
Effect of Clostridium butyricum on Gastrointestinal Infections

{kind=link}
{kind=link}
{kind=link}
Abstract
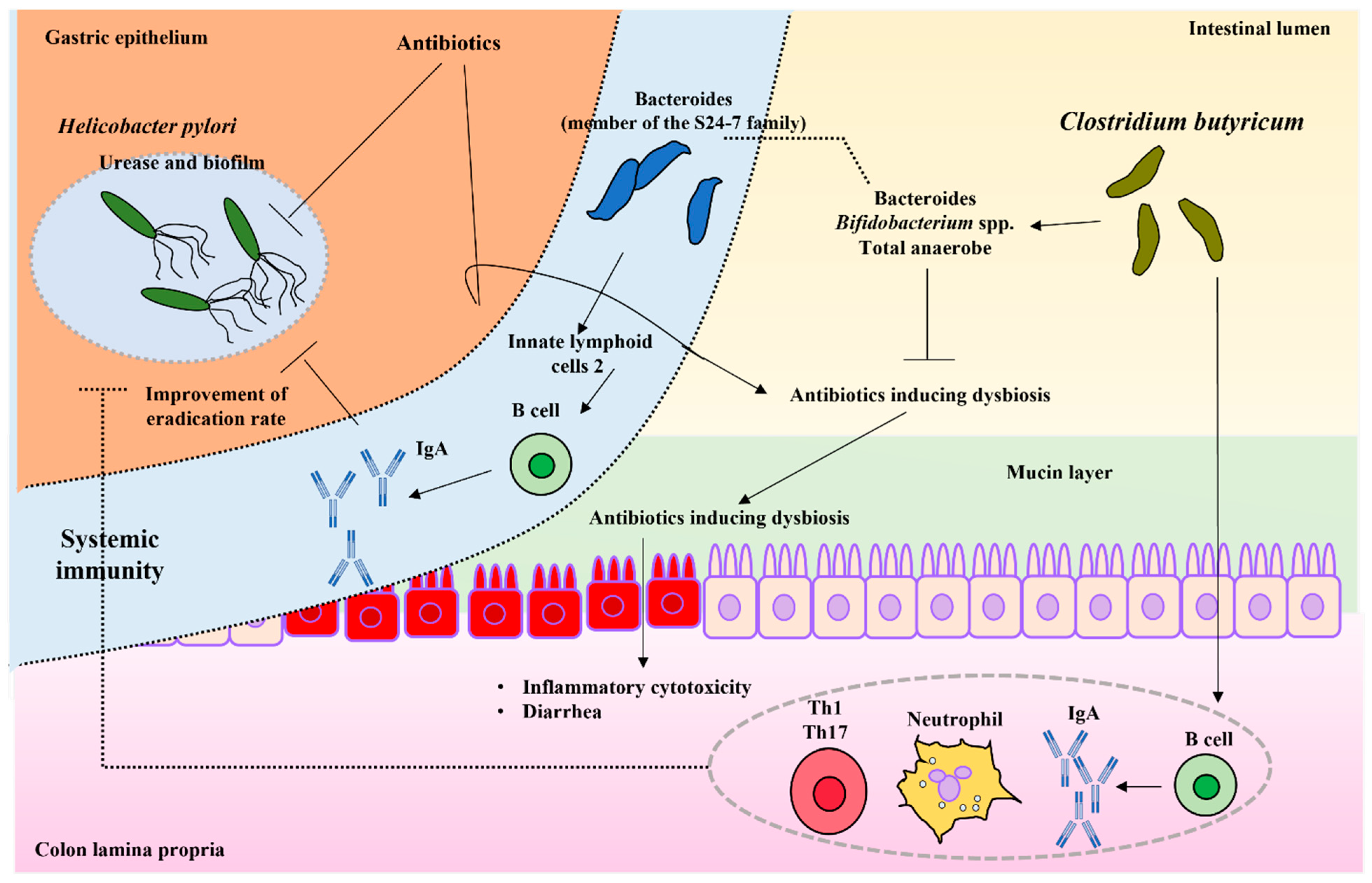
:1. Introduction
2. Clostridium butyricum
3. Gut Dysbiosis and Gastrointestinal Infections
4. Bacterial Gastrointestinal Infections
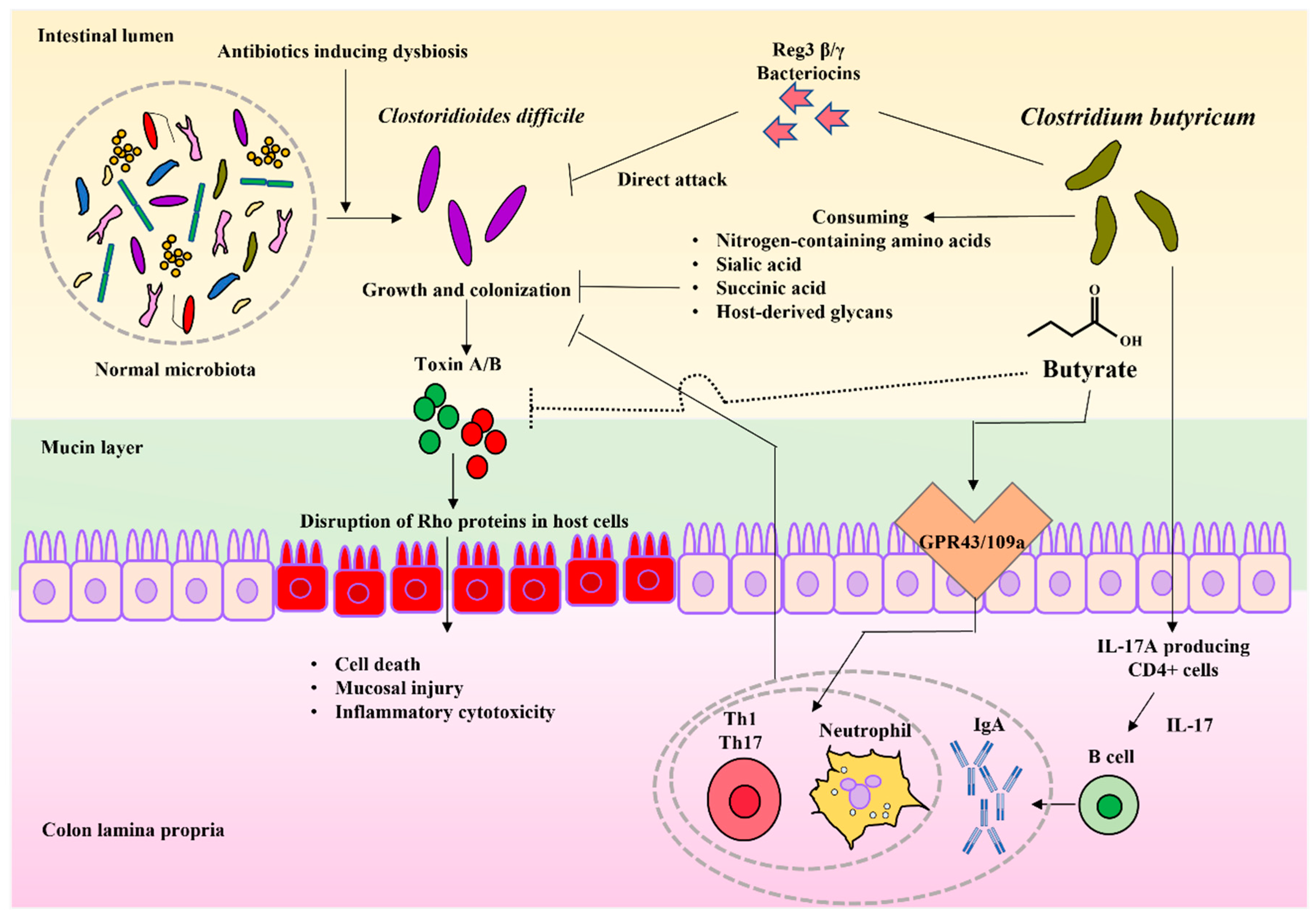
4.1. Clostridioides difficile Infection
4.2. Helicobacter pylori Infection
4.3. Escherichia coli Infection
4.4. Staphylococcus aureus Infection
4.5. Vibrio cholerae Infection
4.6. Salmonella Species Infection
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saberpour, M.; Bakhshi, B.; Najar-Peerayeh, S. Evaluation of the Antimicrobial and Antibiofilm Effect of Chitosan Nanoparticles as Carrier for Supernatant of Mesenchymal Stem Cells on Multidrug-Resistant Vibrio cholerae. Infect. Drug Resist. 2020, 13, 2251–2260. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Knight, D.R.; Riley, T.V. Clostridium difficile and One Health. Clin. Microbiol. Infect. 2020, 26, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Payne, D.C.; Vinjé, J.; Szilagyi, P.G.; Edwards, K.M.; Staat, M.A.; Weinberg, G.A.; Hall, C.B.; Chappell, J.; Bernstein, D.I.; Curns, A.T.; et al. Norovirus and medically attended gastroenteritis in U.S. children. N. Engl. J. Med. 2013, 368, 1121–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marder, E.P.; Cieslak, P.R.; Cronquist, A.B.; Dunn, J.; Lathrop, S.; Rabatsky-Her, T.; Ryan, P.; Smith, K.; Tobin-D’Angelo, M.; Vugia, D.J.; et al. Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food and the Effect of Increasing Use of Culture-Independent Diagnostic Tests on Surveillance-Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2013–2016. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 397–403, Erratum in MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 653. [Google Scholar] [CrossRef] [Green Version]
- Thiagarajah, J.R.; Donowitz, M.; Verkman, A.S. Secretory diarrhea: Mechanisms and emerging therapies. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 446–457. [Google Scholar] [CrossRef] [Green Version]
- Takeda, K.; Suzuki, T.; Shimada, S.I.; Shida, K.; Nanno, M.; Okumura, K. Interleukin-12 is involved in the enhancement of human natural killer cell activity by Lactobacillus casei Shirota. Clin. Exp. Immunol. 2006, 146, 109–115. [Google Scholar] [CrossRef]
- Oka, A.; Sartor, R.B. Microbial-Based and Microbial-Targeted Therapies for Inflammatory Bowel Diseases. Dig. Dis. Sci. 2020, 65, 757–788. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The Influence of the Gut Microbiome on Cancer, Immunity, and Cancer Immunotherapy. Cancer Cell 2018, 33, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, S.; Vlasova, A.N.; Fischer, D.D.; Chattha, K.S.; Shao, L.; Kumar, A.; Langel, S.N.; Rauf, A.; Huang, H.C.; Rajashekara, G.; et al. Unraveling the Differences between Gram-Positive and Gram-Negative Probiotics in Modulating Protective Immunity to Enteric Infections. Front. Immunol. 