
Differential Expression and Localization of EHBP1L1 during the First Wave of Rat Spermatogenesis Suggest Its Involvement in Acrosome Biogenesis †



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care, Tissue Extraction, and Collection of Rat Spermatozoa
2.2. Isolation of Germinal Cells by Centrifugal Elutriation
2.3. Collection of Human Spermatozoa
2.4. Preparation of Total Protein Extracts and Western Blot Analysis
2.5. Immunofluorescence Analysis on Testis
2.6. Immunofluorescence Analysis on SPZ
2.7. Statistical Analysis
3. Results
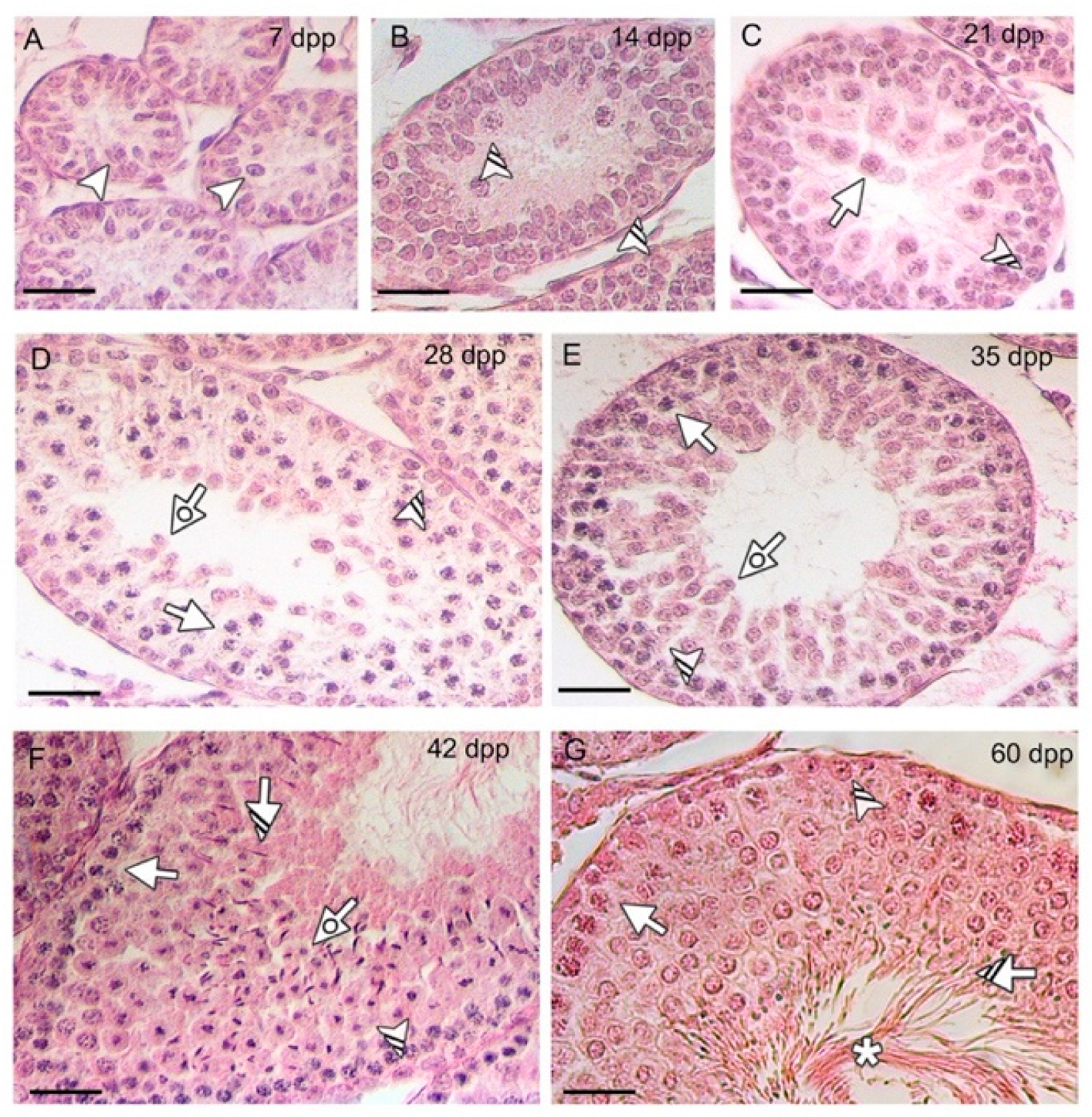
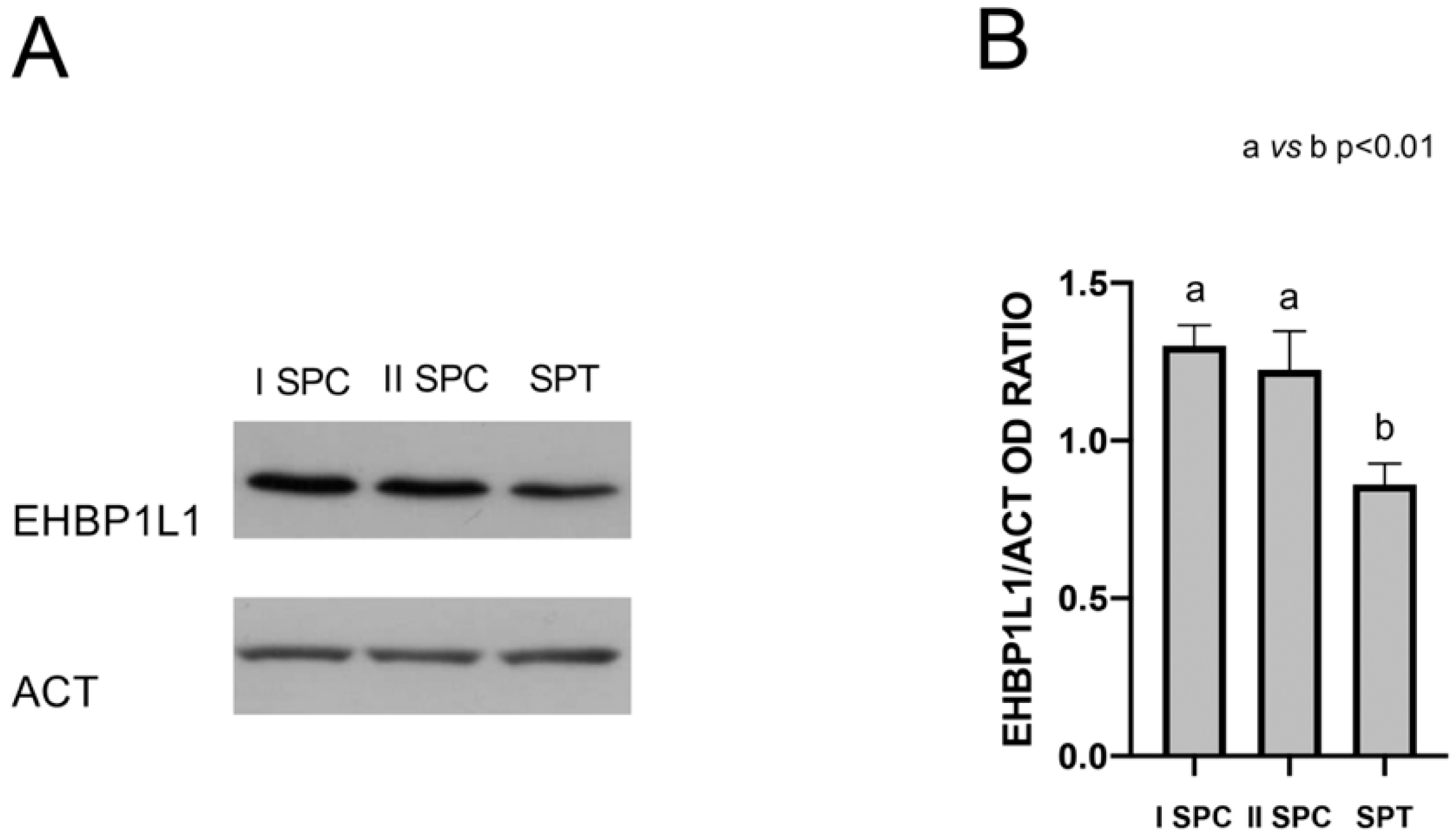
3.1. EHBP1L1 Protein Level during the First Wave of Rat Spermatogenesis
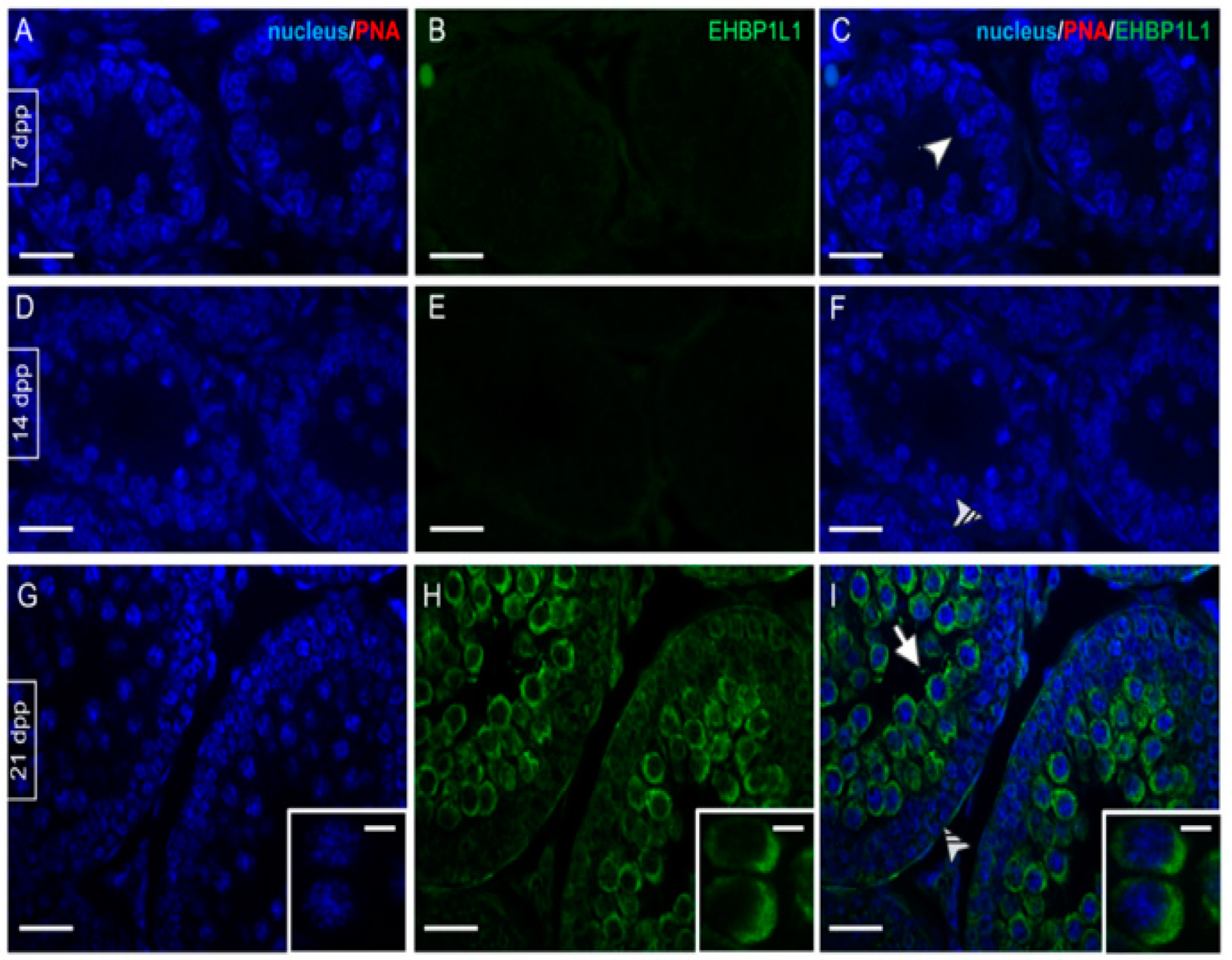
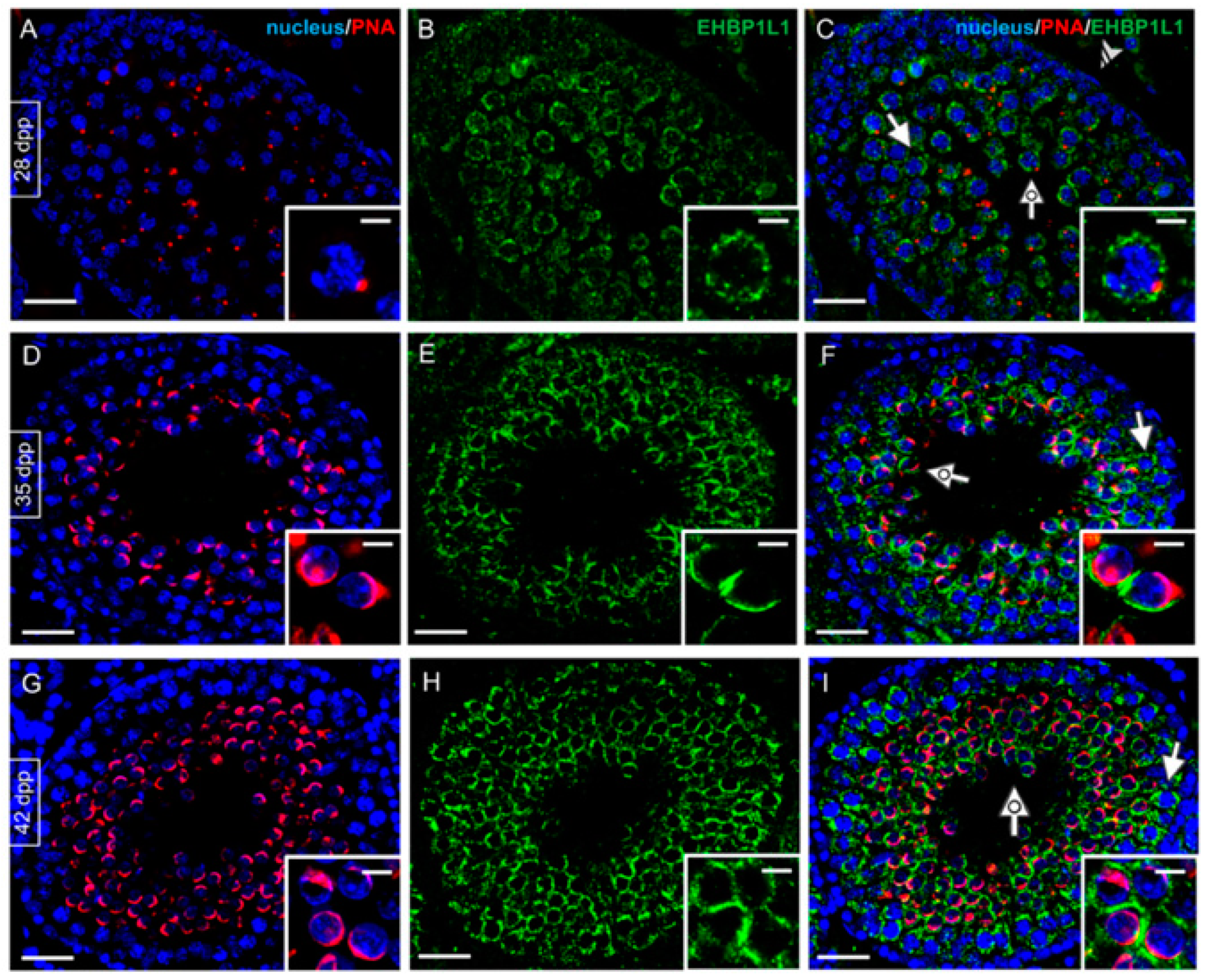
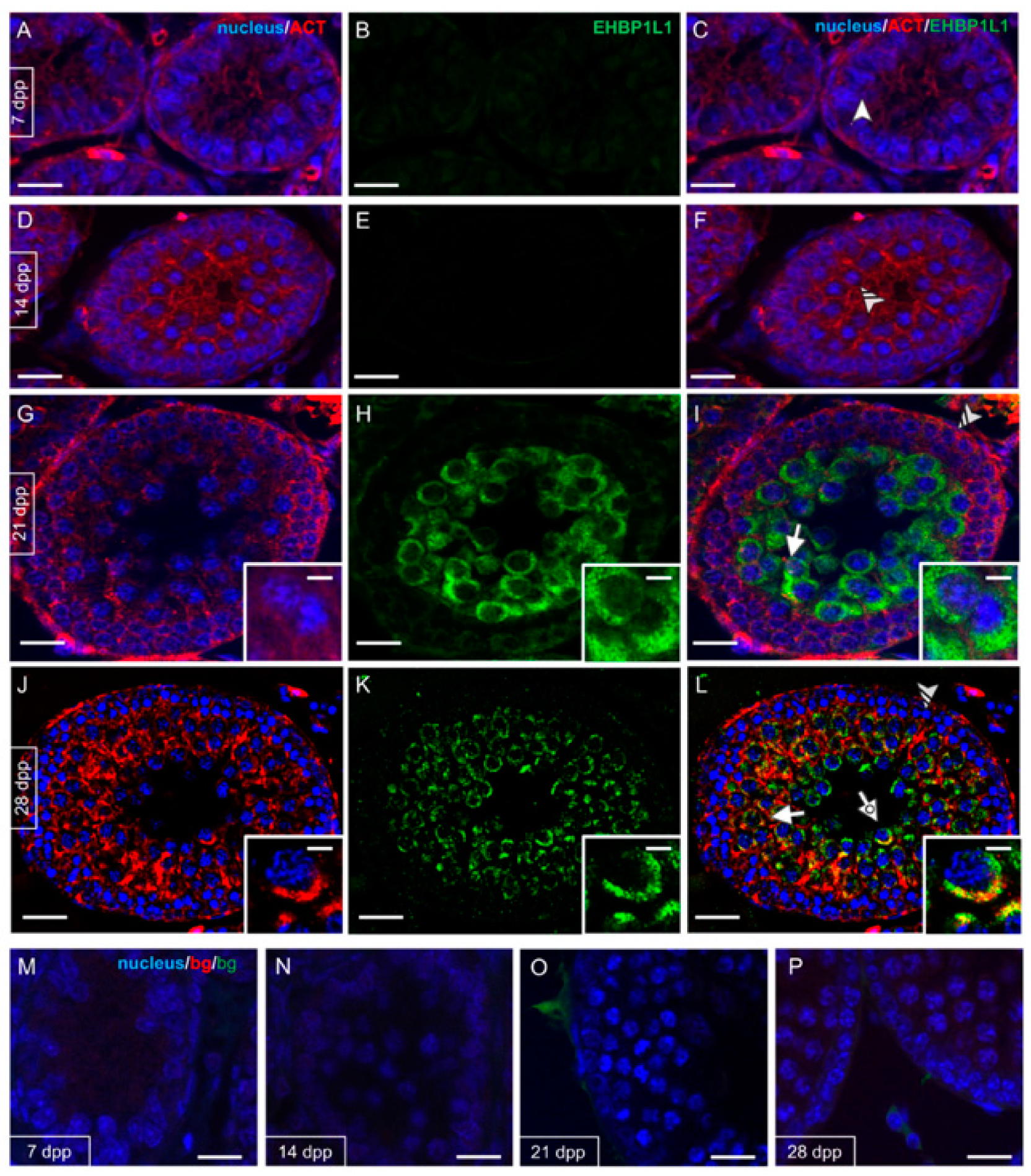
3.2. EHBP1L1 Localization during the First Wave of Rat Spermatogenesis
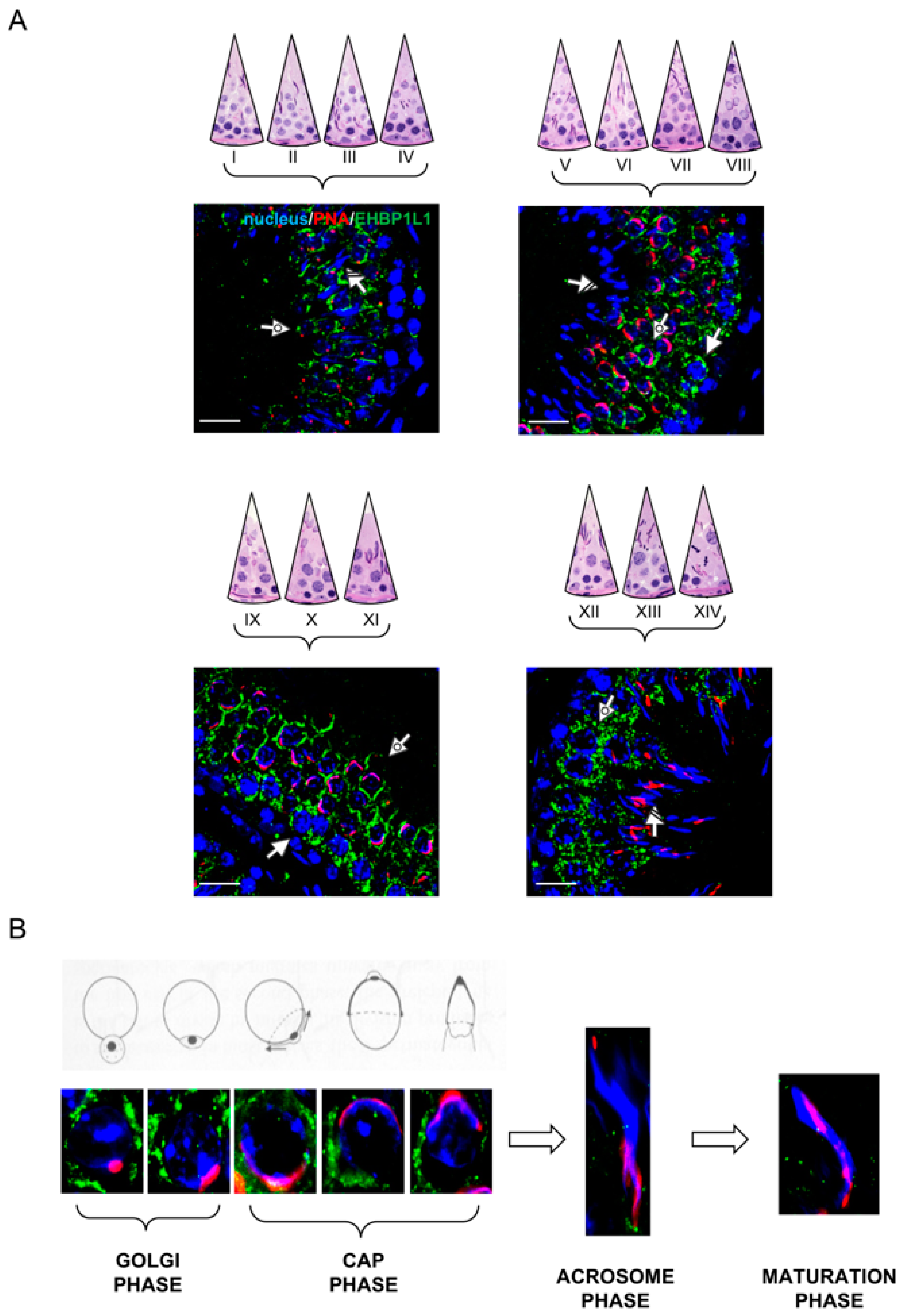
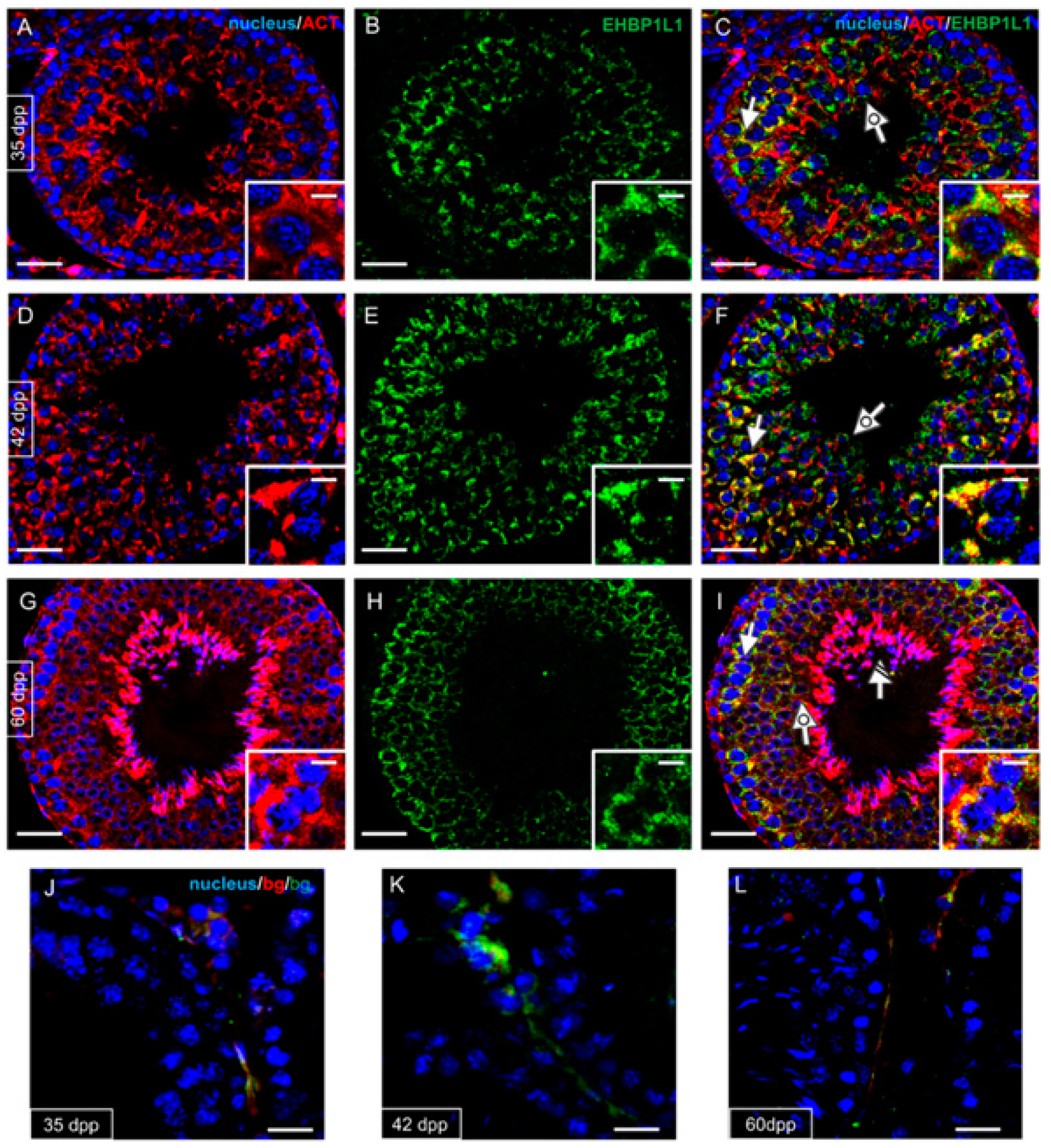
3.3. EHBP1L1 Localization in Adult Rat Testis and during AB
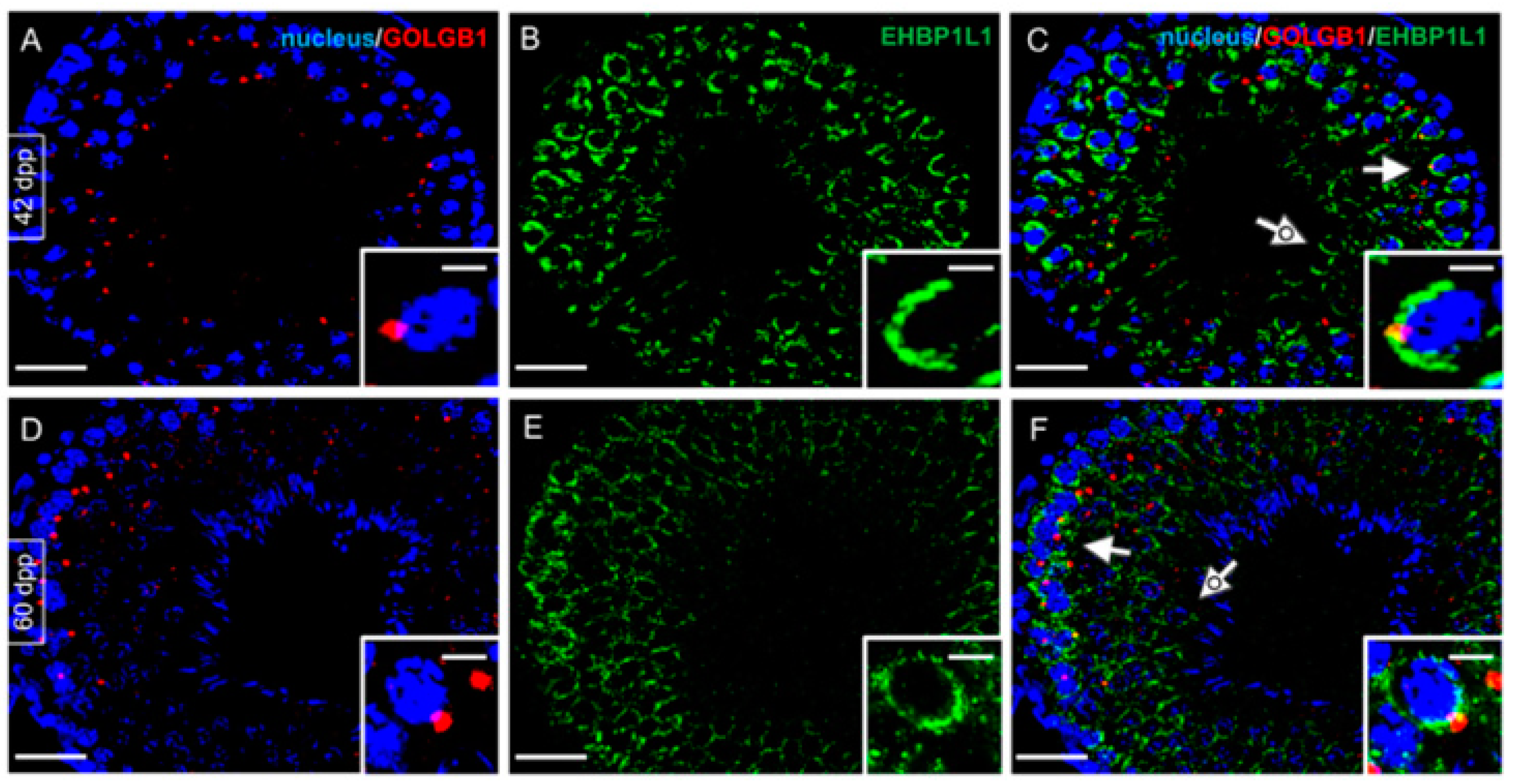
3.4. Co-Localization of EHBP1L1 with Actin and Golgi during the First Wave of Rat Spermatogenesis
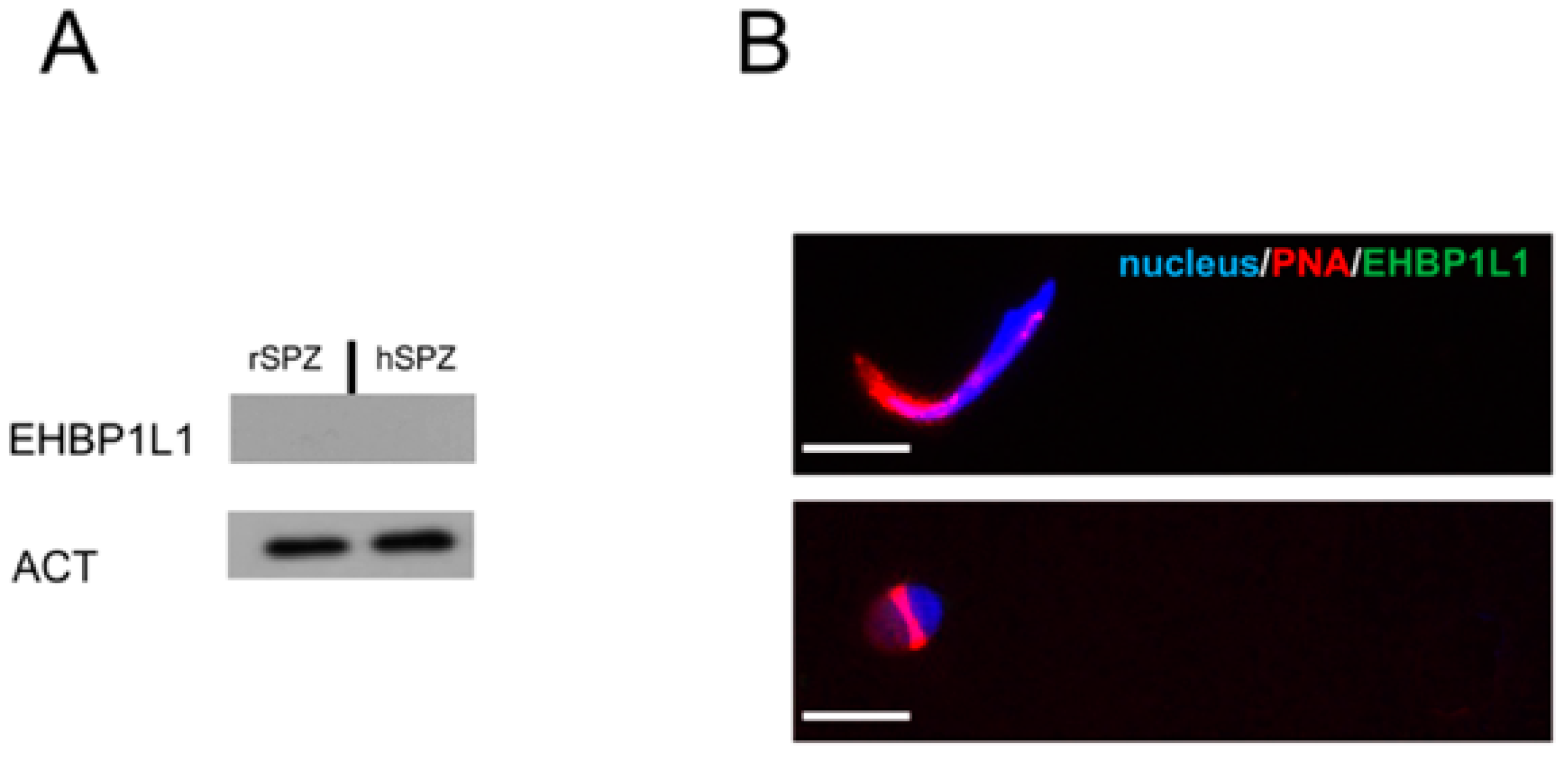
3.5. EHBP1L1 Analysis in Isolated Germ Cells and in Rat and Human SPZ
