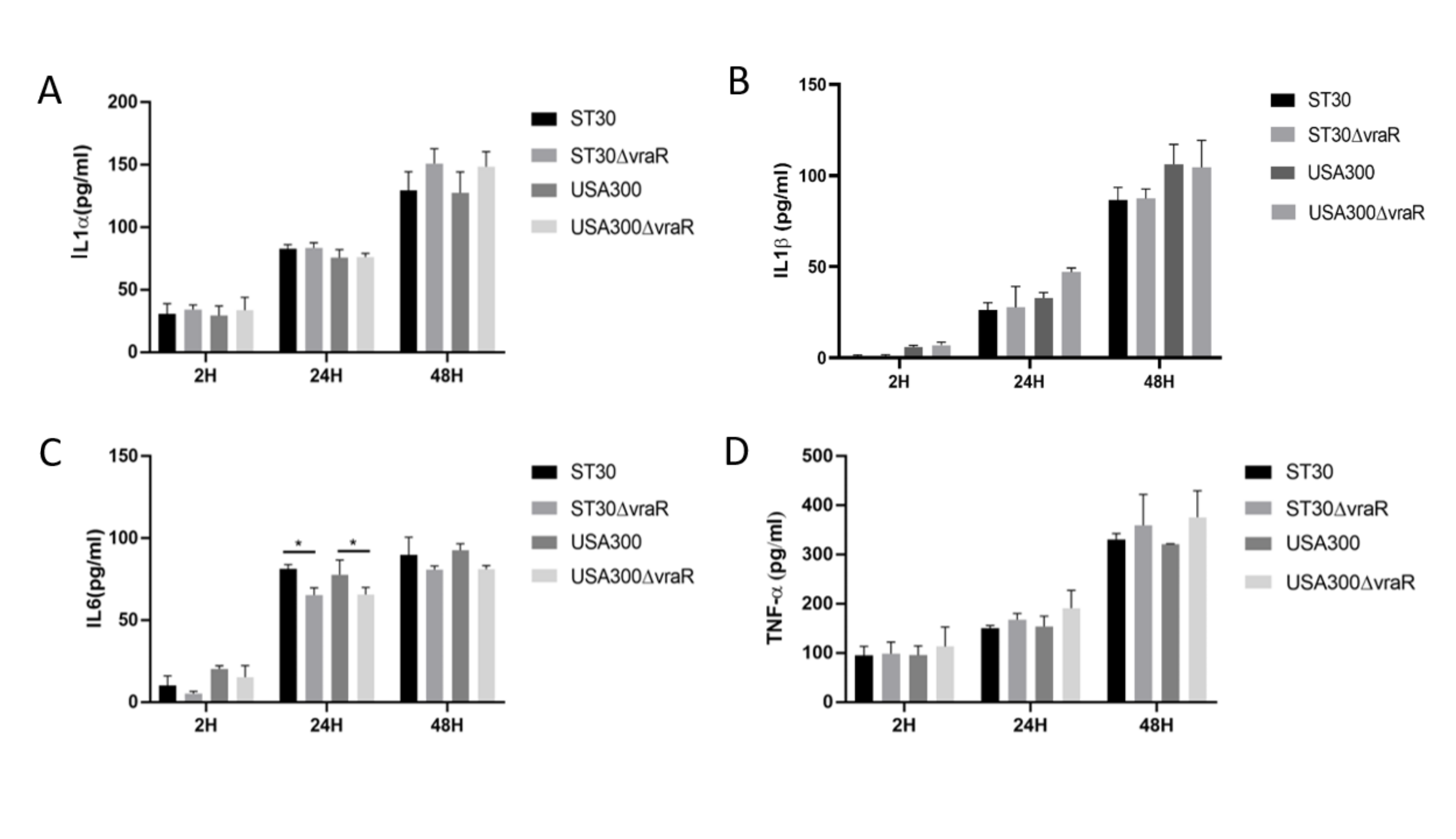
3.3. Cytokines
3.4. Humanized Mouse MRSA-SSTI Model
3.5. CSLM Analysis of Biofilm Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Queck, S.Y.; Jameson-Lee, M.; Villaruz, A.E.; Bach, T.H.L.; Khan, B.A.; Sturdevant, D.E.; Ricklefs, S.M.; Li, M.; Otto, M. RNAIII-Independent Target Gene Control by the agr Quorum-Sensing System: Insight into the Evolution of Virulence Regulation in Staphylococcus aureus. Mol. Cell 2008, 32, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCaig, L.F.; McDonald, L.C.; Mandal, S.; Jernigan, D.B. Staphylococcus aureus-associated skin and soft tissue infections in ambulatory care. Emerg. Infect. Dis. 2006, 12, 1715–1723. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.L.; Chuang, S.K.; Choi, Y.F.; Lee, R.A.; Lit, A.C.H.; Ng, T.K.; Que, T.L.; Shek, K.C.; Tong, H.K.; Tse, C.W.S.; et al. Community-associated methicillin-resistant and methicillin-sensitive Staphylococcus aureus: Skin and soft tissue infections in Hong Kong. Diagn. Microbiol. Infect. Dis. 2008, 61, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Talan, D.A.; Krishnadasan, A.; Gorwitz, R.J.; Fosheim, G.E.; Limbago, B.; Albrecht, V.; Moran, G.J. Comparison of staphylococcus aureus from skin and soft-tissue infections in us emergency Department patients, 2004 and 2008. Clin. Infect. Dis. 2011, 53, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, M.; Kato, F.; Oogai, Y.; Kawai, T.; Sugai, M.; Komatsuzawa, H. Distinct two-component systems in methicillin-resistant Staphylococcus aureus can change the susceptibility to antimicrobial agents. J. Antimicrob. Chemother. 2010, 65, 1536–1537. [Google Scholar] [CrossRef] [Green Version]
- Mascher, T. Intramembrane-sensing histidine kinases: A new family of cell envelope stress sensors in Firmicutes bacteria. FEMS Microbiol. Lett. 2006, 264, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle-Vavra, S.; Yin, S.; Jo, D.S.; Montgomery, C.P.; Daum, R.S. VraT/YvqF is required for methicillin resistance and activation of the VraSR regulon in Staphylococcus aureus. Antimicrob. Agents Chemother. 2013, 57, 83–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Staphylococcus aureus toxins. Curr. Opin. Microbiol. 2014, 17, 32–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonzo, F.; Torres, V.J. Bacterial Survival Amidst an Immune Onslaught: The Contribution of the Staphylococcus aureus Leukotoxins. PLoS Pathog. 2013, 9, 12–15. [Google Scholar] [CrossRef] [Green Version]
- Parker, D. Humanized mouse models of Staphylococcus aureus infection. Front. Immunol. 2017, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, A.R.; Salgado-Pabón, W.; Kohler, P.L.; Horswill, A.R.; Leung, D.Y.M.; Schlievert, P.M. Staphylococcal and streptococcal superantigen exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Zhong, J.; Fang, Y.; Geisinger, E.; Novick, R.P.; Lambowitz, A.M. Use of targetrons to disrupt essential and nonessential genes in Staphylococcus aureus reveals temperature sensitivity of Ll.LtrB group II intron splicing. RNA 2006, 12, 1271–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monk, I.R.; Shah, I.M.; Xu, M.; Tan, M.W.; Foster, T.J. Transforming the untransformable: Application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. mBio 2012, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.; Huang, L.; Wong, Y.P.; Burd, A. HA modulation of epidermal morphogenesis in an organotypic keratinocyte-fibroblast co-culture model. Exp. Dermatol. 2010, 19, 336–339. [Google Scholar] [CrossRef]