2017, 8, 334. [Google Scholar] [CrossRef] [Green Version]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Y. Gut microbiota derived metabolites in cardiovascular health and disease. Protein Cell 2018, 9, 416–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.X.; Wang, S.Y.; Kuo, C.H.; Tsai, I.L. Metabolome analysis for investigating host-gut microbiota interactions. J. Formos. Med. Assoc. 2019, 118 (Suppl. S1), S10–S22. [Google Scholar] [CrossRef] [PubMed]
- Fleming, S.E.; Rodriguez, M.A. Influence of dietary fiber on fecal excretion of volatile fatty acids by human adults. J. Nutr. 1983, 113, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Jiang, Q.; Yin, Y. Butyrate in Energy Metabolism: There Is Still More to Learn. Trends Endocrinol. Metab. 2021, 32, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. Biomed. Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Garg, R. Probiotics. Indian J. Med. Microbiol. 2009, 27, 202–209. [Google Scholar] [CrossRef]
- Sarao, L.K.; Arora, M. Probiotics, prebiotics, and microencapsulation: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 344–371. [Google Scholar] [CrossRef]
- Tonucci, L.B.; Dos Santos, K.M.O.; Ferreira, C.L.D.L.F.; Ribeiro, S.M.R.; De Oliveira, L.L.; Martino, H.S.D. Gut microbiota and probiotics: Focus on diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2017, 57, 2296–2309. [Google Scholar] [CrossRef]
- Williams, N.T. Probiotics. Am. J. Health Syst. Pharm. 2010, 67, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Stoeva, M.K.; Garcia-So, J.; Justice, N.; Myers, J.; Tyagi, S.; Nemchek, M.; McMurdie, P.J.; Kolterman, O.; Eid, J. Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes 2021, 13, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Finegold, S.M.; Sutter, V.L.; Mathisen, G.E. Normal indigenous intestinal flora. In Human Intestinal Microflora in Health and Disease; Hentges, D., Ed.; Academic Press: New York, NY, USA, 1983; pp. 3–28. [Google Scholar]
- Vital, M.; Penton, C.R.; Wang, Q.; Young, V.B.; Antonopoulos, D.A.; Sogin, M.L.; Morrison, H.G.; Raffals, L.; Chang, E.B.; Huffnagle, G.B.; et al. A gene-targeted approach to investigate the intestinal butyrate-producing bacterial community. Microbiome 2013, 1, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyairi, C. Effect of hydrogen ion concentration on the development of intestinal microflora. Characterization of a new spore bacterium isolated from feces. (Suiso ion noudo no tyonaikinso no hatsuiku ni oyobosu eikyo, hunben yori bunriseru ichi shingahoukin no seijo nit suite). J. Chiba Med. School 1935, 3, 1102–1157. (In Japanese) [Google Scholar]