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, S.K. Human Zona Pellucida Glycoproteins: Binding Characteristics With Human Spermatozoa and Induction of Acrosome Reaction. Front. Cell Dev. Biol. 2021, 9, 619868. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.; Xu, F.; Hirschfeld, M.; Brenig, B. Sperm Lipid Markers of Male Fertility in Mammals. Int. J. Mol. Sci. 2021, 22, 8767. [Google Scholar] [CrossRef] [PubMed]
- Nasr-Esfahani, M.H.; Razavi, S.; Javdan, Z.; Tavalaee, M. Artificial oocyte activation in severe teratozoospermia undergoing intracytoplasmic sperm injection. Fertil. Steril. 2008, 90, 2231–2237. [Google Scholar] [CrossRef]
- Nasr-Esfahani, M.H.; Deemeh, M.R.; Tavalaee, M. Artificial oocyte activation and intracytoplasmic sperm injection. Fertil. Steril. 2010, 94, 520–526. [Google Scholar] [CrossRef]
- Nasr-Esfahani, M.H.; Tavalaee, M.; Reza Deemeh, M.; Arbabian, M.; Parrington, J. Can assessment of total acrosin activity help predict failed or low fertilization rate ICSI for implementation of artificial oocyte activation? Open Androl. J. 2010, 2, 19–26. [Google Scholar]
- Khawar, M.B.; Gao, H.; Li, W. Mechanism of Acrosome Biogenesis in Mammals. Front. Cell Dev. Biol. 2019, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Berruti, G. Towards defining an ‘origin’—The case for the mammalian acrosome. Semin. Cell Dev. Biol. 2016, 59, 46–53. [Google Scholar] [CrossRef]