- Van Pham, P.; Le, H.T.; Vu, B.T.; Pham, V.Q.; Le, P.M.; Phan, N.L.C.; Van Trinh, N.; Nguyen, H.T.L.; Nguyen, S.T.; Nguyen, T.L.; et al. Targeting breast cancer stem cells by dendritic cell vaccination in humanized mice with breast tumor: Preliminary results. Onco-Targets Ther. 2016, 9, 4441–4451. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.W.; Biancotti, J.C.; Berg, B.L.; Gate, D.; Kolar, S.L.; Müller, S.; Rodriguez, M.D.; Rezai-Zadeh, K.; Fan, X.; Beenhouwer, D.O.; et al. Increased Susceptibility of Humanized NSG Mice to Panton-Valentine Leukocidin and Staphylococcus aureus Skin Infection. PLoS Pathog. 2015, 11, e1005292. [Google Scholar] [CrossRef] [PubMed]
- García, C.A.; Alcaraz, E.S.; Franco, M.A.; De Rossi, B.N.P. Iron is a signal for Stenotrophomonas maltophilia biofilm formation, oxidative stress response, OMPs expression, and virulence. Front. Microbiol. 2015, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.Y.; Singh, A.; David, M.Z.; Bartsch, S.M.; Slayton, R.B.; Huang, S.S.; Zimmer, S.M.; Potter, M.A.; Macal, C.M.; Lauderdale, D.S.; et al. The economic burden of community-associated methicillin-resistant Staphylococcus aureus (CA-MRSA). Clin. Microbiol. Infect. 2013, 19, 528–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.J.; Bayer, A.S.; Mishra, N.N.; Meehl, M.; Ledala, N.; Yeaman, M.R.; Xiong, Y.Q.; Cheung, A.L. The Staphylococcus aureus two-component regulatory system, grars, senses and confers resistance to selected cationic antimicrobial peptides. Infect. Immun. 2012, 80, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Mainiero, M.; Goerke, C.; Geiger, T.; Gonser, C.; Herbert, S.; Wolz, C. Differential target gene activation by the Staphylococcus aureus two-component system saeRS. J. Bacteriol. 2010, 192, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle-Vavra, S.; Yin, S.; Daum, R.S. The VraS/VraR two-component regulatory system required for oxacillin resistance in community-acquired methicillin-resistant Staphylococcus aureus. FEMS Microbiol. Lett. 2006, 262, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, B.; Klier, A.; Rapoport, G. The two-component system ArlS-ArlR is a regulator of virulence gene expression in Staphylococcus aureus. Mol. Microbiol. 2001, 41, 247–261. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, J.L.; Spinner, J.L.; Minnich, S.A.; Kobayashi, S.D. Yersinia pestis two-component gene regulatory systems promote survival in human neutrophils. Infect. Immun. 2010, 78, 733–782. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.; Li, W.; Shi, G.; Li, H.; Chen, H.; Bei, W. The VraSR regulatory system contributes to virulence in Streptococcus suis via resistance to innate immune defenses. Virulence 2018, 9, 681–682. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Dai, Y.; Chang, W.; Fang, C.; Wang, Z.; Ma, X. VraSR has an important role in immune evasion of Staphylococcus aureus with low level vancomycin resistance. Microbes Infect. 2019, 21, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Chang, W.; Zhao, C.; Peng, J.; Xu, L. VraR Binding to the Promoter Region of agr Inhibits Its Function in Vancomycin-Intermediate Staphylococcus aureus (VISA) and Heterogeneous VISA. Antimicrob. Agents Chemother. 2017, 61, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Missiakas, D.; Schneewind, O. Mouse models for infectious diseases caused by Staphylococcus aureus. J. Immunol. Methods 2014, 410, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Pabón, W.; Schlievert, P.M. Models matter: The search for an effective Staphylococcus aureus vaccine. Nat. Rev. Microbiol. 2014, 12, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Brehm, M.A.; Victor Garcia-Martinez, J.; Greiner, D.L. Humanized mice for immune system investigation: Progress, promise and challenges. Nat. Rev. Immunol. 2012, 12, 786–798. [Google Scholar] [CrossRef]
- Knop, J.; Hanses, F.; Leist, T.; Archin, N.M.; Buchholz, S.; Gläsner, J.; Gessner, A.; Wege, A.K. Staphylococcus aureus Infection in Humanized Mice: A New Model to Study Pathogenicity Associated with Human Immune Response. J. Infect. Dis. 2015, 212, 435–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, A.; Wang, H.; Kitur, K.; Parker, D. Humanized mice exhibit increased susceptibility to staphylococcus aureus pneumonia. J. Infect. Dis. 2017, 215, 1386–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tromp, A.T.; Van Gent, M.; Abrial, P.; Martin, A.; Jansen, J.P.; De Haas, C.J.C.; Van Kessel, K.P.M.; Bardoel, B.W.; Kruse, E.; Bourdonnay, E.; et al. Human CD45 is an f-component-specific receptor for the staphylococcal toxin Panton-Valentine leukocidin. Nat. Microbiol. 2018, 3, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Voyich, J.M.; Braughton, K.R.; Sturdevant, D.E.; Whitney, A.R.; Saïd-Salim, B.; Porcella, S.F.; Long, R.D.; Dorward, D.W.; Gardner, D.J.; Kreiswirth, B.N.; et al. Insights into Mechanisms Used by Staphylococcus aureus to Avoid Destruction by Human Neutrophils. J. Immunol. 2005, 175, 3907–3919. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barua, N.; Yang, Y.; Huang, L.; Ip, M. VraSR Regulatory System Contributes to the Virulence of Community-Associated Methicillin-Resistant Staphylococcus aureus (CA-MRSA) in a 3D-Skin Model and Skin Infection of Humanized Mouse Model. Biomedicines 2022, 10, 35. https://doi.org/10.3390/biomedicines10010035
Barua N, Yang Y, Huang L, Ip M. VraSR Regulatory System Contributes to the Virulence of Community-Associated Methicillin-Resistant Staphylococcus aureus (CA-MRSA) in a 3D-Skin Model and Skin Infection of Humanized Mouse Model. Biomedicines. 2022; 10(1):35. https://doi.org/10.3390/biomedicines10010035
Chicago/Turabian StyleBarua, Nilakshi, Ying Yang, Lin Huang, and Margaret Ip. 2022. "VraSR Regulatory System Contributes to the Virulence of Community-Associated Methicillin-Resistant Staphylococcus aureus (CA-MRSA) in a 3D-Skin Model and Skin Infection of Humanized Mouse Model" Biomedicines 10, no. 1: 35. https://doi.org/10.3390/biomedicines10010035