- Miyarisan Pharmaceutical Co., Ltd. Clostridium Butyricum MIYAIRI Strain [Internet]. Clostridium Butyricum MIYAIRI Strain. Available online: http://www.miyarisan.com/english_index.htm (accessed on 8 January 2022).
- Seki, H.; Shiohara, M.; Matsumura, T.; Miyagawa, N.; Tanaka, M.; Komiyama, A.; Kurata, S. Prevention of antibiotic-associated diarrhea in children by Clostridium butyricum MIYAIRI. Pediatr. Int. 2003, 45, 86–90. [Google Scholar] [CrossRef]
- Yuzawa, T.; Kobayashi, A.; Inoue, H.; Enomoto, M. Twelve-Month Chronic Toxicity Study of Clostridium butyricum Miyairi Powder in Rats. Pharmacometrics 1987, 33, 683–694. [Google Scholar]
- Yuzawa, T.; Kobayashi, A.; Inoue, H.; Enomoto, M. A Five-Week Subacute Oral Toxicity and Recovery Test of Clostridium butyricum Miyairi Powder in Beagle Dogs. Pharmacometrics 1987, 34, 223–237. [Google Scholar]
- Yuzawa, T.; Kobayashi, A.; Yamamoto, T.; Nakajima, M.; Inoue, H. Tests for the Acute Oral Toxicity in Rats and Mutagenicity of Clostridium butyricum Miyairi Powder. Pharmacometrics 1987, 34, 215–221. [Google Scholar]
- Isa, K.; Oka, K.; Beauchamp, N.; Sato, M.; Wada, K.; Ohtani, K.; Nakanishi, S.; McCartney, E.; Tanaka, M.; Shimizu, T.; et al. Safety Assessment of the Clostridium butyricum MIYAIRI 588® Probiotic Strain Including Evaluation of Antimicrobial Sensitivity and Presence of Clostridium Toxin Genes in Vitro and Teratogenicity in Vivo. Hum. Exp. Toxicol. 2016, 35, 818–832. [Google Scholar] [CrossRef]
- Oka, K.; McCartney, E.; Ariyoshi, T.; Kudo, H.; Vilá, B.; de Jong, L.; Kozłowski, K.; Jankowski, J.; Morgan, S.; Kruger, C.; et al. In Vivo Safety Evaluation of the Clostridium butyricum MIYAIRI 588 Strain in Broilers, Piglets, and Turkeys. Toxicol. Res. Appl. 2019, 3, 2397847319826955. [Google Scholar] [CrossRef] [Green Version]
- The European Commission. Commission Implementing Decision of 11 December 2014 authorising the placing on the market of Clostridium butyricum (CBM 588) as a novel food ingredient under Regulation (EC) No 258/97 of the European Parliament and of the Council (notified under document C (2014) 9345). OJEU 2014, 57, 153. [Google Scholar]
- Hagihara, M.; Yamashita, R.; Matsumoto, A.; Mori, T.; Kuroki, Y.; Kudo, H.; Oka, K.; Takahashi, M.; Nonogaki, T.; Yamagishi, Y.; et al. The impact of Clostridium butyricum MIYAIRI 588 on the murine gut microbiome and colonic tissue. Anaerobe 2018, 54, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, M.; Yamashita, R.; Matsumoto, A.; Mori, T.; Kuroki, Y.; Kudo, H.; Oka, K.; Takahashi, M.; Nonogaki, T.; Yamagishi, Y.; et al. The Impact of Clostridium Butyricum MIYAIRI 588 on Murine Gut Microbiome in Colonic Tissue. American Society for Microbiology (ASM) Microbe 2018, Atlanta, 2018, #SUNDAY-85, Poster Presentation. Available online: https://asm.org/Events/ASM-Microbe/Home (accessed on 19 January 2022).