- Xiong, W.; Shen, C.; Wang, Z. The molecular mechanisms underlying acrosome biogenesis elucidated by gene-manipulated mice. Biol. Reprod. 2021, 105, 789–807. [Google Scholar] [CrossRef]
- Berruti, G.; Paiardi, C. Acrosome biogenesis: Revisiting old questions to yield new insights. Spermatogenesis 2011, 1, 95–98. [Google Scholar] [CrossRef]
- Berruti, G.; Paiardi, C. USP8/UBPy-regulated sorting and the development of sperm acrosome: The recruitment of MET. Reproduction 2015, 149, 633–644. [Google Scholar] [CrossRef] [Green Version]
- Venditti, M.; Donizetti, A.; Aniello, F.; Minucci, S. EH domain binding protein 1-like 1 (EHBP1L1), a protein with calponin homology domain, is expressed in the rat testis. Zygote 2020, 28, 441–446. [Google Scholar] [CrossRef]
- Nakajo, A.; Yoshimura, S.; Togawa, H.; Kunii, M.; Iwano, T.; Izumi, A.; Noguchi, Y.; Watanabe, A.; Goto, A.; Sato, T.; et al. EHBP1L1 coordinates Rab8 and Bin1 to regulate apical-directed transport in polarized epithelial cells. J. Cell Biol. 2016, 212, 297–306. [Google Scholar] [CrossRef]
- Eguchi, T.; Kuwahara, T.; Sakurai, M.; Komori, T.; Fujimoto, T.; Ito, G.; Yoshimura, S.I.; Harada, A.; Fukuda, M.; Koike, M.; et al. LRRK2 and its substrate Rab GTPases are sequentially targeted onto stressed lysosomes and maintain their homeostasis. Proc. Natl. Acad. Sci. USA 2018, 115, E9115–E9124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Xu, E.; Zhao, H.T.; Cole, T.; West, A.B. LRRK2 and Rab10 coordinate macropinocytosis to mediate immunological responses in phagocytes. EMBO J. 2020, 39, e104862. [Google Scholar] [CrossRef]