- Hagihara, M.; Yamashita, R.; Matsumoto, A.; Mori, T.; Kuroki, Y.; Kudo, H.; Oka, K.; Takahashi, M.; Nonogaki, T.; Yamagishi, Y.; et al. The Impact of Probiotics Clostridium Butyricum MIYAIRI 588 on Murine Gut Metabolic Alterations. American Society for Microbiology (ASM) Microbe 2018, Atlanta, 2018, #SUNDAY-86, Poster Presentation. Available online: https://asm.org/Events/ASM-Microbe/Home (accessed on 19 January 2022).
- Hagihara, M.; Yamashita, R.; Matsumoto, A.; Mori, T.; Inagaki, T.; Nonogaki, T.; Kuroki, Y.; Higashi, S.; Oka, K.; Takahashi, M.; et al. The impact of probiotic Clostridium butyricum MIYAIRI 588 on murine gut metabolic alterations. J. Infect. Chemother. 2019, 25, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Merga, Y.; Campbell, B.J.; Rhodes, J.M. Mucosal barrier b.bacteria and inflammatory bowel disease: Possibilities for therapy. Dig. Dis. 2014, 32, 475–483. [Google Scholar] [CrossRef]
- Hagihara, M.; Kuroki, Y.; Yamashita, R.; Matsumoto, A.; Mori, T.; Nonogaki, T.; Oka, K.; Takahashi, M.; Yamagishi, Y.; Mikamo, H. Clostridium Butyricum MIYAIRI 588 Protect from the Colon Epithelial Damage Caused by Antimicrobials. American Society for Microbiology (ASM) Microbe 2019, San Francisco, 2019, #FRIDAY-HMB-340, Poster Presentation. Available online: https://asm.org/Events/ASM-Microbe/Home (accessed on 19 January 2022).
- Hagihara, M.; Kuroki, Y.; Ariyoshi, T.; Higashi, S.; Fukuda, K.; Yamashita, R.; Matsumoto, A.; Mori, T.; Mimura, K.; Yamaguchi, N.; et al. Clostridium butyricum Modulates the Microbiome to Protect Intestinal Barrier Function in Mice with Antibiotic-Induced Dysbiosis. iScience 2020, 23, 100772. [Google Scholar] [CrossRef] [Green Version]
- Ariyoshi, T.; Hagihara, M.; Eguchi, S.; Fukuda, A.; Iwasaki, K.; Oka, K.; Takahashi, M.; Yamagishi, Y.; Mikamo, H. Clostridium butyricum MIYAIRI 588-Induced Protectin D1 Has an Anti-inflammatory Effect on Antibiotic-Induced Intestinal Disorder. Front. Microbiol. 2020, 11, 587725. [Google Scholar] [CrossRef]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Vogt, S.L.; Finlay, B.B. Gut microbiota-mediated protect.tion against diarrheal infections. J. Travel Med. 2017, 24 (Suppl. S1), S39–S43. [Google Scholar] [CrossRef]
- Allam-Ndoul, B.; Castonguay-Paradis, S.; Veilleux, A. Gut Microbiota and Intestinal Trans-Epithelial Permeability. Int. J. Mol. Sci. 2020, 21, 6402. [Google Scholar] [CrossRef]
- Ahmadi, S.; Wang, S.; Nagpal, R.; Wang, B.; Jain, S.; Razazan, A.; Mishra, S.P.; Zhu, X.; Wang, Z.; Kavanagh, K.; et al. A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis. JCI Insight 2020, 5, e132055. [Google Scholar] [CrossRef]
- Fukui, H. Leaky Gut and Gut-Liver Axis in Liver Cirrhosis: Clinical Studies Update. Gut Liver 2021, 15, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; Sarkar, A.; McSkimming, D.I. Gut Microbiota and Immune System Interactions. Microorganisms 2020, 8, 1587, Erratum in Microorganisms 2020, 8, 2046. [Google Scholar] [CrossRef] [PubMed]
- Fata, G.L.; Weber, P.; Mohajeri, M.H. Probiotics and the Gut Immune System: Indirect Regulation. Probiot. Antimicrob. Proteins 2018, 10, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Tickler, I.A.; Obradovich, A.E.; Goering, R.V.; Fang, F.C.; Tenover, F.C.; HAI Consortium. Changes in molecular epidemiology and antimicrobial resistance profiles of Clostridioides (Clostridium) difficile strains in the United States between 2011 and 2017. Anaerobe 2019, 60, 102050. [Google Scholar] [CrossRef]
- Chandrasekaran, R.; Lacy, D.B. The role of toxins in Clostridium difficile infection. FEMS Microbiol. Rev. 2017, 41, 723–750. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Savidge, T.; Feng, H. The enterotoxicity of Clostridium difficile toxins. Toxins 2010, 2, 1848–1880. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.P.; LaMont, J.T. Clostridium difficile—more difficult than ever. N. Engl. J. Med. 2008, 359, 1932–1940, Erratum in N. Engl. J. Med. 2010, 363, 1585. [Google Scholar] [CrossRef]
- Evans, C.T.; Safdar, N. Current Trends in the Epidemiology and Outcomes of Clostridium difficile Infection. Clin. Infect. Dis. 2015, 60 (Suppl. S2), S66–S71. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, R.; Ronga, L.; Addati, G.; Magrone, R.; Abbasciano, A.; Decimo, M.; Miragliotta, G. Clostridium difficile. A review on an emerging infection. Clin. Ter. 2019, 170, e41–e47. [Google Scholar] [CrossRef]
- Li, C.; Duan, J.; Liu, S.; Meng, X.; Fu, C.; Zeng, C.; Wu, A. Assessing the risk and disease burden of Clostridium difficile infection among patients with hospital-acquired pneumonia at a University Hospital in Central China. Infection 2017, 45, 621–628. [Google Scholar] [CrossRef]
- Czepiel, J.; Krutova, M.; Mizrahi, A.; Khanafer, N.; Enoch, D.A.; Patyi, M.; Deptuła, A.; Agodi, A.; Nuvials, X.; Pituch, H.; et al. Mortality Following Clostridioides difficile Infection in Europe: A Retrospective Multicenter Case-Control Study. Antibiotics 2021, 10, 299. [Google Scholar] [CrossRef]
- Fujii, H.; Maruyama, K.; Moriguti, M.; Sato, Y.; Takahashi, T.; Ito, K.; Yokoyama, H. Effect of Clostridium butyricum when Combined with Vancomycin in Treatment of Clostridium difficile-Associated Diarrhea. Iryo Yakugaku Jpn. J. Pharm. Health Care Sci. 2006, 32, 1009–1013. [Google Scholar] [CrossRef] [Green Version]
- Oka, K.; Osaki, T.; Hanawa, T.; Kurata, S.; Sugiyama, E.; Takahashi, M.; Tanaka, M.; Taguchi, H.; Kamiya, S. Establishment of an Endogenous Clostridium difficile Rat Infection Model and Evaluation of the Effects of Clostridium butyricum MIYAIRI 588 Probiotic Strain. Front. Microbiol. 2018, 9, 1264. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, S.; Taguchi, H.; Yamaguchi, H.; Osaki, T.; Takahashi, M.; Nakamura, S. Bacterioprophylaxis using Clostridium butyricum for lethal caecitis by Clostridium difficile in gnotobiotic mice. Rev. Med. Microbiol. 1997, 8 (Suppl. S1), S60. [Google Scholar] [CrossRef]
- Woo, T.D.H.; Oka, K.; Takahashi, M.; Hojo, F.; Osaki, T.; Hanawa, T.; Kurata, S.; Yonezawa, H.; Kamiya, S. Inhibition of the cytotoxic effect of Clostridium difficile in vitro by Clostridium butyricum MIYAIRI 588 strain. J. Med. Microbiol. 2011, 60, 1617–1625. [Google Scholar] [CrossRef]
- Hagihara, M.; Ariyoshi, T.; Kuroki, Y.; Eguchi, S.; Higashi, S.; Mori, T.; Nonogaki, T.; Iwasaki, K.; Yamashita, M.; Asai, N.; et al. Clostridium butyricum enhances colonization resistance against Clostridioides difficile by metabolic and immune modulation. Sci. Rep. 2021, 11, 15007. [Google Scholar] [CrossRef]
- Shen, A. A Gut Odyssey: The Impact of the Microbiota on Clostridium difficile Spore Formation and Germination. PLoS Pathog. 2015, 11, e1005157. [Google Scholar] [CrossRef]
- Abbas, A.; Zackular, J.P. Microbe-microbe interactions during Clostridioides difficile infection. Curr. Opin. Microbiol. 2020, 53, 19–25. [Google Scholar] [CrossRef]
- Carasi, P.; Trejo, F.M.; Pérez, P.F.; De Antoni, G.L.; Serradell, M.d.l.A. Surface proteins from Lactobacillus kefir antagonize in vitro cytotoxic effect of Clostridium difficile toxins. Anaerobe 2012, 18, 135–142. [Google Scholar] [CrossRef]
- Castagliuolo, I.; Riegler, M.F.; Valenick, L.; LaMont, J.T.; Pothoulakis, C. Saccharomyces boulardii protease inhibits the effects of Clostridium difficile toxins A and B in human colonic mucosa. Infect. Immun. 1999, 67, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, S.; Tanaka, M. Sequence analysis of a bacteriocinogenic plasmid of Clostridium butyricum and expression of the bacteriocin gene in Escherichia coli. Anaerobe 2010, 16, 253–257. [Google Scholar] [CrossRef]
- Hayashi, A.; Nagao-Kitamoto, H.; Kitamoto, S.; Kim, C.H.; Kamada, N. The Butyrate-Producing Bacterium Clostridium butyricum Suppresses Clostridioides difficile Infection via Neutrophil- and Antimicrobial Cytokine-Dependent but GPR43/109a-Independent Mechanisms. J. Immunol. 2021, 206, 1576–1585. [Google Scholar] [CrossRef]