- Friedberg, F. Singlet CH domain containing human multidomain proteins: An inventory. Mol. Biol. Rep. 2010, 37, 1531–1539. [Google Scholar] [CrossRef]
- World Health Organization. Standard procedures 2010. In WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; Cooper, T.G., Ed.; Cambridge University Press: Cambridge, UK, 2010; p. 68. [Google Scholar]
- Venditti, M.; Aniello, F.; Santillo, A.; Minucci, S. Study on PREP localization in mouse seminal vesicles and its possible involvement during regulated exocytosis. Zygote 2019, 27, 160–165. [Google Scholar] [CrossRef]
- Venditti, M.; Santillo, A.; Falvo, S.; Fiore, M.M.D.; Baccari, G.C.; Minucci, S. D-Aspartate upregulates DAAM1 protein levels in the rat testis and induces its localization in spermatogonia nucleus. Biomolecules 2020, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Malkov, M.; Fisher, Y.; Don, J. Developmental Schedule of the Postnatal Rat Testis Determined by Flow Cytometry. Biol. Reprod. 1998, 59, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Picut, C.A.; Remick, A.K.; de Rijk, E.P.; Simons, M.L.; Stump, D.G.; Parker, G.A. Postnatal development of the testis in the rat: Morphologic study and correlation of morphology to neuroendocrine parameters. Toxicol. Pathol. 2015, 43, 326–342. [Google Scholar] [CrossRef] [PubMed]