- Baj, J.; Forma, A.; Sitarz, M.; Portincasa, P.; Garruti, G.; Krasowska, D.; Maciejewski, R. Helicobacter pylori Virulence Factors-Mechanisms of Bacterial Pathogenicity in the Gastric Microenvironment. Cells 2020, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Handa, O.; Naito, Y.; Yoshikawa, T. Redox biology and gastric carcinogenesis: The role of Helicobacter pylori. Redox Rep. 2011, 16, 1–7, Erratum in Redox Rep. 2014, 19, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, O.; Naito, Y.; Yoshikawa, T. Helicobacter pylori: A ROS-inducing bacterial species in the stomach. Inflamm. Res. 2010, 59, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Sugano, K. Effect of Helicobacter pylori eradication on the incidence of gastric cancer: A systematic review and meta-analysis. Gastric Cancer 2019, 22, 435–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidebotham, R.L.; Worku, M.L.; Karim, Q.N.; Dhir, N.K.; Baron, J.H. How Helicobacter pylori urease may affect external pH and influence growth and motility in the mucus environment: Evidence from in-vitro studies. Eur. J. Gastroenterol. Hepatol. 2003, 15, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, H.; Osaki, T.; Fukutomi, T.; Hanawa, T.; Kurata, S.; Zaman, C.; Hojo, F.; Kamiya, S. Diversification of the AlpB Outer Membrane Protein of Helicobacter pylori Affects Biofilm Formation and Cellular Adhesion. J. Bacteriol. 2017, 199, e00729-16. [Google Scholar] [CrossRef] [Green Version]
- Mukai, R.; Handa, O.; Suyama, Y.; Majima, A.; Naito, Y. Effectiveness of including probiotics to Helicobacter pylori eradication therapies. J. Clin. Biochem. Nutr. 2020, 67, 102–104. [Google Scholar] [CrossRef]
- Imase, K.; Takahashi, M.; Tanaka, A.; Tokunaga, K.; Sugano, H.; Tanaka, M.; Ishida, H.; Kamiya, S.; Takahashi, S. Efficacy of Clostridium butyricum preparation concomitantly with Helicobacter pylori eradication therapy in relation to changes in the intestinal microbiota. Microbiol. Immunol. 2008, 52, 156–161. [Google Scholar] [CrossRef]
- Chen, L.; Xu, W.; Lee, A.; He, J.; Huang, B.; Zheng, W.; Su, T.; Lai, S.; Long, Y.; Chu, H.; et al. The impact of Helicobacter pylori infection, eradication therapy and probiotic supplementation on gut microenvironment homeostasis: An open-label, randomized clinical trial. eBioMedicine 2018, 35, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh-Takayama, N.; Kato, T.; Motomura, Y.; Kageyama, T.; Taguchi-Atarashi, N.; Kinoshita-Daitoku, R.; Kuroda, E.; Di Santo, J.P.; Mimuro, H.; Moro, K.; et al. Bacteria-Induced Group 2 Innate Lymphoid Cells in the Stomach Provide Immune Protection through Induction of IgA. Immunity 2020, 52, 635–649.e4. [Google Scholar] [CrossRef]
- Takahashi, M.; Taguchi, H.; Yamaguchi, H.; Osaki, T.; Kamiya, S. Studies of the effect of Clostridium butyricum on Helicobacter pylori in several test models including gnotobiotic mice. J. Med. Microbiol. 2000, 49, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Bernet, M.F.; Brassart, D.; Neeser, J.R.; Servin, A.L. Lactobacillus acidophilus LA 1 binds to cultured human intestinal cell lines and inhibits cell attachment and cell invasion by enterovirulent bacteria. Gut 1994, 35, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Kabir, A.M.; Aiba, Y.; Takagi, A.; Kamiya, S.; Miwa, T.; Koga, Y. Prevention of Helicobacter pylori infection by lactobacilli in a gnotobiotic murine model. Gut 1997, 41, 49–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erawijantari, P.P.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Saito, Y.; Fukuda, S.; Yachida, S.; Yamada, T. Influence of gastrectomy for gastric cancer treatment on faecal microbiome and metabolome profiles. Gut 2020, 69, 1404–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eslami, M.; Bahar, A.; Keikha, M.; Karbalaei, M.; Kobyliak, N.M.; Yousefi, B. Probiotics function and modulation of the immune system in allergic diseases. Allergol. Immunopathol. 2020, 48, 771–788. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Cazorla, S.I.; Dumit, J.M.L.; Vélez, E.; Perdigón, G. Beneficial Effects of Probiotic Consumption on the Immune System. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Balakrishnan, B.; Barizuddin, S.; Wuliji, T.; El-Dweik, M. A rapid and highly specific immunofluorescence method to detect Escherichia coli O157:H7 in infected meat samples. Int. J. Food Microbiol. 2016, 231, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Fujita, I.; Takahashi, M. Studies on the anti-diarrheal activity of Clostridium butyricum Miyairi II 588. Effects of Clostridium butyricum Miyairi II 588 on the enterotoxicity of the enterotoxins produced by enterotoxigenic Escherichia coli. Jpn. Pharmacol. Ther. 1987, 15, 55–62. (In Japanese) [Google Scholar]
- Takahashi, M.; Taguchi, H.; Yamaguchi, H.; Osaki, T.; Komatsu, A.; Kamiya, S. The effect of probiotic treatment with Clostridium butyricum on enterohemorrhagic Escherichia coli O157:H7 infection in mice. FEMS Immunol. Med. Microbiol. 2004, 41, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunishima, H.; Ishibashi, N.; Wada, K.; Oka, K.; Takahashi, M.; Yamasaki, Y.; Aoyagi, T.; Takemura, H.; Kitagawa, M.; Kaku, M. The effect of gut microbiota and probiotic organisms on the properties of extended spectrum beta-lactamase producing and carbapenem resistant Enterobacteriaceae including growth, beta-lactamase activity and gene transmissibility. J. Infect. Chemother. 2019, 25, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, H. Staphylococcus aureus: The enduring pathogen in surgery. Surgeon 2012, 10, 357–360. [Google Scholar] [CrossRef]
- Asao, T.; Kumeda, Y.; Kawai, T.; Shibata, T.; Oda, H.; Haruki, K.; Nakazawa, H.; Kozaki, S. An extensive outbreak of staphylococcal food poisoning due to low-fat milk in Japan: Estimation of enterotoxin A in the incriminated milk and powdered skim milk. Epidemiol. Infect. 2003, 130, 33–40. [Google Scholar] [CrossRef]
- Bergdoll, M.S. Staphylococcus aureus. In Foodborne Bacterial Pathogens; Doyle, M.P., Ed.; CRC Press: New York, NY, USA, 1989; pp. 463–523. [Google Scholar]
- Hou, Y.; Wang, L.; Yi, D.; Ding, B.; Chen, X.; Wang, Q.; Zhu, H.; Liu, Y.; Yin, Y.; Gong, J.; et al. Dietary supplementation with tributyrin alleviates intestinal injury in piglets challenged with intrarectal administration of acetic acid. Br. J. Nutr. 2014, 111, 1748–1758. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Y.; Chen, D.F.; Wang, B.M.; Cao, H.L. Clostridium butyricum promotes ligands release from intestinal epithelial cells to activate epidermal growth factor receptor. Gastroenterology 2019, 156, S888–S889. [Google Scholar] [CrossRef]
- Ma, M.; Zhao, Z.; Liang, Q.; Shen, H.; Zhao, Z.; Chen, Z.; He, R.; Feng, S.; Cao, D.; Gan, G.; et al. Overexpression of pEGF improved the gut protective function of Clostridium butyricum partly through STAT3 signal pathway. Appl. Microbiol. Biotechnol. 2021, 105, 5973–5991. [Google Scholar] [CrossRef]
- Singleton, F.L.; Attwell, R.W.; Jangi, M.S.; Colwell, R.R. Influence of salinity and organic nutrient concentration on survival and growth of Vibrio cholerae in aquatic microcosms. Appl. Environ. Microbiol. 1982, 43, 1080–1085. [Google Scholar] [CrossRef] [Green Version]
- Huq, A.; Small, E.B.; West, P.A.; Huq, M.I.; Rahman, R.; Colwell, R.R. Ecological relationships between Vibrio cholerae and planktonic crustacean copepods. Appl. Environ. Microbiol. 1983, 45, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Kuroiwa, T.; Kobari, K.; Iwanaga, M. Inhibition of enteropathogens by Clostridium butyricum MIYAIRI 588. Kansenshogaku Zasshi 1990, 64, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, colonization, and virulence differences among serovars. Sci. World J. 2015, 2015, 520179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunstmann, L.; Meedom, L.; Hautekiet, V.; Veken, W.V.; Andersen, A.A. Reduction of Salmonella Serotiters by Clostridum Butyricum in a Commercial Pig Trial. In Proceedings of the ESPHM 2019, Utrecht, The Netherlands, 22–24 May 2019. VPH-PP-01, Poster Presentation. [Google Scholar]
- Zhao, X.; Yang, J.; Ju, Z.; Wu, J.; Wang, L.; Lin, H.; Sun, S. Clostridium butyricum Ameliorates Salmonella Enteritis Induced Inflammation by Enhancing and Improving Immunity of the Intestinal Epithelial Barrier at the Intestinal Mucosal Level. Front. Microbiol. 2020, 11, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.K.; Verma, R.; Ahuja, V.; Paul, J. Real-Time Analysis of Gut Flora in Entamoeba Histolytica Infected Patients of Northern India. BMC Microbiol. 2012, 12, 183. [Google Scholar] [CrossRef] [Green Version]