- Creasy, D.M. Evaluation of Testicular Toxicity in Safety Evaluation Studies: The Appropriate Use of Spermatogenic Staging. Toxicol. Pathol. 1997, 25, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, D.; Menkveld, R. Sperm morphology assessment-historical perspectives and current opinions. J. Androl. 2001, 22, 192–205. [Google Scholar] [PubMed]
- Pariante, P.; Dotolo, R.; Venditti, M.; Ferrara, D.; Donizetti, A.; Aniello, F.; Minucci, S. First evidence of DAAM1 localization during the post-natal development of rat testis and in mammalian sperm. J. Cell Physiol. 2016, 231, 2172–2184. [Google Scholar] [CrossRef] [PubMed]
- Sang, Q.; Zhou, Z.; Mu, J.; Wang, L. Genetic factors as potential molecular markers of human oocyte and embryo quality. J. Assist. Reprod. Genet. 2021, 38, 993–1002. [Google Scholar] [CrossRef]
- Bui, A.D.; Sharma, R.; Henkel, R.; Agarwal, A. Reactive oxygen species impact on sperm DNA and its role in male infertility. Andrologia 2018, 50, e13012. [Google Scholar] [CrossRef] [PubMed]
- Canipari, R.; De Santis, L.; Cecconi, S. Female Fertility and Environmental Pollution. Int. J. Environ. Res. Public. Health 2020, 17, 8802. [Google Scholar] [CrossRef] [PubMed]
- Chemek, M.; Venditti, M.; Boughamoura, S.; Mimouna, S.B.; Messaoudi, I.; Minucci, S. Involvement of testicular DAAM1 expression in zinc protection against cadmium-induced male rat reproductive toxicity. J. Cell Physiol. 2018, 233, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Tirpák, F.; Greifová, H.; Lukáč, N.; Stawarz, R.; Massányi, P. Exogenous Factors Affecting the Functional Integrity of Male Reproduction. Life 2021, 11, 213. [Google Scholar] [CrossRef] [PubMed]
- Venditti, M.; Chemek, M.; Minucci, S.; Messaoudi, I. Cadmium-induced toxicity increases prolyl endopeptidase (PREP) expression in the rat testis. Mol. Reprod. Dev. 2020, 87, 565–573. [Google Scholar] [CrossRef]
- Venditti, M.; Ben Rhouma, M.; Romano, M.Z.; Messaoudi, I.; Reiter, R.J.; Minucci, S. Evidence of melatonin ameliorative effects on the blood-testis barrier and sperm quality alterations induced by cadmium in the rat testis. Ecotoxicol. Environ. Saf. 2021, 226, 112878. [Google Scholar] [CrossRef] [PubMed]
- Krausz, C. Male infertility: Pathogenesis and clinical diagnosis. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 271–285. [Google Scholar] [CrossRef]
- Paiardi, C.; Pasini, M.E.; Gioria, M.; Berruti, G. Failure of acrosome formation and globozoospermia in the wobbler mouse, a Vps54 spontaneous recessivemutant. Spermatogenesis 2011, 1, 52–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venditti, M.; Fasano, C.; Minucci, S.; Serino, I.; Sinisi, A.A.; Dale, B.; Di Matteo, L. DAAM1 