- The, H.C.; de Sessions, P.F.; Jie, S.; Thanh, D.P.; Thompson, C.N.; Minh, C.N.N.; Chu, C.W.; Tran, T.-A.; Thomson, N.R.; Thwaites, G.E.; et al. Assessing Gut Microbiota Perturbations during the Early Phase of Infectious Diarrhea in Vietnamese Children. Gut Microbes 2018, 9, 38–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tachikawa, T.; Seo, G.; Nakazawa, M.; Sueyoshi, M.; Ohishi, T.; Joh, K. Estimation of probiotics by infection model of infant rabbit with enterohemorrhagic Escherichia coli O157:H7. Kansenshogaku Zasshi 1998, 72, 1300–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Jenab, A.; Roghanian, R.; Emtiazi, G. Bacterial Natural Compounds with Anti-Inflammatory and Immunomodulatory Properties (Mini Review). Drug Des. Devel Ther. 2020, 14, 3787–3801. [Google Scholar] [CrossRef]
- Li, M.; van Esch, B.C.A.M.; Wagenaar, G.T.M.; Garssen, J.; Folkerts, G.; Henricks, P.A.J. Pro- and anti-inflammatory effects of short chain fatty acids on immune and endothelial cells. Eur. J. Pharmacol. 2018, 831, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Yajima, T. Sensory mechanisms for SCFA in the colon. In Physiological and Clinical Aspects of Short-Chain Fatty Acids; Cummings, J.H., Rombeau, J.L., Sakata, T., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 209–221. [Google Scholar]
- Yajima, T. Effect of Sodium Propionate on the Contractile Response of the Rat Ileum in Situ. Jpn. J. Pharmacol. 1984, 35, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Grider, J.R.; Piland, B.E. The peristaltic reflex induced by short-chain fatty acids is mediated by sequential release of 5-HT and neuronal CGRP but not BDNF. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G429–G437. [Google Scholar] [CrossRef] [PubMed]
- Cherbut, C. Effects of SCFA from the large intestine. In Physiological and Clinical Aspects of Short-Chain Fatty Acids; Cummings, J.H., Rombeau, J.L., Sakata, T., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 191–207. [Google Scholar]
- Shimotoyodome, A.; Meguro, S.; Hase, T.; Tokumitsu, I.; Sakata, T. Short chain fatty acids but not lactate or succinate stimulate mucus release in the rat colon. Comp. Biochem. Physiol. Part A 2000, 125, 525–531. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; de Roos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450, Erratum in Nature 2014, 506, 254. [Google Scholar] [CrossRef] [PubMed]
- Ariyoshi, T.; Hagihara, M.; Tomono, S.; Eguchi, S.; Minemura, A.; Miura, D.; Oka, K.; Takahashi, M.; Yamagishi, Y.; Mikamo, H. Clostridium butyricum MIYAIRI 588 Modifies Bacterial Composition under Antibiotic-Induced Dysbiosis for the Activation of Interactions via Lipid Metabolism between the Gut Microbiome and the Host. Biomedicines 2021, 9, 1065. [Google Scholar] [CrossRef]
- Fujii, T.; Jounai, K.; Horie, A.; Takahashi, H.; Suzuki, H.; Ohshio, K.; Fujiwara, D.; Yamamoto, N. Effects of Heat-Killed Lactococcus Lactis Subsp. Lactis JCM 5805 on Mucosal and Systemic Immune Parameters, and Antiviral Reactions to Influenza Virus in Healthy Adults; a Randomized Controlled Double-Blind Study. J. Funct. Foods 2017, 35, 513–521. [Google Scholar] [CrossRef]
- Tsuji, R.; Yamamoto, N.; Yamada, S.; Fujii, T.; Yamamoto, N.; Kanauchi, O. Induction of Anti-Viral Genes Mediated by Humoral Factors upon Stimulation with Lactococcus Lactis Strain Plasma Results in Repression of Dengue Virus Replication in Vitro. Antivir. Res. 2018, 160, 101–108. [Google Scholar] [CrossRef]
- Iwabuchi, N.; Xiao, J.-Z.; Yaeshima, T.; Iwatsuki, K. Oral Administration of Bifidobacterium Longum Ameliorates Influenza Virus Infection in Mice. Biol. Pharm. Bull. 2011, 34, 1352–1355. [Google Scholar] [CrossRef] [Green Version]
- Morita, M.; Kuba, K.; Ichikawa, A.; Nakayama, M.; Katahira, J.; Iwamoto, R.; Watanebe, T.; Sakabe, S.; Daidoji, T.; Nakamura, S.; et al. The Lipid Mediator Protectin D1 Inhibits Influenza Virus Replication and Improves Severe Influenza. Cell 2013, 153, 112–125. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ariyoshi, T.; Hagihara, M.; Takahashi, M.; Mikamo, H. Effect of Clostridium butyricum on Gastrointestinal Infections. Biomedicines 2022, 10, 483. https://doi.org/10.3390/biomedicines10020483
Ariyoshi T, Hagihara M, Takahashi M, Mikamo H. Effect of Clostridium butyricum on Gastrointestinal Infections. Biomedicines. 2022; 10(2):483. https://doi.org/10.3390/biomedicines10020483
Chicago/Turabian StyleAriyoshi, Tadashi, Mao Hagihara, Motomichi Takahashi, and Hiroshige Mikamo. 2022. "Effect of Clostridium butyricum on Gastrointestinal Infections" Biomedicines 10, no. 2: 483. https://doi.org/10.3390/biomedicines10020483