and PREP are involved in human spermatogenesis. Reprod. Fertil. Dev. 2020, 32, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Dunleavy, J.E.M.; O’Bryan, M.; Stanton, P.G.; O’Donnell, L. The cytoskeleton in spermatogenesis. Reproduction 2018, 157, R53–R72. [Google Scholar] [CrossRef] [Green Version]
- Venditti, M.; Minucci, S. Subcellular localization of prolyl endopeptidase during the first wave of rat spermatogenesis and in rat and human sperm. J. Histochem. Cytochem. 2019, 67, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Connolly, A.A.; Osterberg, V.; Christensen, S.; Price, M.; Lu, C.; Chicas-Cruz, K.; Lockery, S.; Mains, P.E.; Bowerman, B. Caenorhabditis elegans oocyte meiotic spindle pole assembly requires microtubule severing and the calponin homology domain protein ASPM-1. Mol. Biol. Cell 2014, 25, 1298–1311. [Google Scholar] [CrossRef]
- Mintz, L.; Galperin, E.; Pasmanik-Chor, M.; Tulzinsky, S.; Bromberg, Y.; Kozak, C.A.; Joyner, A.; Fein, A.; Horowitz, M. EHD1—An EH-domain-containing protein with a specific expression pattern. Genomics 1999, 59, 66–76. [Google Scholar] [CrossRef]
- Shannon, K.B.; Li, R. The multiple roles of Cyk1p in the assembly and function of the actomyosin ring in budding yeast. Mol. Biol. Cell 1999, 10, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Monesi, V.; Geremia, R.; D’Agostino, A.; Boitani, C. Biochemistry of male germ cell differentiation in mammals: RNA synthesis in meiotic and postmeiotic cells. Curr. Top. Dev. Biol. 1978, 12, 11–36. [Google Scholar] [CrossRef]
- Ferrara, D.; Izzo, G.; Pariante, P.; Donizetti, A.; d’Istria, M.; Aniello, F.; Minucci, S. Expression of prothymosin alpha in meiotic and post-meiotic germ cells during the first wave of rat spermatogenesis. J. Cell Physiol. 2010, 224, 362–368. [Google Scholar] [CrossRef]
- Venditti, M.; Minucci, S. Prothymosin alpha expression in the vertebrate testis: A comparative review. Zygote 2017, 25, 760–770. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venditti, M.; Minucci, S. Differential Expression and Localization of EHBP1L1 during the First Wave of Rat Spermatogenesis Suggest Its Involvement in Acrosome Biogenesis. Biomedicines 2022, 10, 181. https://doi.org/10.3390/biomedicines10010181
Venditti M, Minucci S. Differential Expression and Localization of EHBP1L1 during the First Wave of Rat Spermatogenesis Suggest Its Involvement in Acrosome Biogenesis. Biomedicines. 2022; 10(1):181. https://doi.org/10.3390/biomedicines10010181
Chicago/Turabian StyleVenditti, Massimo, and Sergio Minucci. 2022. "Differential Expression and Localization of EHBP1L1 during the First Wave of Rat Spermatogenesis Suggest Its Involvement in Acrosome Biogenesis" Biomedicines 10, no. 1: 181. https://doi.org/10.3390/biomedicines10010181